Abstract
To describe the parasitic community of the Magellanic Horned Owl, Bubo magellanicus (Aves, Strigiformes), 19 carcasses from central Chile were analyzed. Ectoparasites were collected through plumage inspection, while endoparasites were collected through traditional techniques of parasitological necropsy. Sixteen owls were infected with at least one species of ectoparasite (84.21%) or endoparasite (31.58%). Eleven of 19 birds (57.89%) harbored feather mites of the three species Pandalura cirrata (42.11%), Glaucalges attenuatus (47.37%), and Kramerella sp. (10.53%), whereas 16 individuals (84.21%) harbored the chewing louse Strigiphilus chilensis. Only six birds (31.58%) were infected with helminths; the nematodes Capillaria tenuissima (26.32%) and Dispharynx nasuta (5.26%); the acanthocephalan Centrorhynchus spinosus (5.26%); and the trematode Neodiplostomum sp. (5.26%). Apart from S . chilensis, all parasites comprised new records for B . magellanicus.
Keywords:
Birds; parasites; Phthiraptera; Acari; helminths
Resumo
Para descrever a comunidade parasitária de coruja-orelhuda Bubo magellanicus (Aves, Strigiformes), foram analisados 19 carcaças das aves do centro do Chile. Os ectoparasitos foram coletados inspecionando-se a plumagem e os endoparasitas extraídos por meio de técnicas tradicionais de necropsia parasitaria. Dezesseis corujas estavam infectadas com pelo menos uma espécie de ectoparasito (84,21%) ou endoparasito (31,58%). Onze de 19 aves (57,89%) abrigavam nas penas ácaros de três espécies: Pandalura cirrata (42,11%), Glaucalges attenuatus (47,37%) e Kramerella sp. (10,53%), enquanto que 16 indivíduos (84,21%) estavam parasitados pelo piolho Strigiphilus chilensis . Apenas seis aves (31,58%) estavam infectadas com helmintos; os nematoides Capillaria tenuissima (26,32%) e Dispharynx nasuta (5,26%); o acantocéfalo Centrorhynchus spinosus (5,26%); e o trematódeo Neodiplostomum sp. (5,26%). Excetuando-se S. chilensis , todos os parasitos incluíam novos registros para B. magellanicus .
Palavras-chave:
Pássaros; parasita; Phthiraptera; Acari; helmintos
Introduction
The Magellanic Horned Owl, Bubo magellanicus (Lesson, 1828), is the largest of the five owl species that inhabit Chile. Its distribution in Chile extends from the Arica and Parinacota region (18°28’30” S, 70°18’52” W) to Tierra del Fuego and Cape Horn (Magallanes region, 54°56’00” S, 67°37’00” W), although these limits are not well defined ( FIGUEROA et al., 2015 Figueroa RA, Alvarado S, Corales S, González-Acuña D, Schlatter R, Martínez D. Los búhos de Chile. In: Enríquez PL. Los búhos neotropicales diversidad y conservación. México: El Colegio de la Frontera Sur; 2015. p. 173-273. ). This owl species inhabits a wide range of habitats, including semi-open forests abundant in Nothofagus and semi-arid areas, which has an altitude of up to 4,500 meters above sea level ( KÖNIG et al., 2008 König C, Weick F, Becking JH. Owls of the world. 2nd ed. London: Christopher Helm Publisher; 2008. ). The owl’s main prey includes rodents, birds, and arthropods ( FIGUEROA et al., 2015 Figueroa RA, Alvarado S, Corales S, González-Acuña D, Schlatter R, Martínez D. Los búhos de Chile. In: Enríquez PL. Los búhos neotropicales diversidad y conservación. México: El Colegio de la Frontera Sur; 2015. p. 173-273. ). The Magellanic Horned Owl was previously considered as a subspecies of the Great Horned Owl, B. virginianus (Gmelin, 1788), until König et al. (1996) König C, Heidrich P, Wink M. Zur taxonomie der Uhus (Strigidae: Bubo spp.) im südlichen Südamerika. Stuttgarter Beitr Naturk 1996; 540: 1-9. showed differences between these two birds based on vocal, morphologic, and genetic evidence, which confirmed that this bird was a separate species ( KÖNIG et al., 2008 König C, Weick F, Becking JH. Owls of the world. 2nd ed. London: Christopher Helm Publisher; 2008. ).
Only two studies have previously focused on the parasites of this bird. Strigiphilus chilensis Carriker, 1966 was described from specimens collected in Santiago, and González-Acuña et al. (2006) González-Acuña D, Muñoz C R, Cicchino A, Figueroa RAR. Lice of Chilean owls: a first description. J Raptor Res 2006; 40(4): 301-302. http://dx.doi.org/10.3356/0892-1016(2006)40[301:LOCOAF]2.0.CO;2.
http://dx.doi.org/10.3356/0892-1016(200...
found this species on B. magellanicus from central and south-central Chile. The aim of this study is to analyze ecto- and endoparasites from this host in central and southern Chile.
Methods
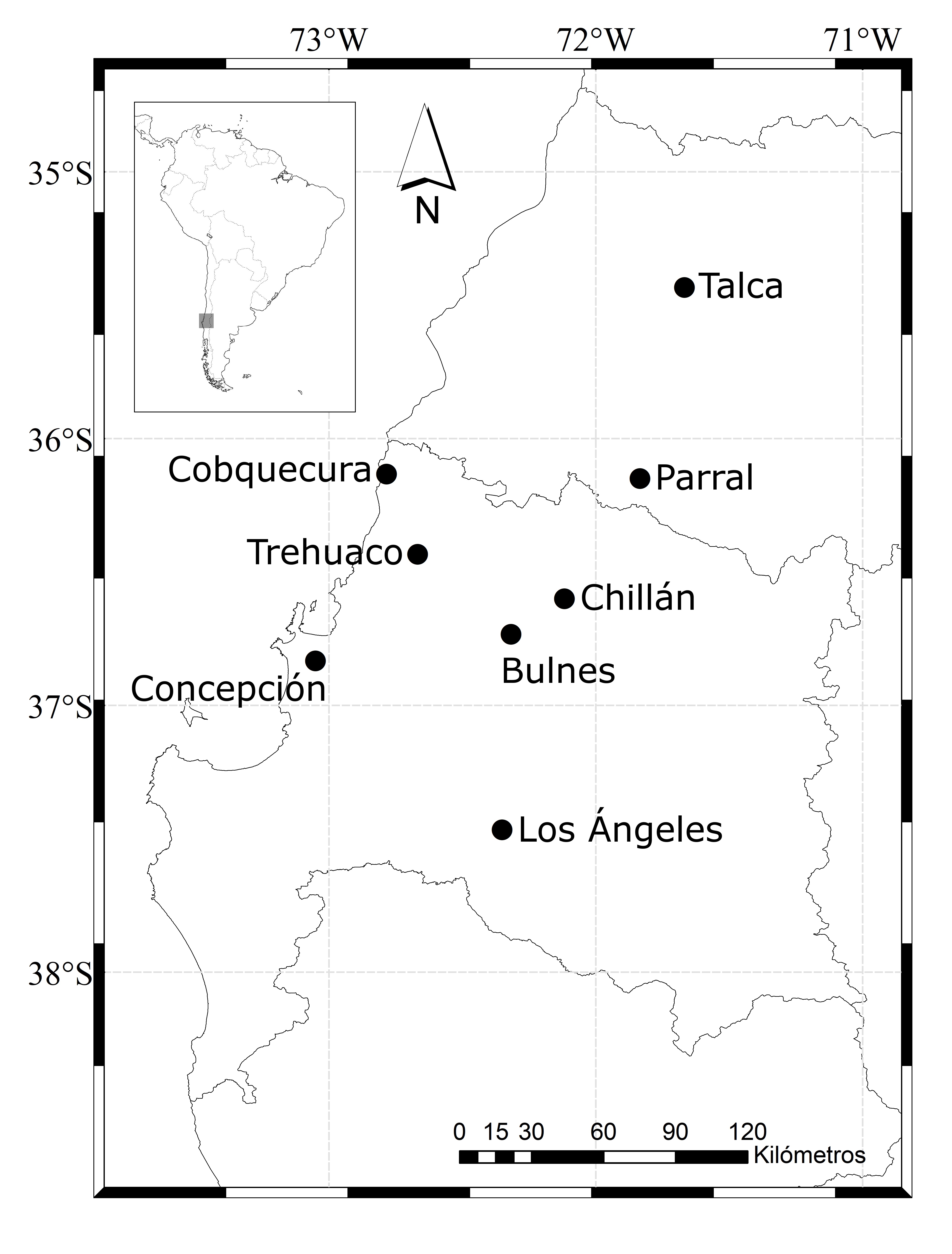
Between 2010 and 2017, 19 specimens of B. magellanicus were collected. The causes of death were mainly attributed to collisions with vehicles, poaching, and poisoning. Eight birds came from Chillán (36°36’ S, 72°07’ W), two from Bulnes (36°44’ S, 72°18’ W), and one from each of the following localities: Concepción (36°50’ S, 73°03’ W); Cobquecura (36°08’ S, 72°47’ W); Los Ángeles (37°28’ S, 72°21’ W); Parral (36°09’ S, 71°50’ W); Talca (35°26’ S, 71°40’ W); and Trehuaco (36°26’ S, 72°40’ W) ( Figure 1 ). The animals used were frozen in bags and then necropsied. The ectoparasites were collected by visually inspecting feathers and they were subsequently preserved in 70% ethanol. Lice were cleared and mounted in Canada balsam, as described by Palma (1978) Palma R. Slide-mounting of lice: a detailed description of the Canada balsam technique. NZ Entomol 1978; 6(4): 432-436. http://dx.doi.org/10.1080/00779962.1978.9722313.
http://dx.doi.org/10.1080/00779962.1978...
and Price et al. (2003) Price RD, Hellenthal RA, Palma RL, Johnson KP, Clayton DH. The chewing lice: world checklist and biological overview. Springfield: Illinois Natural History Survey Special Publication; 2003. . Mites were cleared in Nesbitt’s solution for 72 hours at sub-boiling temperature, and they were finally mounted in Berlese’s medium ( KRANTZ & WALTER, 2009 Krantz GW, Walter DE. A manual of acarology. Lubbock: Texas Tech University Press; 2009. ). The collection and preparation of helminths followed the technique proposed by Kinsella & Forrester (1972) Kinsella JM, Forrester DJ. Helminth parasites of the Florida duck, Anas platyrhynchos fulvigula. Proc Helminthol Soc Wash 1972; 39(2): 173-176. . Nematodes and acanthocephalans were cleared in temporary mounts of lacto-phenol; they were then identified and returned to the preservative.

The taxonomical keys used to identify feather lice followed Clayton and Price (1984) Clayton DH, Price RD. Taxonomy of the Strigiphilus cursitans group (Ischnocera: Philopteridae), parasites of the owls (Strigiformes). Ann Entomol Soc Am 1984; 77(4): 340-363. http://dx.doi.org/10.1093/aesa/77.4.340.
http://dx.doi.org/10.1093/aesa/77.4.340...
and Price et al. (2003) Price RD, Hellenthal RA, Palma RL, Johnson KP, Clayton DH. The chewing lice: world checklist and biological overview. Springfield: Illinois Natural History Survey Special Publication; 2003. ; those for feather mites followed Gaud (1980) Gaud J. Acariens Sarcoptiformes plumicoles parasites sur les oiseaux Psittaciformes, Strigiformes et Caprimulgiformes en Afrique. Tervuren: Musée Royal de l’Afrique Centrale; 1980. (Annales Serie in 8° Sciences Zoologiques; vol. 230). , Atyeo & Philips (1984) Atyeo WT, Philips JR. The feather mite genus Neopetitota (Pterolichoidea: Kramerellidae). J Med Entomol 1984; 21(4): 409-411. http://dx.doi.org/10.1093/jmedent/21.4.409.
http://dx.doi.org/10.1093/jmedent/21.4....
, Gaud & Atyeo (1996) Gaud J, Atyeo WT. Feather mites of the World (Acarina, Astigmata): the supraspecific taxa. Tervuren: Koninklijk Museum voor Midden-Afrika; 1996. , Krantz & Walter (2009) Krantz GW, Walter DE. A manual of acarology. Lubbock: Texas Tech University Press; 2009. , and Mironov (2011) Mironov SV. Feather mites of the genus Pandalura Hull (Astigmata: Psoroptoididae) from owls and caprimulgiforms. Proc Zool Inst RAS 2011; 315(1): 19-37. ; and those for helminths followed Goble & Kutz (1945) Goble FC, Kutz HL. The genus Dispharynx (Nematoda: Acuariidae) in Galliform and Passeriforms birds. J Parasitol 1945; 31(5): 323-331. http://dx.doi.org/10.2307/3273088.
http://dx.doi.org/10.2307/3273088 ...
, Mettrick (1959) Mettrick DF. On the nematode genus Capillaria in British birds. Ann Mag Nat Hist 1959; 2(14): 65-84. http://dx.doi.org/10.1080/00222935908651029.
http://dx.doi.org/10.1080/0022293590865...
, Yamaguti (1963) Yamaguti S. Systema Helminthum. vol. V. New York: Interscience Publisher; 1963. , Skrjabin (1969) Skrjabin K. Key to parasitic nematodes. Israel: Program for Scientific Translations; 1969. , Hong & Shoop (1994) Hong ST, Shoop WL. Neodiplostomum seoulensis n. comb. (Trematoda: Neodiplostomidae). J Parasitol 1994; 80(4): 660-663. http://dx.doi.org/10.2307/3283209. PMid:8064539.
http://dx.doi.org/10.2307/3283209 ...
, Richardson & Nickol (1995) Richardson DJ, Nickol BB. The Genus Centrorhynchus (Acanthocephala) in North America with Description of Centrorhynchus robustus n. sp., redescription of Centrorhynchus conspectus, and a Key to Species. J Parasitol 1995; 81(5): 767-772. http://dx.doi.org/10.2307/3283971. PMid:7472871.
http://dx.doi.org/10.2307/3283971 ...
, and Gómez-Puerta et al. (2009) Gómez-Puerta LA, Enciso MA, Rojas G. Parasitismo natural por Synhimantus (Dispharynx) nasuta (Nematoda: Acuariidae) en Pavo real (Pavo cristatus) en cautiverio. Rev Peru Biol 2009; 16(1): 121-123. .
All collected specimens were stored in the collection at the Laboratory of Zoology, Faculty of Veterinary Science, University of Concepción.
Results and Discussion
All the analyzed birds presented at least one species of parasite. A total of 16/19 (prevalence of 84.21%) of the owls were found infected with ectoparasites and a total of 6/19 (prevalence of 31.57%) were found infected with endoparasites ( Table 1 ). Except for S. chilensis, all of the parasites that were found represented new records for B. magellanicus.
Summary of external and gastrointestinal parasites found in 19 Magellanic Horned Owls, from central Chile.
Acari
A total of 87 specimens of Pandalura cirrata ( MÜLLER, 1860 Müller J. Beitrag zur mährischen Arachnidenfauna. Naturwissenschaftliche Zeitschrift hrsg. vom Deutschen naturwissenschaftlich-medizinischen Verein für Böhmen. Lotos 1860; 10: 44-55. ) ( Figures 2
and 3 ) were collected; they had a prevalence of 42.11% (8/19). This mite belongs to a genus that contains four species, two of which are exclusive to Strigiformes, whereas the other two are associated with Caprimuliformes ( MIRONOV, 2011 Mironov SV. Feather mites of the genus Pandalura Hull (Astigmata: Psoroptoididae) from owls and caprimulgiforms. Proc Zool Inst RAS 2011; 315(1): 19-37. ), as well as with the families Podargidae and Steatornithidae. The ectoparasite P. cirrata was originally described from the Eurasian Eagle-Owl Bubo bubo Linnaeus, 1758 by Müller (1860) Müller J. Beitrag zur mährischen Arachnidenfauna. Naturwissenschaftliche Zeitschrift hrsg. vom Deutschen naturwissenschaftlich-medizinischen Verein für Böhmen. Lotos 1860; 10: 44-55. as Dermaleichus cirratus, and later reassigned to the genus Pandalura Hull, 1934. Like all representatives of the family Psoroptoididae, P. cirrata have the typical appearance of the inhabitants of feathers, which are characterized by having a flattened and moderately widened idiosome, two pairs anterior legs with spines and hook-like structures to join the feathers ( DABERT & MIRONOV, 1999 Dabert J, Mironov SV. Origin and evolution of feather mites (Astigmata). Exp Appl Acarol 1999; 23(6): 437-454. http://dx.doi.org/10.1023/A:1006180705101.
http://dx.doi.org/10.1023/A:10061807051...
). To date, this mite was recently redescribed based on samples obtained from B. bubo in Spain, and it was also reported from B. virginianus in Canada ( MIRONOV, 2011 Mironov SV. Feather mites of the genus Pandalura Hull (Astigmata: Psoroptoididae) from owls and caprimulgiforms. Proc Zool Inst RAS 2011; 315(1): 19-37. ).


Glaucalges attenuatus (Buchholz, 1869) ( Figures 4
and 5 ) had the highest prevalence, mean intensity, and mean abundance among mites, with 131 individuals collected, and being present in half of the birds analyzed (9/19) (47.37). This genus includes the species G. attenuatus and G. tytonis Dabert et al. 2008 that infect Strigiformes, and G. pteropus Gaud et Mouchet, 1959, which was found parasitizing the family Musophagidae ( PHILIPS, 2000 Philips JR. A review and checklist of the parasitic mites (Acarina) of the Falconiformes and Strigiformes. J Raptor Res 2000; 34(3): 210-231. ). Glaucalges attenuatus are characterized by having a flattened and moderately widened idiosome, lateral setae and relatively long caudal, with the previous pairs of legs provided with hooks and spines ( DABERT & MIRONOV, 1999 Dabert J, Mironov SV. Origin and evolution of feather mites (Astigmata). Exp Appl Acarol 1999; 23(6): 437-454. http://dx.doi.org/10.1023/A:1006180705101.
http://dx.doi.org/10.1023/A:10061807051...
). Besides, it is apparently a generalist in host ( DABERT et al., 2008 Dabert J, Ehrnsberger R, Dabert M. Glaucalges tytonis sp. n. (Analgoidea, Xolalgidae) from the barn owl Tyto alba (Strigiformes, Tytonidae): compiling morphology with DNA barcode data for taxon descriptions in mites (Acari). Zootaxa 2008; 1719: 41-52. ); it can occupy both the primaries and secondaries of the wings and the body feathers, presumably explaining the high population parameters that were documented.


Kramerella sp. ( Figures 6
and 7 ) presented the lowest population parameters of the mites; only 11 mites were collected and they had a prevalence of 10.53% (2/19). The genus Kramerella Trouessart, 1916 is presently composed of 14 known species that exclusively affect the bird orders Falconiformes and Strigiformes ( KRANTZ, 1978 Krantz GW. A manual of acarology. Corvallis: Oregon State University; 1978. ; GAUD, 1980 Gaud J. Acariens Sarcoptiformes plumicoles parasites sur les oiseaux Psittaciformes, Strigiformes et Caprimulgiformes en Afrique. Tervuren: Musée Royal de l’Afrique Centrale; 1980. (Annales Serie in 8° Sciences Zoologiques; vol. 230). ; SOHN & NOH, 1994 Sohn BO, Noh YT. Feather mites Kramerellidae and Pterolichidae in Korea (Acari, Pterolichoidea). Korean J Parasitol 1994; 32(2): 75-83. http://dx.doi.org/10.3347/kjp.1994.32.2.75. PMid:8025036.
http://dx.doi.org/10.3347/kjp.1994.32.2...
). Contrary to our findings, mites of this genus are usually abundant on their hosts and primarily infect only the primary feathers of wings ( PHILIPS, 2000 Philips JR. A review and checklist of the parasitic mites (Acarina) of the Falconiformes and Strigiformes. J Raptor Res 2000; 34(3): 210-231. ). The small number of mites collected likely hampered thorough taxonomic identification.


Phthiraptera
Strigiphilus chilensis Carriker, 1966 ( Figures 8
and 9 ) is the only feather louse known to infect B. magellanicus and it has only been reported from this owl to date. This louse presented with a prevalence of 84.21% (16/19), with 315 lice collected. Within the order Phthiraptera, three genera infect owls but, interestingly, the genus Strigiphilus Mjöberg, 1910, which currently includes 52 species, has been exclusively found on Strigiformes ( CLAYTON, 1990 Clayton DH. Host specificity of Strigiphilus owl lice (Ischnocera: Philopteridae), with the description of new species and host associations. J Med Entomol 1990; 27(3): 257-265. http://dx.doi.org/10.1093/jmedent/27.3.257. PMid:2332870.
http://dx.doi.org/10.1093/jmedent/27.3....
). In this genus, males and females of S. chilensis present a longer head comparatively to other species ( CARRIKER, 1966 Carriker MA. New Species and Records of Mallophaga (Insecta) from Neotropical Owls (Strigiformes). Am Midl Nat 1966; 76(1): 74-99. http://dx.doi.org/10.2307/2423236.
http://dx.doi.org/10.2307/2423236 ...
). In other hand, besides the original description documented by Carriker (1966) Carriker MA. New Species and Records of Mallophaga (Insecta) from Neotropical Owls (Strigiformes). Am Midl Nat 1966; 76(1): 74-99. http://dx.doi.org/10.2307/2423236.
http://dx.doi.org/10.2307/2423236 ...
, where this mite was found from this owl in Santiago (Metropolitan Region), the only records of this louse on this host was made by González-Acuña et al. (2006) González-Acuña D, Muñoz C R, Cicchino A, Figueroa RAR. Lice of Chilean owls: a first description. J Raptor Res 2006; 40(4): 301-302. http://dx.doi.org/10.3356/0892-1016(2006)40[301:LOCOAF]2.0.CO;2.
http://dx.doi.org/10.3356/0892-1016(200...
in central and southern Chile (a detailed description of the provenance of the birds is given in that study).


Nematoda
Capillaria tenuissima Rudolphi, 1803 ( Figures 10
and 11 ) had a prevalence of 26.32% (5/19), with 12 individuals collected. This parasite has an indirect life cycle with earthworms as intermediate hosts and small rodents as definitive or paratenic hosts ( RUBILAR et al., 1996 Rubilar L, Bertossi E, Skewes O. Gastrointestinal parasitism in the common partridge (Nothoprocta perdicaria) in the Nuble Zone, Chile. Bol Chil Parasitol 1996; 51(1-2): 35-37. PMid:9196955. ). Some of the important characteristics of this species is that in males the spicule sheath is finely striated transversally and in the case of females its cuticle is covered by a characteristic high pattern ( METTRICK, 1959 Mettrick DF. On the nematode genus Capillaria in British birds. Ann Mag Nat Hist 1959; 2(14): 65-84. http://dx.doi.org/10.1080/00222935908651029.
http://dx.doi.org/10.1080/0022293590865...
). This parasite is common in owls, including the closely related B. virginianus Gmelin, 1788 and it always infects the intestine ( BORGSTEEDE et al., 2003 Borgsteede FH, Okulewicz A, Zoun PE, Okulewicz J. The helminth fauna of birds of prey (Accipitriformes, Falconiformes and Strigiformes) in the Netherlands. Acta Parasitol 2003; 48(3): 200-207. ). Capillaria falconis Goeze, 1782 has also been found in B. virginianus ( KINSELLA et al., 2001 Kinsella JM, Foster GW, Forrester DJ. Parasitic helminths of five species of owls from Florida, U.S.A. Comp Parasitol 2001; 68(1): 130-134. ). In Chile, C. tenuissima was recorded in Chimango Caracara Milvago chimango Vieillot, 1816 sampled in Chillán, Ñuble region ( SAN MARTÍN et al., 2006 San Martín J, Brevis C, Rubilar L, Krone O, González-Acuña D. Parasitismo gastrointestinal en tiuque común Milvago chimango chimango (VIEILLOT, 1816) (Falconidae, Aves) en Ñuble, Chile. Parasitol Latinoam 2006; 61(1-2): 63-68. http://dx.doi.org/10.4067/S0717-77122006000100009.
http://dx.doi.org/10.4067/S0717-7712200...
).


Dispharynx nasuta Rudolphi, 1819 had a prevalence of 5.26% with six parasites collected. This parasite is a generalist with an indirect life cycle of 57 days ( SCHOCK & COOPER, 1978 Schock RC, Cooper R. Internal parasitisms in captive birds. Mod Vet Pract 1978; 59(6): 439-443. PMid:672879. ). This nematode is characterized by a whitish color with a cuticle with finely transverse striations ( OYARZÚN-RUIZ et al., 2016 Oyarzún-Ruiz P, Basso E, Medel J, Muñoz-Alvarado P. Parásitos: los organismos relegados de nuestra biodiversidad. La Chiricoca 2016; 21: 9-15. ) with a muscular and glandular esophagus ( GÓMEZ-PUERTA et al., 2009 Gómez-Puerta LA, Enciso MA, Rojas G. Parasitismo natural por Synhimantus (Dispharynx) nasuta (Nematoda: Acuariidae) en Pavo real (Pavo cristatus) en cautiverio. Rev Peru Biol 2009; 16(1): 121-123. ). The life cycle of D. nasuta commonly requires an intermediate host from the order Isopoda; however, other invertebrates can also be used ( MOORE et al., 1988 Moore J, Freehling M, Crawford J, Cole P. Dispharynx nasuta (Nematoda) in California Quail (Callipepla californica) in Western Oregon. J Wildl Dis 1988; 24(3): 564-567. http://dx.doi.org/10.7589/0090-3558-24.3.564. PMid:3411717.
http://dx.doi.org/10.7589/0090-3558-24....
). According to Yamaguti (1961) Yamaguti S. Systema Helminthum. vol. III. New York: Interscience Publisher; 1961. , the Common Rough Woodlouse (Porcellio scaber Latreille, 1804) is the principal intermediate host for this parasite. Once it reaches the definitive host, it can be found in the proventriculus, esophagus, or small intestine. This parasite was found in phylogenetically distant hosts, as represented by Galliformes, Columbiformes ( BAKER, 2008 Baker DG. Flynn's parasites of laboratory animals. 2nd ed. Hoboken: Blackwell Publishing; 2008. ), and Strigiformes. In this last order, examples of hosts come from both the Old and New World, with Little Owl Athene noctua from Spain ( ILLESCAS GOMEZ et al., 1993 Illescas Gomez MP, Rodríguez Osario M, Aranda Maza F. Parasitation of Falconiform Strigiform and Passeriform (Corvidae) birds by helminths in Spain. Res Rev Parasitol 1993; 53(3-4): 129-135. ) and Eastern Screech Owl, Megascops asio Linnaeus, 1758 from Florida (United States) ( KINSELLA et al., 2001 Kinsella JM, Foster GW, Forrester DJ. Parasitic helminths of five species of owls from Florida, U.S.A. Comp Parasitol 2001; 68(1): 130-134. ), respectively. In Chile, this parasite has also been collected from a wide range of hosts: the Rock Pigeon Columba livia Gmelin, 1789 ( TORO et al., 1999 Toro H, Saucedo C, Borie C, Gough RE, Alcaíno H. Health status of free-living pigeons in the city of Santiago. Avian Pathol 1999; 28(6): 619-623. http://dx.doi.org/10.1080/03079459994416.
http://dx.doi.org/10.1080/0307945999441...
), the California Quail Callipepla californica Shaw, 1798 ( GONZÁLEZ-ACUÑA et al., 2000 González-Acuña D, Skewes-Ramm O, Rubilar-Contreras L, Daugschies A, Pohlmeyer K. Endoparásitos de codorniz (Callipepla californica) en Ñuble (Chile). Boletín Chileno de Ornitología 2000; 7: 23-25. ), the Southern Lapwing Vanellus chilensis Molina, 1782 ( GONZÁLEZ-ACUÑA et al., 2008 González-Acuña D, Olmedo P, Ciccino A. Parásitos de Vanellus chilensis all correspond to new records for this bird in Chile (AVES, CHARADRIIDAE) en Chillán, centrosur de Chile. Boletín Chileno de Ornitología 2008; 14(1): 36-48 ), and the diurnal raptor M. chimango ( OYARZÚN-RUIZ, 2013 Oyarzún-Ruiz PE. Parásitos gastrointestinales del Tiuque Milvago chimango temucoensis Vieillot, 1816, región de los Ríos, Chile [thesis]. Valdivia: Universidad Austral De Chile, Facultad De Ciencias Veterinarias Instituto de Patología Animal; 2013. ).
Acanthocephala
Centrorynchus spinosus Kaiser, 1893 had a prevalence of 5.26% (1/19) with nine individuals collected. Regarding the distinctive characteristics of C. spinosus they present a proboscis with 8-11 (usually 9 in female, 10 in male) longitudinal anterior hooks in each row and a trunk with anterior constriction, but with a lack of visible inflation ( RICHARDSON & NICKOL, 1995 Richardson DJ, Nickol BB. The Genus Centrorhynchus (Acanthocephala) in North America with Description of Centrorhynchus robustus n. sp., redescription of Centrorhynchus conspectus, and a Key to Species. J Parasitol 1995; 81(5): 767-772. http://dx.doi.org/10.2307/3283971. PMid:7472871.
http://dx.doi.org/10.2307/3283971 ...
). The life cycle of this parasite has cockroaches and other arthropods as intermediate hosts, while amphibians, reptiles, and rodents are paratemic hosts; when it reaches the small intestine of birds, the birds become definitive hosts ( OYARZÚN-RUIZ et al., 2016 Oyarzún-Ruiz P, Basso E, Medel J, Muñoz-Alvarado P. Parásitos: los organismos relegados de nuestra biodiversidad. La Chiricoca 2016; 21: 9-15. ).
This acanthocephalan has been reported in the USA ( VAN CLEAVE, 1916 Van Cleave HJ. Acanthocephala of the genera Centrorhynchus and llediorhynchus (new genus) from North American birds. Tr Am Nlicr Soc 1916; 35(4): 221-232. http://dx.doi.org/10.2307/3221908.
http://dx.doi.org/10.2307/3221908 ...
), Ecuador ( VAN CLEAVE, 1940 Van Cleave HJ. The Acanthocephala collected by the Allan Hancock Pacific Expedition, 1934. Rep Hancock Pacific Expedition (1932-1938) 1940; 2: 501-527. ), and Russia ( BYKHOVSKAIA, 1948 Bykhovskaia IE. Acanthocephala of birds of USSR. [in russian]. Parasit Sborn Zool Inst Akad Nauk 1948; 10: 245-257. ), in various species of birds, however, among the Strigiformes that have been recorded as hosts of C. spinosus are Barred Owl, Strix varia Barton, 1799, M. asio and, more frequently, B. virginianus ( KINSELLA et al., 2001 Kinsella JM, Foster GW, Forrester DJ. Parasitic helminths of five species of owls from Florida, U.S.A. Comp Parasitol 2001; 68(1): 130-134. ) all these records made only in USA. Therefore, the present finding means that the parasite can find suitable environmental conditions for its development, including in intermediate hosts, all along the Americas.
Trematoda
Neodiplostomum sp. was the helminth that presented with the lowest population parameters; two parasites were found in one bird (5.26%). Species of the genus Neodiplostomum Railliet, 1919 infects both mammals and birds ( HONG & SHOOP, 1994 Hong ST, Shoop WL. Neodiplostomum seoulensis n. comb. (Trematoda: Neodiplostomidae). J Parasitol 1994; 80(4): 660-663. http://dx.doi.org/10.2307/3283209. PMid:8064539.
http://dx.doi.org/10.2307/3283209 ...
). Nevertheless, some parasite species within the genus appear to be specific to a certain group of hosts; for instance, Richardson & Kinsella (2010) Richardson DJ, Kinsella JM. New host and distribution records for gastrointestinal parasites of raptors from Connecticut, U.S.A. Comp Parasitol 2010; 77(1): 72-82. http://dx.doi.org/10.1654/4411.1.
http://dx.doi.org/10.1654/4411.1 ...
noted that Neodiplostomum delicatum Chandler & Rausch, 1947 is restricted to avian hosts from the order Strigiformes. The species of Neodiplostomum recorded from the genus Bubo included N. americanum ( CHANDLER & RAUSCH, 1947 Chandler AC, Rausch R. A study of strigeids from owls in North Central United States. Trans Am Microsc Soc 1947; 66(3): 283-292. http://dx.doi.org/10.2307/3223396.
http://dx.doi.org/10.2307/3223396 ...
; WOODYARD et al., 2017 Woodyard ET, Rosser TG, Griffin MJ. New data on Neodiplostomum americanum Chandler and Rausch, 1947 (Digenea: Diplostomidae), in the Great Horned Owl Bubo virginianus Gmelin, 1788 and the Eastern Screech Owl Megascops asio Linnaeus, 1758 in Mississippi, USA. Parasitol Res 2017; 116(8): 2075-2089. http://dx.doi.org/10.1007/s00436-017-5503-1. PMid:28634620.
http://dx.doi.org/10.1007/s00436-017-55...
), N. reflexum ( CHANDLER & RAUSCH, 1947 Chandler AC, Rausch R. A study of strigeids from owls in North Central United States. Trans Am Microsc Soc 1947; 66(3): 283-292. http://dx.doi.org/10.2307/3223396.
http://dx.doi.org/10.2307/3223396 ...
; GALLAS & SILVEIRA, 2013 Gallas M, Silveira EF. Neodiplostomum reflexum Chandler & Rausch, 1947 (Digenea, Diplostomidae) em Bubo virginianus Gmelin, 1788 (Aves, Strigidae): primeiro registro para o estado do Rio Grande do Sul, Brasil. Cad Pesqui Sér Biol 2013; 25(1): 50-58. ), and N. delicatum ( KINSELLA et al., 2001 Kinsella JM, Foster GW, Forrester DJ. Parasitic helminths of five species of owls from Florida, U.S.A. Comp Parasitol 2001; 68(1): 130-134. ). The poor condition of these two collected specimens makes species identification impossible.
Conclusion
Bubo magellanicus presents a wide parasitic diversity. Of the eight species of parasites found the following species: Glaucalges attenuatus, Pandalura cirrata, Kramellera sp., Strigiphilus chilensis , Dispharynx nasuta, Capillaria tenuissima , Centrorhynchus spinosus and Neodiplostomum sp.; all correspond to new records for this bird in Chile, with the exception of S . chilensis.
Acknowledgements
We are deeply grateful for the support granted by members of the National Forest Corporation, CONAF, and the Agricultural and Livestock Service, SAG, of Chile. We also thank Karen Ardiles, Carolina Silva, Sebastián Muñoz, Daniela Doussang, Karolina Araya, Marescla Martínez, Danny Fuentes, Felix Varas, Ignacia Najle, Nicolás Fernández, Pablo Olmedo, Diego Barrientos and Sofia González for all their support in the field and laboratory. This research study was funded by FONDECYT, Projects 1130948 and 1170972.
References
- Atyeo WT, Philips JR. The feather mite genus Neopetitota (Pterolichoidea: Kramerellidae). J Med Entomol 1984; 21(4): 409-411. http://dx.doi.org/10.1093/jmedent/21.4.409.
» http://dx.doi.org/10.1093/jmedent/21.4.409 - Baker DG. Flynn's parasites of laboratory animals 2nd ed. Hoboken: Blackwell Publishing; 2008.
- Borgsteede FH, Okulewicz A, Zoun PE, Okulewicz J. The helminth fauna of birds of prey (Accipitriformes, Falconiformes and Strigiformes) in the Netherlands. Acta Parasitol 2003; 48(3): 200-207.
- Bykhovskaia IE. Acanthocephala of birds of USSR. [in russian]. Parasit Sborn Zool Inst Akad Nauk 1948; 10: 245-257.
- Carriker MA. New Species and Records of Mallophaga (Insecta) from Neotropical Owls (Strigiformes). Am Midl Nat 1966; 76(1): 74-99. http://dx.doi.org/10.2307/2423236.
» http://dx.doi.org/10.2307/2423236 - Chandler AC, Rausch R. A study of strigeids from owls in North Central United States. Trans Am Microsc Soc 1947; 66(3): 283-292. http://dx.doi.org/10.2307/3223396.
» http://dx.doi.org/10.2307/3223396 - Clayton DH, Price RD. Taxonomy of the Strigiphilus cursitans group (Ischnocera: Philopteridae), parasites of the owls (Strigiformes). Ann Entomol Soc Am 1984; 77(4): 340-363. http://dx.doi.org/10.1093/aesa/77.4.340.
» http://dx.doi.org/10.1093/aesa/77.4.340 - Clayton DH. Host specificity of Strigiphilus owl lice (Ischnocera: Philopteridae), with the description of new species and host associations. J Med Entomol 1990; 27(3): 257-265. http://dx.doi.org/10.1093/jmedent/27.3.257. PMid:2332870.
» http://dx.doi.org/10.1093/jmedent/27.3.257 - Dabert J, Ehrnsberger R, Dabert M. Glaucalges tytonis sp. n. (Analgoidea, Xolalgidae) from the barn owl Tyto alba (Strigiformes, Tytonidae): compiling morphology with DNA barcode data for taxon descriptions in mites (Acari). Zootaxa 2008; 1719: 41-52.
- Dabert J, Mironov SV. Origin and evolution of feather mites (Astigmata). Exp Appl Acarol 1999; 23(6): 437-454. http://dx.doi.org/10.1023/A:1006180705101.
» http://dx.doi.org/10.1023/A:1006180705101 - Figueroa RA, Alvarado S, Corales S, González-Acuña D, Schlatter R, Martínez D. Los búhos de Chile. In: Enríquez PL. Los búhos neotropicales diversidad y conservación México: El Colegio de la Frontera Sur; 2015. p. 173-273.
- Gallas M, Silveira EF. Neodiplostomum reflexum Chandler & Rausch, 1947 (Digenea, Diplostomidae) em Bubo virginianus Gmelin, 1788 (Aves, Strigidae): primeiro registro para o estado do Rio Grande do Sul, Brasil. Cad Pesqui Sér Biol 2013; 25(1): 50-58.
- Gaud J, Atyeo WT. Feather mites of the World (Acarina, Astigmata): the supraspecific taxa. Tervuren: Koninklijk Museum voor Midden-Afrika; 1996.
- Gaud J. Acariens Sarcoptiformes plumicoles parasites sur les oiseaux Psittaciformes, Strigiformes et Caprimulgiformes en Afrique. Tervuren: Musée Royal de l’Afrique Centrale; 1980. (Annales Serie in 8° Sciences Zoologiques; vol. 230).
- Goble FC, Kutz HL. The genus Dispharynx (Nematoda: Acuariidae) in Galliform and Passeriforms birds. J Parasitol 1945; 31(5): 323-331. http://dx.doi.org/10.2307/3273088.
» http://dx.doi.org/10.2307/3273088 - Gómez-Puerta LA, Enciso MA, Rojas G. Parasitismo natural por Synhimantus (Dispharynx) nasuta (Nematoda: Acuariidae) en Pavo real (Pavo cristatus) en cautiverio. Rev Peru Biol 2009; 16(1): 121-123.
- González-Acuña D, Muñoz C R, Cicchino A, Figueroa RAR. Lice of Chilean owls: a first description. J Raptor Res 2006; 40(4): 301-302. http://dx.doi.org/10.3356/0892-1016(2006)40[301:LOCOAF]2.0.CO;2.
» http://dx.doi.org/10.3356/0892-1016(2006)40[301:LOCOAF]2.0.CO;2 - González-Acuña D, Olmedo P, Ciccino A. Parásitos de Vanellus chilensis all correspond to new records for this bird in Chile (AVES, CHARADRIIDAE) en Chillán, centrosur de Chile. Boletín Chileno de Ornitología 2008; 14(1): 36-48
- González-Acuña D, Skewes-Ramm O, Rubilar-Contreras L, Daugschies A, Pohlmeyer K. Endoparásitos de codorniz (Callipepla californica) en Ñuble (Chile). Boletín Chileno de Ornitología 2000; 7: 23-25.
- Hong ST, Shoop WL. Neodiplostomum seoulensis n. comb. (Trematoda: Neodiplostomidae). J Parasitol 1994; 80(4): 660-663. http://dx.doi.org/10.2307/3283209. PMid:8064539.
» http://dx.doi.org/10.2307/3283209 - Illescas Gomez MP, Rodríguez Osario M, Aranda Maza F. Parasitation of Falconiform Strigiform and Passeriform (Corvidae) birds by helminths in Spain. Res Rev Parasitol 1993; 53(3-4): 129-135.
- Kinsella JM, Forrester DJ. Helminth parasites of the Florida duck, Anas platyrhynchos fulvigula. Proc Helminthol Soc Wash 1972; 39(2): 173-176.
- Kinsella JM, Foster GW, Forrester DJ. Parasitic helminths of five species of owls from Florida, U.S.A. Comp Parasitol 2001; 68(1): 130-134.
- König C, Heidrich P, Wink M. Zur taxonomie der Uhus (Strigidae: Bubo spp.) im südlichen Südamerika. Stuttgarter Beitr Naturk 1996; 540: 1-9.
- König C, Weick F, Becking JH. Owls of the world. 2nd ed. London: Christopher Helm Publisher; 2008.
- Krantz GW, Walter DE. A manual of acarology Lubbock: Texas Tech University Press; 2009.
- Krantz GW. A manual of acarology Corvallis: Oregon State University; 1978.
- Mettrick DF. On the nematode genus Capillaria in British birds. Ann Mag Nat Hist 1959; 2(14): 65-84. http://dx.doi.org/10.1080/00222935908651029.
» http://dx.doi.org/10.1080/00222935908651029 - Mironov SV. Feather mites of the genus Pandalura Hull (Astigmata: Psoroptoididae) from owls and caprimulgiforms. Proc Zool Inst RAS 2011; 315(1): 19-37.
- Moore J, Freehling M, Crawford J, Cole P. Dispharynx nasuta (Nematoda) in California Quail (Callipepla californica) in Western Oregon. J Wildl Dis 1988; 24(3): 564-567. http://dx.doi.org/10.7589/0090-3558-24.3.564. PMid:3411717.
» http://dx.doi.org/10.7589/0090-3558-24.3.564 - Müller J. Beitrag zur mährischen Arachnidenfauna. Naturwissenschaftliche Zeitschrift hrsg. vom Deutschen naturwissenschaftlich-medizinischen Verein für Böhmen Lotos 1860; 10: 44-55.
- Oyarzún-Ruiz P, Basso E, Medel J, Muñoz-Alvarado P. Parásitos: los organismos relegados de nuestra biodiversidad. La Chiricoca 2016; 21: 9-15.
- Oyarzún-Ruiz PE. Parásitos gastrointestinales del Tiuque Milvago chimango temucoensis Vieillot, 1816, región de los Ríos, Chile [thesis]. Valdivia: Universidad Austral De Chile, Facultad De Ciencias Veterinarias Instituto de Patología Animal; 2013.
- Palma R. Slide-mounting of lice: a detailed description of the Canada balsam technique. NZ Entomol 1978; 6(4): 432-436. http://dx.doi.org/10.1080/00779962.1978.9722313.
» http://dx.doi.org/10.1080/00779962.1978.9722313 - Philips JR. A review and checklist of the parasitic mites (Acarina) of the Falconiformes and Strigiformes. J Raptor Res 2000; 34(3): 210-231.
- Price RD, Hellenthal RA, Palma RL, Johnson KP, Clayton DH. The chewing lice: world checklist and biological overview Springfield: Illinois Natural History Survey Special Publication; 2003.
- Richardson DJ, Kinsella JM. New host and distribution records for gastrointestinal parasites of raptors from Connecticut, U.S.A. Comp Parasitol 2010; 77(1): 72-82. http://dx.doi.org/10.1654/4411.1.
» http://dx.doi.org/10.1654/4411.1 - Richardson DJ, Nickol BB. The Genus Centrorhynchus (Acanthocephala) in North America with Description of Centrorhynchus robustus n. sp., redescription of Centrorhynchus conspectus, and a Key to Species. J Parasitol 1995; 81(5): 767-772. http://dx.doi.org/10.2307/3283971. PMid:7472871.
» http://dx.doi.org/10.2307/3283971 - Rubilar L, Bertossi E, Skewes O. Gastrointestinal parasitism in the common partridge (Nothoprocta perdicaria) in the Nuble Zone, Chile. Bol Chil Parasitol 1996; 51(1-2): 35-37. PMid:9196955.
- San Martín J, Brevis C, Rubilar L, Krone O, González-Acuña D. Parasitismo gastrointestinal en tiuque común Milvago chimango chimango (VIEILLOT, 1816) (Falconidae, Aves) en Ñuble, Chile Parasitol Latinoam 2006; 61(1-2): 63-68. http://dx.doi.org/10.4067/S0717-77122006000100009.
» http://dx.doi.org/10.4067/S0717-77122006000100009 - Schock RC, Cooper R. Internal parasitisms in captive birds. Mod Vet Pract 1978; 59(6): 439-443. PMid:672879.
- Skrjabin K. Key to parasitic nematodes. Israel: Program for Scientific Translations; 1969.
- Sohn BO, Noh YT. Feather mites Kramerellidae and Pterolichidae in Korea (Acari, Pterolichoidea). Korean J Parasitol 1994; 32(2): 75-83. http://dx.doi.org/10.3347/kjp.1994.32.2.75. PMid:8025036.
» http://dx.doi.org/10.3347/kjp.1994.32.2.75 - Toro H, Saucedo C, Borie C, Gough RE, Alcaíno H. Health status of free-living pigeons in the city of Santiago. Avian Pathol 1999; 28(6): 619-623. http://dx.doi.org/10.1080/03079459994416.
» http://dx.doi.org/10.1080/03079459994416 - Van Cleave HJ. Acanthocephala of the genera Centrorhynchus and llediorhynchus (new genus) from North American birds. Tr Am Nlicr Soc 1916; 35(4): 221-232. http://dx.doi.org/10.2307/3221908.
» http://dx.doi.org/10.2307/3221908 - Van Cleave HJ. The Acanthocephala collected by the Allan Hancock Pacific Expedition, 1934. Rep Hancock Pacific Expedition (1932-1938) 1940; 2: 501-527.
- Woodyard ET, Rosser TG, Griffin MJ. New data on Neodiplostomum americanum Chandler and Rausch, 1947 (Digenea: Diplostomidae), in the Great Horned Owl Bubo virginianus Gmelin, 1788 and the Eastern Screech Owl Megascops asio Linnaeus, 1758 in Mississippi, USA. Parasitol Res 2017; 116(8): 2075-2089. http://dx.doi.org/10.1007/s00436-017-5503-1. PMid:28634620.
» http://dx.doi.org/10.1007/s00436-017-5503-1 - Yamaguti S. Systema Helminthum. vol. III. New York: Interscience Publisher; 1961.
- Yamaguti S. Systema Helminthum. vol. V. New York: Interscience Publisher; 1963.
Publication Dates
-
Publication in this collection
14 June 2018 -
Date of issue
Apr-Jun 2018
History
-
Received
23 Oct 2017 -
Accepted
07 Feb 2018