Abstract
The helminthological examination of nine individuals of Aetobatus cf. narinari (spotted eagle ray; raya pinta; arraia pintada) revealed the presence of an undescribed species of cestode of the genus Acanthobothrium. The stingrays were collected from four locations in México: Laguna Términos, south of Isla del Carmen and the marine waters north of Isla del Carmen and Champotón, in the State of Campeche, and Isla Holbox, State of Quintana Roo. The new species, nominated Acanthobothrium marquesi, is a category 3 species (i.e, the strobila is long, has more than 50 proglottids, the numerous testicles greater than 80, and has asymmetrically-lobed ovaries); at the present, the only category 3 species that has been reported in the Western Atlantic Ocean is Acanthobothrium tortum. Acanthobothrium marquesi n. sp. can be distinguished from A. tortum by length (26.1 cm vs. 10.6 cm), greater number of proglottids (1,549 vs. 656), a larger scolex (707 µm long by 872 µm wide vs. 699 µm long by 665 µm wide), larger bothridia (626 µm long by 274 µm wide vs. 563 µm long by 238 µm wide). This is the first report of a species of Acanthobothrium from the Mexican coast of the Gulf México.
Keywords:
Onchobothriidae; Acanthobothrium; Chondrichthyes; Myliobatiformes; helminth; México
Resumo
O exame helmintológico do trato digestivo de nove espécimes de Aetobatus cf. narinari (arraia pintada) revelou a presença de uma nova espécie de cestódeo do gênero Acanthobothrium. As arraias foram coletadas de quatro locais no México: Laguna Términos, ao sul de Isla del Carmen e nas águas marinhas ao norte de Isla del Carmen e Champotón, no estado de Campeche, e Isla Holbox, estado de Quintana Roo. A nova espécie foi denominada Acanthobothrium marquesi, pertencente a uma espécie da categoria 3 (estrobilo longo, tendo mais de 50 proglotes, numerosos testículos, superiores a 80, e ovários assimetricamente lobados). Apenas outra espécie deste gênero, Acanthobothrium tortum pertence a categoria 3, no Oceano Atlântico Ocidental. Acanthobothrium marquesi n. sp. se distingue de A. tortum por ser mais longo (26,1 cm vs. 10,6 cm), possuir maior número de proglotes (1.549 vs. 656), ter um escolex maior (707 µm de comprimento por 872 µm de largura vs. 699 µm de comprimento por 666 µm de largura), e botridias maiores (626 µm de comprimento por 274 µm de largura vs. 563 µm de comprimento por 238 µm de largura). Este é o primeiro relato de uma espécie de Acanthobothrium do Golfo do México.
Palavras-chave:
Onchobothriidae; Acanthobothrium; Chondrichthyes; Myliobatiformes; helminto; México
Introduction
Acanthobothrium is one of the richest genera within Onchoproteocephalidea, currently comprised of more than 188 nominal species (CAIRA & JENSEN, 2017Caira JN, Jensen K. Planetary biodiversity inventory (2008-2017): tapeworms from vertebrate bowels of the earth. Lawrence, Kansas: The University of Kansas, Museum of Natural History; 2017. Special Publication, n. 25.). Only a few occurrences have been documented in México, and most for the Pacific coast. In the first report of this genus in Mexican waters, Monks et al. (1996)Monks S, Brooks DR, Pérez-Ponce de Leon G. A new species of Acanthobothrium Van Beneden, 1849 (Eucestoda: Tetraphyllidea: Onchobothriidae) in Dasyatis longus Garman (Chondrichthyes: Myliobatiformes: Dasyatididae) from Chamela Bay, Jalisco, Mexico. J Parasitol 1996; 82(3): 484-488. PMid:8636857. http://dx.doi.org/10.2307/3284090.
http://dx.doi.org/10.2307/3284090...
described A. cleofanus Monks, Brooks, and Pérez-Ponce de León, 1996, from Hypanus longus (Garman, 1880). In three studies derived from the same project, Ghoshroy & Caira (2001)Ghoshroy S, Caira JN. Four new species of Acanthobothrium (Cestoda: Tetraphyllidea) from the whiptail stingray Dasyatis brevis in the Gulf of California, Mexico. J Parasitol 2001; 87(2): 354-372. PMid:11318566. http://dx.doi.org/10.1645/0022-3395(2001)087[0354:FNSOAC]2.0.CO;2.
http://dx.doi.org/10.1645/0022-3395(2001...
, Caira & Burge (2001)Caira JN, Burge AN. Three new species of Acanthobothrium (Cestoda: Tetraphyllidea) from the ocellated electric ray, Diplobatis ommata, in the Gulf of California, México. Comp Parasitol 2001; 68(1): 52-65. and Caira & Zahner (2001)Caira JN, Zahner SD. Two new species of Acanthobothrium Beneden, 1849 (Tetraphyllidea: Onchobothriidae) from horn sharks in the Gulf of California, Mexico. Syst Parasitol 2001; 50(3): 219-229. PMid:11590308. http://dx.doi.org/10.1023/A:1012241913722.
http://dx.doi.org/10.1023/A:101224191372...
described nine species, all from the Gulf of California (Figure 1; Table 1). Only A. cartegenensis Brooks and Mayes, 1980 has been reported from the eastern coast of México (BROOKS & MAYES, 1980Brooks DR, Mayes MA. Cestodes in four species of euryhaline stingrays from Colombia. Proc Helminthol Soc Wash 1980; 47(1): 22-29.; MONKS et al., 2015Monks S, Pulido-Flores G, Lara-Sánchez M. Distribution extension of Acanthobothrium cartagenensis Brooks & Mayes, 1980 (Tetraphyllidea: Onchobothriidae) in Urobatis jamaicensis (Cuvier, 1816) (Myliobatiformes: Urotrygonidae) from Quintana Roo, México. Check List 2015; 11(4): 1-3. https://doi.org/10.15560/11.4.1707.
https://doi.org/10.15560/11.4.1707...
). To date, no members of the genus have been reported from the Mexican coasts of Gulf of Mexico, but Linton (1908)Linton EIX. Helminth fauna of the Dry Tortugas. I. Cestodes. In: Carnegie Institution of Washington. Papers from the Tortugas Laboratory. Washington: Carnegie Institution of Washington; 1908. p. 157-190., Goldstein (1964)Goldstein RJ. Species of Acanthobothrium (Cestoda: Tetraphyllidea) from the Gulf of Mexico. J Parasitol 1964; 50(5): 656-661. PMid:14215486. http://dx.doi.org/10.2307/3276123.
http://dx.doi.org/10.2307/3276123...
, Goldstein et al. (1969)Goldstein RJ, Henson RN, Schlicht FG. Acanthobothrium lintoni sp. n. (Cestoda: Tetraphyllidea) from the electric ray, Narcine brasiliensis (Olfers) in the Gulf of Mexico. Zool Anz 1969; 181(5-6): 435-438. and Vardo-Zalik & Campbell (2011)Vardo-Zalik AM, Campbell RA. Five new species of Acanthobothrium van Beneden, 1849 (Cestoda: Tetraphyllidea) in elasmobranchs from the northwest Atlantic and Gulf of Mexico with the first records from smooth-hound sharks and guitarfish. Zootaxa 2011; 2838: 41-64. collectively have reported nine species of Acanthobothrium from the northern coast of the Gulf of Mexico in waters of the USA. This suggests that members of the genus likely occur in more southern parts of the Gulf. As part of a parasitological survey of elasmobranchs in the Gulf of Mexico, adult cestodes were found in the spiral valve of Aetobatus cf. narinari (Euphrasen, 1790) that could be assigned to Acanthobothrium but not to any known species; this species is described herein. It represents the first record of the genus from the Mexican coast of the Gulf of Mexico.

The type localities of the species of Acanthobothrium reported from the coasts of Mexico (see Table 1); a and b refers to the sampling areas reported in this study, where ☒ is assigned to individuals of Aetobatus cf. narinari negative for A. marquesi n. sp. and ☑ for individuals of Aetobatus cf. narinari that were positive for the new species.
Species of Acanthobothrium reported from the Eastern Pacific Ocean and from the Western Atlantic Ocean of the Americas (amphi-American species). Numbers in parentheses for each taxon name refer to localities in Figure 1.
Materials and Methods
From 1999 to 2005 the digestive tracts of nine stingrays, Aetobatus cf. narinari (Euphrasen, 1790) (spotted eagle ray; raya pinta; arraia pintada) were purchased from local fishermen at four localities: five rays from Isla del Carmen, Campeche (three from open water North of Ciudad del Carmen and two rays from Laguna de Términos) (18°36’ N, 91°33’ W and 18°37’58”N; 91°49’57”W, respectively) (2000 and 2005); three specimens from Champotón, Campeche, (19°21’N; 90°54’W) (1999); and one specimen from Holbox, Quintana Roo, northwest of the island, (21°34’N; 86°14’W) (2000) (Insets a and b in Figure 1, Table 2). Cestodes were removed from the spiral valve of the host, killed with hot tap water, transferred immediately to AFA (alcohol-formalin-acetic acid) for 24-48 hr, and then stored in 70°GL ethanol. Specimens were stained either with Mayer’s carmalum or Ehrlich’s hematoxylin and mounted in Canada balsam for examination as whole mounts; worms that were too large to be mounted on a single slide were cut into sections and mounted sequentially on separate slides. Stained specimens were examined using a compound photomicroscope equipped with Nomarski differential interference contrast optics (Leica DM LB2). Taxonomic information for the host was taken from McEachran & Dunn (1998)McEachran JD, Dunn KA. Phylogenetic analysis of skates, a morphologically conservative clade of elasmobranchs (Chondrichthyes: Rajidae). Copeia 1998; 2(2): 271-290. http://dx.doi.org/10.2307/1447424.
http://dx.doi.org/10.2307/1447424...
, White et al. (2010)White WT, Last PR, Naylor GJP, Jensen K, Caira JN. Clarification of Aetobatus ocellatus (Kuhl, 1823) as a valid species, and a comparison with Aetobatus narinari (Euphrasen, 1790) (Rajiformes: Myliobatidae). CSIRO Marine Atmospheric Res Paper 2010; 32: 141-164., Richards et al. (2009)Richards VP, Henning M, Witzell W, Shivji MS. Species delineation and evolutionary history of the globally distributed spotted eagle ray (Aetobatus narinari). J Hered 2009; 100(3): 273-283. PMid:19304741. http://dx.doi.org/10.1093/jhered/esp005.
http://dx.doi.org/10.1093/jhered/esp005...
, Froese & Pauly (2017)Froese R, Pauly D. FishBase.World Wide Web electronic publication [online]. 2017 [cited 2017 Oct 5]. Available from: www.fishbase.org
www.fishbase.org...
. Some specimens were prepared for scanning electron microscopy (SEM) by dehydration in a graded ethanol series and low-vacuum dried. Dried specimens were mounted on aluminum stubs with carbon tape and grounded with carbon paint before being sputter-coated with approximately 100 Å of gold/palladium (about two minutes). Specimens were examined using a Jeol JSM-6300 scanning electron microscope. Illustrations were made with the aid of a drawing tube. Measurements are in micrometers unless specified otherwise, and expressed as length by width. For most characters, ranges are given, followed in parentheses by the sample mean and sample size (n). Mean values ± 1 standard deviation is provided for some characters (i.e., mean ± sd; n = sample size). Hook measurements follow Euzet (1959)Euzet L. Recherches sur les Cestodes Tétraphyllides des Sélaciens de côtes de France. Docteur ès Sciences Naturelles (Thesis). Montpellier: University of Montpellier; 1959. 263 p. as modified by Monks et al. (1996)Monks S, Brooks DR, Pérez-Ponce de Leon G. A new species of Acanthobothrium Van Beneden, 1849 (Eucestoda: Tetraphyllidea: Onchobothriidae) in Dasyatis longus Garman (Chondrichthyes: Myliobatiformes: Dasyatididae) from Chamela Bay, Jalisco, Mexico. J Parasitol 1996; 82(3): 484-488. PMid:8636857. http://dx.doi.org/10.2307/3284090.
http://dx.doi.org/10.2307/3284090...
(i.e., mean value ± 1 standard deviation is given followed by, in parentheses, the range).
The categorical method suggested by Ghoshroy & Caira (2001)Ghoshroy S, Caira JN. Four new species of Acanthobothrium (Cestoda: Tetraphyllidea) from the whiptail stingray Dasyatis brevis in the Gulf of California, Mexico. J Parasitol 2001; 87(2): 354-372. PMid:11318566. http://dx.doi.org/10.1645/0022-3395(2001)087[0354:FNSOAC]2.0.CO;2.
http://dx.doi.org/10.1645/0022-3395(2001...
and by Fyler & Caira (2006)Fyler CA, Caira JN. Five new species of Acanthobothrium (Tetraphyllidea: Onchobothriidae) from the freshwater stingray Himantura chaophraya (Batoidea: Dasyatidae) in Malaysian Borneo. J Parasitol 2006; 92(1): 105-125. PMid:16629324. http://dx.doi.org/10.1645/GE-3522.1.
http://dx.doi.org/10.1645/GE-3522.1...
was used to facilitate comparisons among species from the same geographic area in conjunction with the current literature. This method was proposed for the comparison and differentiation of species by categories based upon the combination of four variables: total length ≤ 15 mm = S (short) or > 15 mm = L (long); number of proglottids comprising the strobili ≤ 50 = F (few) or > 50 = M (many); number of testes per proglottid ≤ 80 = F (few) or > 80 = M (many); and ovarian lobes symmetrical = S or asymmetrical = A (GHOSHROY & CAIRA, 2001Ghoshroy S, Caira JN. Four new species of Acanthobothrium (Cestoda: Tetraphyllidea) from the whiptail stingray Dasyatis brevis in the Gulf of California, Mexico. J Parasitol 2001; 87(2): 354-372. PMid:11318566. http://dx.doi.org/10.1645/0022-3395(2001)087[0354:FNSOAC]2.0.CO;2.
http://dx.doi.org/10.1645/0022-3395(2001...
). Of the possible combinations the following 10 categories currently are recognized and coded as follows: 1 = SFFS; 2 = SFFA; 3 = LMMA; 4 = LMMS; 5 = LMFS; 6 = LMFA; 7 = LFFA; 8 = SMFS; 9 = LFFS; 10 = SMMS.
Specimens from three collections were examined: CNHE refers to the Colección Nacional de Helmintos, Instituto de Biología, Universidad Nacional Autónoma de México, Mexico City, México; CHE refers to the Colección de Helmintos, Centro de Investigaciones Biológicas, Universidad Autónoma del Estado de Hidalgo, Pachuca, México; and HWML refers to the Harold W. Manter Laboratory of Parasitology, University of Nebraska-Lincoln, Lincoln, Nebraska, USA.
Results
Survey data
Nine stingrays, all large adults, were collected from the four localities (Figure 1, insets a and b). Of these, two of three rays from Champotón (in 1999) were infected with two and six individuals of Acanthobothrium marquesi n. sp., two rays from south of Isla del Carmen in Laguna de Términos (collected in 2005) were infected with 10 and 12 worms, and one ray from north of Isla del Carmen was infected with 64 worms (Table 2). One ray collected in 2000 from Isla Holbox and two rays from the marine waters north of Isla del Carmen (2005) were uninfected.
Description
Acanthobothrium marquesi n. sp.

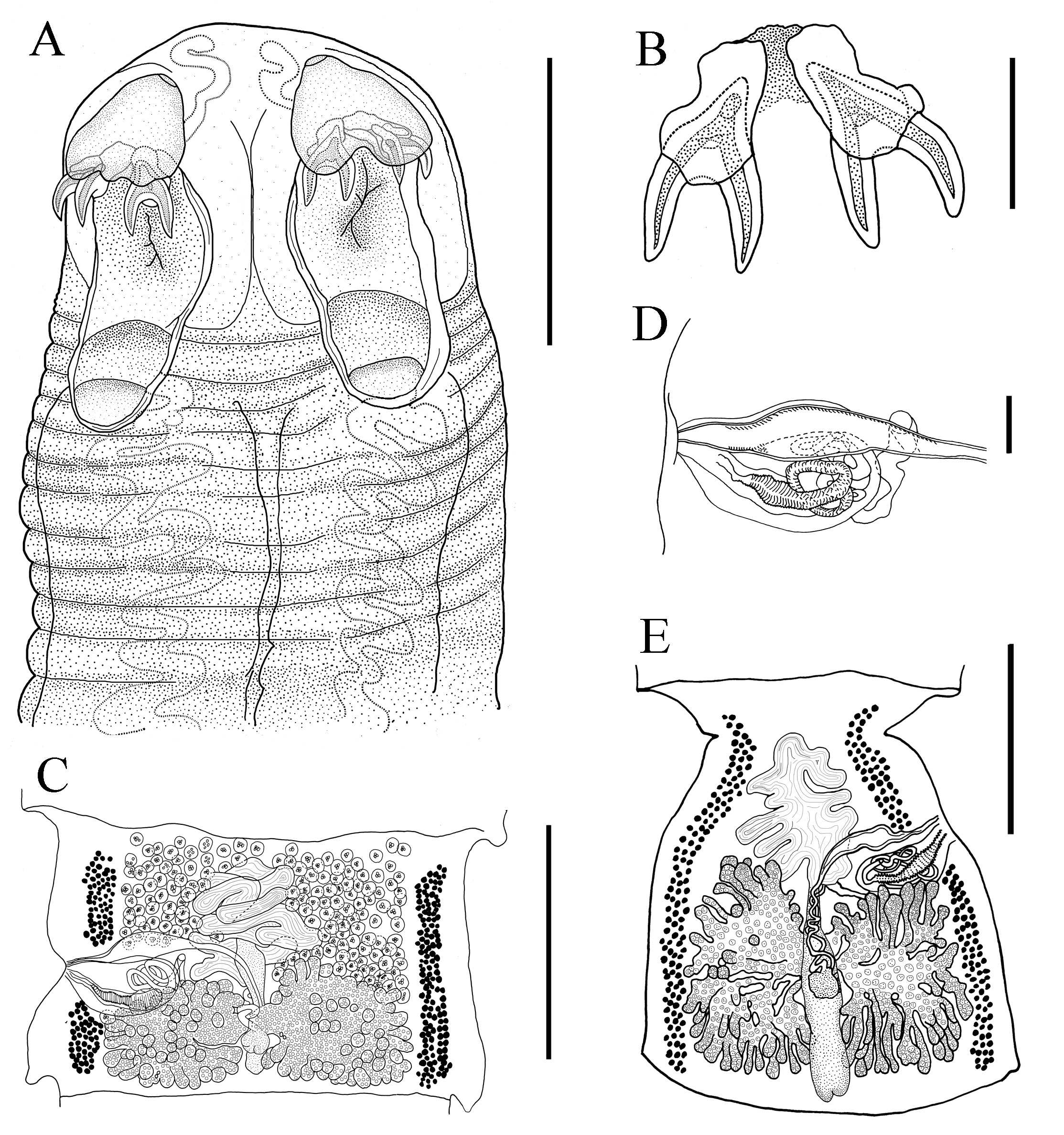
Acanthobothrium marquesi n. sp. A. Scolex. B. Hooks. C. Mature problotide. D. Cirrus sack. E. Terminal proglotide. Scale bar: A. 750 μm; B. 150 μm; C. 1000 μm; D. 133 μm; E. 880 μm.

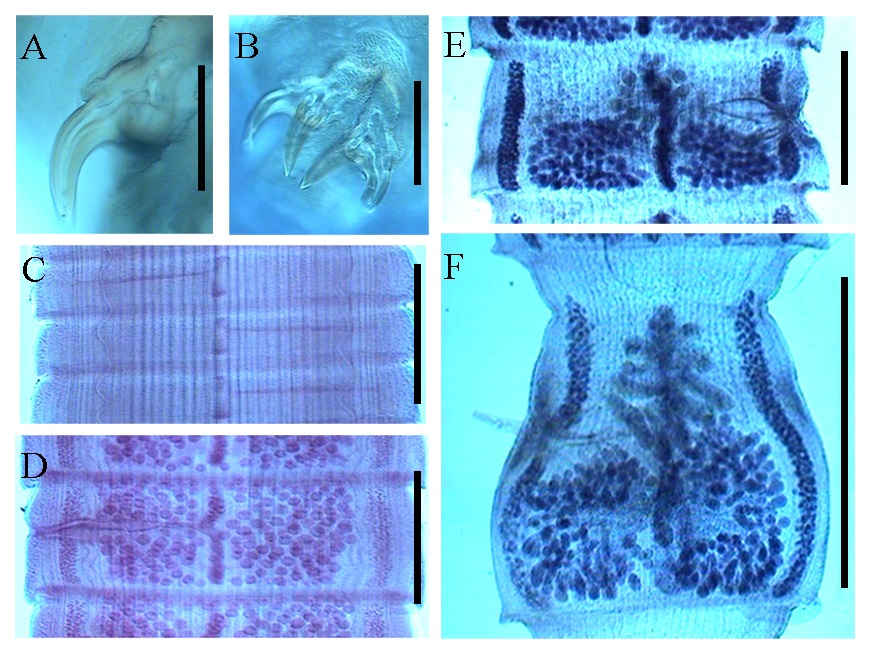
Photographic images of Acanthobothrium marquesi n. sp. from Aetobatus cf. narinari taken using a compound microscope equipped with normal light and Nomarski differential optics. A. Abaxial view of hook (lateral prong). B. Formal view of hooks with sclerotic plaques. C. Immature proglottid. D. Mature proglottid showing testicles. E. Mature proglottid. F. Terminal proglottid. Scale bars: A. 115 μm; B. 160 μm; C. 300 μm; D. 535 μm; E. 1170 μm; F. 1725 μm.

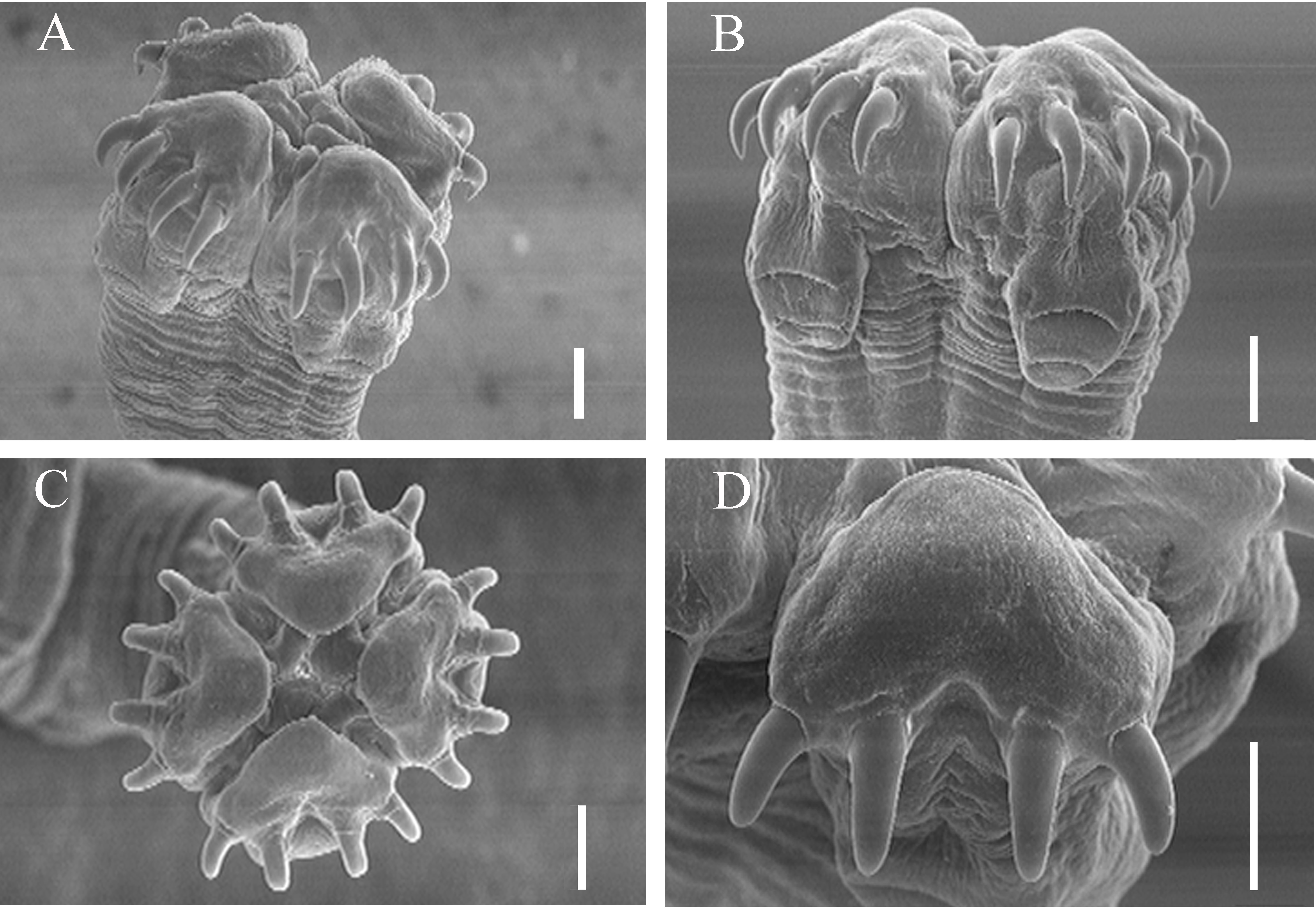
Scanning electron micrographs of Acanthobothrium marquesi n. sp. from Aetobatus cf. narinari A. Scolex. B. Front view of pad, hooks and bothridia. C. Superficial apical view. D. Front view of pad and hooks. Scale bars: A, B, C, D 100 μm.
Description: Based upon measurements of 10 complete worms and 20 partial worms mounted on slides, and two scoleces mounted on stubs for SEM. Strobila craspedote, anapolytic, 7.9 cm to 51.0 cm (26.1 cm ± 17.1 cm, n = 10) long; composed of 903-2,379 (1,549 ± 491, n = 11) proglottids. Scolex proper 500-1,000 (707 ± 151; n = 20) long by 670–1,130 (872 ± 122; n = 19) wide, composed of 4 sessile triloculate bothridia; each bothridium with apical sucker and pad, armed with pair of bifid hooks (Figures 2 -4). Bothridia with thick muscular walls; 490-915 (626 ± 136; n = 19) long by 233-338 (274 ± 27, n = 20) wide. Locular wall dividing anterior and middle loculi relatively thick and muscular; wall between middle and posterior loculi relatively thin. Anterior loculus 263–480 (373 ± 72, n = 20) long, middle loculus 75-150 (113 ± 22, n = 20) long, posterior loculus 88-140 (110 ± 15, n = 19) long (Figures 2 -4). Ratio of lengths of loculi 1:0.3:0.3.
Apical suckers not strongly defined, 118-170 (143 ± 15, n = 18) in diameter; pads muscular, 258-365 (309 ± 32, n = 19) wide. Posterior edge of apical muscular pads extending posteriorly over handles of hooks, divided in middle to form two straight extensions; posterior edge not well defined. Hooks bipronged, handle stout, extended almost directly opposite prongs (Figures 2 -4). Hook formula for external hooks (n = 18):
Hook formula for internal hooks (n = 16):
Cephalic peduncle unspined, 5,750-13,650 (9,255 ± 1,945, n = 21) long by 510-1,290 (742 ± 235, n = 21). Immature proglottids wider than long, 200-950 (468 ± 193, n = 23) long by 220-2,720 (1,655 ± 617, n = 24) wide (Figure 3); mature proglottids, 300-1,450 (763 ± 296, n = 23) long by 830-2,000 (1,405 ± 327, n = 23) wide (Figures 2 and 3); terminal proglottids 980-2,250 (1,388 ± 381, n = 13) long by 780-1,900 (1,152 ± 324, n = 12) wide (Figures 2 and 3). Testes 109–171 (138 ± 17, n = 18) in number, 25-45 (33 ± 7, n = 17) preporal, 14-39 (27 ± 9, n = 18) postporal, 62-95 (81 ± 8, n = 18) antiporal. Testes 43-150 (75 ± 26, n = 21) long by 40-130 (71 ± 24, n = 21) wide (Figures 2 and 3). Cirrus sac slightly posterior to mid-proglottid, extending medially to mid-proglottid, curved posteriorly in terminal proglottids, 325-750 (425 ± 90, n = 22) long, 100-450 (197 ± 76, n = 22) wide (Figures 2 and 3). Cirrus 556 μm long when extended, slightly swollen at base. Cirrus sac containing spined eversible cirrus. Genital atrium shallow. Genital pore located 41%-59% (51% ± 6%, n = 21) of total length of proglottid from anterior end, irregularly alternating. Vagina anterior to cirrus sac, vaginal wall glandular. Vagina canal can form loops, descends on the aporal, relatively to the midline of proglottid. Ovary near posterior end of proglottid, with foliose lobes. Shape in frontal view changing as proglottids mature: nearly U–shaped in immature proglottids, H–shaped in mature proglottids, and inverted A– or V–shaped in terminal proglottids. Ovary 460-1400 (974 ± 305, n = 22) wide at isthmus; arms unequal in length; in proglottids mature, and terminal the isthmus often located in middle position regarding the uterus, gravid proglottid not observed (Figures 2 and 3). Aporal arm 250-810 (435 ± 158, n = 15) long, extending to anterior margin of cirrus sac; poral arm 250-920 (473 ± 180, n = 15) long. Mehlis’ gland immediately posterior to ovarian isthmus; external seminal receptacle at level of or slightly anterior to isthmus (Figures 2 and 3). Vitelline follicles elongate oval in shape, 120-640 (305 ± 146, n = 21) wide, 160-640 (323 ± 136, n = 21) long, extending as narrow bands on each side of proglottid. Uterus saccate, narrowly elongate in terminal attached proglottids. Eggs not observed.
Taxonomic Summary
Type host: Aetobatus cf. narinari (Euphrasen, 1790); the host belongs to a currently unresolved species complex (RICHARDS et al., 2009Richards VP, Henning M, Witzell W, Shivji MS. Species delineation and evolutionary history of the globally distributed spotted eagle ray (Aetobatus narinari). J Hered 2009; 100(3): 273-283. PMid:19304741. http://dx.doi.org/10.1093/jhered/esp005.
http://dx.doi.org/10.1093/jhered/esp005...
; WHITE et al., 2010White WT, Last PR, Naylor GJP, Jensen K, Caira JN. Clarification of Aetobatus ocellatus (Kuhl, 1823) as a valid species, and a comparison with Aetobatus narinari (Euphrasen, 1790) (Rajiformes: Myliobatidae). CSIRO Marine Atmospheric Res Paper 2010; 32: 141-164.).
Site of infection: spiral valve.
Type locality: Laguna de Términos, Ciudad del Carmen, Campeche, México (18°35’19”N; 91°33’30”W).
Other locality: Champotón, Campeche, México (19°21’N; 90°54’W).
Holotype: CNHE–10554.
Paratypes: CNHE–10555 to 10556; HWML–139377 to 139384; CHE–P00061 to P00063.
Zoobank registration: B4DAA0D5-284B-4B66-9477-CDDEAA98B7BE (ZOOBANK, 2018The Official Registry of Zoological Nomenclature International – ZOOBANK. Acanthobothrium marquesi Rodríguez-Ibarra, Pulido-Flores, Violante-González & Monks 2018 [online]. Commission on Zoological Nomenclature; 2018 [cited 2018 Feb 8]. Available from: http://zoobank.org/References/B4DAA0D5-284B-4B66-9477-CDDEAA98B7BE
http://zoobank.org/References/B4DAA0D5-2...
)
Etymology: the species is named in honor of Dr. Fernando Marques (Departamento de Zoologia, Universidade de São Paulo, São Paulo, Brazil) for his friendship and for his contributions to our knowledge of the helminths of stingrays.
Remarks: There are 11 species reported from the Pacific Coast of México (Figure 1; Table 1); A. bajaensis Appy and Dailey, 1973; A. bullardi Ghoshroy and Caira, 2001; A. cleofanus Monks, Brooks, and Pérez-Ponce de León, 1996; A. dasi Ghoshroy and Caira, 2001; A. dollyae Caira and Burge, 2001; A. maryanskii Caira and Burge, 2001; A. puertecitense Caira and Zahner, 2001; A. rajivi Ghoshroy and Caira, 2001; A. royi Caira and Burge, 2001; A. santarosaliense Caira and Zahner, 2001; A. soberoni Ghoshroy and Caira, 2001 (CAIRA & BURGE, 2001Caira JN, Burge AN. Three new species of Acanthobothrium (Cestoda: Tetraphyllidea) from the ocellated electric ray, Diplobatis ommata, in the Gulf of California, México. Comp Parasitol 2001; 68(1): 52-65.; CAIRA & ZAHNER, 2001Caira JN, Zahner SD. Two new species of Acanthobothrium Beneden, 1849 (Tetraphyllidea: Onchobothriidae) from horn sharks in the Gulf of California, Mexico. Syst Parasitol 2001; 50(3): 219-229. PMid:11590308. http://dx.doi.org/10.1023/A:1012241913722.
http://dx.doi.org/10.1023/A:101224191372...
; GHOSHROY & CAIRA, 2001Ghoshroy S, Caira JN. Four new species of Acanthobothrium (Cestoda: Tetraphyllidea) from the whiptail stingray Dasyatis brevis in the Gulf of California, Mexico. J Parasitol 2001; 87(2): 354-372. PMid:11318566. http://dx.doi.org/10.1645/0022-3395(2001)087[0354:FNSOAC]2.0.CO;2.
http://dx.doi.org/10.1645/0022-3395(2001...
; MONKS et al., 1996Monks S, Brooks DR, Pérez-Ponce de Leon G. A new species of Acanthobothrium Van Beneden, 1849 (Eucestoda: Tetraphyllidea: Onchobothriidae) in Dasyatis longus Garman (Chondrichthyes: Myliobatiformes: Dasyatididae) from Chamela Bay, Jalisco, Mexico. J Parasitol 1996; 82(3): 484-488. PMid:8636857. http://dx.doi.org/10.2307/3284090.
http://dx.doi.org/10.2307/3284090...
). None of these have been reported from Aetobatus cf. narinari in Mexican waters.
Acanthobothrium cartagenensis Brooks and Mayes, 1980 is the only species that has been reported from the Eastern Coast of México (includes the Gulf of Mexico and Caribbean Sea). It can be distinguished from the new species because it is a Category 9 species (LFFS: L, >15 mm; F, ≤50 segments; F, ≤80 testes; S, left and right lobes symmetrical) (categories sensuGHOSHROY & CAIRA, 2001Ghoshroy S, Caira JN. Four new species of Acanthobothrium (Cestoda: Tetraphyllidea) from the whiptail stingray Dasyatis brevis in the Gulf of California, Mexico. J Parasitol 2001; 87(2): 354-372. PMid:11318566. http://dx.doi.org/10.1645/0022-3395(2001)087[0354:FNSOAC]2.0.CO;2.
http://dx.doi.org/10.1645/0022-3395(2001...
).
The new species is the third member of the genus reported from Aetobatus cf. narinari in the greater Atlantic Ocean. The species that have been reported from Aetobatus cf. narinari from the eastern Pacific Ocean are A. monksi Marques, Brooks, and Barriga, 1997, and A. nicoyaense Brooks and McCorquodale, 1995 (both are Category 1 species; i.e., much smaller than the new species). None of these species have been reported from Mexican waters (MARGUEE et al., 1997Marguee F, Brooks DR, Barriga R. Six species of Acanthobothrium (Eucestoda: Tetraphyllidea) in stingrays (Chondrichthyes: Rajiformes: Myliobatoidei) from Ecuador. J Parasitol 1997; 83(3): 475-484. PMid:9194831. http://dx.doi.org/10.2307/3284414.
http://dx.doi.org/10.2307/3284414...
; BROOKS & MCCORQUODALE, 1995Brooks DR, McCorquodale S. Acanthobothrium nicoyaense n. sp. (Eucestoda: Tetraphyllidea: Onchobothriidae) in Aetobatus narinari (Euphrasen) (Chondrichthyes: Myliobatiformes: Myliobatidae) from the Gulf of Nicoya, Costa Rica. J Parasitol 1995; 81(2): 244-246. PMid:7707201. http://dx.doi.org/10.2307/3283927.
http://dx.doi.org/10.2307/3283927...
). The species of Acanthobothrium that have been reported from Aetobatus cf. narinari from the western Atlantic Ocean are, A. colombianum Brooks and Mayes, 1980 and A. tortum (Linton, 1916) Baer and Euzet, 1962.
Acanthobothrium marquesi is a Category 3 species (LMMA) (sensuGHOSHROY & CAIRA, 2001Ghoshroy S, Caira JN. Four new species of Acanthobothrium (Cestoda: Tetraphyllidea) from the whiptail stingray Dasyatis brevis in the Gulf of California, Mexico. J Parasitol 2001; 87(2): 354-372. PMid:11318566. http://dx.doi.org/10.1645/0022-3395(2001)087[0354:FNSOAC]2.0.CO;2.
http://dx.doi.org/10.1645/0022-3395(2001...
): total length > 15 mm (specimens of the new species are 26,100 mm long); number of proglottids > 50 (average of 1,549 proglottids); number of testes per proglottid > 80 (138 testes per proglottid); and ovarian lobes asymmetrical (aporal lobe reaches anteriorly from the posterior end of the proglottid to about the anterior margin of the cirrus sac and the poral lobe reaches anteriorly to the posterior margin of the cirrus sac).
Acanthobothrium colombianum is a Category 9 species (L = >15 mm; F = ≤50 proglottids; F = ≤80 testes per proglottid; S = ovarian lobes symmetrical), differing from the new species in the number of proglottids, number of testes, and the symmetry of the ovarian lobes. Acanthobothrium monksi and A. nicoyaense are Category 1 species (S = ≤15 mm; F = ≤50 proglottids; F = ≤80 testes per proglottid; S = ovarian lobes symmetrical).
Acanthobothrium tortum, as described by Campbell (1970)Campbell RA. Notes on tetraphyllidean cestodes from the Atlantic Coast of North America, with descriptions of two new species. J Parasitol 1970; 56(3): 498-508. http://dx.doi.org/10.2307/3277613.
http://dx.doi.org/10.2307/3277613...
is a Category 3 species and is the most similar species in the Western Atlantic Ocean (Eastern coast of North America). Acanthobothrium marquesi n. sp. can be distinguished from A. tortum by being longer (26.1 cm vs. 10.6 cm), having a greater number of proglottids (1,549 proglottids vs. 656), having a larger scolex (872 long by 872 wide vs. 699 long by 665 wide), larger bothridia (626 long by 274 wide vs. 563 long by 238 wide). In specimens of the new species the sizes and relative proportions of the bothridia are different from those of A. tortum (373:113:110 = 1:0.3:0.3 vs. 259:87:117 = 1:0.3:0.5, respectively). Finally, the cirrus sac is larger in the new species (425 long by 197 wide vs. 350 long by 122 wide) and the number of testes is less (138 total = 33 preporal, 27 postporal, and 81 aporal vs. 163 total = 44 preporal, 32 postporal, and 87 aporal).
Other amphi-American Category 3 species are A. holorhini Alexander, 1953 and A. maculatum Riser, 1955, both from Myliobatis californica Gill; neither has been reported from México. These two species are smaller than the new species in all structures and the number of testes is less in the former two species than in the new species. Finally, these two species and their hosts are known only from the Pacific coast.
Discussion
Only seven species of Acanthobothrium have been reported previously from México, six species from the Pacific coast (A. bajaensis Appy and Dailey, 1973; A. bullardi Ghoshroy and Caira, 2001; A. cleofanus; A. dasi Ghoshroy and Caira, 2001; A. rajivi Ghoshroy and Caira, 2001; and A. soberoni Ghoshroy and Caira, 2001) and one from the Caribbean coast (A. cartagenensisBrooks and Mayes, 1980Brooks DR, Mayes MA. Cestodes in four species of euryhaline stingrays from Colombia. Proc Helminthol Soc Wash 1980; 47(1): 22-29.) (APPY & DAILEY, 1973Appy RG, Dailey MD. Two new species of Acanthobothrium (Cestoda: Tetraphyllidea) from elasmobranchs of the eastern Pacific. J Parasitol 1973; 59(5): 817-820. http://dx.doi.org/10.2307/3278414.
http://dx.doi.org/10.2307/3278414...
; MONKS et al., 1996Monks S, Brooks DR, Pérez-Ponce de Leon G. A new species of Acanthobothrium Van Beneden, 1849 (Eucestoda: Tetraphyllidea: Onchobothriidae) in Dasyatis longus Garman (Chondrichthyes: Myliobatiformes: Dasyatididae) from Chamela Bay, Jalisco, Mexico. J Parasitol 1996; 82(3): 484-488. PMid:8636857. http://dx.doi.org/10.2307/3284090.
http://dx.doi.org/10.2307/3284090...
, 2015Monks S, Pulido-Flores G, Lara-Sánchez M. Distribution extension of Acanthobothrium cartagenensis Brooks & Mayes, 1980 (Tetraphyllidea: Onchobothriidae) in Urobatis jamaicensis (Cuvier, 1816) (Myliobatiformes: Urotrygonidae) from Quintana Roo, México. Check List 2015; 11(4): 1-3. https://doi.org/10.15560/11.4.1707.
https://doi.org/10.15560/11.4.1707...
; GHOSHROY & CAIRA, 2001Ghoshroy S, Caira JN. Four new species of Acanthobothrium (Cestoda: Tetraphyllidea) from the whiptail stingray Dasyatis brevis in the Gulf of California, Mexico. J Parasitol 2001; 87(2): 354-372. PMid:11318566. http://dx.doi.org/10.1645/0022-3395(2001)087[0354:FNSOAC]2.0.CO;2.
http://dx.doi.org/10.1645/0022-3395(2001...
); A. marquesi is the eleventh species reported from México and the second species from the eastern coast of México.
The nominal species Aetobatus narinari (sensu Euphrasen) has classically been thought to be a globally-distributed species inhabiting tropical seas, but recent studies have suggested that it represents a species complex (RICHARDS et al., 2009Richards VP, Henning M, Witzell W, Shivji MS. Species delineation and evolutionary history of the globally distributed spotted eagle ray (Aetobatus narinari). J Hered 2009; 100(3): 273-283. PMid:19304741. http://dx.doi.org/10.1093/jhered/esp005.
http://dx.doi.org/10.1093/jhered/esp005...
; WHITE et al., 2010White WT, Last PR, Naylor GJP, Jensen K, Caira JN. Clarification of Aetobatus ocellatus (Kuhl, 1823) as a valid species, and a comparison with Aetobatus narinari (Euphrasen, 1790) (Rajiformes: Myliobatidae). CSIRO Marine Atmospheric Res Paper 2010; 32: 141-164.; WHITE et al., 2013White WT, Furumitsu K, Yamaguchi A. A new species of eagle ray Aetobatus narutobiei from the Northwest Pacific: an example of the critical role taxonomy plays in fisheries and ecological sciences. PLoS One 2013; 8(12): e83785. PMid:24391827. http://dx.doi.org/10.1371/journal.pone.0083785.
http://dx.doi.org/10.1371/journal.pone.0...
), an hypothesis supported by parasitological studies (MARIE & JUSTINE, 2005Marie AD, Justine J-L. Monocotylids (Monogenea: Monopisthocotylea) from Aetobatus cf. narinari off New Caledonia, with a description of Decacotyle elpora n. sp. Syst Parasitol 2005; 60(3): 175-185. PMid:15864455. http://dx.doi.org/10.1007/s11230-004-6345-7.
http://dx.doi.org/10.1007/s11230-004-634...
; MARIE & JUSTINE, 2006Marie AD, Justine J-L. Thaumatocotyle pseudodasybatis Hargis, 1955 (Monogenea: Monocotylidae) from Aetobatus cf. narinari, with a comparison of specimens from Australia, French Polynesia and New Caledonia. Syst Parasitol 2006; 64(1): 47-55. PMid:16612660. http://dx.doi.org/10.1007/s11230-005-9017-3.
http://dx.doi.org/10.1007/s11230-005-901...
). No molecular data exist for the particular populations of Aetobatus cf. narinari that are hosts to A. tortum and A. marquesi n. sp., but they are on the opposite north-south coasts of the Gulf of México; the presence of different species of Acanthobothrium in each suggests that these two population of Aetobatus cf. narinari do not mix. The study of divergence of cryptic populations of species that serve as hosts and that of their parasites is in initial processes, but studies of water currents in the Gulf of México/Caribbean Sea region suggest that more isolated species of helminths are waiting to be discovered (SHULMAN & BERMINGHAM, 1995Shulman MJ, Bermingham E. Early life histories, ocean currents, and the population genetics of caribbean reef fishes. Evolution 1995; 49(5): 897-910. PMid:28564869. http://dx.doi.org/10.1111/j.1558-5646.1995.tb02325.x.
http://dx.doi.org/10.1111/j.1558-5646.19...
; CARTON & CHAO, 1999Carton JA, Chao Y. Caribbean Sea eddies inferred from TOPEX/POSEIDON altimetry and a 1/6º Atlantic Ocean model simulation. J Geophys Res 1999; 104(C4): 7743-7752. http://dx.doi.org/10.1029/1998JC900081.
http://dx.doi.org/10.1029/1998JC900081...
; SANDOVAL-CASTILLO & ROCHA-OLIVARES, 2011Sandoval-Castillo J, Rocha-Olivares A. Deep mitochondrial divergence in Baja California populations of an aquilopelagic elasmobranch: the golden cownose ray. J Hered 2011; 102(3): 269-274. PMid:21414964. http://dx.doi.org/10.1093/jhered/esr004.
http://dx.doi.org/10.1093/jhered/esr004...
).
Members of Aetobatus Blainville, 1816 traditionally have been considered to be part of Myliobatidae (FROESE & PAULY, 2017Froese R, Pauly D. FishBase.World Wide Web electronic publication [online]. 2017 [cited 2017 Oct 5]. Available from: www.fishbase.org
www.fishbase.org...
). However, White & Naylor (2016)White WT, Naylor GJP. Resurrection of the family Aetobatidae (Myliobatiformes) for the pelagic eagle rays, genus Aetobatus. Zootaxa 2016; 4139(3): 435-438. PMid:27470816. http://dx.doi.org/10.11646/zootaxa.4139.3.10.
http://dx.doi.org/10.11646/zootaxa.4139....
recently moved the genus to Aetobatidae. Although this arrangement has begun to be followed (LAST et al, 2016Last PR, White WT, Carvalho MR, Séret B, Stehman MFW, Naylor GJP, et al. Rays of the world. Ithaca: Comstock Publishing Associates; 2016.), we chose to follow the most widely accepted classification (FROESE & PAULY, 2017Froese R, Pauly D. FishBase.World Wide Web electronic publication [online]. 2017 [cited 2017 Oct 5]. Available from: www.fishbase.org
www.fishbase.org...
) pending further evaluations of the status of the family.
Acknowledgements
The material for this study was collected during the fieldwork of GP-F as part of her Doctorate in Science studies in Ecology and Sustainable Development at El Colegio de la Frontera Sur during 1999-2001 (CONACyT Scholarship No. 137558). Likewise, ER-I thanks CONACyT for a doctoral scholarship (No. 626250); this study is part of his Doctoral studies in Biodiversity and Conservation, Universidad Autónoma del Estado de Hidalgo. The authors would like to thank the curators of the HWML and CNHE (Scott L. Gardner and Luis García-Prieto, respectively) for providing access to laboratories and specimens.
References
- Appy RG, Dailey MD. Two new species of Acanthobothrium (Cestoda: Tetraphyllidea) from elasmobranchs of the eastern Pacific. J Parasitol 1973; 59(5): 817-820. http://dx.doi.org/10.2307/3278414
» http://dx.doi.org/10.2307/3278414 - Brooks DR, Mayes MA. Cestodes in four species of euryhaline stingrays from Colombia. Proc Helminthol Soc Wash 1980; 47(1): 22-29.
- Brooks DR, McCorquodale S. Acanthobothrium nicoyaense n. sp. (Eucestoda: Tetraphyllidea: Onchobothriidae) in Aetobatus narinari (Euphrasen) (Chondrichthyes: Myliobatiformes: Myliobatidae) from the Gulf of Nicoya, Costa Rica. J Parasitol 1995; 81(2): 244-246. PMid:7707201. http://dx.doi.org/10.2307/3283927
» http://dx.doi.org/10.2307/3283927 - Caira JN, Burge AN. Three new species of Acanthobothrium (Cestoda: Tetraphyllidea) from the ocellated electric ray, Diplobatis ommata, in the Gulf of California, México. Comp Parasitol 2001; 68(1): 52-65.
- Caira JN, Jensen K. Planetary biodiversity inventory (2008-2017): tapeworms from vertebrate bowels of the earth Lawrence, Kansas: The University of Kansas, Museum of Natural History; 2017. Special Publication, n. 25.
- Caira JN, Zahner SD. Two new species of Acanthobothrium Beneden, 1849 (Tetraphyllidea: Onchobothriidae) from horn sharks in the Gulf of California, Mexico. Syst Parasitol 2001; 50(3): 219-229. PMid:11590308. http://dx.doi.org/10.1023/A:1012241913722
» http://dx.doi.org/10.1023/A:1012241913722 - Campbell RA. Notes on tetraphyllidean cestodes from the Atlantic Coast of North America, with descriptions of two new species. J Parasitol 1970; 56(3): 498-508. http://dx.doi.org/10.2307/3277613
» http://dx.doi.org/10.2307/3277613 - Carton JA, Chao Y. Caribbean Sea eddies inferred from TOPEX/POSEIDON altimetry and a 1/6º Atlantic Ocean model simulation. J Geophys Res 1999; 104(C4): 7743-7752. http://dx.doi.org/10.1029/1998JC900081
» http://dx.doi.org/10.1029/1998JC900081 - Euzet L. Recherches sur les Cestodes Tétraphyllides des Sélaciens de côtes de France. Docteur ès Sciences Naturelles (Thesis). Montpellier: University of Montpellier; 1959. 263 p.
- Froese R, Pauly D. FishBase.World Wide Web electronic publication [online]. 2017 [cited 2017 Oct 5]. Available from: www.fishbase.org
» www.fishbase.org - Fyler CA, Caira JN. Five new species of Acanthobothrium (Tetraphyllidea: Onchobothriidae) from the freshwater stingray Himantura chaophraya (Batoidea: Dasyatidae) in Malaysian Borneo. J Parasitol 2006; 92(1): 105-125. PMid:16629324. http://dx.doi.org/10.1645/GE-3522.1
» http://dx.doi.org/10.1645/GE-3522.1 - Ghoshroy S, Caira JN. Four new species of Acanthobothrium (Cestoda: Tetraphyllidea) from the whiptail stingray Dasyatis brevis in the Gulf of California, Mexico. J Parasitol 2001; 87(2): 354-372. PMid:11318566. http://dx.doi.org/10.1645/0022-3395(2001)087[0354:FNSOAC]2.0.CO;2
» http://dx.doi.org/10.1645/0022-3395(2001)087[0354:FNSOAC]2.0.CO;2 - Goldstein RJ, Henson RN, Schlicht FG. Acanthobothrium lintoni sp. n. (Cestoda: Tetraphyllidea) from the electric ray, Narcine brasiliensis (Olfers) in the Gulf of Mexico. Zool Anz 1969; 181(5-6): 435-438.
- Goldstein RJ. Species of Acanthobothrium (Cestoda: Tetraphyllidea) from the Gulf of Mexico. J Parasitol 1964; 50(5): 656-661. PMid:14215486. http://dx.doi.org/10.2307/3276123
» http://dx.doi.org/10.2307/3276123 - Last PR, White WT, Carvalho MR, Séret B, Stehman MFW, Naylor GJP, et al. Rays of the world Ithaca: Comstock Publishing Associates; 2016.
- Linton EIX. Helminth fauna of the Dry Tortugas. I. Cestodes. In: Carnegie Institution of Washington. Papers from the Tortugas Laboratory. Washington: Carnegie Institution of Washington; 1908. p. 157-190.
- Marie AD, Justine J-L. Monocotylids (Monogenea: Monopisthocotylea) from Aetobatus cf. narinari off New Caledonia, with a description of Decacotyle elpora n. sp. Syst Parasitol 2005; 60(3): 175-185. PMid:15864455. http://dx.doi.org/10.1007/s11230-004-6345-7
» http://dx.doi.org/10.1007/s11230-004-6345-7 - Marie AD, Justine J-L. Thaumatocotyle pseudodasybatis Hargis, 1955 (Monogenea: Monocotylidae) from Aetobatus cf. narinari, with a comparison of specimens from Australia, French Polynesia and New Caledonia. Syst Parasitol 2006; 64(1): 47-55. PMid:16612660. http://dx.doi.org/10.1007/s11230-005-9017-3
» http://dx.doi.org/10.1007/s11230-005-9017-3 - Marguee F, Brooks DR, Barriga R. Six species of Acanthobothrium (Eucestoda: Tetraphyllidea) in stingrays (Chondrichthyes: Rajiformes: Myliobatoidei) from Ecuador. J Parasitol 1997; 83(3): 475-484. PMid:9194831. http://dx.doi.org/10.2307/3284414
» http://dx.doi.org/10.2307/3284414 - McEachran JD, Dunn KA. Phylogenetic analysis of skates, a morphologically conservative clade of elasmobranchs (Chondrichthyes: Rajidae). Copeia 1998; 2(2): 271-290. http://dx.doi.org/10.2307/1447424
» http://dx.doi.org/10.2307/1447424 - Monks S, Brooks DR, Pérez-Ponce de Leon G. A new species of Acanthobothrium Van Beneden, 1849 (Eucestoda: Tetraphyllidea: Onchobothriidae) in Dasyatis longus Garman (Chondrichthyes: Myliobatiformes: Dasyatididae) from Chamela Bay, Jalisco, Mexico. J Parasitol 1996; 82(3): 484-488. PMid:8636857. http://dx.doi.org/10.2307/3284090
» http://dx.doi.org/10.2307/3284090 - Monks S, Pulido-Flores G, Lara-Sánchez M. Distribution extension of Acanthobothrium cartagenensis Brooks & Mayes, 1980 (Tetraphyllidea: Onchobothriidae) in Urobatis jamaicensis (Cuvier, 1816) (Myliobatiformes: Urotrygonidae) from Quintana Roo, México. Check List 2015; 11(4): 1-3. https://doi.org/10.15560/11.4.1707
» https://doi.org/10.15560/11.4.1707 - Richards VP, Henning M, Witzell W, Shivji MS. Species delineation and evolutionary history of the globally distributed spotted eagle ray (Aetobatus narinari). J Hered 2009; 100(3): 273-283. PMid:19304741. http://dx.doi.org/10.1093/jhered/esp005
» http://dx.doi.org/10.1093/jhered/esp005 - Sandoval-Castillo J, Rocha-Olivares A. Deep mitochondrial divergence in Baja California populations of an aquilopelagic elasmobranch: the golden cownose ray. J Hered 2011; 102(3): 269-274. PMid:21414964. http://dx.doi.org/10.1093/jhered/esr004
» http://dx.doi.org/10.1093/jhered/esr004 - Shulman MJ, Bermingham E. Early life histories, ocean currents, and the population genetics of caribbean reef fishes. Evolution 1995; 49(5): 897-910. PMid:28564869. http://dx.doi.org/10.1111/j.1558-5646.1995.tb02325.x
» http://dx.doi.org/10.1111/j.1558-5646.1995.tb02325.x - The Official Registry of Zoological Nomenclature International – ZOOBANK. Acanthobothrium marquesi Rodríguez-Ibarra, Pulido-Flores, Violante-González & Monks 2018 [online]. Commission on Zoological Nomenclature; 2018 [cited 2018 Feb 8]. Available from: http://zoobank.org/References/B4DAA0D5-284B-4B66-9477-CDDEAA98B7BE
» http://zoobank.org/References/B4DAA0D5-284B-4B66-9477-CDDEAA98B7BE - Vardo-Zalik AM, Campbell RA. Five new species of Acanthobothrium van Beneden, 1849 (Cestoda: Tetraphyllidea) in elasmobranchs from the northwest Atlantic and Gulf of Mexico with the first records from smooth-hound sharks and guitarfish. Zootaxa 2011; 2838: 41-64.
- White WT, Furumitsu K, Yamaguchi A. A new species of eagle ray Aetobatus narutobiei from the Northwest Pacific: an example of the critical role taxonomy plays in fisheries and ecological sciences. PLoS One 2013; 8(12): e83785. PMid:24391827. http://dx.doi.org/10.1371/journal.pone.0083785
» http://dx.doi.org/10.1371/journal.pone.0083785 - White WT, Last PR, Naylor GJP, Jensen K, Caira JN. Clarification of Aetobatus ocellatus (Kuhl, 1823) as a valid species, and a comparison with Aetobatus narinari (Euphrasen, 1790) (Rajiformes: Myliobatidae). CSIRO Marine Atmospheric Res Paper 2010; 32: 141-164.
- White WT, Naylor GJP. Resurrection of the family Aetobatidae (Myliobatiformes) for the pelagic eagle rays, genus Aetobatus. Zootaxa 2016; 4139(3): 435-438. PMid:27470816. http://dx.doi.org/10.11646/zootaxa.4139.3.10
» http://dx.doi.org/10.11646/zootaxa.4139.3.10
Publication Dates
-
Publication in this collection
12 Mar 2018 -
Date of issue
Jan-Mar 2018
History
-
Received
05 Oct 2017 -
Accepted
30 Jan 2018