
Abstract
Endocrine-disrupting compounds (EDCs) and foodborne contaminants are environmental pollutants that are considered reproductive toxicants due to their deleterious effects on female and male gametes. Among the EDCs, the phthalate plasticizers are of growing concern. In-vivo and in-vitro models indicate that the oocyte is highly sensitive to phthalates. This review summarizes the effects of di(2-ethylhexyl) phthalate and its major metabolite mono(2-ethyhexyl) phthalate (MEHP) on the oocyte. MEHP reduces the proportion of oocytes that fertilize, cleave and develop to the blastocyst stage. This is associated with negative effects on meiotic progression, and disruption of cortical granules, endoplasmic reticulum and mitochondrial reorganization. MEHP alters mitochondrial membrane polarity, increases reactive oxygen species levels and induces alterations in genes associated with oxidative phosphorylation. A carryover effect from the oocyte to the blastocyst is manifested by alterations in the transcriptomic profile of blastocysts developed from MEHP-treated oocytes. Among foodborne contaminants, the pesticide atrazine (ATZ) and the mycotoxin aflatoxin B1 (AFB1) are of high concern. The potential hazards associated with exposure of spermatozoa to these contaminants and their carryover effect to the blastocyst are described. AFB1 and ATZ reduce spermatozoa's viability, as reflected by a high proportion of cells with damaged plasma membrane; induce acrosome reaction, expressed as damage to the acrosomal membrane; and interfere with mitochondrial function, characterized by hyperpolarization of the membrane. ATZ and AFB1-treated spermatozoa show a high proportion of cells with fragmented DNA. Exposure of spermatozoa to AFB1 and ATZ reduces fertilization and cleavage rates, but not that of blastocyst formation. However, fertilization with AFB1- or ATZ-treated spermatozoa impairs transcript expression in the formed blastocysts, implying a carryover effect. Taken together, the review indicates the risk of exposing farm animals to environmental contaminants, and their deleterious effects on female and male gametes and the developing embryo.
Keywords:
environmental contaminant; bovine; oocyte; spermatozoa; embryonic development
Introduction
Female and male reproductive health can be affected by various environmental stressors. While much attention has been paid to the increase in ambient temperature, i.e., global warming, less attention has been given to environmental contamination and its consequences on animal fertility. Reproductive health can be affected by a variety of manmade environmental contaminants (Diamanti-Kandarakis et al., 2009Diamanti-Kandarakis E, Bourguignon J-P, Giudice LC, Hauser R, Prins GS, Soto AM, Zoeller RT, Gore AC. Endocrine-disrupting chemicals: an Endocrine Society scientific statement. Endocr Rev. 2009;30(4):293-342. http://dx.doi.org/10.1210/er.2009-0002. PMid:19502515.
http://dx.doi.org/10.1210/er.2009-0002...
) or foodborne toxins (Jin et al., 2015Jin Y, Lin X, Miao W, Wang L, Wu Y, Fu Z. Oral exposure of pubertal male mice to endocrine-disrupting chemicals alters fat metabolism in adult livers. Environ Toxicol. 2015;30(12):1434-44. http://dx.doi.org/10.1002/tox.22013. PMid:24916741.
http://dx.doi.org/10.1002/tox.22013...
; Pflieger-Bruss et al., 2004Pflieger-Bruss S, Schuppe H-C, Schill W-B. The male reproductive system and its susceptibility to endocrine disrupting chemicals. Andrologia. 2004;36(6):337-45. http://dx.doi.org/10.1111/j.1439-0272.2004.00641.x. PMid:15541049.
http://dx.doi.org/10.1111/j.1439-0272.20...
), some of which are defined as endocrine-disrupting compounds (EDCs).
EDCs are exogenous chemicals that can interfere with natural hormone activities, such as synthesis, secretion, transport, metabolism, and binding action, thereby impairing homeostasis, reproduction, and developmental processes. EDCs act by binding to nuclear receptors [e.g., aryl-hydrocarbon (dioxin) receptor, peroxisome proliferator-activated receptor family]. EDCs can also act via other mechanisms that are associated with endocrine and reproductive systems by binding to non-nuclear steroid hormone receptors (e.g., membrane estrogen and androgen receptors), non-steroid receptors (e.g., serotonin, dopamine and norepinephrine receptor) (Diamanti-Kandarakis et al., 2009Diamanti-Kandarakis E, Bourguignon J-P, Giudice LC, Hauser R, Prins GS, Soto AM, Zoeller RT, Gore AC. Endocrine-disrupting chemicals: an Endocrine Society scientific statement. Endocr Rev. 2009;30(4):293-342. http://dx.doi.org/10.1210/er.2009-0002. PMid:19502515.
http://dx.doi.org/10.1210/er.2009-0002...
).
Throughout the female’s lifetime, follicular growth is characterized by recruitment of a cohort of primordial follicles into the pool of growing follicles from which a preovulatory follicle develops. In bovine, this course of follicular development is lengthy, taking approximately 180 days (Lussier et al., 1987Lussier JG, Matton P, Dufour JJ. Growth rates of follicles in the ovary of the cow. J Reprod Fertil. 1987;81(2):301-7. http://dx.doi.org/10.1530/jrf.0.0810301.
http://dx.doi.org/10.1530/jrf.0.0810301...
). Therefore, exposure to environmental contaminants during early stages of follicular development might further affect the function of the follicle and its enclosed oocytes. EDCs act on the hypothalamic–pituitary–ovarian axis, and their effect on the female reproductive tract is therefore multifactorial in nature. The deleterious effect is expressed as ovarian failure, impaired follicular development, accelerated atresia and depletion of antral growing follicles, ovulation failure and impaired steroidogenesis. These, in turn, might lead to reduced fertility (Bhattacharya and Keating, 2012Bhattacharya P, Keating AF. Impact of environmental exposures on ovarian function and role of xenobiotic metabolism during ovotoxicity. Toxicol Appl Pharmacol. 2012;261(3):227-35. http://dx.doi.org/10.1016/j.taap.2012.04.009. PMid:22531813.
http://dx.doi.org/10.1016/j.taap.2012.04...
; Craig et al., 2011Craig ZR, Wang W, Flaws JA. Endocrine-disrupting chemicals in ovarian function: effects on steroidogenesis, metabolism and nuclear receptor signaling. Reproduction. 2011;142(5):633-46. http://dx.doi.org/10.1530/REP-11-0136. PMid:21862696.
http://dx.doi.org/10.1530/REP-11-0136...
; Zama et al., 2016Zama AM, Bhurke A, Uzumcu M. Effects of Endocrine-disrupting Chemicals on Female Reproductive Health. Open Biotechnol J. 2016;10(1):54-75. http://dx.doi.org/10.2174/1874070701610010054.
http://dx.doi.org/10.2174/18740707016100...
).
Similarly, spermatogenesis is a highly intricate and coordinated process that produces thousands of spermatozoa daily in the seminiferous tubules of the testes, which then pass through the epididymal caput (head) and corpus (body) and are stored in the cauda (tail) until ejaculation (Reyes-Moreno et al., 2002Reyes-Moreno C, Boilard M, Sullivan R, Sirard MA. Characterization and Identification of Epididymal Factors That Protect Ejaculated Bovine Sperm During In Vitro Storage. Biol Reprod. 2002;66(1):159-66. http://dx.doi.org/10.1095/biolreprod66.1.159. PMid:11751277.
http://dx.doi.org/10.1095/biolreprod66.1...
; Cornwall, 2014Cornwall GA. Role of posttranslational protein modifications in epididymal sperm maturation and extracellular quality control. Adv Exp Med Biol. 2014;759:159-80. http://dx.doi.org/10.1007/978-1-4939-0817-2_8. PMid:25030764.
http://dx.doi.org/10.1007/978-1-4939-081...
). In the bull, this process takes about 60 days (Staub and Johnson, 2018Staub C, Johnson L. Review: spermatogenesis in the bull. Animal. 2018;12(s1):s27-35. http://dx.doi.org/10.1017/S1751731118000435. PMid:29882505.
http://dx.doi.org/10.1017/S1751731118000...
). During their passage through the epididymis compartments, the spermatozoa undergo extensive changes, which are completed in the female reproductive tract during the capacitation process. Given that spermatogenesis is also a lengthy process, spermatozoa are potentially exposed to a variety of environmental stressors at their different developmental stages. Indeed, epidemiological studies suggest an association between exposure to pesticides and semen quality (Swan et al., 2003Swan SH, Kruse RL, Liu F, Barr DB, Drobnis EZ, Redmon JB, Wang C, Brazil C, Overstreet JW. Semen Quality in Relation to Biomarkers of Pesticide Exposure. Environ Health Perspect. 2003;111(12):1478-84. http://dx.doi.org/10.1289/ehp.6417. PMid:12948887.
http://dx.doi.org/10.1289/ehp.6417...
). In that respect, the global decline in male fertility is mainly attributed to the exposure of spermatozoa to environmental EDCs, pharmaceutical products, pesticides and mycotoxins of food origin (Eze et al., 2018Eze UA, Huntriss J, Routledge MN, Gong YY. Toxicological effects of regulated mycotoxins and persistent organochloride pesticides: in vitro cytotoxic assessment of single and defined mixtures on MA-10 murine Leydig cell line. Toxicol Vitr. 2018;48:93-103. http://dx.doi.org/10.1016/j.tiv.2017.12.019. PMid:29307701.
http://dx.doi.org/10.1016/j.tiv.2017.12....
; Mann et al., 2020Mann U, Shiff B, Patel P. Reasons for worldwide decline in male fertility. Curr Opin Urol. 2020;1(3):296-301. http://dx.doi.org/10.1097/MOU.0000000000000745. PMid:32168194.
http://dx.doi.org/10.1097/MOU.0000000000...
; Tavares et al., 2016Tavares RS, Escada-Rebelo S, Correia M, Mota PC, Ramalho-Santos J. The non-genomic effects of endocrine-disrupting chemicals on mammalian sperm. Reproduction. 2016;151(1):R1-13. http://dx.doi.org/10.1530/REP-15-0355. PMid:26585413.
http://dx.doi.org/10.1530/REP-15-0355...
).
The aim of the current review is to describe the risk associated with exposing farm animals to environmental contaminants. The review focuses on the effects of representative EDCs on female and male gametes, in particular the effects of the plasticizers phthalates on the oocyte, and of foodborne contaminants on the spermatozoa. A carryover effect from the gametes to the developed embryo is also discussed.
Endocrine-disrupting compounds
The list of EDCs consists of more than 80,000 heterogeneous chemicals that are used for numerous applications. Some of these are the polychlorinated biphenyls, polybrominated biphenyls, dioxins, plastics (bisphenol A;BPA), pesticides [methoxychlor, chlorpyrifos, atrazine (ATZ), dichlorodiphenyltrichloroethane (DDT)], fungicides (vinclozolin), pharmaceutical agents (diethylstilbestrol) and plasticizers (phthalates) (Diamanti-Kandarakis et al., 2009Diamanti-Kandarakis E, Bourguignon J-P, Giudice LC, Hauser R, Prins GS, Soto AM, Zoeller RT, Gore AC. Endocrine-disrupting chemicals: an Endocrine Society scientific statement. Endocr Rev. 2009;30(4):293-342. http://dx.doi.org/10.1210/er.2009-0002. PMid:19502515.
http://dx.doi.org/10.1210/er.2009-0002...
). The current section focuses on phthalates, a group of chemical reproductive toxicants, and discusses the mechanism by which phthalates impair oocyte competence and further, the formed embryo.
Phthalates
Phthalate esters have been widely used in manufacturing for about a century (Koch and Calafat, 2009Koch HM, Calafat AM. Human body burdens of chemicals used in plastic manufacture. Philos Trans R Soc Lond B Biol Sci. 2009;364(1526):2063-78. http://dx.doi.org/10.1098/rstb.2008.0208. PMid:19528056.
http://dx.doi.org/10.1098/rstb.2008.0208...
). The annual production of phthalate is estimated at over 6 million tons worldwide, making these chemicals ubiquitous (Net et al., 2015aNet S, Delmont A, Sempéré R, Paluselli A, Ouddane B. Reliable quantification of phthalates in environmental matrices (air, water, sludge, sediment and soil): A review. Sci Total Environ. 2015a;515-516:162-80. http://dx.doi.org/10.1016/j.scitotenv.2015.02.013. PMid:25723871.
http://dx.doi.org/10.1016/j.scitotenv.20...
, bNet S, Sempéré R, Delmont A, Paluselli A, Ouddane B. Occurrence, fate, behavior and ecotoxicological state of phthalates in different environmental matrices. Environ Sci Technol. 2015b;49(7):4019-35. http://dx.doi.org/10.1021/es505233b. PMid:25730609.
http://dx.doi.org/10.1021/es505233b...
). More than 25 types of phthalates are used for commercial applications such as toys, vinyl flooring, wall covering, detergents, lubricating oils, food packaging, pharmaceuticals, blood bags and personal care products (Koch and Calafat, 2009Koch HM, Calafat AM. Human body burdens of chemicals used in plastic manufacture. Philos Trans R Soc Lond B Biol Sci. 2009;364(1526):2063-78. http://dx.doi.org/10.1098/rstb.2008.0208. PMid:19528056.
http://dx.doi.org/10.1098/rstb.2008.0208...
). The top 10 are dimethyl phthalate, diethyl phthalate, dibutyl phthalate, diisobutyl phthalate, benzylbutyl phthalate, dicyclohexyl phthalate, di(2-ethylhexyl) phthalate (DEHP), di-n-octyl phthalate, di-isononyl phthalate, and di-isodecyl phthalate. With respect to animal exposure, in particular dairy cows, phthalates can be found in milking equipment such as milking pipes, liners, teat dip cups, food packing and silage wrap (Pure Strategies, 2018Pure Strategies. Sources of phthalates in dairy farm equipment. Prepared for environmental health strategy center and the coalition for safer food processing & packaging [Internet] Pure Strategies; 2018 [cited 2020 June 23]. Available from: http://kleanupkraft.org/Phthalates-Farm-Equipment.pdf.
http://kleanupkraft.org/Phthalates-Farm-...
).
Among phthalates, DEHP is predominant, providing softness and flexibility to polyvinyl chloride products (ATSDR, 2002ATSDR. Toxicological profile for Di(2-ethylhexyl)phthalate (DEHP). Atlanta, GA: U.S. Department of Health and Human Services, Public Health Service; 2002.). DEHP does not bind covalently to the polyvinyl chloride polymer; it therefore leaches into the environment and is frequently found in the atmosphere, soil, sediment, and water sources (Bergé et al., 2013Bergé A, Cladière M, Gasperi J, Coursimault A, Tassin B, Moilleron R. Meta-analysis of environmental contamination by phthalates. Environ Sci Pollut Res Int. 2013;20(11):8057-76. http://dx.doi.org/10.1007/s11356-013-1982-5. PMid:23917738.
http://dx.doi.org/10.1007/s11356-013-198...
; Hongjun et al., 2013Hongjun Y, Wenjun X, Qing L, Jingtao L, Hongwen Y, Zhaohua L. Distribution of phthalate esters in topsoil: a case study in the Yellow River Delta, China. Environ Monit Assess. 2013;185(10):8489-500. http://dx.doi.org/10.1007/s10661-013-3190-7. PMid:23609921.
http://dx.doi.org/10.1007/s10661-013-319...
; Martine et al., 2013Martine B, Marie-Jeanne T, Cendrine D, Fabrice A, Marc C. Assessment of Adult Human Exposure to Phthalate Esters in the Urban Centre of Paris (France). Bull Environ Contam Toxicol. 2013;90(1):91-6. http://dx.doi.org/10.1007/s00128-012-0859-5. PMid:23090363.
http://dx.doi.org/10.1007/s00128-012-085...
; Net et al., 2015bNet S, Sempéré R, Delmont A, Paluselli A, Ouddane B. Occurrence, fate, behavior and ecotoxicological state of phthalates in different environmental matrices. Environ Sci Technol. 2015b;49(7):4019-35. http://dx.doi.org/10.1021/es505233b. PMid:25730609.
http://dx.doi.org/10.1021/es505233b...
). It is estimated that humans are exposed to DEHP at around 1.7–52.1 µg/kg body weight (BW) per day (McKee et al., 2004McKee RH, Butala JH, David RM, Gans G. NTP center for the evaluation of risks to human reproduction reports on phthalates: addressing the data gaps. Reprod Toxicol. 2004;18(1):1-22. http://dx.doi.org/10.1016/j.reprotox.2003.09.002. PMid:15013060.
http://dx.doi.org/10.1016/j.reprotox.200...
; Petersen and Breindahl, 2000Petersen JH, Breindahl T. Plasticizers in total diet samples, baby food and infant formulae. Food Addit Contam. 2000;17(2):133-41. http://dx.doi.org/10.1080/026520300283487. PMid:10793844.
http://dx.doi.org/10.1080/02652030028348...
). Infants are suggested to be at higher risk under certain circumstances (Kavlock et al., 2006Kavlock R, Barr D, Boekelheide K, Breslin W, Breysse P, Chapin R, Gaido K, Hodgson E, Marcus M, Shea K, Williams P. NTP-CERHR Expert Panel Update on the Reproductive and Developmental Toxicity of di(2-ethylhexyl) phthalate. Reprod Toxicol. 2006;22(3):291-399. http://dx.doi.org/10.1016/j.reprotox.2006.04.007. PMid:17068859.
http://dx.doi.org/10.1016/j.reprotox.200...
). In the body, DEHP is metabolized into several metabolites, including mono(2-ethyl-5-hydroxyhexyl) phthalate (5OH-MEHP), mono(2-ethyl-5-carboxypentyl) phthalate (5cx-MEPP), mono(2-ethyl-5-oxohexyl) phthalate (5oxo-MEHP), mono[2-(carboxymethyl)hexyl] phthalate (2cx-MMHP), and mono(2-ethyhexyl) phthalate (MEHP) (Koch et al., 2005Koch HM, Bolt HM, Preuss R, Angerer J. New metabolites of di(2-ethylhexyl)phthalate (DEHP) in human urine and serum after single oral doses of deuterium-labelled DEHP. Arch Toxicol. 2005;79(7):367-76. http://dx.doi.org/10.1007/s00204-004-0642-4. PMid:15700144.
http://dx.doi.org/10.1007/s00204-004-064...
). According to the US Food and Drug Administration, the metabolites have more adverse and toxic effects than DEHP.
While the effects of phthalates on human health have been extensively studied (Benjamin et al., 2017Benjamin S, Masai E, Kamimura N, Takahashi K, Anderson RC, Faisal PA. Phthalates impact human health: Epidemiological evidences and plausible mechanism of action. J Hazard Mater. 2017;340:360-83. http://dx.doi.org/10.1016/j.jhazmat.2017.06.036. PMid:28800814.
http://dx.doi.org/10.1016/j.jhazmat.2017...
; Hauser and Calafat, 2005Hauser R, Calafat AM. Phthalates and human health. Occup Environ Med. 2005;62(11):806-18. http://dx.doi.org/10.1136/oem.2004.017590. PMid:16234408.
http://dx.doi.org/10.1136/oem.2004.01759...
), the magnitude and risk of exposure of domestic animals to phthalates is less known, most likely due to less awareness or concern. Nevertheless, recent studies have reported a deleterious effect of phthalates on animal health. In vivo studies in farm animals demonstrated the clearance pattern of DEHP. Oral administration to young male pigs of 1000 mg/kg BW DEHP resulted in a rapid increase in MEHP level (14 ng/L) in the blood with a half-life of ~6 h (Ljungvall et al., 2004Ljungvall K, Tienpont B, David F, Magnusson U, Törneke K. Kinetics of orally administered di(2-ethylhexyl) phthalate and its metabolite, mono(2-ethylhexyl) phthalate, in male pigs. Arch Toxicol. 2004;78(7):384-9. http://dx.doi.org/10.1007/s00204-004-0558-z. PMid:15022035.
http://dx.doi.org/10.1007/s00204-004-055...
). Oral administration of DEHP to cows resulted in low levels of the metabolites 5OH-MEHP, MEHP, 5oxo-MEHP, 2cx-MMHP, and 5cx-MEPP were found in the milk, urine, and plasma samples of DEHP-treated cows, 20 days after DEHP administration (Kalo et al., 2015Kalo D, Hadas R, Furman O, Ben-Ari J, Maor Y, Patterson DG, Tomey C, Roth Z. Carryover effects of acute DEHP exposure on ovarian function and oocyte developmental competence in lactating cows. PLoS One. 2015;10(7):e0130896. http://dx.doi.org/10.1371/journal.pone.0130896. PMid:26154164.
http://dx.doi.org/10.1371/journal.pone.0...
). A level of ~20 nM MEHP was detected in the follicular fluid aspirated from the DEHP-treated cows (Kalo et al., 2015Kalo D, Hadas R, Furman O, Ben-Ari J, Maor Y, Patterson DG, Tomey C, Roth Z. Carryover effects of acute DEHP exposure on ovarian function and oocyte developmental competence in lactating cows. PLoS One. 2015;10(7):e0130896. http://dx.doi.org/10.1371/journal.pone.0130896. PMid:26154164.
http://dx.doi.org/10.1371/journal.pone.0...
). Similarly, a level of ~10 nM DEHP was recorded in equine follicular fluid (Marzano et al., 2019Marzano G, Mastrorocco A, Zianni R, Mangiacotti M, Chiaravalle AE, Lacalandra GM, Minervini F, Cardinali A, Macciocca M, Vicenti R, Fabbri R, Hinrichs K, Dell’Aquila ME, Martino NA. Altered morphokinetics in equine embryos from oocytes exposed to DEHP during IVM. Mol Reprod Dev. 2019;86(10):1388-404. http://dx.doi.org/10.1002/mrd.23156. PMid:31025442.
http://dx.doi.org/10.1002/mrd.23156...
). Non-DEHP-originated metabolites such as monoethyl phthalate and monobutyl phthalate were also detected in the follicular fluid, presumably of environmental origin (Kalo et al., 2015Kalo D, Hadas R, Furman O, Ben-Ari J, Maor Y, Patterson DG, Tomey C, Roth Z. Carryover effects of acute DEHP exposure on ovarian function and oocyte developmental competence in lactating cows. PLoS One. 2015;10(7):e0130896. http://dx.doi.org/10.1371/journal.pone.0130896. PMid:26154164.
http://dx.doi.org/10.1371/journal.pone.0...
). In addition, DEHP has been reported in bovine milk (Jarošová, 2006Jarošová A. Phthalic acid esters (PAEs) in the Food Chain. Czech J Food Sci. 2006;24(5):223-31. http://dx.doi.org/10.17221/3318-CJFS.
http://dx.doi.org/10.17221/3318-CJFS...
; Krejčíková and Jarošová, 2013Krejčíková M, Jarošová A. Phthalate in cow milk depending on the method of milking. Medelnet [serial on the Internet]. 2013:592-6. [cited 2020 June 23]. Available from: http://mnet.mendelu.cz/mendelnet2013/articles/44_krejcikova_795.pdf
http://mnet.mendelu.cz/mendelnet2013/art...
) and fat tissue (Jarošová, 2006Jarošová A. Phthalic acid esters (PAEs) in the Food Chain. Czech J Food Sci. 2006;24(5):223-31. http://dx.doi.org/10.17221/3318-CJFS.
http://dx.doi.org/10.17221/3318-CJFS...
) and in ewe (Rhind et al., 2005Rhind SM, Kyle CE, Telfer G, Duff EI, Smith A. Alkyl phenols and diethylhexyl phthalate in tissues of sheep grazing pastures fertilized with sewage sludge or inorganic fertilizer. Environ Health Perspect. 2005;113(4):447-53. http://dx.doi.org/10.1289/ehp.7469. PMid:15811823.
http://dx.doi.org/10.1289/ehp.7469...
) and swine tissues (Jarošová, 2006Jarošová A. Phthalic acid esters (PAEs) in the Food Chain. Czech J Food Sci. 2006;24(5):223-31. http://dx.doi.org/10.17221/3318-CJFS.
http://dx.doi.org/10.17221/3318-CJFS...
; Ljungvall et al., 2004Ljungvall K, Tienpont B, David F, Magnusson U, Törneke K. Kinetics of orally administered di(2-ethylhexyl) phthalate and its metabolite, mono(2-ethylhexyl) phthalate, in male pigs. Arch Toxicol. 2004;78(7):384-9. http://dx.doi.org/10.1007/s00204-004-0558-z. PMid:15022035.
http://dx.doi.org/10.1007/s00204-004-055...
).
Effect of phthalates on female reproduction
There is accumulating evidence associating the presence of phthalates in the environment with impaired reproduction and reduced female fertility. In women undergoing in-vitro fertilization treatments, the presence of DEHP and its metabolites in the urine has been associated with a low number of retrieved, matured and fertilized oocytes and a reduced number of high-quality embryos (Machtinger et al., 2018Machtinger R, Gaskins AJ, Racowsky C, Mansur A, Adir M, Baccarelli AA, Calafat AM, Hauser R. Urinary concentrations of biomarkers of phthalates and phthalate alternatives and IVF outcomes. Environ Int. 2018;111:23-31. http://dx.doi.org/10.1016/j.envint.2017.11.011. PMid:29161633.
http://dx.doi.org/10.1016/j.envint.2017....
). Exposure of women to MEHP close to conception was associated with early pregnancy loss (Toft et al., 2011Toft G, Jönsson BAG, Lindh CH, Jensen TK, Hjollund NH, Vested A, Bonde JP. Association between Pregnancy Loss and Urinary Phthalate Levels around the Time of Conception. Environ Health Perspect. 2011;120(3):458-63. http://dx.doi.org/10.1289/ehp.1103552. PMid:22113848.
http://dx.doi.org/10.1289/ehp.1103552...
). High levels of MEHP and 5oxo-MEHP in the urine have been associated with long gestation and interruption in parturition (Adibi et al., 2009Adibi JJ, Hauser R, Williams PL, Whyatt RM, Calafat AM, Nelson H, Herrick R, Swan SH. Maternal urinary metabolites of Di-(2-Ethylhexyl) phthalate in relation to the timing of labor in a US multicenter pregnancy cohort study. Am J Epidemiol. 2009;169(8):1015-24. http://dx.doi.org/10.1093/aje/kwp001. PMid:19251754.
http://dx.doi.org/10.1093/aje/kwp001...
). Tranfo et al. (2012)Tranfo G, Caporossi L, Paci E, Aragona C, Romanzi D, De Carolis C, De Rosa M, Capanna S, Papaleo B, Pera A. Urinary phthalate monoesters concentration in couples with infertility problems. Toxicol Lett. 2012;213(1):15-20. http://dx.doi.org/10.1016/j.toxlet.2011.11.033. PMid:22197707.
http://dx.doi.org/10.1016/j.toxlet.2011....
found a correlation between the level of DEHP metabolites in the urine and infertility, in both men and women. In mice, DEHP and MEHP deleteriously affected the ovarian pool of follicles, expressed as a reduction in the number of primordial and antral follicles (Hannon et al., 2016Hannon PR, Niermann S, Flaws JA. Acute exposure to Di(2-Ethylhexyl) phthalate in adulthood causes adverse reproductive outcomes later in life and accelerates reproductive aging in female mice. Toxicol Sci. 2016;150(1):97-108. http://dx.doi.org/10.1093/toxsci/kfv317. PMid:26678702.
http://dx.doi.org/10.1093/toxsci/kfv317...
; Li et al., 2016Li L, Liu J-C, Lai F-N, Liu H-Q, Zhang X-F, Dyce PW, Shen W, Chen H. Di (2-ethylhexyl) Phthalate Exposure Impairs Growth of Antral Follicle in Mice. PLoS One. 2016;11(2):e0148350. http://dx.doi.org/10.1371/journal.pone.0148350. PMid:26845775.
http://dx.doi.org/10.1371/journal.pone.0...
; Moyer and Hixon, 2012Moyer B, Hixon ML. Reproductive effects in F1 adult females exposed in utero to moderate to high doses of mono-2-ethylhexylphthalate (MEHP). Reprod Toxicol. 2012;34(1):43-50. http://dx.doi.org/10.1016/j.reprotox.2012.02.006. PMid:22401849.
http://dx.doi.org/10.1016/j.reprotox.201...
; Niermann et al., 2015Niermann S, Rattan S, Brehm E, Flaws JA. Prenatal exposure to di-(2-ethylhexyl) phthalate (DEHP) affects reproductive outcomes in female mice. Reprod Toxicol. 2015;53:23-32. http://dx.doi.org/10.1016/j.reprotox.2015.02.013. PMid:25765777.
http://dx.doi.org/10.1016/j.reprotox.201...
; Zhang et al., 2013Zhang X-F, Zhang L-J, Li L, Feng Y-N, Chen B, Ma J-M, Huynh E, Shi QH, De Felici M, Shen W. Diethylhexyl phthalate exposure impairs follicular development and affects oocyte maturation in the mouse. Environ Mol Mutagen. 2013;54(5):354-61. http://dx.doi.org/10.1002/em.21776. PMid:23625783.
http://dx.doi.org/10.1002/em.21776...
). Chronic exposure of female mice, at implantation, to a high level of phthalate (250 mg/kg BW) was associated with decreased pregnancy rates and increased miscarriage rates (Aldyreva et al., 1975Aldyreva MV, Klimova TS, Iziumova AS, Timofeevskaia LA. The effect of phthalate plasticizers on the generative function. Gig Tr Prof Zabol. 1975;(12):25-9. PMid:1213487.). Exposure of female rats to high doses of DEHP (up to 2 g/kg BW) altered estradiol concentration, inhibited follicular growth, disrupted the estrous cycle, and impaired ovulation (Davis et al., 1994Davis BJ, Maronpot RR, Di Heindel JJ. -(2-ethylhexyl) phthalate suppresses estradiol and ovulation in cycling rats. Toxicol Appl Pharmacol. 1994;128(2):216-23. http://dx.doi.org/10.1006/taap.1994.1200. PMid:7940536.
http://dx.doi.org/10.1006/taap.1994.1200...
; Gupta et al., 2010Gupta RK, Singh JM, Leslie TC, Meachum S, Flaws JA, Yao HH. C-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl) phthalate inhibit growth and reduce estradiol levels of antral follicles in vitro. Toxicol Appl Pharmacol. 2010;242(2):224-30. http://dx.doi.org/10.1016/j.taap.2009.10.011. PMid:19874833.
http://dx.doi.org/10.1016/j.taap.2009.10...
; Lovekamp-Swan and Davis, 2003Lovekamp-Swan T, Davis BJ. Mechanisms of phthalate ester toxicity in the female reproductive system. Environ Health Perspect. 2003;111(2):139-45. http://dx.doi.org/10.1289/ehp.5658. PMid:12573895.
http://dx.doi.org/10.1289/ehp.5658...
). With respect to DEHP metabolites, studies in mice have reported that MEHP inhibits follicular growth and reduces estradiol production in antral follicles (Gupta et al., 2010Gupta RK, Singh JM, Leslie TC, Meachum S, Flaws JA, Yao HH. C-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl) phthalate inhibit growth and reduce estradiol levels of antral follicles in vitro. Toxicol Appl Pharmacol. 2010;242(2):224-30. http://dx.doi.org/10.1016/j.taap.2009.10.011. PMid:19874833.
http://dx.doi.org/10.1016/j.taap.2009.10...
; Wang et al., 2012Wang W, Craig ZR, Basavarajappa MS, Hafner KS, Flaws JA. Mono-(2-ethylhexyl) phthalate induces oxidative stress and inhibits growth of mouse ovarian antral follicles. Biol Reprod. 2012;87(6):152. http://dx.doi.org/10.1095/biolreprod.112.102467. PMid:23077170.
http://dx.doi.org/10.1095/biolreprod.112...
). In vivo exposure of cows to 100 mg/kg DEHP per day for 3 days impairs the pattern of follicular development, decreases estradiol concentration and increase follicular pathologies (Kalo et al., 2015Kalo D, Hadas R, Furman O, Ben-Ari J, Maor Y, Patterson DG, Tomey C, Roth Z. Carryover effects of acute DEHP exposure on ovarian function and oocyte developmental competence in lactating cows. PLoS One. 2015;10(7):e0130896. http://dx.doi.org/10.1371/journal.pone.0130896. PMid:26154164.
http://dx.doi.org/10.1371/journal.pone.0...
).
Taken together, it is becoming clear that both DEHP and MEHP are reproductive toxicants (Balabanič et al., 2011Balabanič D, Rupnik M, Klemenčič AK. Negative impact of endocrine-disrupting compounds on human reproductive health. Reprod Fertil Dev. 2011;23(3):403-16. http://dx.doi.org/10.1071/RD09300. PMid:21426858.
http://dx.doi.org/10.1071/RD09300...
; Heudorf et al., 2007Heudorf U, Mersch-Sundermann V, Angerer J. Phthalates: toxicology and exposure. Int J Hyg Environ Health. 2007;210(5):623-34. http://dx.doi.org/10.1016/j.ijheh.2007.07.011. PMid:17889607.
http://dx.doi.org/10.1016/j.ijheh.2007.0...
; Johnson et al., 2012Johnson KJ, Heger NE, Boekelheide K. Of mice and men (and rats): phthalate-induced fetal testis endocrine disruption is species-dependent. Toxicol Sci. 2012;129(2):235-48. http://dx.doi.org/10.1093/toxsci/kfs206. PMid:22700540.
http://dx.doi.org/10.1093/toxsci/kfs206...
; Kay et al., 2013Kay VR, Chambers C, Foster WG. Reproductive and developmental effects of phthalate diesters in females. Crit Rev Toxicol. 2013;43(3):200-19. http://dx.doi.org/10.3109/10408444.2013.766149. PMid:23405971.
http://dx.doi.org/10.3109/10408444.2013....
).
Effects of DEHP and MEHP on the oocyte
Gathering evidence indicates that the oocyte is highly sensitive to phthalates. Oral administration of mice with 50–200 µL of 2.56 µM DEHP for 12 days reduced the number of mature oocytes, and decreased fertilization rate and embryonic development (Eimani et al., 2005Eimani H, Dalman A, Sepehri H, Kazemi S, Hassani F, Rezazadeh M, Baharvand H, Shahverdi A, Samani RO. Effect of DEHP (Di-2-Ethyl Hexyl-phthalate) on resumption of meiosis and in-vitro maturation of mouse oocytes and development of resulting embryos. Yakhth Med J. 2005;7:56-61.). Exposing the ovaries of newborn mice, in vitro, to DEHP (10 or 100 µM) induced apoptosis, as it increased the mRNA expression of Bax, a proapoptotic factor, and the proportion of TUNEL-positive oocytes (Zhang et al., 2014Zhang T, Li L, Qin X-S, Zhou Y, Zhang X-F, Wang L-Q, De Felici M, Chen H, Qin GQ, Shen W. Di-(2-ethylhexyl) phthalate and bisphenol A exposure impairs mouse primordial follicle assembly in vitro. Environ Mol Mutagen. 2014;55(4):343-53. http://dx.doi.org/10.1002/em.21847. PMid:24458533.
http://dx.doi.org/10.1002/em.21847...
). Oral administration of DEHP (50, 100 or 200 μL of 2.56 μM) to mice dose-dependently reduced the number of oocytes progressing to metaphase II. This was accompanied by a decrease in oocyte developmental competence, an increase in early apoptosis, and a decrease in the expression levels of the genes Ccna1, Asah1 and Pou5f1 (Absalan et al., 2017Absalan F, Saremy S, Mansori E, Taheri Moghadam M, Eftekhari Moghadam AR, Ghanavati R. Effects of Mono-(2-Ethylhexyl) Phthalate and Di-(2-Ethylhexyl) Phthalate administrations on Oocyte Meiotic Maturation, Apoptosis and Gene Quantification in mouse model. Cell J. 2017;18(4):503-13. PMid:28042535.). Oral administration of 2000 μg/kg BW DEHP per day to mice resulted in a lower number of retrieved oocytes, a higher number of unfertilized oocytes, a higher number of fragmented zygotes, and decreased embryonic development (Parra-Forero et al., 2019Parra-Forero LY, Veloz-Contreras A, Vargas-Marín S, Mojica-Villegas MA, Alfaro-Pedraza E, Urióstegui-Acosta M, Hernández-Ochoa I. Alterations in oocytes and early zygotes following oral exposure to di(2-ethylhexyl) phthalate in young adult female mice. Reprod Toxicol. 2019;90:53-61. http://dx.doi.org/10.1016/j.reprotox.2019.08.012. PMid:31442482.
http://dx.doi.org/10.1016/j.reprotox.201...
).
While much data have been reported for rodents, data from farm animals are limited. Acute exposure of equine oocytes to DEHP (12 and 1200 µM) induced apoptosis and increased reactive oxygen species (ROS) levels in the cumulus cells, but did not affect nuclear maturation (Ambruosi et al., 2011Ambruosi B, Uranio M, Sardanelli A, Pocar P, Martino N, Parternoster M, Amati F, Dell'Aquila ME. In Vitro acute exposure to DEHP affects Oocyte Meiotic maturation, energy and oxidative stress parameters in a large animal model. PLoS One. 2011;6(11):e27452. http://dx.doi.org/10.1371/journal.pone.0027452. PMid:22076161.
http://dx.doi.org/10.1371/journal.pone.0...
). In-vitro culture of equine oocytes with 0.12 µM DEHP inhibited their nuclear maturation, and increased apoptosis and ROS levels in the surrounding cumulus cells (Ambruosi et al., 2011Ambruosi B, Uranio M, Sardanelli A, Pocar P, Martino N, Parternoster M, Amati F, Dell'Aquila ME. In Vitro acute exposure to DEHP affects Oocyte Meiotic maturation, energy and oxidative stress parameters in a large animal model. PLoS One. 2011;6(11):e27452. http://dx.doi.org/10.1371/journal.pone.0027452. PMid:22076161.
http://dx.doi.org/10.1371/journal.pone.0...
). On the other hand, exposure of porcine cumulus oocyte complexes (COCs) to extremely high doses of DEHP (250 µM to 5 mM) did not affect the proportion of oocytes with extruded polar body (Wang et al., 2016Wang T, Han J, Duan X, Xiong B, Cui X-S, Kim N-H, Liu HL, Sun SC. The toxic effects and possible mechanisms of Bisphenol A on oocyte maturation of porcine in vitro. Oncotarget. 2016;7(22):32554-65. http://dx.doi.org/10.18632/oncotarget.8689. PMid:27086915.
http://dx.doi.org/10.18632/oncotarget.86...
). Exposure of bovine COCs to 50 µM DEHP during oocyte maturation did not affect their cleavage rate into 2- to 4-cell-stage embryos, but decreased blastocyst-formation rate (Grossman et al., 2012Grossman D, Kalo D, Gendelman M, Roth Z. Effect of di-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl) phthalate on in vitro developmental competence of bovine oocytes. Cell Biol Toxicol. 2012;28(6):383-96. http://dx.doi.org/10.1007/s10565-012-9230-1. PMid:22956148.
http://dx.doi.org/10.1007/s10565-012-923...
).
In-vitro exposure of bovine oocytes to MEHP (50 µM) impaired nuclear maturation, reflected by a decreased proportion of oocytes that resumed meiosis and progressed to the metaphase II stage (Grossman et al., 2012Grossman D, Kalo D, Gendelman M, Roth Z. Effect of di-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl) phthalate on in vitro developmental competence of bovine oocytes. Cell Biol Toxicol. 2012;28(6):383-96. http://dx.doi.org/10.1007/s10565-012-9230-1. PMid:22956148.
http://dx.doi.org/10.1007/s10565-012-923...
; Kalo and Roth, 2015Kalo D, Roth Z. Effects of mono(2-ethylhexyl)phthalate on cytoplasmic maturation of oocytes-the bovine model. Reprod Toxicol. 2015;53:141-51. http://dx.doi.org/10.1016/j.reprotox.2015.04.007. PMid:25900598.
http://dx.doi.org/10.1016/j.reprotox.201...
). In-vitro culturing of bovine oocytes in follicular fluid containing ~20 nM MEHP resulted in a low proportion of oocytes that resumed meiosis. In particular, a higher proportion of MEHP-treated oocytes were blocked in the telophase I stage (Figure 1A; Kalo et al., 2015Kalo D, Hadas R, Furman O, Ben-Ari J, Maor Y, Patterson DG, Tomey C, Roth Z. Carryover effects of acute DEHP exposure on ovarian function and oocyte developmental competence in lactating cows. PLoS One. 2015;10(7):e0130896. http://dx.doi.org/10.1371/journal.pone.0130896. PMid:26154164.
http://dx.doi.org/10.1371/journal.pone.0...
). In support of this, exposure of COCs or denuded bovine oocytes to 5–100 µM MEHP increased the proportion of oocytes expressing a germinal vesicle stage nucleus by the end of culture, i.e., they did not resume meiosis (Anas et al., 2003Anas M-KI, Suzuki C, Yoshioka K, Iwamura S. Effect of mono-(2-ethylhexyl) phthalate on bovine oocyte maturation in vitro. Reprod Toxicol. 2003;17(3):305-10. http://dx.doi.org/10.1016/S0890-6238(03)00014-5. PMid:12759099.
http://dx.doi.org/10.1016/S0890-6238(03)...
). Similar findings were reported for bovine COCs cultured with a high concentration of MEHP (75 µM) (Beker van Woudenberg et al., 2012Beker van Woudenberg A, Gröllers-Mulderij M, Snel C, Jeurissen N, Stierum R, Wolterbeek A. The bovine oocyte in vitro maturation model: A potential tool for reproductive toxicology screening. Reprod Toxicol. 2012;34(2):251-60. http://dx.doi.org/10.1016/j.reprotox.2012.05.098. PMid:22664270.
http://dx.doi.org/10.1016/j.reprotox.201...
) and for porcine COCs cultured with 50–100 μM MEHP (Zhang et al., 2018Zhang Y, Wang T, Lan M, Zang XW, Li YL, Cui XS, Kim NH, Sun SC. Melatonin protects oocytes from MEHP exposure-induced meiosis defects in porcine. Biol Reprod. 2018;98(3):286-98. http://doi.org//10.1093/biolre/iox185
http://doi.org//10.1093/biolre/iox185...
).
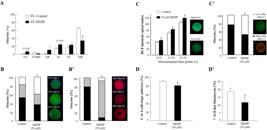
Effect of the EDC MEHP on bovine oocytes. (A) Nuclear maturation of oocytes cultured in follicular fluid (FF) aspirated from control (FF-control) and DEHP-treated (FF-DEHP) cows. Presented is the distribution of oocytes into meiotic stages: germinal vesicle (GV), GV breakdown (GVBD), metaphase I (MI), anaphase I (AI), telophase I (TI), and metaphase II (MII) (adapted with permission from Kalo et al., 2015Kalo D, Hadas R, Furman O, Ben-Ari J, Maor Y, Patterson DG, Tomey C, Roth Z. Carryover effects of acute DEHP exposure on ovarian function and oocyte developmental competence in lactating cows. PLoS One. 2015;10(7):e0130896. http://dx.doi.org/10.1371/journal.pone.0130896. PMid:26154164.
http://dx.doi.org/10.1371/journal.pone.0... ). (B,B') Cytoplasmic maturation is affected by MEHP. (B) Distribution of mature oocytes among three types of cortical granule (CG) patterns (fluorescein-conjugated peanut agglutinin [FITC–PNA] dye): CG-Type I (immature), CG-Type II (partially mature) and CG-Type III (fully mature). (B') Distribution of mature oocytes among three classes of endoplasmic reticulum (ER) pattern (red dye): ER-Class III (degenerative), ER-Class II (immature) and ER-Class I (fully mature). (C,C') Mitochondrial function is affected by MEHP. (C) ROS level expressed by dichlorofluorescein (DCF) fluorescence intensity, measured from 0–10, 11–20 and 21–30 s in mature oocytes, and representative images of live oocyte stained with dichlorodihydrofluorescein diacetate (H2DCFDA) expressing low (bottom) or high (top) DCF fluorescence intensity. (C') Proportion of oocytes with high- and low-polarized mitochondrial membrane (∆Ψm) along with representative images of live oocytes stained with MitoCapture mitochondrial apoptosis kit expressing high (red) and low (green) ∆Ψm (adapted with permission from Kalo and Roth, 2015Kalo D, Roth Z. Effects of mono(2-ethylhexyl)phthalate on cytoplasmic maturation of oocytes-the bovine model. Reprod Toxicol. 2015;53:141-51. http://dx.doi.org/10.1016/j.reprotox.2015.04.007. PMid:25900598.
http://dx.doi.org/10.1016/j.reprotox.201... ). (D,D') Developmental competence of oocytes in-vitro matured without or with 50 µM MEHP, further fertilized and cultured for 8 days. (D) Cleavage rate into 2- to 4-cell-stage embryos. (D') Blastocyst-formation rate (adopted with permission from Grossman et al., 2012Grossman D, Kalo D, Gendelman M, Roth Z. Effect of di-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl) phthalate on in vitro developmental competence of bovine oocytes. Cell Biol Toxicol. 2012;28(6):383-96. http://dx.doi.org/10.1007/s10565-012-9230-1. PMid:22956148.
http://dx.doi.org/10.1007/s10565-012-923... ).
One of the suggested mechanisms by which phthalates impair oocyte nuclear maturation is alterations in the cAMP-signaling pathway. An accurate cAMP level is required for chromatin transition, and synchronization between nuclear and cytoplasmic maturation (Gilchrist et al., 2011Gilchrist RB, Aktas H, Wheeler MB. Recent insights into oocyte - follicle cell interactions provide opportunities for the development of new approaches to in vitro maturation. Reprod Fertil Dev. 2011;23(1):23-31. http://dx.doi.org/10.1071/RD10225. PMid:21366977.
http://dx.doi.org/10.1071/RD10225...
; Luciano et al., 2011Luciano AM, Franciosi F, Modina SC, Lodde V. Gap junction-mediated communications regulate chromatin remodeling during bovine oocyte growth and differentiation through cAMP-Dependent Mechanism(s). Biol Reprod. 2011;85(6):1252-9. http://dx.doi.org/10.1095/biolreprod.111.092858. PMid:21816847.
http://dx.doi.org/10.1095/biolreprod.111...
). Microarray analysis of MEHP-treated bovine oocytes revealed alterations in seven transcripts associated with the cAMP-signaling pathway (Kalo et al., 2019Kalo D, Vitorino Carvalho A, Archilla C, Duranthon V, Moroldo M, Levin Y, Kupervaser M, Smith Y, Roth Z. Mono(2-ethylhexyl) phthalate (MEHP) induces transcriptomic alterations in oocytes and their derived blastocysts. Toxicology. 2019;421:59-73. http://dx.doi.org/10.1016/j.tox.2019.04.016. PMid:31059758.
http://dx.doi.org/10.1016/j.tox.2019.04....
). In rats, MEHP inhibited the accumulation of cAMP in Sertoli cells (Heindel and Chapin, 1989Heindel JJ, Chapin RE. Inhibition of FSH-stimulated cAMP accumulation by mono(2-ethylhexyl) phthalate in primary rat Sertoli cell cultures. Toxicol Appl Pharmacol. 1989;97(2):377-85. http://dx.doi.org/10.1016/0041-008X(89)90342-6. PMid:2538009.
http://dx.doi.org/10.1016/0041-008X(89)9...
). In addition, exposure of rat granulosa cells to 100 µM MEHP decreased progesterone production and follicle-stimulating hormone-induced cAMP accumulation (Treinen et al., 1990Treinen KA, Dodson WC, Heindel JJ. Inhibition of FSH-stimulated cAMP accumulation and progesterone production by mono(2-ethylhexyl) phthalate in rat granulosa cell cultures. Toxicol Appl Pharmacol. 1990;106(2):334-40. http://dx.doi.org/10.1016/0041-008X(90)90252-P. PMid:2175055.
http://dx.doi.org/10.1016/0041-008X(90)9...
). MEHP-induced impairment of the cell cycle has also been suggested as a potential mechanism, because exposure of bovine oocytes to 50 µM MEHP altered the expression level of CCNA2 (Grossman et al., 2012Grossman D, Kalo D, Gendelman M, Roth Z. Effect of di-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl) phthalate on in vitro developmental competence of bovine oocytes. Cell Biol Toxicol. 2012;28(6):383-96. http://dx.doi.org/10.1007/s10565-012-9230-1. PMid:22956148.
http://dx.doi.org/10.1007/s10565-012-923...
). In mice, oral administration of 100 µL DEHP or MEHP (2.56 µM) resulted in reduced expression of the pluripotency (Pou5f1), cell cycle (Ccna1) and apoptotic (Asah1) related genes (Absalan et al., 2017Absalan F, Saremy S, Mansori E, Taheri Moghadam M, Eftekhari Moghadam AR, Ghanavati R. Effects of Mono-(2-Ethylhexyl) Phthalate and Di-(2-Ethylhexyl) Phthalate administrations on Oocyte Meiotic Maturation, Apoptosis and Gene Quantification in mouse model. Cell J. 2017;18(4):503-13. PMid:28042535.).
Recent studies have provided evidence that phthalates induce alterations in cytoplasmic maturation in association with reduced oocyte competence. The maturation process includes multiple activities that are highly important to the formation of a mature and fertilizable oocyte. These include organelle redistribution (mitochondria, endoplasmic reticulum, cortical granules and lipid droplets), cytoskeletal rearrangement, and cytoplasmic polyadenylation (Eppig, 1996Eppig JJ. Coordination of nuclear and cytoplasmic oocyte maturation in eutherian mammals. Reprod Fertil Dev. 1996;8(4):485-9. http://dx.doi.org/10.1071/RD9960485. PMid:8870074.
http://dx.doi.org/10.1071/RD9960485...
; Ferreira et al., 2009Ferreira EM, Vireque AA, Adona PR, Meirelles FV, Ferriani RA, Navarro PAAS. Cytoplasmic maturation of bovine oocytes: structural and biochemical modifications and acquisition of developmental competence. Theriogenology. 2009;71(5):836-48. http://dx.doi.org/10.1016/j.theriogenology.2008.10.023. PMid:19121865.
http://dx.doi.org/10.1016/j.theriogenolo...
). Culturing of bovine COCs with 50 µM MEHP altered oocyte cytoplasmic maturation, reflected by alterations in the reorganization of the mitochondria, cortical granules and endoplasmic reticulum (Figure 1B,B'; Kalo and Roth, 2015Kalo D, Roth Z. Effects of mono(2-ethylhexyl)phthalate on cytoplasmic maturation of oocytes-the bovine model. Reprod Toxicol. 2015;53:141-51. http://dx.doi.org/10.1016/j.reprotox.2015.04.007. PMid:25900598.
http://dx.doi.org/10.1016/j.reprotox.201...
). Given the pivotal function of the mitochondria in early stages of embryonic development (Acton et al., 2004Acton BM, Jurisicova A, Jurisica I, Casper RF. Alterations in mitochondrial membrane potential during preimplantation stages of mouse and human embryo development. Mol Hum Reprod. 2004;10(1):23-32. http://dx.doi.org/10.1093/molehr/gah004. PMid:14665703.
http://dx.doi.org/10.1093/molehr/gah004...
; Bavister and Squirrell, 2000Bavister BD, Squirrell JM. Mitochondrial distribution and function in oocytes and early embryos. Hum Reprod. 2000;15(Suppl 2):189-98. http://dx.doi.org/10.1093/humrep/15.suppl_2.189. PMid:11041524.
http://dx.doi.org/10.1093/humrep/15.supp...
; Thouas et al., 2004Thouas GA, Trounson AO, Wolvetang EJ, Jones GM. Mitochondrial Dysfunction in Mouse Oocytes Results in Preimplantation Embryo Arrest in Vitro. Biol Reprod. 2004;71(6):1936-42. http://dx.doi.org/10.1095/biolreprod.104.033589. PMid:15286028.
http://dx.doi.org/10.1095/biolreprod.104...
), MEHP-induced impairment in oocyte mitochondrial function is suggested to be one of the main mechanisms underlying reduced developmental competence (Roth, 2018Roth Z. Symposium review: Reduction in oocyte developmental competence by stress is associated with alterations in mitochondrial function1. J Dairy Sci. 2018;101(4):3642-54. http://dx.doi.org/10.3168/jds.2017-13389. PMid:29395145.
http://dx.doi.org/10.3168/jds.2017-13389...
). Alterations in mitochondrial distribution were associated with increased ROS levels and decreased mitochondrial membrane potential (ΔΨm) (Figure 1C,C': Kalo and Roth, 2015Kalo D, Roth Z. Effects of mono(2-ethylhexyl)phthalate on cytoplasmic maturation of oocytes-the bovine model. Reprod Toxicol. 2015;53:141-51. http://dx.doi.org/10.1016/j.reprotox.2015.04.007. PMid:25900598.
http://dx.doi.org/10.1016/j.reprotox.201...
). In mice, oral treatment with DEHP (40 μg/kg BW) increased ROS levels in the oocytes, along with disruption of mitochondrial function and reduction in ATP levels (Lu et al., 2019Lu Z, Zhang C, Han C, An Q, Cheng Y, Chen Y, Chen Y, Meng R, Zhang Y, Su J. Plasticizer Bis(2-ethylhexyl) Phthalate causes meiosis defects and decreases fertilization ability of mouse oocytes in vivo. J Agric Food Chem. 2019;67(12):3459-68. https://doi.org/10.1021/acs.jafc.9b00121.
https://doi.org/10.1021/acs.jafc.9b00121...
). In equine, an increase in ROS and ATP levels was recorded in oocytes that were exposed to 12 µM DEHP (Ambruosi et al., 2011Ambruosi B, Uranio M, Sardanelli A, Pocar P, Martino N, Parternoster M, Amati F, Dell'Aquila ME. In Vitro acute exposure to DEHP affects Oocyte Meiotic maturation, energy and oxidative stress parameters in a large animal model. PLoS One. 2011;6(11):e27452. http://dx.doi.org/10.1371/journal.pone.0027452. PMid:22076161.
http://dx.doi.org/10.1371/journal.pone.0...
). Zhang et al. (2018)Zhang Y, Wang T, Lan M, Zang XW, Li YL, Cui XS, Kim NH, Sun SC. Melatonin protects oocytes from MEHP exposure-induced meiosis defects in porcine. Biol Reprod. 2018;98(3):286-98. http://doi.org//10.1093/biolre/iox185
http://doi.org//10.1093/biolre/iox185...
reported that in-vitro culturing of porcine COCs with 75 µM MEHP results in increased ROS production.
Changes in mitochondrial function along with the increased levels of ROS might further lead to apoptosis. Culturing of bovine COCs with 50 µM MEHP increased the proportion of apoptotic oocytes (TUNEL+) (Kalo and Roth, 2015Kalo D, Roth Z. Effects of mono(2-ethylhexyl)phthalate on cytoplasmic maturation of oocytes-the bovine model. Reprod Toxicol. 2015;53:141-51. http://dx.doi.org/10.1016/j.reprotox.2015.04.007. PMid:25900598.
http://dx.doi.org/10.1016/j.reprotox.201...
) and was associated with alteration in the expression of ASAH1 gene encodes the acid ceramidase enzyme, an antiapoptotic factor (Grossman et al., 2012Grossman D, Kalo D, Gendelman M, Roth Z. Effect of di-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl) phthalate on in vitro developmental competence of bovine oocytes. Cell Biol Toxicol. 2012;28(6):383-96. http://dx.doi.org/10.1007/s10565-012-9230-1. PMid:22956148.
http://dx.doi.org/10.1007/s10565-012-923...
). In-depth transcript screening and proteomic analysis strongly support the oocyte transcripts' sensitivity to MEHP, since alterations in several genes associated with mitochondria and the apoptotic cascade have been reported (Kalo et al., 2019Kalo D, Vitorino Carvalho A, Archilla C, Duranthon V, Moroldo M, Levin Y, Kupervaser M, Smith Y, Roth Z. Mono(2-ethylhexyl) phthalate (MEHP) induces transcriptomic alterations in oocytes and their derived blastocysts. Toxicology. 2019;421:59-73. http://dx.doi.org/10.1016/j.tox.2019.04.016. PMid:31059758.
http://dx.doi.org/10.1016/j.tox.2019.04....
). In support of this, MEHP induced alterations in the expression of genes associated with mitochondrial function in bovine COCs (cytochrome bc1 complex; CYC1, Mitochondrially Encoded Cytochrome C Oxidase I ;MT-CO1 and ATP synthase subunit beta; ATP5B), mouse fetal oocytes (superoxide dismutase 1 ;Sod1 and NADH dehydrogenase 1; Nd1) and porcine COCs (SOD1 and Catalase; CAT) (Kalo and Roth, 2015Kalo D, Roth Z. Effects of mono(2-ethylhexyl)phthalate on cytoplasmic maturation of oocytes-the bovine model. Reprod Toxicol. 2015;53:141-51. http://dx.doi.org/10.1016/j.reprotox.2015.04.007. PMid:25900598.
http://dx.doi.org/10.1016/j.reprotox.201...
; Bonilla and del Mazo, 2010Bonilla E, del Mazo J. Deregulation of the Sod1 and Nd1 genes in mouse fetal oocytes exposed to mono-(2-ethylhexyl) phthalate (MEHP). Reprod Toxicol. 2010;30(3):387-92. http://dx.doi.org/10.1016/j.reprotox.2010.04.008. PMid:20438828.
http://dx.doi.org/10.1016/j.reprotox.201...
; Zhang et al., 2018Zhang Y, Wang T, Lan M, Zang XW, Li YL, Cui XS, Kim NH, Sun SC. Melatonin protects oocytes from MEHP exposure-induced meiosis defects in porcine. Biol Reprod. 2018;98(3):286-98. http://doi.org//10.1093/biolre/iox185
http://doi.org//10.1093/biolre/iox185...
).
Carryover effect of DEHP and MEHP from the oocyte to the embryo
The early developing embryo, as well as the growing fetus, can also be exposed to phthalates during their growth in the uterus. In humans, DEHP and MEHP have been detected in maternal serum and cord blood, peritoneal fluid, meconium and amniotic fluid (Arbuckle et al., 2016Arbuckle TE, Fisher M, MacPherson S, Lang C, Provencher G, LeBlanc A, Hauser R, Feeley M, Ayotte P, Neisa A, Ramsay T, Tawagi G. Maternal and early life exposure to phthalates: The plastics and personal-care Products use in Pregnancy (P4) study. Sci Total Environ. 2016;551-552:344-56. http://dx.doi.org/10.1016/j.scitotenv.2016.02.022. PMid:26878646.
http://dx.doi.org/10.1016/j.scitotenv.20...
; Huang et al., 2014Huang Y, Li J, Garcia JM, Lin H, Wang Y, Yan P, Wang L, Tan Y, Luo J, Qiu Z, Chen JA, Shu W. Phthalate levels in cord blood are associated with preterm delivery and fetal growth parameters in chinese women. PLoS One. 2014;9(2):e87430. http://dx.doi.org/10.1371/journal.pone.0087430. PMid:24503621.
http://dx.doi.org/10.1371/journal.pone.0...
; Latini et al., 2003Latini G, De Felice C, Presta G, Del Vecchio A, Paris I, Ruggieri F, Mazzeo P. In utero exposure to di-(2-ethylhexyl)phthalate and duration of human pregnancy. Environ Health Perspect. 2003;111(14):1783-5. http://dx.doi.org/10.1289/ehp.6202. PMid:14594632.
http://dx.doi.org/10.1289/ehp.6202...
; Silva et al., 2004Silva MJ, Reidy JA, Herbert AR, Preau JL Jr, Needham LL, Calafat AM. Detection of phthalate metabolites in human amniotic fluid. Bull Environ Contam Toxicol. 2004;72(6):1226-31. http://dx.doi.org/10.1007/s00128-004-0374-4. PMid:15362453.
http://dx.doi.org/10.1007/s00128-004-037...
). Oral administration of DEHP (10 mg/kg BW) to female mice on day 7 or 8 of gestation led to a high incidence of death or malformed fetuses, whereas exposure before or after these crucial days resulted in fewer defective fetuses (Yagi et al., 1980Yagi Y, Nakamura Y, Tomita I, Tsuchikawa K, Shimoi N. Teratogenic potential of di- and mono-(2-ethylhexyl)phthalate in mice. J Environ Pathol Toxicol. 1980;4(2-3):533-44. PMid:7462917.). Exposure of mice on day 8 of gestation to 1 mL/kg BW MEHP resulted in skeletal abnormalities and fetal deaths (Tomita et al., 1982Tomita I, Nakamura Y, Aoki N, Inui N. Mutagenic/carcinogenic potential of DEHP and MEHP. Environ Health Perspect. 1982;45:119-25. http://dx.doi.org/10.1289/ehp.8245119. PMid:6814903.
http://dx.doi.org/10.1289/ehp.8245119...
; Yagi et al., 1980Yagi Y, Nakamura Y, Tomita I, Tsuchikawa K, Shimoi N. Teratogenic potential of di- and mono-(2-ethylhexyl)phthalate in mice. J Environ Pathol Toxicol. 1980;4(2-3):533-44. PMid:7462917.). In-vitro culture of 8-day-old mouse embryos with DEHP and MEHP (100–1000 µg/mL) impaired their development and increased the proportion of malformed embryos (Sant et al., 2016Sant KE, Dolinoy DC, Jilek JL, Sartor MA, Harris C. Mono-2-ethylhexyl phthalate disrupts neurulation and modifies the embryonic redox environment and gene expression. Reprod Toxicol. 2016;63:32-48. http://dx.doi.org/10.1016/j.reprotox.2016.03.042. PMid:27167697.
http://dx.doi.org/10.1016/j.reprotox.201...
).
Moreover, carryover effects from the oocyte to the developed embryo have been documented. Culturing of bovine COCs with either 50 µM DEHP or MEHP resulted in reduced oocyte developmental competence, expressed as a lower cleavage rate and reduced blastocyst-formation rate (Figure 1D, D'; Grossman et al., 2012Grossman D, Kalo D, Gendelman M, Roth Z. Effect of di-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl) phthalate on in vitro developmental competence of bovine oocytes. Cell Biol Toxicol. 2012;28(6):383-96. http://dx.doi.org/10.1007/s10565-012-9230-1. PMid:22956148.
http://dx.doi.org/10.1007/s10565-012-923...
). In-vitro exposure of bovine oocytes to relatively low levels of MEHP (20–1000 nM) decreased their developmental competence, reflected by a low proportion of oocytes that fertilized, cleaved and developed to the blastocyst stage (Kalo and Roth, 2017Kalo D, Roth Z. Low level of mono(2-ethylhexyl) phthalate reduces oocyte developmental competence in association with impaired gene expression. Toxicology. 2017;377:38-48. http://dx.doi.org/10.1016/j.tox.2016.12.005. PMid:27989758.
http://dx.doi.org/10.1016/j.tox.2016.12....
). In-vitro maturation of oocytes in follicular fluid aspirated from DEHP-treated cows (i.e., containing 20 nM MEHP) reduced cleavage and blastocyst-formation rates (Kalo et al., 2015Kalo D, Hadas R, Furman O, Ben-Ari J, Maor Y, Patterson DG, Tomey C, Roth Z. Carryover effects of acute DEHP exposure on ovarian function and oocyte developmental competence in lactating cows. PLoS One. 2015;10(7):e0130896. http://dx.doi.org/10.1371/journal.pone.0130896. PMid:26154164.
http://dx.doi.org/10.1371/journal.pone.0...
). Blastocysts developed from MEHP-treated oocytes showed different expression patterns of genes involved in intracellular mechanisms such as pluripotency, apoptosis, and placental development (Kalo et al., 2019Kalo D, Vitorino Carvalho A, Archilla C, Duranthon V, Moroldo M, Levin Y, Kupervaser M, Smith Y, Roth Z. Mono(2-ethylhexyl) phthalate (MEHP) induces transcriptomic alterations in oocytes and their derived blastocysts. Toxicology. 2019;421:59-73. http://dx.doi.org/10.1016/j.tox.2019.04.016. PMid:31059758.
http://dx.doi.org/10.1016/j.tox.2019.04....
). Culturing of bovine oocytes with 50 µM MEHP altered the levels of POU5F1 in 2-cell-stage embryos (Grossman et al., 2012Grossman D, Kalo D, Gendelman M, Roth Z. Effect of di-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl) phthalate on in vitro developmental competence of bovine oocytes. Cell Biol Toxicol. 2012;28(6):383-96. http://dx.doi.org/10.1007/s10565-012-9230-1. PMid:22956148.
http://dx.doi.org/10.1007/s10565-012-923...
). MEHP-induced impairment in POU5F1 and SOX2 transcript abundance which associated with pluripotency, was found in blastocysts developed from oocytes exposed to relatively low MEHP concentrations (Kalo and Roth, 2017Kalo D, Roth Z. Low level of mono(2-ethylhexyl) phthalate reduces oocyte developmental competence in association with impaired gene expression. Toxicology. 2017;377:38-48. http://dx.doi.org/10.1016/j.tox.2016.12.005. PMid:27989758.
http://dx.doi.org/10.1016/j.tox.2016.12....
), suggesting that these embryos are of inferior quality. In-vitro maturation of bovine oocytes with 20 nM MEHP induced a carryover effect on the transcriptomic profile of the developing embryo, expressed by the altered expression of 260 genes (Kalo et al., 2019Kalo D, Vitorino Carvalho A, Archilla C, Duranthon V, Moroldo M, Levin Y, Kupervaser M, Smith Y, Roth Z. Mono(2-ethylhexyl) phthalate (MEHP) induces transcriptomic alterations in oocytes and their derived blastocysts. Toxicology. 2019;421:59-73. http://dx.doi.org/10.1016/j.tox.2019.04.016. PMid:31059758.
http://dx.doi.org/10.1016/j.tox.2019.04....
). Direct exposure of mouse embryos to 100–1000 μg/mL MEHP (corresponding to 0.4–3.6 mM) impaired the expression of metabolic genes, in particular those related to the electron transport chain (Sant et al., 2016Sant KE, Dolinoy DC, Jilek JL, Sartor MA, Harris C. Mono-2-ethylhexyl phthalate disrupts neurulation and modifies the embryonic redox environment and gene expression. Reprod Toxicol. 2016;63:32-48. http://dx.doi.org/10.1016/j.reprotox.2016.03.042. PMid:27167697.
http://dx.doi.org/10.1016/j.reprotox.201...
). In agreement with this, a carryover effect was also recorded in female mice treated with DEHP or MEHP, manifested in a reduction in the proportion of developing blastocysts (Absalan et al., 2017Absalan F, Saremy S, Mansori E, Taheri Moghadam M, Eftekhari Moghadam AR, Ghanavati R. Effects of Mono-(2-Ethylhexyl) Phthalate and Di-(2-Ethylhexyl) Phthalate administrations on Oocyte Meiotic Maturation, Apoptosis and Gene Quantification in mouse model. Cell J. 2017;18(4):503-13. PMid:28042535.).
Summary
This section provides broad information on the direct and indirect effects of phthalates on ovarian function, oocyte maturation and embryonic developmental competence. Note that when the oocyte was exposed to MEHP, deleterious effects were further recorded in the developed embryos. In addition, phthalate-induced damage involves multiple mechanisms which might also be relevant to other EDCs.
Foodborne contaminants
A wide range of chemicals have been identified as foodborne contaminants that can be potentially found in food and water and might be consumed by humans and/or animals. These include pesticides and herbicides (ATZ, methoxychlor, chlorpyrifos, DDT) (Diamanti-Kandarakis et al., 2009Diamanti-Kandarakis E, Bourguignon J-P, Giudice LC, Hauser R, Prins GS, Soto AM, Zoeller RT, Gore AC. Endocrine-disrupting chemicals: an Endocrine Society scientific statement. Endocr Rev. 2009;30(4):293-342. http://dx.doi.org/10.1210/er.2009-0002. PMid:19502515.
http://dx.doi.org/10.1210/er.2009-0002...
). Among the most concerning foodborne contaminants are the naturally occurring toxins, such as mycotoxins, and specifically aflatoxins (El Khoury et al., 2019El Khoury D, Fayjaloun S, Nassar M, Sahakian J, Aad PY. Updates on the effect of mycotoxins on male reproductive efficiency in mammals. Toxins (Basel). 2019;11(9). http://dx.doi.org/10.3390/toxins11090515. PMid:31484408.
http://dx.doi.org/10.3390/toxins11090515...
). The current section focuses on two representative foodborne contaminants—the mycotoxin aflatoxin B1 (AFB1) and the herbicide ATZ—and discusses their toxic effects on bovine spermatozoa which carry over to the developed embryo.
Effect of foodborne contaminants on male reproduction
Studies in occupationally exposed humans, as well as animals, have shown that pesticides may cause pathological changes in the male reproductive system, such as testicular damage, decreased spermatogenesis, and reduced semen quality (Zamkowska et al., 2018Zamkowska D, Karwacka A, Jurewicz J, Radwan M. Environmental exposure to non-persistent endocrine disrupting chemicals and semen quality: an overview of the current epidemiological evidence. Int J Occup Med Environ Health. 2018;31(4):377-414. http://dx.doi.org/10.13075/ijomeh.1896.01195. PMid:30160090.
http://dx.doi.org/10.13075/ijomeh.1896.0...
). Pesticides and certain toxins, including mycotoxins, may pass through the blood–testis barrier and negatively affect spermatogenesis (Ataman et al., 2014Ataman MB, Bucak MN, Coyan K. Esterified glucomannan improves aflatoxin-induced damage of sperm parameters during liquid storage of ram semen at 5°C. Cryobiology. 2014;68(3):405-10. http://dx.doi.org/10.1016/j.cryobiol.2014.03.007. PMid:24662029.
http://dx.doi.org/10.1016/j.cryobiol.201...
; Perry, 2008Perry MJ. Effects of environmental and occupational pesticide exposure on human sperm: A systematic review. Hum Reprod Update. 2008;14(3):233-42. http://dx.doi.org/10.1093/humupd/dmm039. PMid:18281240.
http://dx.doi.org/10.1093/humupd/dmm039...
). Nevertheless, the direct effect of these foodborne contaminants on spermatozoa has been less studied, and the existing data are limited to basic parameters, such as spermatozoon concentration and motility. Moreover, the mechanism by which foodborne contaminants induce damage in spermatozoa is not fully understood.
Sperm fertilization potential depends on multiple factors. Among these are the integrity and functionality of sperm membranes, which indicate the sperm's viability and fertilization potential (Figure 2A; Komsky-Elbaz and Roth, 2018Komsky-Elbaz A, Roth Z. Fluorimetric techniques for the assessment of sperm membranes. J. Vis. Exp. 2018(141), e58622. http://dx.doi.org/10.3791/58 622.).
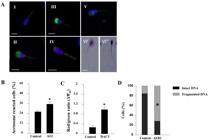
Effect of foodborne contaminants on spermatozoa. (A) Epifluorescence photomicrography of sperm cells stained with fluorescent probes: (I) live spermatozoon, nucleus stained with 4′,6-diamidino-2-phenylindole (DAPI; blue); (II) dead spermatozoon with damaged plasma membrane (propidium iodide staining, purple) and damaged acrosome (fluorescein-conjugated Pisum sativum agglutinin [FITC–PSA] staining, green); (III) live spermatozoon with damaged acrosome membrane (green); (IV) acrosome-reacted sperm cell with residual upper staining (green) and high mitochondrial membrane potential stained by JC-1 (red); (V) acrosome-reacted sperm cell with residual equatorial FITC–PSA staining (green); (VI) spermatozoon DNA fragmentation determined by Halosperm kit: (VI') spermatozoon with intact DNA expresses large halo and (VI”) spermatozoon with fragmented DNA expresses small or no halo. Scale bars = 10 µM (adopted with permission from Komsky-Elbaz and Roth, 2018Komsky-Elbaz A, Roth Z. Fluorimetric techniques for the assessment of sperm membranes. J. Vis. Exp. 2018(141), e58622. http://dx.doi.org/10.3791/58 622.). Spermatozoa were isolated from fresh ejaculate and incubated for 4 h without or with (B) 0.1 µM ATZ, presented is the proportion of acrosome-reacted cells (FITC–PSA); (C) 1 µM DACT, mitochondrial membrane potential (ΔΨm) is presented as the mean proportion of red-stained (high potential) to green-stained (low potential) (JC-1) (adopted with permission from Komsky-Elbaz and Roth, 2017Komsky-Elbaz A, Roth Z. Effect of the herbicide atrazine and its metabolite DACT on bovine sperm quality. Reprod Toxicol. 2017;67:15-25. http://dx.doi.org/10.1016/j.reprotox.2016.11.001. PMid:27836535.
http://dx.doi.org/10.1016/j.reprotox.201... ); (D) 10 µM AFB1, spermatozoon DNA fragmentation determined by Halosperm kit (adopted with permission from Komsky-Elbaz and Roth, 2018Komsky-Elbaz A, Roth Z. Fluorimetric techniques for the assessment of sperm membranes. J. Vis. Exp. 2018(141), e58622. http://dx.doi.org/10.3791/58 622.). Data are presented as mean proportion ± SD, *P < 0.05.
-
i
Plasma membrane Damage to plasma membrane integrity may reduce spermatozoa's viability and thus reduce their fertilization capacity (Komsky-Elbaz and Roth, 2017Komsky-Elbaz A, Roth Z. Effect of the herbicide atrazine and its metabolite DACT on bovine sperm quality. Reprod Toxicol. 2017;67:15-25. http://dx.doi.org/10.1016/j.reprotox.2016.11.001. PMid:27836535.
http://dx.doi.org/10.1016/j.reprotox.201... ). Various EDCs, such as BPA and phthalates, have been reported to decrease spermatozoon viability in humans (Pant et al., 2011Pant N, Pant A, Shukla M, Mathur N, Gupta Y, Saxena D. Environmental and experimental exposure of phthalate esters: the toxicological consequence on human sperm. Hum Exp Toxicol. 2011;30(6):507-14. http://dx.doi.org/10.1177/0960327110374205. PMid:20551087.
http://dx.doi.org/10.1177/09603271103742... ; Knez et al., 2014Knez J, Kranvogl R, Breznik BP, Vončina E, Vlaisavljević V. Are urinary bisphenol A levels in men related to semen quality and embryo development after medically assisted reproduction? Fertil Steril. 2014;101(1):215-221.e5. http://dx.doi.org/10.1016/j.fertnstert.2013.09.030. PMid:24182411.
http://dx.doi.org/10.1016/j.fertnstert.2... ), and the herbicide, fenoxaprop-ethyl, in porcine (Betancourt et al., 2006Betancourt M, Reséndiz A, Fierro ECYR. Effect of two insecticides and two herbicides on the porcine sperm motility patterns using computer-assisted semen analysis (CASA) in vitro. Reprod Toxicol. 2006;22(3):508-12. http://dx.doi.org/10.1016/j.reprotox.2006.03.001. PMid:16713176.
http://dx.doi.org/10.1016/j.reprotox.200... ). -
ii
Acrosome integrity Disruption of spermatozoon acrosomal membrane integrity leads to loss of its ability to undergo acrosome reaction (AR), resulting in reduced competence to attach to and penetrate the zona pellucida and fertilize the oocyte. Pseudo-AR (i.e., a non-controlled event which occurs via unknown factors such as environmental contaminants) can also lead to loss of fertilization ability (Wiser et al., 2014Wiser A, Sachar S, Ghetler Y, Shulman A, Breitbart H. Assessment of sperm hyperactivated motility and acrosome reaction can discriminate the use of spermatozoa for conventional in vitro fertilisation or intracytoplasmic sperm injection: preliminary results. Andrologia. 2014;46(3):313-5. http://dx.doi.org/10.1111/and.12068. PMid:23356546.
http://dx.doi.org/10.1111/and.12068... ). -
iii
Mitochondrial function Alterations in mitochondrial function are associated with physiological dysfunction, including male infertility (Ramalho-Santos et al., 2009Ramalho-Santos J, Varum S, Amaral S, Mota PC, Sousa AP, Amaral A. Mitochondrial functionality in reproduction: from gonads and gametes to embryos and embryonic stem cells. Hum Reprod Update. 2009;15(5):553-72. http://dx.doi.org/10.1093/humupd/dmp016. PMid:19414527.
http://dx.doi.org/10.1093/humupd/dmp016... ). In humans, loss of ΔΨm is associated with low spermatozoon quality and caspase activation, which might eventually lead to apoptosis. Spermatozoa with altered ΔΨm generally have reduced fertilization potential and exhibit excess production of ROS (Espinoza et al., 2009Espinoza JA, Paasch U, Villegas JV. Mitochondrial membrane potential disruption pattern in human sperm. Hum Reprod. 2009;24(9):2079-85. http://dx.doi.org/10.1093/humrep/dep120. PMid:19465458.
http://dx.doi.org/10.1093/humrep/dep120... ). Various environmental compounds, such as EDCs, have been shown to induce cellular stress and lead to a transient increase in ΔΨm, i.e., hyperpolarization (Hüttemann et al., 2008Hüttemann M, Lee I, Pecinova A, Pecina P, Przyklenk K, Doan JW. Regulation of oxidative phosphorylation, the mitochondrial membrane potential, and their role in human disease. J Bioenerg Biomembr. 2008;40(5):445-56. http://dx.doi.org/10.1007/s10863-008-9169-3. PMid:18843528.
http://dx.doi.org/10.1007/s10863-008-916... ), thereby leading to the generation of ROS (Saradha and Mathur, 2006Saradha B, Mathur PP. Effect of environmental contaminants on male reproduction. Environ Toxicol Pharmacol. 2006;21(1):34-41. http://dx.doi.org/10.1016/j.etap.2005.06.004. PMid:21783636.
http://dx.doi.org/10.1016/j.etap.2005.06... ). In turn, ROS can directly attack unsaturated fatty acids on the spermatozoon membrane, induce lipid peroxidation, damage membrane integrity, and eventually reduce the spermatozoa's fertilization potential (Lavranos et al., 2012Lavranos G, Balla M, Tzortzopoulou A, Syriou V, Angelopoulou R. Investigating ROS sources in male infertility: a common end for numerous pathways. Reprod Toxicol. 2012;34(3):298-307. http://dx.doi.org/10.1016/j.reprotox.2012.06.007. PMid:22749934.
http://dx.doi.org/10.1016/j.reprotox.201... ). Mammalian spermatozoon membranes are rich in polyunsaturated fatty acids and thus are more susceptible to ROS attack, resulting in decreased sperm motility and viability, leading to infertility (Saradha and Mathur, 2006Saradha B, Mathur PP. Effect of environmental contaminants on male reproduction. Environ Toxicol Pharmacol. 2006;21(1):34-41. http://dx.doi.org/10.1016/j.etap.2005.06.004. PMid:21783636.
http://dx.doi.org/10.1016/j.etap.2005.06... ). -
iv
DNA fragmentation The proportion of sperm with DNA fragmentation correlates with male fertility and is considered a practical parameter for characterizing semen quality (Sergerie et al., 2005Sergerie M, Laforest G, Bujan L, Bissonnette F, Bleau G. Sperm DNA fragmentation: threshold value in male fertility. Hum Reprod. 2005;20(12):3446-51. http://dx.doi.org/10.1093/humrep/dei231. PMid:16085665.
http://dx.doi.org/10.1093/humrep/dei231... ). DNA molecules are a known target for various drugs, pesticides and toxic compounds (Ma et al., 2017Ma L, Wang J, Zhang Y. Probing the characterization of the interaction of Aflatoxins B1 and G1 with Calf Thymus DNA In Vitro. Toxins (Basel). 2017;9(7):209. http://dx.doi.org/10.3390/toxins9070209. PMid:28671585.
http://dx.doi.org/10.3390/toxins9070209... ; Virro et al., 2004Virro MR, Larson-Cook KL, Evenson DP. Sperm chromatin structure assay (SCSA®) parameters are related to fertilization, blastocyst development, and ongoing pregnancy in in vitro fertilization and intracytoplasmic sperm injection cycles. Fertil Steril. 2004;81(5):1289-95. http://dx.doi.org/10.1016/j.fertnstert.2003.09.063. PMid:15136092.
http://dx.doi.org/10.1016/j.fertnstert.2... ). DNA damage was shown in subfertile males with increased BPA concentration in the urine (Meeker et al., 2010Meeker JD, Ehrlich S, Toth TL, Wright DL, Calafat AM, Trisini AT, Ye X, Hauser R. Semen quality and sperm DNA damage in relation to urinary bisphenol A among men from an infertility clinic. Reprod Toxicol. 2010;30(4):532-9. http://dx.doi.org/10.1016/j.reprotox.2010.07.005. PMid:20656017.
http://dx.doi.org/10.1016/j.reprotox.201... ). In humans, increased DNA damage was documented after environmental and occupational exposure to the pesticide fenvalerate (Bian et al., 2004Bian Q, Xu LC, Wang SL, Xia YK, Tan LF, Chen JF, Song L, Chang HC, Wang XR. Study on the relation between occupational fenvalerate exposure and spermatozoa DNA damage of pesticide factory workers. Occup Environ Med. 2004;61(12):999-1005. http://dx.doi.org/10.1136/oem.2004.014597. PMid:15550606.
http://dx.doi.org/10.1136/oem.2004.01459... ) and to a mixture of organophosphorous substances (Sánchez-Peña et al., 2004Sánchez-Peña LC, Reyes BE, López-Carrillo L, Recio R, Morán-Martínez J, Cebrián ME, Quintanilla-Vega B. Organophosphorous pesticide exposure alters sperm chromatin structure in Mexican agricultural workers. Toxicol Appl Pharmacol. 2004;196(1):108-13. http://dx.doi.org/10.1016/j.taap.2003.11.023. PMid:15050412.
http://dx.doi.org/10.1016/j.taap.2003.11... ). -
v
Epigenetic modifications The environmental effect on the gametes is not limited to the obvious physiological and genetic effects. Epigenetic modifications and DNA methylation in the germline can be altered by various environmental factors (Gely-Pernot et al., 2015Gely-Pernot A, Hao C, Becker E, Stuparevic I, Kervarrec C, Chalmel F, Primig M, Jégou B, Smagulova F. The epigenetic processes of meiosis in male mice are broadly affected by the widely used herbicide atrazine. BMC Genomics. 2015;16(1):885. http://dx.doi.org/10.1186/s12864-015-2095-y. PMid:26518232.
http://dx.doi.org/10.1186/s12864-015-209... ; Siddeek et al., 2018Siddeek B, Mauduit C, Simeoni U, Benahmed M. Sperm epigenome as a marker of environmental exposure and lifestyle, at the origin of diseases inheritance. Mutat Res Mutat Res. 2018;778:38-44. http://dx.doi.org/10.1016/j.mrrev.2018.09.001. PMid:30454681.
http://dx.doi.org/10.1016/j.mrrev.2018.0... ). Changes in differentially methylated regions were recorded in spermatozoa exposed to the fungicide vinclozolin (Skinner et al., 2019Skinner MK, Nilsson E, Sadler-Riggleman I, Beck D, Ben Maamar M, McCarrey JR. Transgenerational sperm DNA methylation epimutation developmental origins following ancestral vinclozolin exposure. Epigenetics. 2019;14(7):721-39. http://dx.doi.org/10.1080/15592294.2019.1614417. PMid:31079544.
http://dx.doi.org/10.1080/15592294.2019.... ). Alterations in spermatozoa's DNA-methylation profile were reported upon paternal exposure to PBA and phthalates (Manikkam et al., 2013Manikkam M, Tracey R, Guerrero-Bosagna C, Skinner MK. Plastics derived endocrine disruptors (BPA, DEHP and DBP) induce epigenetic transgenerational inheritance of obesity, reproductive disease and sperm epimutations. PLoS One. 2013;8(1):e55387. http://dx.doi.org/10.1371/journal.pone.0055387. PMid:23359474.
http://dx.doi.org/10.1371/journal.pone.0... ).
Effect of aflatoxins on the spermatozoa
Aflatoxins are low-molecular-weight compounds produced mainly by the fungi Aspergillus flavus and Aspergillus parasiticus (Dai et al., 2017Dai Y, Huang K, Zhang B, Zhu L, Xu W. Aflatoxin B1-induced epigenetic alterations: an overview. Food Chem Toxicol. 2017;109(Pt 1):683-9. http://dx.doi.org/10.1016/j.fct.2017.06.034. PMid:28645871.
http://dx.doi.org/10.1016/j.fct.2017.06....
; Kew, 2013Kew MC. Aflatoxins as a cause of hepatocellular carcinoma. J Gastrointestin Liver Dis. 2013;22(3):305-10. PMid:24078988.). Under humid conditions, these fungi grow on food grains, fruit and nuts, among others (Schenzel et al., 2012Schenzel J, Forrer H-R, Vogelgsang S, Hungerbühler K, Bucheli TD. Mycotoxins in the Environment: I. Production and Emission from an Agricultural Test Field. Environ Sci Technol. 2012;46(24):13067-75. http://dx.doi.org/10.1021/es301557m. PMid:23145781.
http://dx.doi.org/10.1021/es301557m...
; Strosnider et al., 2006Strosnider H, Azziz-Baumgartner E, Banziger M, Bhat RV, Breiman R, Brune M-N, DeCock K, Dilley A, Groopman J, Hell K, Henry SH, Jeffers D, Jolly C, Jolly P, Kibata GN, Lewis L, Liu X, Luber G, McCoy L, Mensah P, Miraglia M, Misore A, Njapau H, Ong CN, Onsongo MT, Page SW, Park D, Patel M, Phillips T, Pineiro M, Pronczuk J, Rogers HS, Rubin C, Sabino M, Schaafsma A, Shephard G, Stroka J, Wild C, Williams JT, Wilson D. Workgroup Report: Public Health Strategies for Reducing Aflatoxin Exposure in Developing Countries. Environ Health Perspect. 2006;114(12):1898-903. http://dx.doi.org/10.1289/ehp.9302. PMid:17185282.
http://dx.doi.org/10.1289/ehp.9302...
). Aflatoxin contamination can occur at any stage of the food chain (Verma, 2004Verma RJ. Aflatoxin cause DNA damage. Int J Hum Genet. 2004;4(4):231-6. http://dx.doi.org/10.1080/09723757.2004.11885899.
http://dx.doi.org/10.1080/09723757.2004....
), from preharvest to storage (Iimura et al., 2017Iimura K, Furukawa T, Yamamoto T, Negishi L, Suzuki M, Sakuda S. The mode of action of Cyclo(l-Ala-l-Pro) in Inhibiting Aflatoxin Production of Aspergillus flavus. Toxins (Basel). 2017;9(7):219. http://dx.doi.org/10.3390/toxins9070219. PMid:28704973.
http://dx.doi.org/10.3390/toxins9070219...
; Shuaib et al., 2010Shuaib FMB, Ehiri J, Abdullahi A, Williams JH, Jolly PE. Reproductive health effects of aflatoxins: a review of the literature. Reprod Toxicol. 2010;29(3):262-70. http://dx.doi.org/10.1016/j.reprotox.2009.12.005. PMid:20045048.
http://dx.doi.org/10.1016/j.reprotox.200...
). According to the US Food and Drug Administration the allowed amount of aflatoxins in human feed, is 4 to 30 ppb (0.01–0.1 μM), while grains for animal feeding can have up to 300 ppb (1 μM) (Williams et al., 2004Williams JH, Phillips TD, Jolly PE, Stiles JK, Jolly CM, Aggarwal D. Human aflatoxicosis in developing countries: a review of toxicology, exposure, potential health consequences, and interventions. Am J Clin Nutr. 2004;80(5):1106-22. http://dx.doi.org/10.1093/ajcn/80.5.1106. PMid:15531656.
http://dx.doi.org/10.1093/ajcn/80.5.1106...
).
Of the aflatoxins, AFB1 is considered the most toxic to mammals, owing to its hepatotoxic, teratogenic, mutagenic and immunosuppressive properties (Raisuddin et al., 1993Raisuddin S, Singh KP, Zaidi SI, Paul BN, Ray PK. Immunosuppressive effects of aflatoxin in growing rats. Mycopathologia. 1993;124(3):189-94. http://dx.doi.org/10.1007/BF01103737. PMid:8022466.
http://dx.doi.org/10.1007/BF01103737...
; Shen et al., 1994Shen HM, Shi CY, Lee HP, Ong CN. Aflatoxin B1-induced lipid peroxidation in rat liver. Toxicol Appl Pharmacol. 1994;127(1):145-50. http://dx.doi.org/10.1006/taap.1994.1148. PMid:8048046.
http://dx.doi.org/10.1006/taap.1994.1148...
; Meissonnier et al., 2008Meissonnier GM, Pinton P, Laffitte J, Cossalter A-M, Gong YY, Wild CP, Bertin G, Galtier P, Oswald IP. Immunotoxicity of aflatoxin B1: impairment of the cell-mediated response to vaccine antigen and modulation of cytokine expression. Toxicol Appl Pharmacol. 2008;231(2):142-9. http://dx.doi.org/10.1016/j.taap.2008.04.004. PMid:18501398.
http://dx.doi.org/10.1016/j.taap.2008.04...
). AFB1 is also defined as an EDC as it affects the cytochrome P450 enzymes, which are involved in steroid synthesis (Storvik et al., 2011Storvik M, Huuskonen P, Kyllönen T, Lehtonen S, El-Nezami H, Auriola S, Pasanen M. Aflatoxin B1 – a potential endocrine disruptor – up-regulates CYP19A1 in JEG-3 cells. Toxicol Lett. 2011;202(3):161-7. http://dx.doi.org/10.1016/j.toxlet.2011.01.028. PMid:21296134.
http://dx.doi.org/10.1016/j.toxlet.2011....
). Consumption of AFB1 leads to rapid metabolism into several metabolites, including aflatoxin M1 (AFM1).
Aflatoxins are of great concern to public health because they accumulate in the body and can be found in edible tissues, such as liver and muscles, as well as in animal food products such as milk and eggs (Fan et al., 2015Fan S, Li Q, Sun L, Du Y, Xia J, Zhang Y. Simultaneous determination of aflatoxin B 1 and M 1 in milk, fresh milk and milk powder by LC-MS/MS utilising online turbulent flow chromatography. Food Addit Contam Part A Chem Anal Control Expo Risk Assess. 2015;32(7):1175-84. http://dx.doi.org/10.1080/19440049.2015.1048311. PMid:25952817.
http://dx.doi.org/10.1080/19440049.2015....
; Giovati et al., 2015Giovati L, Magliani W, Ciociola T, Santinoli C, Conti S, Polonelli L. AFM1 in Milk: Physical, biological, and prophylactic methods to mitigate contamination. Toxins (Basel). 2015;7(10):4330-49. http://dx.doi.org/10.3390/toxins7104330. PMid:26512694.
http://dx.doi.org/10.3390/toxins7104330...
; Monson et al., 2016Monson M, Cardona C, Coulombe R, Reed K. Hepatic Transcriptome Responses of Domesticated and Wild Turkey Embryos to Aflatoxin B1. Toxins (Basel). 2016;8(1):16. http://dx.doi.org/10.3390/toxins8010016. PMid:26751476.
http://dx.doi.org/10.3390/toxins8010016...
; Rajkovic et al., 2007Rajkovic A, Uyttendaele M, Debevere J. Computer aided boar semen motility analysis for cereulide detection in different food matrices. Int J Food Microbiol. 2007;114(1):92-9. http://dx.doi.org/10.1016/j.ijfoodmicro.2006.09.031. PMid:17174428.
http://dx.doi.org/10.1016/j.ijfoodmicro....
; Verma and Nair, 2001Verma RJ, Nair A. Ameliorative effect of vitamin E on aflatoxin-induced lipid peroxidation in the testis of mice. Asian J Androl. 2001;3(3):217-21. PMid:11561193.; Yuan et al., 2016Yuan S, Wu B, Yu Z, Fang J, Liang N, Zhou M, Huang C, Peng X. The mitochondrial and endoplasmic reticulum pathways involved in the apoptosis of bursa of Fabricius cells in broilers exposed to dietary aflatoxin B1. Oncotarget. 2016;7(40):65295-306. http://dx.doi.org/10.18632/oncotarget.11321. PMid:27542244.
http://dx.doi.org/10.18632/oncotarget.11...
). Moreover, aflatoxins have been found in human maternal breast milk, and maternal and cord blood (Shuaib et al., 2010Shuaib FMB, Ehiri J, Abdullahi A, Williams JH, Jolly PE. Reproductive health effects of aflatoxins: a review of the literature. Reprod Toxicol. 2010;29(3):262-70. http://dx.doi.org/10.1016/j.reprotox.2009.12.005. PMid:20045048.
http://dx.doi.org/10.1016/j.reprotox.200...
), and can apparently enter the developing fetus in humans and animals (Verma, 2004Verma RJ. Aflatoxin cause DNA damage. Int J Hum Genet. 2004;4(4):231-6. http://dx.doi.org/10.1080/09723757.2004.11885899.
http://dx.doi.org/10.1080/09723757.2004....
). AFM1 can also be found in milk and in other dairy products. It is resistant to heat and not fully inactivated by pasteurization or sterilization (Becker-Algeri et al., 2016Becker-Algeri TA, Castagnaro D, Bortoli K, Souza C, Drunkler DA, Badiale-Furlong E. Mycotoxins in bovine milk and dairy products: A review. J Food Sci. 2016;81(3):R544-52. http://dx.doi.org/10.1111/1750-3841.13204. PMid:26799355.
http://dx.doi.org/10.1111/1750-3841.1320...
). In the USA and some Asian countries, the maximum residual level of AFM1 in milk is 0.5 ppb (Becker-Algeri et al., 2016Becker-Algeri TA, Castagnaro D, Bortoli K, Souza C, Drunkler DA, Badiale-Furlong E. Mycotoxins in bovine milk and dairy products: A review. J Food Sci. 2016;81(3):R544-52. http://dx.doi.org/10.1111/1750-3841.13204. PMid:26799355.
http://dx.doi.org/10.1111/1750-3841.1320...
). The Israeli Public Health Services – Food Control Services (2008-2012) have reported milk samples containing AFM1 at levels of 0.08–0.10 ppb.
Although aflatoxins are considered one of the major risk factors for infertility (Gong et al., 2016Gong YY, Watson S, Routledge MN. Aflatoxin exposure and associated human health effects, a review of epidemiological studies. Food Saf. 2016;4(1):14-27. http://dx.doi.org/10.14252/foodsafetyfscj.2015026. PMid:32231900.
http://dx.doi.org/10.14252/foodsafetyfsc...
), there is limited information on their effect on the male gametes (i.e., spermatozoa), in particular following exposure to allowable regulated levels. Evaluation of semen samples revealed the presence of aflatoxins in 8% of the samples collected from fertile men, compared to 40% of those collected from infertile men (Ibeh et al., 1994Ibeh IN, Uraih N, Ogonar JI. Dietary exposure to aflatoxin in human male infertility in Benin City, Nigeria. Int J Fertil Menopausal Stud. 1994;39(4):208-14. PMid:7951403.). Human exposure to aflatoxin is associated with abnormal spermatozoon count and morphology, as well as reduced motility (Ibeh et al., 1994Ibeh IN, Uraih N, Ogonar JI. Dietary exposure to aflatoxin in human male infertility in Benin City, Nigeria. Int J Fertil Menopausal Stud. 1994;39(4):208-14. PMid:7951403.; Nduka et al., 2001Nduka U, Ibeh NI, Oluwafemi F. A Study on the Impact of Aflatoxin on Human Reproduction. Afr J Reprod Health. 2001;5(1):106-10. http://dx.doi.org/10.2307/3583204.
http://dx.doi.org/10.2307/3583204...
). Administration of 50 µg/kg BW AFB1 daily to mice for 45 days led to upregulation of genes involved in cell differentiation, immunity and the extracellular matrix in the testes. These alterations led to disruption of androgen production in Leydig cells (Austin et al., 2012Austin KJ, Cockrum RR, Jons AM, Alexander BM, Cammack KM. Renin mRNA is upregulated in testes and testicular cells in response to treatment with aflatoxin B1. Theriogenology. 2012;77:331-7.e1-7. http://dx.doi.org/10.1016/j.theriogenology.2011.08.004.
http://dx.doi.org/10.1016/j.theriogenolo...
). In rats, a dose-dependent decrease in the number of spermatozoa that developed in the seminiferous tubules was observed following administration with 0.8–3.2 ppm AFB1 for 48 days (Hasanzadeh and Rezazadeh, 2013Hasanzadeh S, Rezazadeh L. Effects of aflatoxin B1 on the growth processes of spermatogenic cell series in adult male rats. Comp Clin Pathol. 2013;22(4):555-62. http://dx.doi.org/10.1007/s00580-012-1445-2.
http://dx.doi.org/10.1007/s00580-012-144...
). Rats administered 10–50 µg AFB1 for 60 days showed decreased steroidogenesis, and reduced reproductive organ weight, sperm quantity, and sperm quality (Supriya et al., 2014Supriya C, Girish BP, Reddy PS. Aflatoxin B1-Induced Reproductive Toxicity in Male Rats: Possible Mechanism of Action. Int J Toxicol. 2014;33(3):155-61. http://dx.doi.org/10.1177/1091581814530764. PMid:24728861.
http://dx.doi.org/10.1177/10915818145307...
). Oral administration of AFB1 (50 µg/kg BW per day for 35 days) resulted in spermatotoxic effects on mouse epididymal sperm, expressed as reduced sperm concentration and motility and increased sperm abnormalities (Agnes and Akbarsha, 2003Agnes VF, Akbarsha MA. Spermatotoxic effect of aflatoxin B(1) in the albino mouse. Food Chem Toxicol. 2003;41(1):119-30. http://dx.doi.org/10.1016/S0278-6915(02)00171-0. PMid:12453736.
http://dx.doi.org/10.1016/S0278-6915(02)...
). Intramuscular injection of AFB1 (0.2–0.25 mL/day for 55 days) into male rats resulted in extrusion of the outer dense fibers and axonemal microtubule doublets of the sperm flagellum (Faisal et al., 2008aFaisal K, Periasamy VS, Sahabudeen S, Radha A, Anandhi R, Akbarsha MA. Spermatotoxic effect of aflatoxin B1 in rat: extrusion of outer dense fibres and associated axonemal microtubule doublets of sperm flagellum. Reproduction. 2008a;135(3):303-10. http://dx.doi.org/10.1530/REP-07-0367. PMid:18299423.
http://dx.doi.org/10.1530/REP-07-0367...
), suggesting negative effects on spermatozoa throughout the early stages of spermatogenesis.
In bovine, exposing spermatozoa to 10 µM AFB1 for 4 h decreased their viability, but did not activate the AR. In addition, hyperpolarization of the mitochondrial membrane was documented (Komsky-Elbaz et al., 2018Komsky-Elbaz A, Saktsier M, Roth Z. Aflatoxin B1 impairs sperm quality and fertilization competence. Toxicology. 2018;393:42-50. http://dx.doi.org/10.1016/j.tox.2017.11.007. PMid:29113834.
http://dx.doi.org/10.1016/j.tox.2017.11....
). Direct interaction of AFB1 with the cells' DNA, generating reversible non-covalent interactions, was demonstrated (Ma et al., 2017Ma L, Wang J, Zhang Y. Probing the characterization of the interaction of Aflatoxins B1 and G1 with Calf Thymus DNA In Vitro. Toxins (Basel). 2017;9(7):209. http://dx.doi.org/10.3390/toxins9070209. PMid:28671585.
http://dx.doi.org/10.3390/toxins9070209...
). It was suggested that binding of AFB1 to the DNA of mammalian spermatogenic cells (Sotomayor and Sega, 2000Sotomayor RE, Sega GA. Unscheduled DNA synthesis assay in mammalian spermatogenic cells: an update. Environ Mol Mutagen. 2000;36(4):255-65. http://dx.doi.org/10.1002/1098-2280(2000)36:4<255::AID-EM1>3.0.CO;2-O. PMid:11152558.
http://dx.doi.org/10.1002/1098-2280(2000...
) and epididymal spermatozoa (Faisal et al., 2008bFaisal K, Faridha A, Akbarsha MA. Induction of meiotic micronuclei in spermatocytes in vivo by aflatoxin B1: light and transmission electron microscopic study in Swiss mouse. Reprod Toxicol. 2008b;26:303-9. http://dx.doi.org/10.1016/j.reprotox.2008.09.003.
http://dx.doi.org/10.1016/j.reprotox.200...
) interferes with normal spermatogenesis, resulting in the production of abnormal spermatozoa and reduced fertilization. Exposure of bovine spermatozoa to 10 µM AFB1 significantly increased the proportion of DNA damage (Figure 2D; Komsky-Elbaz et al., 2018Komsky-Elbaz A, Saktsier M, Roth Z. Aflatoxin B1 impairs sperm quality and fertilization competence. Toxicology. 2018;393:42-50. http://dx.doi.org/10.1016/j.tox.2017.11.007. PMid:29113834.
http://dx.doi.org/10.1016/j.tox.2017.11....
).
To date, very few studies have addressed the effect of mycotoxins on the male reproductive epigenome (El Khoury et al., 2019El Khoury D, Fayjaloun S, Nassar M, Sahakian J, Aad PY. Updates on the effect of mycotoxins on male reproductive efficiency in mammals. Toxins (Basel). 2019;11(9). http://dx.doi.org/10.3390/toxins11090515. PMid:31484408.
http://dx.doi.org/10.3390/toxins11090515...
). In porcine, AFB1 at 50 µM induced alterations in histone, DNA methylation and apoptosis, and increased ROS levels (Liu et al., 2015Liu J, Wang Q-C, Han J, Xiong B, Sun S-C. Aflatoxin B1 is toxic to porcine oocyte maturation. Mutagenesis. 2015;30(4):527-35. http://dx.doi.org/10.1093/mutage/gev015. PMid:25778688.
http://dx.doi.org/10.1093/mutage/gev015...
). A recent proteomics approach to explore possible mechanisms by which AFB1 induces alterations in spermatozoa revealed differential expression of multiple proteins related to several cellular pathways and biological processes, and associated with genetic and epigenetic processes (Komsky-Elbaz and Roth, 2019Komsky-Elbaz A, Roth Z. Atrazine-induced DNA fragmentation in bovine spermatozoa is associated with alterations in the transcriptome profile of in-vitro-derived embryos. 2019. In: Abstracts of the 35th Annual Meeting of the Association of Embryo Technology in Europe (AETE); 2019 Sept 13-14; Murcia, Spain. Murcia: AETE. ).
Effect of ATZ on the spermatozoa
ATZ is one of the best known chlorotriazine herbicides, extensively used to control growth of broadleaf and grassy weeds in agricultural crops (Severi-Aguiar and Silva-Zacarin, 2011Severi-Aguiar GDC, Silva-Zacarin ECM. Effects of herbicide atrazine in experimental animal models. In: Hasaneen MN. Herbicides: Properties, synthesis and control of weeds. Rijeka: InTech; 2011. p. 285-96.). ATZ is considered a ubiquitous environmental contaminant, as it is frequently detected in ground and surface water, as a result of its mobility in the soil (Barr et al., 2007Barr DB, Panuwet P, Nguyen JV, Udunka S, Needham LL. Assessing exposure to atrazine and its metabolites using biomonitoring. Environ Health Perspect. 2007;115(10):1474-8. http://dx.doi.org/10.1289/ehp.10141. PMid:17938738.
http://dx.doi.org/10.1289/ehp.10141...
). ATZ has been banned for use in Europe since 2004, but is still utilized in about 70 countries, including US, Brazil, Argentina, Mexico, China, and Israel (LeBaron et al., 2008LeBaron HM, McFarland JE, Burnside OC. The Triazine Herbicides. Amsterdam: Elsevier; 2008. https://doi.org/10.1016/B978-044451167-6.50004-0.
https://doi.org/10.1016/B978-044451167-6...
). Once it enters the body, ATZ is rapidly metabolized in the liver by cytochrome P450 enzymes into several metabolites, which are detected in the urine, serum and various tissues (Barr et al., 2007Barr DB, Panuwet P, Nguyen JV, Udunka S, Needham LL. Assessing exposure to atrazine and its metabolites using biomonitoring. Environ Health Perspect. 2007;115(10):1474-8. http://dx.doi.org/10.1289/ehp.10141. PMid:17938738.
http://dx.doi.org/10.1289/ehp.10141...
; Dooley et al., 2013Dooley GP, Tjalkens RB, Hanneman WH. The atrazine metabolite diaminochlorotriazine suppresses LH release from murine LβT2 cells by suppressing GnRH-induced intracellular calcium transients. Toxicol Res (Camb). 2013;2(3):180-6. http://dx.doi.org/10.1039/c3tx20088d. PMid:24052811.
http://dx.doi.org/10.1039/c3tx20088d...
; Ross et al., 2009Ross MK, Jones TL, Filipov NM. Disposition of the Herbicide 2-Chloro-4-(ethylamino)-6-(isopropylamino)-s-triazine (Atrazine) and its major metabolites in mice: A liquid chromatography / mass spectrometry analysis of urine. Plasma, and Tissue Levels. 2009;37(4):776-86. http://dx.doi.org/10.1124/dmd.108.024927. PMid:19116264.
http://dx.doi.org/10.1124/dmd.108.024927...
); thus, an indirect effect of ATZ through its metabolites cannot be ruled out. In mammals, the major and most frequently detected metabolite is diaminochlorotriazine (DACT), shown to induce oxidative stress and disrupt endocrine function (Forgacs et al., 2013Forgacs AL, D’Souza ML, Huhtaniemi IT, Rahman NA, Zacharewski TR. Triazine Herbicides and Their Chlorometabolites Alter Steroidogenesis in BLTK1 Murine Leydig Cells. ToxSci 2013;134:155-67. https://doi.org/10.1093/toxsci/kft096.
https://doi.org/10.1093/toxsci/kft096...
; Jin et al., 2014Jin Y, Wang L, Chen G, Lin X, Miao W, Fu Z. Exposure of mice to atrazine and its metabolite diaminochlorotriazine elicits oxidative stress and endocrine disruption. Environ Toxicol Pharmacol. 2014;37(2):782-90. http://dx.doi.org/10.1016/j.etap.2014.02.014. PMid:24632104.
http://dx.doi.org/10.1016/j.etap.2014.02...
). DACT forms covalent adducts with various proteins, presumably as a chemical-induced toxicity step (Dooley et al., 2008Dooley GP, Reardon KF, Prenni JE, Tjalkens RB, Legare ME, Foradori CD, Tessari JE, Hanneman WH. Proteomic analysis of diaminochlorotriazine adducts in wister rat pituitary glands and L T2 rat pituitary cells 2008:21(4):844-51.).
Studies in humans have provided evidence for the transfer of trace amounts of ATZ to the circulation via the food chain (Diamanti-Kandarakis et al., 2009Diamanti-Kandarakis E, Bourguignon J-P, Giudice LC, Hauser R, Prins GS, Soto AM, Zoeller RT, Gore AC. Endocrine-disrupting chemicals: an Endocrine Society scientific statement. Endocr Rev. 2009;30(4):293-342. http://dx.doi.org/10.1210/er.2009-0002. PMid:19502515.
http://dx.doi.org/10.1210/er.2009-0002...
; Peighambarzadeh et al., 2011Peighambarzadeh S, Safi S, Shahtaheri S, Javanbakht M, Rahimi Forushani A. Rahimi Forushani a. Presence of atrazine in the biological samples of cattle and its consequence adversity in human health. Iran J Public Health. 2011;40(4):112-21. PMid:23113110.). For instance, ATZ has been detected in human amniotic fluid (0.6 µg/L) and urine (0.1 µg/L) (Bradman et al., 2003Bradman A, Barr DB, Claus Henn BG, Drumheller T, Curry C, Eskenazi B. Measurement of pesticides and other toxicants in amniotic fluid as a potential biomarker of prenatal exposure: A validation study. Environ Health Perspect. 2003;111(14):1779-82. http://dx.doi.org/10.1289/ehp.6259. PMid:14594631.
http://dx.doi.org/10.1289/ehp.6259...
). Humans, wildlife, and domesticated animals can be exposed to ATZ via consumption of contaminated food or water, inhalation of pesticide spray, or absorption through the skin (Barr et al., 2007Barr DB, Panuwet P, Nguyen JV, Udunka S, Needham LL. Assessing exposure to atrazine and its metabolites using biomonitoring. Environ Health Perspect. 2007;115(10):1474-8. http://dx.doi.org/10.1289/ehp.10141. PMid:17938738.
http://dx.doi.org/10.1289/ehp.10141...
; Chevrier et al., 2011Chevrier C, Limon G, Monfort C, Rouget F, Garlantézec R, Petit C, Durand G, Cordier S. Urinary biomarkers of prenatal atrazine exposure and adverse birth outcomes in the PELAGIE birth cohort. Environ Health Perspect. 2011;119(7):1034-41. http://dx.doi.org/10.1289/ehp.1002775. PMid:21367690.
http://dx.doi.org/10.1289/ehp.1002775...
). Several studies have suggested that ATZ can alter reproductive functions, even at low, ecologically relevant doses (0.1–3 µg/L) (Abarikwu et al., 2010Abarikwu SO, Adesiyan AC, Oyeloja TO, Oyeyemi MO, Farombi EO. Changes in sperm characteristics and induction of oxidative stress in the testis and epididymis of experimental rats by a herbicide, atrazine. Arch Environ Contam Toxicol. 2010;58(3):874-82. http://dx.doi.org/10.1007/s00244-009-9371-2. PMid:19672647.
http://dx.doi.org/10.1007/s00244-009-937...
; Hayes, 2005Hayes TB. Welcome to the revolution: Integrative biology and assessing the impact of endocrine disruptors on environmental and public health. Integr Comp Biol. 2005;45(2):321-9. http://dx.doi.org/10.1093/icb/45.2.321. PMid:21676775.
http://dx.doi.org/10.1093/icb/45.2.321...
; Jin et al., 2010Jin Y, Zhang X, Shu L, Chen L, Sun L, Qian H, Liu W, Fu Z. Oxidative stress response and gene expression with atrazine exposure in adult female zebrafish (Danio rerio). Chemosphere. 2010;78(7):846-52. http://dx.doi.org/10.1016/j.chemosphere.2009.11.044. PMid:20036412.
http://dx.doi.org/10.1016/j.chemosphere....
). In amphibians, ATZ has a demasculinizing/feminizing effect (Hayes, 2005Hayes TB. Welcome to the revolution: Integrative biology and assessing the impact of endocrine disruptors on environmental and public health. Integr Comp Biol. 2005;45(2):321-9. http://dx.doi.org/10.1093/icb/45.2.321. PMid:21676775.
http://dx.doi.org/10.1093/icb/45.2.321...
). Studies in rats have shown that ATZ elicits depletion of the antioxidant defense system in the testes, which in turn leads to oxidative stress (Abarikwu et al., 2010Abarikwu SO, Adesiyan AC, Oyeloja TO, Oyeyemi MO, Farombi EO. Changes in sperm characteristics and induction of oxidative stress in the testis and epididymis of experimental rats by a herbicide, atrazine. Arch Environ Contam Toxicol. 2010;58(3):874-82. http://dx.doi.org/10.1007/s00244-009-9371-2. PMid:19672647.
http://dx.doi.org/10.1007/s00244-009-937...
; Jin et al., 2010Jin Y, Zhang X, Shu L, Chen L, Sun L, Qian H, Liu W, Fu Z. Oxidative stress response and gene expression with atrazine exposure in adult female zebrafish (Danio rerio). Chemosphere. 2010;78(7):846-52. http://dx.doi.org/10.1016/j.chemosphere.2009.11.044. PMid:20036412.
http://dx.doi.org/10.1016/j.chemosphere....
).
Gely-Pernot et al. (2015)Gely-Pernot A, Hao C, Becker E, Stuparevic I, Kervarrec C, Chalmel F, Primig M, Jégou B, Smagulova F. The epigenetic processes of meiosis in male mice are broadly affected by the widely used herbicide atrazine. BMC Genomics. 2015;16(1):885. http://dx.doi.org/10.1186/s12864-015-2095-y. PMid:26518232.
http://dx.doi.org/10.1186/s12864-015-209...
reported that ATZ interferes with the process of meiosis in male mice. Interruption of meiosis, a key step in gametogenesis, may lead to the production of abnormal spermatozoa and reduced sperm quality, as found in ATZ-treated animals (Abarikwu et al., 2015Abarikwu SO, Duru QC, Chinonso OV, Njoku R-C. Antioxidant enzymes activity, lipid peroxidation, oxidative damage in the testis and epididymis, and steroidogenesis in rats after co-exposure to atrazine and ethanol. Andrologia. 2015;48(5):548-57. http://dx.doi.org/10.1111/and.12478. PMid:26364937.
http://dx.doi.org/10.1111/and.12478...
; Feyzi-Dehkhargani et al., 2012Feyzi-Dehkhargani S, Shahrooz R, Malekinejad H, Sadrkhanloo R-A. Atrazine in sub-acute exposure results in sperm DNA disintegrity and nuclear immaturity in rats. Vet Res Forum an Int Q J 2012;3:19-26.). Supplementation of male goats with 15 mg/kg BW ATZ daily for 6 months impaired morphology, viability, ΔΨm, and cell lipid composition of ejaculated and epididymal spermatozoa (Komsky-Elbaz et al., 2019aKomsky-Elbaz A, Saktsier M, Biran D, Argov-Argaman N, Azaizeh H, Landau YS, Roth Z. Atrazine-induced toxicity in goat spermatozoa is alleviated to some extent by polyphenol-enriched feed. Chemosphere. 2019a;236:124858. http://dx.doi.org/10.1016/j.chemosphere.2019.124858. PMid:31549674.
http://dx.doi.org/10.1016/j.chemosphere....
). Daily administration of 25 mg/kg BW ATZ to mice impaired the production of spermatozoa by interfering with meiosis in the testes (Gely-Pernot et al., 2015Gely-Pernot A, Hao C, Becker E, Stuparevic I, Kervarrec C, Chalmel F, Primig M, Jégou B, Smagulova F. The epigenetic processes of meiosis in male mice are broadly affected by the widely used herbicide atrazine. BMC Genomics. 2015;16(1):885. http://dx.doi.org/10.1186/s12864-015-2095-y. PMid:26518232.
http://dx.doi.org/10.1186/s12864-015-209...
). In rats, daily administration of 120 mg/kg BW ATZ depleted the antioxidant defense system (Abarikwu et al., 2010Abarikwu SO, Adesiyan AC, Oyeloja TO, Oyeyemi MO, Farombi EO. Changes in sperm characteristics and induction of oxidative stress in the testis and epididymis of experimental rats by a herbicide, atrazine. Arch Environ Contam Toxicol. 2010;58(3):874-82. http://dx.doi.org/10.1007/s00244-009-9371-2. PMid:19672647.
http://dx.doi.org/10.1007/s00244-009-937...
) and induced oxidative stress (Abdel Aziz et al., 2018Abdel Aziz RL, Abdel-Wahab A, Abo El-Ela FI, Hassan NE-HY, El-Nahass E-S, Ibrahim MA, Khalil AAY. Dose- dependent ameliorative effects of quercetin and l-Carnitine against atrazine - induced reproductive toxicity in adult male Albino rats. Biomed Pharmacother. 2018;102:855-64. http://dx.doi.org/10.1016/j.biopha.2018.03.136. PMid:29710542.
http://dx.doi.org/10.1016/j.biopha.2018....
) in the testes.
ATZ has been shown to decrease viability and increase the occurrence of pseudo-AR in bovine (Figure 2B; Komsky-Elbaz and Roth, 2017Komsky-Elbaz A, Roth Z. Effect of the herbicide atrazine and its metabolite DACT on bovine sperm quality. Reprod Toxicol. 2017;67:15-25. http://dx.doi.org/10.1016/j.reprotox.2016.11.001. PMid:27836535.
http://dx.doi.org/10.1016/j.reprotox.201...
) and boar (Maravilla-Galván et al., 2009Maravilla-Galván R, Fierro R, González-Márquez H, Gómez-Arroyo S, Jiménez I, Betancourt M. Effects of atrazine and fenoxaprop-ethyl on capacitation and the acrosomal reaction in boar sperm. Int J Toxicol. 2009;28(1):24-32. http://dx.doi.org/10.1177/1091581809333138. PMid:19482828.
http://dx.doi.org/10.1177/10915818093331...
) spermatozoa. In this context, daily administration of 15 mg/kg BW ATZ to male goats for 6 months impaired the ejaculated spermatozoa’s membrane composition, expressed as an increased proportion of saturated fatty acids and reduced proportion of polyunsaturated fatty acids, which were associated with spermatozoon quality (Komsky-Elbaz et al., 2019aKomsky-Elbaz A, Saktsier M, Biran D, Argov-Argaman N, Azaizeh H, Landau YS, Roth Z. Atrazine-induced toxicity in goat spermatozoa is alleviated to some extent by polyphenol-enriched feed. Chemosphere. 2019a;236:124858. http://dx.doi.org/10.1016/j.chemosphere.2019.124858. PMid:31549674.
http://dx.doi.org/10.1016/j.chemosphere....
). Mitochondrial membrane hyperpolarization was documented in ejaculated bovine spermatozoa after 4 h exposure to ATZ or DACT (Figure 2C; Komsky-Elbaz and Roth, 2017Komsky-Elbaz A, Roth Z. Effect of the herbicide atrazine and its metabolite DACT on bovine sperm quality. Reprod Toxicol. 2017;67:15-25. http://dx.doi.org/10.1016/j.reprotox.2016.11.001. PMid:27836535.
http://dx.doi.org/10.1016/j.reprotox.201...
). InDrosophila spermatozoa, ATZ has been shown to affect mitochondrial electron transport (Hase et al., 2008Hase Y, Tatsuno M, Nishi T, Kataoka K, Kabe Y, Yamaguchi Y, Ozawa N, Natori M, Handa H, Watanabe H. Atrazine binds to F1F0-ATP synthase and inhibits mitochondrial function in sperm. Biochem Biophys Res Commun. 2008;366(1):66-72. http://dx.doi.org/10.1016/j.bbrc.2007.11.107. PMid:18060860.
http://dx.doi.org/10.1016/j.bbrc.2007.11...
). In addition, ATZ is suggested to inhibit mitochondrial function in human spermatozoa by binding to F1F0–ATP synthase (Hase et al., 2008Hase Y, Tatsuno M, Nishi T, Kataoka K, Kabe Y, Yamaguchi Y, Ozawa N, Natori M, Handa H, Watanabe H. Atrazine binds to F1F0-ATP synthase and inhibits mitochondrial function in sperm. Biochem Biophys Res Commun. 2008;366(1):66-72. http://dx.doi.org/10.1016/j.bbrc.2007.11.107. PMid:18060860.
http://dx.doi.org/10.1016/j.bbrc.2007.11...
). In terms of practical aspects, i.e., artificial insemination, exposure to ATZ (0.1–1 µM) or its metabolite, DACT (1–10 µM), reduced spermatozoa's cryotolerance and survival post-thawing, by interfering with membrane integrity (Komsky-Elbaz et al., 2019bKomsky-Elbaz A, Zubov A, Roth Z. Effect of the herbicide atrazine and its major metabolite, DACT, on bovine sperm cryotolerance. Theriogenology. 2019b;140:117-23. http://dx.doi.org/10.1016/j.theriogenology.2019.08.026. PMid:31473494.
http://dx.doi.org/10.1016/j.theriogenolo...
).
Epigenetic interference with spermatogenesis was documented in male mice (Gely-Pernot et al., 2015Gely-Pernot A, Hao C, Becker E, Stuparevic I, Kervarrec C, Chalmel F, Primig M, Jégou B, Smagulova F. The epigenetic processes of meiosis in male mice are broadly affected by the widely used herbicide atrazine. BMC Genomics. 2015;16(1):885. http://dx.doi.org/10.1186/s12864-015-2095-y. PMid:26518232.
http://dx.doi.org/10.1186/s12864-015-209...
). Exposure of rat to 25 mg/kg BW/day ATZ was shown to promote epigenetic transgenerational inheritance, expressed by sperm differential DNA methylation regions over three generations (McBirney et al., 2017McBirney M, King SE, Pappalardo M, Houser E, Unkefer M, Nilsson E, Sadler-Riggleman I, Beck D, Winchester P, Skinner MK. Atrazine induced epigenetic transgenerational inheritance of disease, lean phenotype and sperm epimutation pathology biomarkers. PLoS One. 2017;12(9):e0184306. http://dx.doi.org/10.1371/journal.pone.0184306. PMid:28931070.
http://dx.doi.org/10.1371/journal.pone.0...
). Significant transgenerational differences in expression of genes involved in DNA methylation were reported in Medaka fish exposed to 5 or 50 μg/L ATZ (Cleary et al., 2019Cleary JA, Tillitt DE, vom Saal FS, Nicks DK, Claunch RA, Bhandari RK. Atrazine induced transgenerational reproductive effects in medaka (Oryzias latipes). Environ Pollut. 2019;251:639-50. http://dx.doi.org/10.1016/j.envpol.2019.05.013. PMid:31108297.
http://dx.doi.org/10.1016/j.envpol.2019....
). Zebrafish larvae exposure to 3 ppb ATZ through embryogenesis lead to decreased expression of DNA methyltransferase and decreased DNA methylation levels (Wirbisky-Hershberger et al., 2017Wirbisky-Hershberger SE, Sanchez OF, Horzmann KA, Thanki D, Yuan C, Freeman JL. Atrazine exposure decreases the activity of DNMTs, global DNA methylation levels, and dnmt expression. Food Chem Toxicol. 2017;109(Pt 1):727-34. http://dx.doi.org/10.1016/j.fct.2017.08.041. PMid:28859886.
http://dx.doi.org/10.1016/j.fct.2017.08....
). Evaluation of the proteome of spermatozoa exposed to ATZ or DACT revealed differential expression of multiple proteins related to several cellular pathways and biological processes, associated with genetic and epigenetic processes (Komsky-Elbaz et al., unpublished data).
Carryover effect of foodborne contaminants from the sperm to the embryo
During fertilization, the spermatozoon delivers paternal components to the oocyte which are crucial for oocyte activation, zygote formation, and further embryonic development (Dogan et al., 2015Dogan S, Vargovic P, Oliveira R, Belser LE, Kaya A, Moura A, Sutovsky P, Parrish J, Topper E, Memili E. Sperm protamine-status correlates to the fertility of breeding bulls. Biol Reprod. 2015;92(4):92. http://dx.doi.org/10.1095/biolreprod.114.124255. PMid:25673563.
http://dx.doi.org/10.1095/biolreprod.114...
). Thus, paternal exposure to environmental contaminants plays a critical role in the offspring’s health (Siddeek et al., 2018Siddeek B, Mauduit C, Simeoni U, Benahmed M. Sperm epigenome as a marker of environmental exposure and lifestyle, at the origin of diseases inheritance. Mutat Res Mutat Res. 2018;778:38-44. http://dx.doi.org/10.1016/j.mrrev.2018.09.001. PMid:30454681.
http://dx.doi.org/10.1016/j.mrrev.2018.0...
). In-vitro fertilization of rat oocytes with epididymal sperm capacitated with AFB1 (2–16 ppb) resulted in significantly lower fertilization rates relative to fertilization with untreated sperm (Ibeh et al., 2000Ibeh IN, Saxena DK, Uraih N. Toxicity of aflatoxin: Effects on spermatozoa, oocytes, and in vitro fertilization. J Environ Pathol Toxicol Oncol. 2000;19(4):357-61. PMid:11213017.). In bovine, the proportion of oocytes that were fertilized and cleaved to 2- to 4-cell-stage embryos was lower after fertilization with spermatozoa exposed to 10 µM AFB1 compared to non-treated spermatozoa, but blastocyst-formation rate was relatively fair (Komsky-Elbaz et al., 2018Komsky-Elbaz A, Saktsier M, Roth Z. Aflatoxin B1 impairs sperm quality and fertilization competence. Toxicology. 2018;393:42-50. http://dx.doi.org/10.1016/j.tox.2017.11.007. PMid:29113834.
http://dx.doi.org/10.1016/j.tox.2017.11....
). One possible explanation is that although the spermatozoa were exposed to AFB1 through capacitation, fertilization was performed by those that had not been impaired.
The spermatozoa's contribution to the embryo is not limited to the paternal DNA; it also includes epigenetic marks on the DNA, as well as RNA and proteins that are transferred through fertilization (Castillo et al., 2018Castillo J, Jodar M, Oliva R. The contribution of human sperm proteins to the development and epigenome of the preimplantation embryo. Hum Reprod Update. 2018;24(5):535-55. http://dx.doi.org/10.1093/humupd/dmy017. PMid:29800303.
http://dx.doi.org/10.1093/humupd/dmy017...
). About 108 sperm-originated proteins have been identified in the human blastocyst (Castillo et al., 2018Castillo J, Jodar M, Oliva R. The contribution of human sperm proteins to the development and epigenome of the preimplantation embryo. Hum Reprod Update. 2018;24(5):535-55. http://dx.doi.org/10.1093/humupd/dmy017. PMid:29800303.
http://dx.doi.org/10.1093/humupd/dmy017...
), and these might be crucial for embryonic development and health (Oliva et al., 2015Oliva R, Castillo J, Estanyol J, Ballescà J. Human sperm chromatin epigenetic potential: genomics, proteomics, and male infertility. Asian J Androl. 2015;17(4):601-609. http://dx.doi.org/10.4103/1008-682X.153302.
http://dx.doi.org/10.4103/1008-682X.1533...
; Rando, 2012Rando OJ. Daddy issues: paternal effects on phenotype. Cell. 2012;151(4):702-8. http://dx.doi.org/10.1016/j.cell.2012.10.020. PMid:23141533.
http://dx.doi.org/10.1016/j.cell.2012.10...
). It is also possible that the reduced cleavage rate reported for AFB1-treated spermatozoa expresses a delay in embryonic development rather than fertilization failure. In support of this assumption, it has been recently reported that sperm with DNA damage can fertilize the oocyte (Bungum et al., 2004Bungum M, Humaidan P, Spano M, Jepson K, Bungum L, Giwercman A. The predictive value of sperm chromatin structure assay (SCSA) parameters for the outcome of intrauterine insemination, IVF and ICSI. Hum Reprod. 2004;19:1401-8. http://dx.doi.org/10.1093/humrep/deh280.
http://dx.doi.org/10.1093/humrep/deh280...
). Moreover, soon after fertilization, a repair process begins in the oocyte (Uppangala et al., 2016Uppangala S, Pudakalakatti S, D’souza F, Salian SR, Kalthur G, Kumar P, Atreya H, Adiga SK. Influence of sperm DNA damage on human preimplantation embryo metabolism. Reprod Biol. 2016;16(3):234-41. http://dx.doi.org/10.1016/j.repbio.2016.07.004. PMid:27492188.
http://dx.doi.org/10.1016/j.repbio.2016....
), suggesting that the embryo can repair DNA damage of sperm origin. On the other hand, it has been reported that a high proportion of sperm with DNA damage is associated with altered metabolism in the developing blastocyst (Uppangala et al., 2016Uppangala S, Pudakalakatti S, D’souza F, Salian SR, Kalthur G, Kumar P, Atreya H, Adiga SK. Influence of sperm DNA damage on human preimplantation embryo metabolism. Reprod Biol. 2016;16(3):234-41. http://dx.doi.org/10.1016/j.repbio.2016.07.004. PMid:27492188.
http://dx.doi.org/10.1016/j.repbio.2016....
). These findings suggest that damaged DNA not only reduces sperm fertilization competence, but also negatively affects embryonic development (Ioannou et al., 2016Ioannou D, Miller D, Griffin DK, Tempest HG. Impact of sperm DNA chromatin in the clinic. J Assist Reprod Genet. 2016;33(2):157-66. http://dx.doi.org/10.1007/s10815-015-0624-x. PMid:26678492.
http://dx.doi.org/10.1007/s10815-015-062...
) and increases the number of embryonic cells undergoing apoptosis (Simões et al., 2013Simões R, Feitosa WB, Siqueira AFP, Nichi M, Paula-Lopes FF, Marques MG, Peres MA, Barnabe VH, Visintin JA, Assumpção ME. Influence of bovine sperm DNA fragmentation and oxidative stress on early embryo in vitro development outcome. Reproduction. 2013;146(5):433-41. http://dx.doi.org/10.1530/REP-13-0123. PMid:23940385.
http://dx.doi.org/10.1530/REP-13-0123...
). Exposure of spermatozoa of the marine invertebrate Galeolaria caespitosa to dibutyl phthalate inhibited fertilization and embryogenesis (Lu et al., 2017Lu Y, Lin M, Aitken RJ. Exposure of spermatozoa to dibutyl phthalate induces abnormal embryonic development in a marine invertebrate Galeolaria caespitosa (Polychaeta: serpulidae). Aquat Toxicol. 2017;191:189-200. http://dx.doi.org/10.1016/j.aquatox.2017.08.008. PMid:28843738.
http://dx.doi.org/10.1016/j.aquatox.2017...
). Although sperm with damaged DNA can fertilize the oocyte, genomic instability in the developed embryo cannot be ruled out (Adiga et al., 2010Adiga SK, Upadhya D, Kalthur G, Bola Sadashiva SR, Kumar P. Transgenerational changes in somatic and germ line genetic integrity of first-generation offspring derived from the DNA damaged sperm. Fertil Steril. 2010;93(8):2486-90. http://dx.doi.org/10.1016/j.fertnstert.2009.06.015. PMid:19591998.
http://dx.doi.org/10.1016/j.fertnstert.2...
). Evaluating the transcriptome profiles of blastocysts derived following in-vitro fertilization with AFB1-treated spermatozoa revealed a significantly different profile from that of the control, suggesting a carryover effect of AFB1 from the spermatozoa to the developing embryo (Komsky-Elbaz and Roth, unpublished data).
Traditional studies claim that inheritance from one generation to the next occurs solely through genetic information. However, there is increasing evidence of an important role for epigenetic factors in the transmission of certain phenotypes from parents to their offspring. Environmentally induced epigenetic alterations can be inherited through the male gametes (Wei et al., 2015Wei Y, Schatten H, Sun Q-Y. Environmental epigenetic inheritance through gametes and implications for human reproduction. Hum Reprod Update. 2015;21(2):194-208. http://dx.doi.org/10.1093/humupd/dmu061. PMid:25416302.
http://dx.doi.org/10.1093/humupd/dmu061...
) and might regulate gene expression at transcriptional as well as post-transcriptional levels in the early developing embryo (Castillo et al., 2018Castillo J, Jodar M, Oliva R. The contribution of human sperm proteins to the development and epigenome of the preimplantation embryo. Hum Reprod Update. 2018;24(5):535-55. http://dx.doi.org/10.1093/humupd/dmy017. PMid:29800303.
http://dx.doi.org/10.1093/humupd/dmy017...
). For instance, paternal exposure to EDCs may cause alterations in the spermatozoa's RNA, and some of these changes might persist in the offspring (Uppangala et al., 2016Uppangala S, Pudakalakatti S, D’souza F, Salian SR, Kalthur G, Kumar P, Atreya H, Adiga SK. Influence of sperm DNA damage on human preimplantation embryo metabolism. Reprod Biol. 2016;16(3):234-41. http://dx.doi.org/10.1016/j.repbio.2016.07.004. PMid:27492188.
http://dx.doi.org/10.1016/j.repbio.2016....
). In mice, paternal prediabetes altered overall methylome patterns in spermatozoa, with a large portion of differentially methylated genes overlapping with genes of pancreatic islets in the offspring (Wei et al., 2014Wei Y, Yang CR, Wei YP, Zhao ZA, Hou Y, Schatten H, Sun QY. Paternally induced transgenerational inheritance of susceptibility to diabetes in mammals. Proc Natl Acad Sci USA. 2014;111(5):1873-8. http://dx.doi.org/10.1073/pnas.1321195111. PMid:24449870.
http://dx.doi.org/10.1073/pnas.132119511...
). Thus, it is highly possible that alterations in the transcriptomic profile of embryos derived after fertilization with AFB1-treated spermatozoa will include not only genetic but also epigenetic alterations. This point should be further evaluated.
Summary
There is a wide range of evidence for the potential hazard associated with exposure of spermatozoa to foodborne contaminants. The damage is already initiated in the testes during spermatogenesis, persists during the sperm's passage through the epididymis and finally, is expressed in the mature spermatozoa. Exposure of the spermatozoa to contaminants may also occur after ejaculation, in the female reproductive tract. The damage seems to be multifactorial in nature, involving alterations in the spermatozoa’s membranes, DNA and proteome. Interestingly, different contaminants do not necessarily have the same effect on spermatozoa, suggesting different modes of action. Evidence of the damage in the spermatozoa carrying over to the developing embryo is of great concern, and warrants further investigation.
General Summary
This review provides a broad screening of the information on the effects of EDCs and foodborne contaminants on both the oocyte and spermatozoa (Figure 3). When the oocyte is exposed to MEHP, deleterious effects are further recorded in the developed embryos. Similarly, exposing spermatozoa to foodborne contaminants further affects the formed embryos. While this information is directly related to the presented environmental contaminants, these data might also be relevant to other EDCs. Exploring the risk associated with exposing animals to environmental contaminants might lead to better management for reproductive health preservation.

Schematic illustration of the effects of EDCs and foodborne contaminants on female and male gametes. ATZ and AFB1 induce alterations in the spermatozoa. These include cellular and molecular alterations, which in turn impair fertilization competence. MEHP induces cellular and molecular alterations that affect oocyte developmental competence. Alterations in both male and female gametes are further expressed in the developed blastocyst.
-
Funding disclosure statement: ZR received funding for this research from the Environmental Health fund (EHF), Jerusalem, Israel.
-
How to cite: Roth Z, Komsky-Elbaz A, Kalo D. Effect of environmental contamination on female and male gametes – A lesson from bovines. Anim Reprod. 2020;17(3):e20200041. https://doi.org/10.1590/1984-3143-AR2020-0041
References
- Abarikwu SO, Adesiyan AC, Oyeloja TO, Oyeyemi MO, Farombi EO. Changes in sperm characteristics and induction of oxidative stress in the testis and epididymis of experimental rats by a herbicide, atrazine. Arch Environ Contam Toxicol. 2010;58(3):874-82. http://dx.doi.org/10.1007/s00244-009-9371-2 PMid:19672647.
» http://dx.doi.org/10.1007/s00244-009-9371-2 - Abarikwu SO, Duru QC, Chinonso OV, Njoku R-C. Antioxidant enzymes activity, lipid peroxidation, oxidative damage in the testis and epididymis, and steroidogenesis in rats after co-exposure to atrazine and ethanol. Andrologia. 2015;48(5):548-57. http://dx.doi.org/10.1111/and.12478 PMid:26364937.
» http://dx.doi.org/10.1111/and.12478 - Abdel Aziz RL, Abdel-Wahab A, Abo El-Ela FI, Hassan NE-HY, El-Nahass E-S, Ibrahim MA, Khalil AAY. Dose- dependent ameliorative effects of quercetin and l-Carnitine against atrazine - induced reproductive toxicity in adult male Albino rats. Biomed Pharmacother. 2018;102:855-64. http://dx.doi.org/10.1016/j.biopha.2018.03.136 PMid:29710542.
» http://dx.doi.org/10.1016/j.biopha.2018.03.136 - Absalan F, Saremy S, Mansori E, Taheri Moghadam M, Eftekhari Moghadam AR, Ghanavati R. Effects of Mono-(2-Ethylhexyl) Phthalate and Di-(2-Ethylhexyl) Phthalate administrations on Oocyte Meiotic Maturation, Apoptosis and Gene Quantification in mouse model. Cell J. 2017;18(4):503-13. PMid:28042535.
- Acton BM, Jurisicova A, Jurisica I, Casper RF. Alterations in mitochondrial membrane potential during preimplantation stages of mouse and human embryo development. Mol Hum Reprod. 2004;10(1):23-32. http://dx.doi.org/10.1093/molehr/gah004 PMid:14665703.
» http://dx.doi.org/10.1093/molehr/gah004 - Adibi JJ, Hauser R, Williams PL, Whyatt RM, Calafat AM, Nelson H, Herrick R, Swan SH. Maternal urinary metabolites of Di-(2-Ethylhexyl) phthalate in relation to the timing of labor in a US multicenter pregnancy cohort study. Am J Epidemiol. 2009;169(8):1015-24. http://dx.doi.org/10.1093/aje/kwp001 PMid:19251754.
» http://dx.doi.org/10.1093/aje/kwp001 - Adiga SK, Upadhya D, Kalthur G, Bola Sadashiva SR, Kumar P. Transgenerational changes in somatic and germ line genetic integrity of first-generation offspring derived from the DNA damaged sperm. Fertil Steril. 2010;93(8):2486-90. http://dx.doi.org/10.1016/j.fertnstert.2009.06.015 PMid:19591998.
» http://dx.doi.org/10.1016/j.fertnstert.2009.06.015 - Agnes VF, Akbarsha MA. Spermatotoxic effect of aflatoxin B(1) in the albino mouse. Food Chem Toxicol. 2003;41(1):119-30. http://dx.doi.org/10.1016/S0278-6915(02)00171-0 PMid:12453736.
» http://dx.doi.org/10.1016/S0278-6915(02)00171-0 - Aldyreva MV, Klimova TS, Iziumova AS, Timofeevskaia LA. The effect of phthalate plasticizers on the generative function. Gig Tr Prof Zabol. 1975;(12):25-9. PMid:1213487.
- Ambruosi B, Uranio M, Sardanelli A, Pocar P, Martino N, Parternoster M, Amati F, Dell'Aquila ME. In Vitro acute exposure to DEHP affects Oocyte Meiotic maturation, energy and oxidative stress parameters in a large animal model. PLoS One. 2011;6(11):e27452. http://dx.doi.org/10.1371/journal.pone.0027452 PMid:22076161.
» http://dx.doi.org/10.1371/journal.pone.0027452 - Anas M-KI, Suzuki C, Yoshioka K, Iwamura S. Effect of mono-(2-ethylhexyl) phthalate on bovine oocyte maturation in vitro. Reprod Toxicol. 2003;17(3):305-10. http://dx.doi.org/10.1016/S0890-6238(03)00014-5 PMid:12759099.
» http://dx.doi.org/10.1016/S0890-6238(03)00014-5 - Arbuckle TE, Fisher M, MacPherson S, Lang C, Provencher G, LeBlanc A, Hauser R, Feeley M, Ayotte P, Neisa A, Ramsay T, Tawagi G. Maternal and early life exposure to phthalates: The plastics and personal-care Products use in Pregnancy (P4) study. Sci Total Environ. 2016;551-552:344-56. http://dx.doi.org/10.1016/j.scitotenv.2016.02.022 PMid:26878646.
» http://dx.doi.org/10.1016/j.scitotenv.2016.02.022 - Ataman MB, Bucak MN, Coyan K. Esterified glucomannan improves aflatoxin-induced damage of sperm parameters during liquid storage of ram semen at 5°C. Cryobiology. 2014;68(3):405-10. http://dx.doi.org/10.1016/j.cryobiol.2014.03.007 PMid:24662029.
» http://dx.doi.org/10.1016/j.cryobiol.2014.03.007 - ATSDR. Toxicological profile for Di(2-ethylhexyl)phthalate (DEHP). Atlanta, GA: U.S. Department of Health and Human Services, Public Health Service; 2002.
- Austin KJ, Cockrum RR, Jons AM, Alexander BM, Cammack KM. Renin mRNA is upregulated in testes and testicular cells in response to treatment with aflatoxin B1. Theriogenology. 2012;77:331-7.e1-7. http://dx.doi.org/10.1016/j.theriogenology.2011.08.004
» http://dx.doi.org/10.1016/j.theriogenology.2011.08.004 - Balabanič D, Rupnik M, Klemenčič AK. Negative impact of endocrine-disrupting compounds on human reproductive health. Reprod Fertil Dev. 2011;23(3):403-16. http://dx.doi.org/10.1071/RD09300 PMid:21426858.
» http://dx.doi.org/10.1071/RD09300 - Barr DB, Panuwet P, Nguyen JV, Udunka S, Needham LL. Assessing exposure to atrazine and its metabolites using biomonitoring. Environ Health Perspect. 2007;115(10):1474-8. http://dx.doi.org/10.1289/ehp.10141 PMid:17938738.
» http://dx.doi.org/10.1289/ehp.10141 - Bavister BD, Squirrell JM. Mitochondrial distribution and function in oocytes and early embryos. Hum Reprod. 2000;15(Suppl 2):189-98. http://dx.doi.org/10.1093/humrep/15.suppl_2.189 PMid:11041524.
» http://dx.doi.org/10.1093/humrep/15.suppl_2.189 - Becker-Algeri TA, Castagnaro D, Bortoli K, Souza C, Drunkler DA, Badiale-Furlong E. Mycotoxins in bovine milk and dairy products: A review. J Food Sci. 2016;81(3):R544-52. http://dx.doi.org/10.1111/1750-3841.13204 PMid:26799355.
» http://dx.doi.org/10.1111/1750-3841.13204 - Beker van Woudenberg A, Gröllers-Mulderij M, Snel C, Jeurissen N, Stierum R, Wolterbeek A. The bovine oocyte in vitro maturation model: A potential tool for reproductive toxicology screening. Reprod Toxicol. 2012;34(2):251-60. http://dx.doi.org/10.1016/j.reprotox.2012.05.098 PMid:22664270.
» http://dx.doi.org/10.1016/j.reprotox.2012.05.098 - Benjamin S, Masai E, Kamimura N, Takahashi K, Anderson RC, Faisal PA. Phthalates impact human health: Epidemiological evidences and plausible mechanism of action. J Hazard Mater. 2017;340:360-83. http://dx.doi.org/10.1016/j.jhazmat.2017.06.036 PMid:28800814.
» http://dx.doi.org/10.1016/j.jhazmat.2017.06.036 - Bergé A, Cladière M, Gasperi J, Coursimault A, Tassin B, Moilleron R. Meta-analysis of environmental contamination by phthalates. Environ Sci Pollut Res Int. 2013;20(11):8057-76. http://dx.doi.org/10.1007/s11356-013-1982-5 PMid:23917738.
» http://dx.doi.org/10.1007/s11356-013-1982-5 - Betancourt M, Reséndiz A, Fierro ECYR. Effect of two insecticides and two herbicides on the porcine sperm motility patterns using computer-assisted semen analysis (CASA) in vitro. Reprod Toxicol. 2006;22(3):508-12. http://dx.doi.org/10.1016/j.reprotox.2006.03.001 PMid:16713176.
» http://dx.doi.org/10.1016/j.reprotox.2006.03.001 - Bhattacharya P, Keating AF. Impact of environmental exposures on ovarian function and role of xenobiotic metabolism during ovotoxicity. Toxicol Appl Pharmacol. 2012;261(3):227-35. http://dx.doi.org/10.1016/j.taap.2012.04.009 PMid:22531813.
» http://dx.doi.org/10.1016/j.taap.2012.04.009 - Bian Q, Xu LC, Wang SL, Xia YK, Tan LF, Chen JF, Song L, Chang HC, Wang XR. Study on the relation between occupational fenvalerate exposure and spermatozoa DNA damage of pesticide factory workers. Occup Environ Med. 2004;61(12):999-1005. http://dx.doi.org/10.1136/oem.2004.014597 PMid:15550606.
» http://dx.doi.org/10.1136/oem.2004.014597 - Bonilla E, del Mazo J. Deregulation of the Sod1 and Nd1 genes in mouse fetal oocytes exposed to mono-(2-ethylhexyl) phthalate (MEHP). Reprod Toxicol. 2010;30(3):387-92. http://dx.doi.org/10.1016/j.reprotox.2010.04.008 PMid:20438828.
» http://dx.doi.org/10.1016/j.reprotox.2010.04.008 - Bradman A, Barr DB, Claus Henn BG, Drumheller T, Curry C, Eskenazi B. Measurement of pesticides and other toxicants in amniotic fluid as a potential biomarker of prenatal exposure: A validation study. Environ Health Perspect. 2003;111(14):1779-82. http://dx.doi.org/10.1289/ehp.6259 PMid:14594631.
» http://dx.doi.org/10.1289/ehp.6259 - Bungum M, Humaidan P, Spano M, Jepson K, Bungum L, Giwercman A. The predictive value of sperm chromatin structure assay (SCSA) parameters for the outcome of intrauterine insemination, IVF and ICSI. Hum Reprod. 2004;19:1401-8. http://dx.doi.org/10.1093/humrep/deh280
» http://dx.doi.org/10.1093/humrep/deh280 - Castillo J, Jodar M, Oliva R. The contribution of human sperm proteins to the development and epigenome of the preimplantation embryo. Hum Reprod Update. 2018;24(5):535-55. http://dx.doi.org/10.1093/humupd/dmy017 PMid:29800303.
» http://dx.doi.org/10.1093/humupd/dmy017 - Chevrier C, Limon G, Monfort C, Rouget F, Garlantézec R, Petit C, Durand G, Cordier S. Urinary biomarkers of prenatal atrazine exposure and adverse birth outcomes in the PELAGIE birth cohort. Environ Health Perspect. 2011;119(7):1034-41. http://dx.doi.org/10.1289/ehp.1002775 PMid:21367690.
» http://dx.doi.org/10.1289/ehp.1002775 - Cleary JA, Tillitt DE, vom Saal FS, Nicks DK, Claunch RA, Bhandari RK. Atrazine induced transgenerational reproductive effects in medaka (Oryzias latipes). Environ Pollut. 2019;251:639-50. http://dx.doi.org/10.1016/j.envpol.2019.05.013 PMid:31108297.
» http://dx.doi.org/10.1016/j.envpol.2019.05.013 - Cornwall GA. Role of posttranslational protein modifications in epididymal sperm maturation and extracellular quality control. Adv Exp Med Biol. 2014;759:159-80. http://dx.doi.org/10.1007/978-1-4939-0817-2_8 PMid:25030764.
» http://dx.doi.org/10.1007/978-1-4939-0817-2_8 - Craig ZR, Wang W, Flaws JA. Endocrine-disrupting chemicals in ovarian function: effects on steroidogenesis, metabolism and nuclear receptor signaling. Reproduction. 2011;142(5):633-46. http://dx.doi.org/10.1530/REP-11-0136 PMid:21862696.
» http://dx.doi.org/10.1530/REP-11-0136 - Dai Y, Huang K, Zhang B, Zhu L, Xu W. Aflatoxin B1-induced epigenetic alterations: an overview. Food Chem Toxicol. 2017;109(Pt 1):683-9. http://dx.doi.org/10.1016/j.fct.2017.06.034 PMid:28645871.
» http://dx.doi.org/10.1016/j.fct.2017.06.034 - Davis BJ, Maronpot RR, Di Heindel JJ. -(2-ethylhexyl) phthalate suppresses estradiol and ovulation in cycling rats. Toxicol Appl Pharmacol. 1994;128(2):216-23. http://dx.doi.org/10.1006/taap.1994.1200 PMid:7940536.
» http://dx.doi.org/10.1006/taap.1994.1200 - Diamanti-Kandarakis E, Bourguignon J-P, Giudice LC, Hauser R, Prins GS, Soto AM, Zoeller RT, Gore AC. Endocrine-disrupting chemicals: an Endocrine Society scientific statement. Endocr Rev. 2009;30(4):293-342. http://dx.doi.org/10.1210/er.2009-0002 PMid:19502515.
» http://dx.doi.org/10.1210/er.2009-0002 - Dogan S, Vargovic P, Oliveira R, Belser LE, Kaya A, Moura A, Sutovsky P, Parrish J, Topper E, Memili E. Sperm protamine-status correlates to the fertility of breeding bulls. Biol Reprod. 2015;92(4):92. http://dx.doi.org/10.1095/biolreprod.114.124255 PMid:25673563.
» http://dx.doi.org/10.1095/biolreprod.114.124255 - Dooley GP, Reardon KF, Prenni JE, Tjalkens RB, Legare ME, Foradori CD, Tessari JE, Hanneman WH. Proteomic analysis of diaminochlorotriazine adducts in wister rat pituitary glands and L T2 rat pituitary cells 2008:21(4):844-51.
- Dooley GP, Tjalkens RB, Hanneman WH. The atrazine metabolite diaminochlorotriazine suppresses LH release from murine LβT2 cells by suppressing GnRH-induced intracellular calcium transients. Toxicol Res (Camb). 2013;2(3):180-6. http://dx.doi.org/10.1039/c3tx20088d PMid:24052811.
» http://dx.doi.org/10.1039/c3tx20088d - Eimani H, Dalman A, Sepehri H, Kazemi S, Hassani F, Rezazadeh M, Baharvand H, Shahverdi A, Samani RO. Effect of DEHP (Di-2-Ethyl Hexyl-phthalate) on resumption of meiosis and in-vitro maturation of mouse oocytes and development of resulting embryos. Yakhth Med J. 2005;7:56-61.
- El Khoury D, Fayjaloun S, Nassar M, Sahakian J, Aad PY. Updates on the effect of mycotoxins on male reproductive efficiency in mammals. Toxins (Basel). 2019;11(9). http://dx.doi.org/10.3390/toxins11090515 PMid:31484408.
» http://dx.doi.org/10.3390/toxins11090515 - Eppig JJ. Coordination of nuclear and cytoplasmic oocyte maturation in eutherian mammals. Reprod Fertil Dev. 1996;8(4):485-9. http://dx.doi.org/10.1071/RD9960485 PMid:8870074.
» http://dx.doi.org/10.1071/RD9960485 - Espinoza JA, Paasch U, Villegas JV. Mitochondrial membrane potential disruption pattern in human sperm. Hum Reprod. 2009;24(9):2079-85. http://dx.doi.org/10.1093/humrep/dep120 PMid:19465458.
» http://dx.doi.org/10.1093/humrep/dep120 - Eze UA, Huntriss J, Routledge MN, Gong YY. Toxicological effects of regulated mycotoxins and persistent organochloride pesticides: in vitro cytotoxic assessment of single and defined mixtures on MA-10 murine Leydig cell line. Toxicol Vitr. 2018;48:93-103. http://dx.doi.org/10.1016/j.tiv.2017.12.019 PMid:29307701.
» http://dx.doi.org/10.1016/j.tiv.2017.12.019 - Faisal K, Faridha A, Akbarsha MA. Induction of meiotic micronuclei in spermatocytes in vivo by aflatoxin B1: light and transmission electron microscopic study in Swiss mouse. Reprod Toxicol. 2008b;26:303-9. http://dx.doi.org/10.1016/j.reprotox.2008.09.003
» http://dx.doi.org/10.1016/j.reprotox.2008.09.003 - Faisal K, Periasamy VS, Sahabudeen S, Radha A, Anandhi R, Akbarsha MA. Spermatotoxic effect of aflatoxin B1 in rat: extrusion of outer dense fibres and associated axonemal microtubule doublets of sperm flagellum. Reproduction. 2008a;135(3):303-10. http://dx.doi.org/10.1530/REP-07-0367 PMid:18299423.
» http://dx.doi.org/10.1530/REP-07-0367 - Fan S, Li Q, Sun L, Du Y, Xia J, Zhang Y. Simultaneous determination of aflatoxin B 1 and M 1 in milk, fresh milk and milk powder by LC-MS/MS utilising online turbulent flow chromatography. Food Addit Contam Part A Chem Anal Control Expo Risk Assess. 2015;32(7):1175-84. http://dx.doi.org/10.1080/19440049.2015.1048311 PMid:25952817.
» http://dx.doi.org/10.1080/19440049.2015.1048311 - Ferreira EM, Vireque AA, Adona PR, Meirelles FV, Ferriani RA, Navarro PAAS. Cytoplasmic maturation of bovine oocytes: structural and biochemical modifications and acquisition of developmental competence. Theriogenology. 2009;71(5):836-48. http://dx.doi.org/10.1016/j.theriogenology.2008.10.023 PMid:19121865.
» http://dx.doi.org/10.1016/j.theriogenology.2008.10.023 - Feyzi-Dehkhargani S, Shahrooz R, Malekinejad H, Sadrkhanloo R-A. Atrazine in sub-acute exposure results in sperm DNA disintegrity and nuclear immaturity in rats. Vet Res Forum an Int Q J 2012;3:19-26.
- Forgacs AL, D’Souza ML, Huhtaniemi IT, Rahman NA, Zacharewski TR. Triazine Herbicides and Their Chlorometabolites Alter Steroidogenesis in BLTK1 Murine Leydig Cells. ToxSci 2013;134:155-67. https://doi.org/10.1093/toxsci/kft096
» https://doi.org/10.1093/toxsci/kft096 - Gely-Pernot A, Hao C, Becker E, Stuparevic I, Kervarrec C, Chalmel F, Primig M, Jégou B, Smagulova F. The epigenetic processes of meiosis in male mice are broadly affected by the widely used herbicide atrazine. BMC Genomics. 2015;16(1):885. http://dx.doi.org/10.1186/s12864-015-2095-y PMid:26518232.
» http://dx.doi.org/10.1186/s12864-015-2095-y - Gilchrist RB, Aktas H, Wheeler MB. Recent insights into oocyte - follicle cell interactions provide opportunities for the development of new approaches to in vitro maturation. Reprod Fertil Dev. 2011;23(1):23-31. http://dx.doi.org/10.1071/RD10225 PMid:21366977.
» http://dx.doi.org/10.1071/RD10225 - Giovati L, Magliani W, Ciociola T, Santinoli C, Conti S, Polonelli L. AFM1 in Milk: Physical, biological, and prophylactic methods to mitigate contamination. Toxins (Basel). 2015;7(10):4330-49. http://dx.doi.org/10.3390/toxins7104330 PMid:26512694.
» http://dx.doi.org/10.3390/toxins7104330 - Gong YY, Watson S, Routledge MN. Aflatoxin exposure and associated human health effects, a review of epidemiological studies. Food Saf. 2016;4(1):14-27. http://dx.doi.org/10.14252/foodsafetyfscj.2015026 PMid:32231900.
» http://dx.doi.org/10.14252/foodsafetyfscj.2015026 - Grossman D, Kalo D, Gendelman M, Roth Z. Effect of di-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl) phthalate on in vitro developmental competence of bovine oocytes. Cell Biol Toxicol. 2012;28(6):383-96. http://dx.doi.org/10.1007/s10565-012-9230-1 PMid:22956148.
» http://dx.doi.org/10.1007/s10565-012-9230-1 - Gupta RK, Singh JM, Leslie TC, Meachum S, Flaws JA, Yao HH. C-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl) phthalate inhibit growth and reduce estradiol levels of antral follicles in vitro. Toxicol Appl Pharmacol. 2010;242(2):224-30. http://dx.doi.org/10.1016/j.taap.2009.10.011 PMid:19874833.
» http://dx.doi.org/10.1016/j.taap.2009.10.011 - Hannon PR, Niermann S, Flaws JA. Acute exposure to Di(2-Ethylhexyl) phthalate in adulthood causes adverse reproductive outcomes later in life and accelerates reproductive aging in female mice. Toxicol Sci. 2016;150(1):97-108. http://dx.doi.org/10.1093/toxsci/kfv317 PMid:26678702.
» http://dx.doi.org/10.1093/toxsci/kfv317 - Hasanzadeh S, Rezazadeh L. Effects of aflatoxin B1 on the growth processes of spermatogenic cell series in adult male rats. Comp Clin Pathol. 2013;22(4):555-62. http://dx.doi.org/10.1007/s00580-012-1445-2
» http://dx.doi.org/10.1007/s00580-012-1445-2 - Hase Y, Tatsuno M, Nishi T, Kataoka K, Kabe Y, Yamaguchi Y, Ozawa N, Natori M, Handa H, Watanabe H. Atrazine binds to F1F0-ATP synthase and inhibits mitochondrial function in sperm. Biochem Biophys Res Commun. 2008;366(1):66-72. http://dx.doi.org/10.1016/j.bbrc.2007.11.107 PMid:18060860.
» http://dx.doi.org/10.1016/j.bbrc.2007.11.107 - Hauser R, Calafat AM. Phthalates and human health. Occup Environ Med. 2005;62(11):806-18. http://dx.doi.org/10.1136/oem.2004.017590 PMid:16234408.
» http://dx.doi.org/10.1136/oem.2004.017590 - Hayes TB. Welcome to the revolution: Integrative biology and assessing the impact of endocrine disruptors on environmental and public health. Integr Comp Biol. 2005;45(2):321-9. http://dx.doi.org/10.1093/icb/45.2.321 PMid:21676775.
» http://dx.doi.org/10.1093/icb/45.2.321 - Heindel JJ, Chapin RE. Inhibition of FSH-stimulated cAMP accumulation by mono(2-ethylhexyl) phthalate in primary rat Sertoli cell cultures. Toxicol Appl Pharmacol. 1989;97(2):377-85. http://dx.doi.org/10.1016/0041-008X(89)90342-6 PMid:2538009.
» http://dx.doi.org/10.1016/0041-008X(89)90342-6 - Heudorf U, Mersch-Sundermann V, Angerer J. Phthalates: toxicology and exposure. Int J Hyg Environ Health. 2007;210(5):623-34. http://dx.doi.org/10.1016/j.ijheh.2007.07.011 PMid:17889607.
» http://dx.doi.org/10.1016/j.ijheh.2007.07.011 - Hongjun Y, Wenjun X, Qing L, Jingtao L, Hongwen Y, Zhaohua L. Distribution of phthalate esters in topsoil: a case study in the Yellow River Delta, China. Environ Monit Assess. 2013;185(10):8489-500. http://dx.doi.org/10.1007/s10661-013-3190-7 PMid:23609921.
» http://dx.doi.org/10.1007/s10661-013-3190-7 - Huang Y, Li J, Garcia JM, Lin H, Wang Y, Yan P, Wang L, Tan Y, Luo J, Qiu Z, Chen JA, Shu W. Phthalate levels in cord blood are associated with preterm delivery and fetal growth parameters in chinese women. PLoS One. 2014;9(2):e87430. http://dx.doi.org/10.1371/journal.pone.0087430 PMid:24503621.
» http://dx.doi.org/10.1371/journal.pone.0087430 - Hüttemann M, Lee I, Pecinova A, Pecina P, Przyklenk K, Doan JW. Regulation of oxidative phosphorylation, the mitochondrial membrane potential, and their role in human disease. J Bioenerg Biomembr. 2008;40(5):445-56. http://dx.doi.org/10.1007/s10863-008-9169-3 PMid:18843528.
» http://dx.doi.org/10.1007/s10863-008-9169-3 - Ibeh IN, Saxena DK, Uraih N. Toxicity of aflatoxin: Effects on spermatozoa, oocytes, and in vitro fertilization. J Environ Pathol Toxicol Oncol. 2000;19(4):357-61. PMid:11213017.
- Ibeh IN, Uraih N, Ogonar JI. Dietary exposure to aflatoxin in human male infertility in Benin City, Nigeria. Int J Fertil Menopausal Stud. 1994;39(4):208-14. PMid:7951403.
- Iimura K, Furukawa T, Yamamoto T, Negishi L, Suzuki M, Sakuda S. The mode of action of Cyclo(l-Ala-l-Pro) in Inhibiting Aflatoxin Production of Aspergillus flavus. Toxins (Basel). 2017;9(7):219. http://dx.doi.org/10.3390/toxins9070219 PMid:28704973.
» http://dx.doi.org/10.3390/toxins9070219 - Ioannou D, Miller D, Griffin DK, Tempest HG. Impact of sperm DNA chromatin in the clinic. J Assist Reprod Genet. 2016;33(2):157-66. http://dx.doi.org/10.1007/s10815-015-0624-x PMid:26678492.
» http://dx.doi.org/10.1007/s10815-015-0624-x - Jarošová A. Phthalic acid esters (PAEs) in the Food Chain. Czech J Food Sci. 2006;24(5):223-31. http://dx.doi.org/10.17221/3318-CJFS
» http://dx.doi.org/10.17221/3318-CJFS - Jin Y, Lin X, Miao W, Wang L, Wu Y, Fu Z. Oral exposure of pubertal male mice to endocrine-disrupting chemicals alters fat metabolism in adult livers. Environ Toxicol. 2015;30(12):1434-44. http://dx.doi.org/10.1002/tox.22013 PMid:24916741.
» http://dx.doi.org/10.1002/tox.22013 - Jin Y, Wang L, Chen G, Lin X, Miao W, Fu Z. Exposure of mice to atrazine and its metabolite diaminochlorotriazine elicits oxidative stress and endocrine disruption. Environ Toxicol Pharmacol. 2014;37(2):782-90. http://dx.doi.org/10.1016/j.etap.2014.02.014 PMid:24632104.
» http://dx.doi.org/10.1016/j.etap.2014.02.014 - Jin Y, Zhang X, Shu L, Chen L, Sun L, Qian H, Liu W, Fu Z. Oxidative stress response and gene expression with atrazine exposure in adult female zebrafish (Danio rerio). Chemosphere. 2010;78(7):846-52. http://dx.doi.org/10.1016/j.chemosphere.2009.11.044 PMid:20036412.
» http://dx.doi.org/10.1016/j.chemosphere.2009.11.044 - Johnson KJ, Heger NE, Boekelheide K. Of mice and men (and rats): phthalate-induced fetal testis endocrine disruption is species-dependent. Toxicol Sci. 2012;129(2):235-48. http://dx.doi.org/10.1093/toxsci/kfs206 PMid:22700540.
» http://dx.doi.org/10.1093/toxsci/kfs206 - Kalo D, Hadas R, Furman O, Ben-Ari J, Maor Y, Patterson DG, Tomey C, Roth Z. Carryover effects of acute DEHP exposure on ovarian function and oocyte developmental competence in lactating cows. PLoS One. 2015;10(7):e0130896. http://dx.doi.org/10.1371/journal.pone.0130896 PMid:26154164.
» http://dx.doi.org/10.1371/journal.pone.0130896 - Kalo D, Roth Z. Effects of mono(2-ethylhexyl)phthalate on cytoplasmic maturation of oocytes-the bovine model. Reprod Toxicol. 2015;53:141-51. http://dx.doi.org/10.1016/j.reprotox.2015.04.007 PMid:25900598.
» http://dx.doi.org/10.1016/j.reprotox.2015.04.007 - Kalo D, Roth Z. Low level of mono(2-ethylhexyl) phthalate reduces oocyte developmental competence in association with impaired gene expression. Toxicology. 2017;377:38-48. http://dx.doi.org/10.1016/j.tox.2016.12.005 PMid:27989758.
» http://dx.doi.org/10.1016/j.tox.2016.12.005 - Kalo D, Vitorino Carvalho A, Archilla C, Duranthon V, Moroldo M, Levin Y, Kupervaser M, Smith Y, Roth Z. Mono(2-ethylhexyl) phthalate (MEHP) induces transcriptomic alterations in oocytes and their derived blastocysts. Toxicology. 2019;421:59-73. http://dx.doi.org/10.1016/j.tox.2019.04.016 PMid:31059758.
» http://dx.doi.org/10.1016/j.tox.2019.04.016 - Kavlock R, Barr D, Boekelheide K, Breslin W, Breysse P, Chapin R, Gaido K, Hodgson E, Marcus M, Shea K, Williams P. NTP-CERHR Expert Panel Update on the Reproductive and Developmental Toxicity of di(2-ethylhexyl) phthalate. Reprod Toxicol. 2006;22(3):291-399. http://dx.doi.org/10.1016/j.reprotox.2006.04.007 PMid:17068859.
» http://dx.doi.org/10.1016/j.reprotox.2006.04.007 - Kay VR, Chambers C, Foster WG. Reproductive and developmental effects of phthalate diesters in females. Crit Rev Toxicol. 2013;43(3):200-19. http://dx.doi.org/10.3109/10408444.2013.766149 PMid:23405971.
» http://dx.doi.org/10.3109/10408444.2013.766149 - Kew MC. Aflatoxins as a cause of hepatocellular carcinoma. J Gastrointestin Liver Dis. 2013;22(3):305-10. PMid:24078988.
- Knez J, Kranvogl R, Breznik BP, Vončina E, Vlaisavljević V. Are urinary bisphenol A levels in men related to semen quality and embryo development after medically assisted reproduction? Fertil Steril. 2014;101(1):215-221.e5. http://dx.doi.org/10.1016/j.fertnstert.2013.09.030 PMid:24182411.
» http://dx.doi.org/10.1016/j.fertnstert.2013.09.030 - Koch HM, Bolt HM, Preuss R, Angerer J. New metabolites of di(2-ethylhexyl)phthalate (DEHP) in human urine and serum after single oral doses of deuterium-labelled DEHP. Arch Toxicol. 2005;79(7):367-76. http://dx.doi.org/10.1007/s00204-004-0642-4 PMid:15700144.
» http://dx.doi.org/10.1007/s00204-004-0642-4 - Koch HM, Calafat AM. Human body burdens of chemicals used in plastic manufacture. Philos Trans R Soc Lond B Biol Sci. 2009;364(1526):2063-78. http://dx.doi.org/10.1098/rstb.2008.0208 PMid:19528056.
» http://dx.doi.org/10.1098/rstb.2008.0208 - Komsky-Elbaz A, Roth Z. Effect of the herbicide atrazine and its metabolite DACT on bovine sperm quality. Reprod Toxicol. 2017;67:15-25. http://dx.doi.org/10.1016/j.reprotox.2016.11.001 PMid:27836535.
» http://dx.doi.org/10.1016/j.reprotox.2016.11.001 - Komsky-Elbaz A, Roth Z. Fluorimetric techniques for the assessment of sperm membranes. J. Vis. Exp. 2018(141), e58622. http://dx.doi.org/10.3791/58 622.
- Komsky-Elbaz A, Saktsier M, Roth Z. Aflatoxin B1 impairs sperm quality and fertilization competence. Toxicology. 2018;393:42-50. http://dx.doi.org/10.1016/j.tox.2017.11.007 PMid:29113834.
» http://dx.doi.org/10.1016/j.tox.2017.11.007 - Komsky-Elbaz A, Roth Z. Atrazine-induced DNA fragmentation in bovine spermatozoa is associated with alterations in the transcriptome profile of in-vitro-derived embryos. 2019. In: Abstracts of the 35th Annual Meeting of the Association of Embryo Technology in Europe (AETE); 2019 Sept 13-14; Murcia, Spain. Murcia: AETE.
- Komsky-Elbaz A, Saktsier M, Biran D, Argov-Argaman N, Azaizeh H, Landau YS, Roth Z. Atrazine-induced toxicity in goat spermatozoa is alleviated to some extent by polyphenol-enriched feed. Chemosphere. 2019a;236:124858. http://dx.doi.org/10.1016/j.chemosphere.2019.124858 PMid:31549674.
» http://dx.doi.org/10.1016/j.chemosphere.2019.124858 - Komsky-Elbaz A, Zubov A, Roth Z. Effect of the herbicide atrazine and its major metabolite, DACT, on bovine sperm cryotolerance. Theriogenology. 2019b;140:117-23. http://dx.doi.org/10.1016/j.theriogenology.2019.08.026 PMid:31473494.
» http://dx.doi.org/10.1016/j.theriogenology.2019.08.026 - Krejčíková M, Jarošová A. Phthalate in cow milk depending on the method of milking. Medelnet [serial on the Internet]. 2013:592-6. [cited 2020 June 23]. Available from: http://mnet.mendelu.cz/mendelnet2013/articles/44_krejcikova_795.pdf
» http://mnet.mendelu.cz/mendelnet2013/articles/44_krejcikova_795.pdf - Latini G, De Felice C, Presta G, Del Vecchio A, Paris I, Ruggieri F, Mazzeo P. In utero exposure to di-(2-ethylhexyl)phthalate and duration of human pregnancy. Environ Health Perspect. 2003;111(14):1783-5. http://dx.doi.org/10.1289/ehp.6202 PMid:14594632.
» http://dx.doi.org/10.1289/ehp.6202 - Lavranos G, Balla M, Tzortzopoulou A, Syriou V, Angelopoulou R. Investigating ROS sources in male infertility: a common end for numerous pathways. Reprod Toxicol. 2012;34(3):298-307. http://dx.doi.org/10.1016/j.reprotox.2012.06.007 PMid:22749934.
» http://dx.doi.org/10.1016/j.reprotox.2012.06.007 - LeBaron HM, McFarland JE, Burnside OC. The Triazine Herbicides. Amsterdam: Elsevier; 2008. https://doi.org/10.1016/B978-044451167-6.50004-0
» https://doi.org/10.1016/B978-044451167-6.50004-0 - Li L, Liu J-C, Lai F-N, Liu H-Q, Zhang X-F, Dyce PW, Shen W, Chen H. Di (2-ethylhexyl) Phthalate Exposure Impairs Growth of Antral Follicle in Mice. PLoS One. 2016;11(2):e0148350. http://dx.doi.org/10.1371/journal.pone.0148350 PMid:26845775.
» http://dx.doi.org/10.1371/journal.pone.0148350 - Liu J, Wang Q-C, Han J, Xiong B, Sun S-C. Aflatoxin B1 is toxic to porcine oocyte maturation. Mutagenesis. 2015;30(4):527-35. http://dx.doi.org/10.1093/mutage/gev015 PMid:25778688.
» http://dx.doi.org/10.1093/mutage/gev015 - Ljungvall K, Tienpont B, David F, Magnusson U, Törneke K. Kinetics of orally administered di(2-ethylhexyl) phthalate and its metabolite, mono(2-ethylhexyl) phthalate, in male pigs. Arch Toxicol. 2004;78(7):384-9. http://dx.doi.org/10.1007/s00204-004-0558-z PMid:15022035.
» http://dx.doi.org/10.1007/s00204-004-0558-z - Lovekamp-Swan T, Davis BJ. Mechanisms of phthalate ester toxicity in the female reproductive system. Environ Health Perspect. 2003;111(2):139-45. http://dx.doi.org/10.1289/ehp.5658 PMid:12573895.
» http://dx.doi.org/10.1289/ehp.5658 - Lu Y, Lin M, Aitken RJ. Exposure of spermatozoa to dibutyl phthalate induces abnormal embryonic development in a marine invertebrate Galeolaria caespitosa (Polychaeta: serpulidae). Aquat Toxicol. 2017;191:189-200. http://dx.doi.org/10.1016/j.aquatox.2017.08.008 PMid:28843738.
» http://dx.doi.org/10.1016/j.aquatox.2017.08.008 - Lu Z, Zhang C, Han C, An Q, Cheng Y, Chen Y, Chen Y, Meng R, Zhang Y, Su J. Plasticizer Bis(2-ethylhexyl) Phthalate causes meiosis defects and decreases fertilization ability of mouse oocytes in vivo. J Agric Food Chem. 2019;67(12):3459-68. https://doi.org/10.1021/acs.jafc.9b00121
» https://doi.org/10.1021/acs.jafc.9b00121 - Luciano AM, Franciosi F, Modina SC, Lodde V. Gap junction-mediated communications regulate chromatin remodeling during bovine oocyte growth and differentiation through cAMP-Dependent Mechanism(s). Biol Reprod. 2011;85(6):1252-9. http://dx.doi.org/10.1095/biolreprod.111.092858 PMid:21816847.
» http://dx.doi.org/10.1095/biolreprod.111.092858 - Lussier JG, Matton P, Dufour JJ. Growth rates of follicles in the ovary of the cow. J Reprod Fertil. 1987;81(2):301-7. http://dx.doi.org/10.1530/jrf.0.0810301
» http://dx.doi.org/10.1530/jrf.0.0810301 - Ma L, Wang J, Zhang Y. Probing the characterization of the interaction of Aflatoxins B1 and G1 with Calf Thymus DNA In Vitro. Toxins (Basel). 2017;9(7):209. http://dx.doi.org/10.3390/toxins9070209 PMid:28671585.
» http://dx.doi.org/10.3390/toxins9070209 - Machtinger R, Gaskins AJ, Racowsky C, Mansur A, Adir M, Baccarelli AA, Calafat AM, Hauser R. Urinary concentrations of biomarkers of phthalates and phthalate alternatives and IVF outcomes. Environ Int. 2018;111:23-31. http://dx.doi.org/10.1016/j.envint.2017.11.011 PMid:29161633.
» http://dx.doi.org/10.1016/j.envint.2017.11.011 - Manikkam M, Tracey R, Guerrero-Bosagna C, Skinner MK. Plastics derived endocrine disruptors (BPA, DEHP and DBP) induce epigenetic transgenerational inheritance of obesity, reproductive disease and sperm epimutations. PLoS One. 2013;8(1):e55387. http://dx.doi.org/10.1371/journal.pone.0055387 PMid:23359474.
» http://dx.doi.org/10.1371/journal.pone.0055387 - Mann U, Shiff B, Patel P. Reasons for worldwide decline in male fertility. Curr Opin Urol. 2020;1(3):296-301. http://dx.doi.org/10.1097/MOU.0000000000000745 PMid:32168194.
» http://dx.doi.org/10.1097/MOU.0000000000000745 - Maravilla-Galván R, Fierro R, González-Márquez H, Gómez-Arroyo S, Jiménez I, Betancourt M. Effects of atrazine and fenoxaprop-ethyl on capacitation and the acrosomal reaction in boar sperm. Int J Toxicol. 2009;28(1):24-32. http://dx.doi.org/10.1177/1091581809333138 PMid:19482828.
» http://dx.doi.org/10.1177/1091581809333138 - Martine B, Marie-Jeanne T, Cendrine D, Fabrice A, Marc C. Assessment of Adult Human Exposure to Phthalate Esters in the Urban Centre of Paris (France). Bull Environ Contam Toxicol. 2013;90(1):91-6. http://dx.doi.org/10.1007/s00128-012-0859-5 PMid:23090363.
» http://dx.doi.org/10.1007/s00128-012-0859-5 - Marzano G, Mastrorocco A, Zianni R, Mangiacotti M, Chiaravalle AE, Lacalandra GM, Minervini F, Cardinali A, Macciocca M, Vicenti R, Fabbri R, Hinrichs K, Dell’Aquila ME, Martino NA. Altered morphokinetics in equine embryos from oocytes exposed to DEHP during IVM. Mol Reprod Dev. 2019;86(10):1388-404. http://dx.doi.org/10.1002/mrd.23156 PMid:31025442.
» http://dx.doi.org/10.1002/mrd.23156 - McBirney M, King SE, Pappalardo M, Houser E, Unkefer M, Nilsson E, Sadler-Riggleman I, Beck D, Winchester P, Skinner MK. Atrazine induced epigenetic transgenerational inheritance of disease, lean phenotype and sperm epimutation pathology biomarkers. PLoS One. 2017;12(9):e0184306. http://dx.doi.org/10.1371/journal.pone.0184306 PMid:28931070.
» http://dx.doi.org/10.1371/journal.pone.0184306 - McKee RH, Butala JH, David RM, Gans G. NTP center for the evaluation of risks to human reproduction reports on phthalates: addressing the data gaps. Reprod Toxicol. 2004;18(1):1-22. http://dx.doi.org/10.1016/j.reprotox.2003.09.002 PMid:15013060.
» http://dx.doi.org/10.1016/j.reprotox.2003.09.002 - Meeker JD, Ehrlich S, Toth TL, Wright DL, Calafat AM, Trisini AT, Ye X, Hauser R. Semen quality and sperm DNA damage in relation to urinary bisphenol A among men from an infertility clinic. Reprod Toxicol. 2010;30(4):532-9. http://dx.doi.org/10.1016/j.reprotox.2010.07.005 PMid:20656017.
» http://dx.doi.org/10.1016/j.reprotox.2010.07.005 - Meissonnier GM, Pinton P, Laffitte J, Cossalter A-M, Gong YY, Wild CP, Bertin G, Galtier P, Oswald IP. Immunotoxicity of aflatoxin B1: impairment of the cell-mediated response to vaccine antigen and modulation of cytokine expression. Toxicol Appl Pharmacol. 2008;231(2):142-9. http://dx.doi.org/10.1016/j.taap.2008.04.004 PMid:18501398.
» http://dx.doi.org/10.1016/j.taap.2008.04.004 - Monson M, Cardona C, Coulombe R, Reed K. Hepatic Transcriptome Responses of Domesticated and Wild Turkey Embryos to Aflatoxin B1. Toxins (Basel). 2016;8(1):16. http://dx.doi.org/10.3390/toxins8010016 PMid:26751476.
» http://dx.doi.org/10.3390/toxins8010016 - Moyer B, Hixon ML. Reproductive effects in F1 adult females exposed in utero to moderate to high doses of mono-2-ethylhexylphthalate (MEHP). Reprod Toxicol. 2012;34(1):43-50. http://dx.doi.org/10.1016/j.reprotox.2012.02.006 PMid:22401849.
» http://dx.doi.org/10.1016/j.reprotox.2012.02.006 - Nduka U, Ibeh NI, Oluwafemi F. A Study on the Impact of Aflatoxin on Human Reproduction. Afr J Reprod Health. 2001;5(1):106-10. http://dx.doi.org/10.2307/3583204
» http://dx.doi.org/10.2307/3583204 - Net S, Delmont A, Sempéré R, Paluselli A, Ouddane B. Reliable quantification of phthalates in environmental matrices (air, water, sludge, sediment and soil): A review. Sci Total Environ. 2015a;515-516:162-80. http://dx.doi.org/10.1016/j.scitotenv.2015.02.013 PMid:25723871.
» http://dx.doi.org/10.1016/j.scitotenv.2015.02.013 - Net S, Sempéré R, Delmont A, Paluselli A, Ouddane B. Occurrence, fate, behavior and ecotoxicological state of phthalates in different environmental matrices. Environ Sci Technol. 2015b;49(7):4019-35. http://dx.doi.org/10.1021/es505233b PMid:25730609.
» http://dx.doi.org/10.1021/es505233b - Niermann S, Rattan S, Brehm E, Flaws JA. Prenatal exposure to di-(2-ethylhexyl) phthalate (DEHP) affects reproductive outcomes in female mice. Reprod Toxicol. 2015;53:23-32. http://dx.doi.org/10.1016/j.reprotox.2015.02.013 PMid:25765777.
» http://dx.doi.org/10.1016/j.reprotox.2015.02.013 - Oliva R, Castillo J, Estanyol J, Ballescà J. Human sperm chromatin epigenetic potential: genomics, proteomics, and male infertility. Asian J Androl. 2015;17(4):601-609. http://dx.doi.org/10.4103/1008-682X.153302
» http://dx.doi.org/10.4103/1008-682X.153302 - Pant N, Pant A, Shukla M, Mathur N, Gupta Y, Saxena D. Environmental and experimental exposure of phthalate esters: the toxicological consequence on human sperm. Hum Exp Toxicol. 2011;30(6):507-14. http://dx.doi.org/10.1177/0960327110374205 PMid:20551087.
» http://dx.doi.org/10.1177/0960327110374205 - Parra-Forero LY, Veloz-Contreras A, Vargas-Marín S, Mojica-Villegas MA, Alfaro-Pedraza E, Urióstegui-Acosta M, Hernández-Ochoa I. Alterations in oocytes and early zygotes following oral exposure to di(2-ethylhexyl) phthalate in young adult female mice. Reprod Toxicol. 2019;90:53-61. http://dx.doi.org/10.1016/j.reprotox.2019.08.012 PMid:31442482.
» http://dx.doi.org/10.1016/j.reprotox.2019.08.012 - Peighambarzadeh S, Safi S, Shahtaheri S, Javanbakht M, Rahimi Forushani A. Rahimi Forushani a. Presence of atrazine in the biological samples of cattle and its consequence adversity in human health. Iran J Public Health. 2011;40(4):112-21. PMid:23113110.
- Perry MJ. Effects of environmental and occupational pesticide exposure on human sperm: A systematic review. Hum Reprod Update. 2008;14(3):233-42. http://dx.doi.org/10.1093/humupd/dmm039 PMid:18281240.
» http://dx.doi.org/10.1093/humupd/dmm039 - Petersen JH, Breindahl T. Plasticizers in total diet samples, baby food and infant formulae. Food Addit Contam. 2000;17(2):133-41. http://dx.doi.org/10.1080/026520300283487 PMid:10793844.
» http://dx.doi.org/10.1080/026520300283487 - Pflieger-Bruss S, Schuppe H-C, Schill W-B. The male reproductive system and its susceptibility to endocrine disrupting chemicals. Andrologia. 2004;36(6):337-45. http://dx.doi.org/10.1111/j.1439-0272.2004.00641.x PMid:15541049.
» http://dx.doi.org/10.1111/j.1439-0272.2004.00641.x - Pure Strategies. Sources of phthalates in dairy farm equipment. Prepared for environmental health strategy center and the coalition for safer food processing & packaging [Internet] Pure Strategies; 2018 [cited 2020 June 23]. Available from: http://kleanupkraft.org/Phthalates-Farm-Equipment.pdf
» http://kleanupkraft.org/Phthalates-Farm-Equipment.pdf - Raisuddin S, Singh KP, Zaidi SI, Paul BN, Ray PK. Immunosuppressive effects of aflatoxin in growing rats. Mycopathologia. 1993;124(3):189-94. http://dx.doi.org/10.1007/BF01103737 PMid:8022466.
» http://dx.doi.org/10.1007/BF01103737 - Rajkovic A, Uyttendaele M, Debevere J. Computer aided boar semen motility analysis for cereulide detection in different food matrices. Int J Food Microbiol. 2007;114(1):92-9. http://dx.doi.org/10.1016/j.ijfoodmicro.2006.09.031 PMid:17174428.
» http://dx.doi.org/10.1016/j.ijfoodmicro.2006.09.031 - Ramalho-Santos J, Varum S, Amaral S, Mota PC, Sousa AP, Amaral A. Mitochondrial functionality in reproduction: from gonads and gametes to embryos and embryonic stem cells. Hum Reprod Update. 2009;15(5):553-72. http://dx.doi.org/10.1093/humupd/dmp016 PMid:19414527.
» http://dx.doi.org/10.1093/humupd/dmp016 - Rando OJ. Daddy issues: paternal effects on phenotype. Cell. 2012;151(4):702-8. http://dx.doi.org/10.1016/j.cell.2012.10.020 PMid:23141533.
» http://dx.doi.org/10.1016/j.cell.2012.10.020 - Reyes-Moreno C, Boilard M, Sullivan R, Sirard MA. Characterization and Identification of Epididymal Factors That Protect Ejaculated Bovine Sperm During In Vitro Storage. Biol Reprod. 2002;66(1):159-66. http://dx.doi.org/10.1095/biolreprod66.1.159 PMid:11751277.
» http://dx.doi.org/10.1095/biolreprod66.1.159 - Rhind SM, Kyle CE, Telfer G, Duff EI, Smith A. Alkyl phenols and diethylhexyl phthalate in tissues of sheep grazing pastures fertilized with sewage sludge or inorganic fertilizer. Environ Health Perspect. 2005;113(4):447-53. http://dx.doi.org/10.1289/ehp.7469 PMid:15811823.
» http://dx.doi.org/10.1289/ehp.7469 - Ross MK, Jones TL, Filipov NM. Disposition of the Herbicide 2-Chloro-4-(ethylamino)-6-(isopropylamino)-s-triazine (Atrazine) and its major metabolites in mice: A liquid chromatography / mass spectrometry analysis of urine. Plasma, and Tissue Levels. 2009;37(4):776-86. http://dx.doi.org/10.1124/dmd.108.024927 PMid:19116264.
» http://dx.doi.org/10.1124/dmd.108.024927 - Roth Z. Symposium review: Reduction in oocyte developmental competence by stress is associated with alterations in mitochondrial function1. J Dairy Sci. 2018;101(4):3642-54. http://dx.doi.org/10.3168/jds.2017-13389 PMid:29395145.
» http://dx.doi.org/10.3168/jds.2017-13389 - Sánchez-Peña LC, Reyes BE, López-Carrillo L, Recio R, Morán-Martínez J, Cebrián ME, Quintanilla-Vega B. Organophosphorous pesticide exposure alters sperm chromatin structure in Mexican agricultural workers. Toxicol Appl Pharmacol. 2004;196(1):108-13. http://dx.doi.org/10.1016/j.taap.2003.11.023 PMid:15050412.
» http://dx.doi.org/10.1016/j.taap.2003.11.023 - Sant KE, Dolinoy DC, Jilek JL, Sartor MA, Harris C. Mono-2-ethylhexyl phthalate disrupts neurulation and modifies the embryonic redox environment and gene expression. Reprod Toxicol. 2016;63:32-48. http://dx.doi.org/10.1016/j.reprotox.2016.03.042 PMid:27167697.
» http://dx.doi.org/10.1016/j.reprotox.2016.03.042 - Saradha B, Mathur PP. Effect of environmental contaminants on male reproduction. Environ Toxicol Pharmacol. 2006;21(1):34-41. http://dx.doi.org/10.1016/j.etap.2005.06.004 PMid:21783636.
» http://dx.doi.org/10.1016/j.etap.2005.06.004 - Schenzel J, Forrer H-R, Vogelgsang S, Hungerbühler K, Bucheli TD. Mycotoxins in the Environment: I. Production and Emission from an Agricultural Test Field. Environ Sci Technol. 2012;46(24):13067-75. http://dx.doi.org/10.1021/es301557m PMid:23145781.
» http://dx.doi.org/10.1021/es301557m - Sergerie M, Laforest G, Bujan L, Bissonnette F, Bleau G. Sperm DNA fragmentation: threshold value in male fertility. Hum Reprod. 2005;20(12):3446-51. http://dx.doi.org/10.1093/humrep/dei231 PMid:16085665.
» http://dx.doi.org/10.1093/humrep/dei231 - Severi-Aguiar GDC, Silva-Zacarin ECM. Effects of herbicide atrazine in experimental animal models. In: Hasaneen MN. Herbicides: Properties, synthesis and control of weeds. Rijeka: InTech; 2011. p. 285-96.
- Shen HM, Shi CY, Lee HP, Ong CN. Aflatoxin B1-induced lipid peroxidation in rat liver. Toxicol Appl Pharmacol. 1994;127(1):145-50. http://dx.doi.org/10.1006/taap.1994.1148 PMid:8048046.
» http://dx.doi.org/10.1006/taap.1994.1148 - Shuaib FMB, Ehiri J, Abdullahi A, Williams JH, Jolly PE. Reproductive health effects of aflatoxins: a review of the literature. Reprod Toxicol. 2010;29(3):262-70. http://dx.doi.org/10.1016/j.reprotox.2009.12.005 PMid:20045048.
» http://dx.doi.org/10.1016/j.reprotox.2009.12.005 - Siddeek B, Mauduit C, Simeoni U, Benahmed M. Sperm epigenome as a marker of environmental exposure and lifestyle, at the origin of diseases inheritance. Mutat Res Mutat Res. 2018;778:38-44. http://dx.doi.org/10.1016/j.mrrev.2018.09.001 PMid:30454681.
» http://dx.doi.org/10.1016/j.mrrev.2018.09.001 - Silva MJ, Reidy JA, Herbert AR, Preau JL Jr, Needham LL, Calafat AM. Detection of phthalate metabolites in human amniotic fluid. Bull Environ Contam Toxicol. 2004;72(6):1226-31. http://dx.doi.org/10.1007/s00128-004-0374-4 PMid:15362453.
» http://dx.doi.org/10.1007/s00128-004-0374-4 - Simões R, Feitosa WB, Siqueira AFP, Nichi M, Paula-Lopes FF, Marques MG, Peres MA, Barnabe VH, Visintin JA, Assumpção ME. Influence of bovine sperm DNA fragmentation and oxidative stress on early embryo in vitro development outcome. Reproduction. 2013;146(5):433-41. http://dx.doi.org/10.1530/REP-13-0123 PMid:23940385.
» http://dx.doi.org/10.1530/REP-13-0123 - Skinner MK, Nilsson E, Sadler-Riggleman I, Beck D, Ben Maamar M, McCarrey JR. Transgenerational sperm DNA methylation epimutation developmental origins following ancestral vinclozolin exposure. Epigenetics. 2019;14(7):721-39. http://dx.doi.org/10.1080/15592294.2019.1614417 PMid:31079544.
» http://dx.doi.org/10.1080/15592294.2019.1614417 - Sotomayor RE, Sega GA. Unscheduled DNA synthesis assay in mammalian spermatogenic cells: an update. Environ Mol Mutagen. 2000;36(4):255-65. http://dx.doi.org/10.1002/1098-2280(2000)36:4<255::AID-EM1>3.0.CO;2-O PMid:11152558.
» http://dx.doi.org/10.1002/1098-2280(2000)36:4<255::AID-EM1>3.0.CO;2-O - Staub C, Johnson L. Review: spermatogenesis in the bull. Animal. 2018;12(s1):s27-35. http://dx.doi.org/10.1017/S1751731118000435 PMid:29882505.
» http://dx.doi.org/10.1017/S1751731118000435 - Storvik M, Huuskonen P, Kyllönen T, Lehtonen S, El-Nezami H, Auriola S, Pasanen M. Aflatoxin B1 – a potential endocrine disruptor – up-regulates CYP19A1 in JEG-3 cells. Toxicol Lett. 2011;202(3):161-7. http://dx.doi.org/10.1016/j.toxlet.2011.01.028 PMid:21296134.
» http://dx.doi.org/10.1016/j.toxlet.2011.01.028 - Strosnider H, Azziz-Baumgartner E, Banziger M, Bhat RV, Breiman R, Brune M-N, DeCock K, Dilley A, Groopman J, Hell K, Henry SH, Jeffers D, Jolly C, Jolly P, Kibata GN, Lewis L, Liu X, Luber G, McCoy L, Mensah P, Miraglia M, Misore A, Njapau H, Ong CN, Onsongo MT, Page SW, Park D, Patel M, Phillips T, Pineiro M, Pronczuk J, Rogers HS, Rubin C, Sabino M, Schaafsma A, Shephard G, Stroka J, Wild C, Williams JT, Wilson D. Workgroup Report: Public Health Strategies for Reducing Aflatoxin Exposure in Developing Countries. Environ Health Perspect. 2006;114(12):1898-903. http://dx.doi.org/10.1289/ehp.9302 PMid:17185282.
» http://dx.doi.org/10.1289/ehp.9302 - Supriya C, Girish BP, Reddy PS. Aflatoxin B1-Induced Reproductive Toxicity in Male Rats: Possible Mechanism of Action. Int J Toxicol. 2014;33(3):155-61. http://dx.doi.org/10.1177/1091581814530764 PMid:24728861.
» http://dx.doi.org/10.1177/1091581814530764 - Swan SH, Kruse RL, Liu F, Barr DB, Drobnis EZ, Redmon JB, Wang C, Brazil C, Overstreet JW. Semen Quality in Relation to Biomarkers of Pesticide Exposure. Environ Health Perspect. 2003;111(12):1478-84. http://dx.doi.org/10.1289/ehp.6417 PMid:12948887.
» http://dx.doi.org/10.1289/ehp.6417 - Tavares RS, Escada-Rebelo S, Correia M, Mota PC, Ramalho-Santos J. The non-genomic effects of endocrine-disrupting chemicals on mammalian sperm. Reproduction. 2016;151(1):R1-13. http://dx.doi.org/10.1530/REP-15-0355 PMid:26585413.
» http://dx.doi.org/10.1530/REP-15-0355 - Thouas GA, Trounson AO, Wolvetang EJ, Jones GM. Mitochondrial Dysfunction in Mouse Oocytes Results in Preimplantation Embryo Arrest in Vitro. Biol Reprod. 2004;71(6):1936-42. http://dx.doi.org/10.1095/biolreprod.104.033589 PMid:15286028.
» http://dx.doi.org/10.1095/biolreprod.104.033589 - Toft G, Jönsson BAG, Lindh CH, Jensen TK, Hjollund NH, Vested A, Bonde JP. Association between Pregnancy Loss and Urinary Phthalate Levels around the Time of Conception. Environ Health Perspect. 2011;120(3):458-63. http://dx.doi.org/10.1289/ehp.1103552 PMid:22113848.
» http://dx.doi.org/10.1289/ehp.1103552 - Tomita I, Nakamura Y, Aoki N, Inui N. Mutagenic/carcinogenic potential of DEHP and MEHP. Environ Health Perspect. 1982;45:119-25. http://dx.doi.org/10.1289/ehp.8245119 PMid:6814903.
» http://dx.doi.org/10.1289/ehp.8245119 - Tranfo G, Caporossi L, Paci E, Aragona C, Romanzi D, De Carolis C, De Rosa M, Capanna S, Papaleo B, Pera A. Urinary phthalate monoesters concentration in couples with infertility problems. Toxicol Lett. 2012;213(1):15-20. http://dx.doi.org/10.1016/j.toxlet.2011.11.033 PMid:22197707.
» http://dx.doi.org/10.1016/j.toxlet.2011.11.033 - Treinen KA, Dodson WC, Heindel JJ. Inhibition of FSH-stimulated cAMP accumulation and progesterone production by mono(2-ethylhexyl) phthalate in rat granulosa cell cultures. Toxicol Appl Pharmacol. 1990;106(2):334-40. http://dx.doi.org/10.1016/0041-008X(90)90252-P PMid:2175055.
» http://dx.doi.org/10.1016/0041-008X(90)90252-P - Uppangala S, Pudakalakatti S, D’souza F, Salian SR, Kalthur G, Kumar P, Atreya H, Adiga SK. Influence of sperm DNA damage on human preimplantation embryo metabolism. Reprod Biol. 2016;16(3):234-41. http://dx.doi.org/10.1016/j.repbio.2016.07.004 PMid:27492188.
» http://dx.doi.org/10.1016/j.repbio.2016.07.004 - Verma RJ, Nair A. Ameliorative effect of vitamin E on aflatoxin-induced lipid peroxidation in the testis of mice. Asian J Androl. 2001;3(3):217-21. PMid:11561193.
- Verma RJ. Aflatoxin cause DNA damage. Int J Hum Genet. 2004;4(4):231-6. http://dx.doi.org/10.1080/09723757.2004.11885899
» http://dx.doi.org/10.1080/09723757.2004.11885899 - Virro MR, Larson-Cook KL, Evenson DP. Sperm chromatin structure assay (SCSA®) parameters are related to fertilization, blastocyst development, and ongoing pregnancy in in vitro fertilization and intracytoplasmic sperm injection cycles. Fertil Steril. 2004;81(5):1289-95. http://dx.doi.org/10.1016/j.fertnstert.2003.09.063 PMid:15136092.
» http://dx.doi.org/10.1016/j.fertnstert.2003.09.063 - Wang T, Han J, Duan X, Xiong B, Cui X-S, Kim N-H, Liu HL, Sun SC. The toxic effects and possible mechanisms of Bisphenol A on oocyte maturation of porcine in vitro. Oncotarget. 2016;7(22):32554-65. http://dx.doi.org/10.18632/oncotarget.8689 PMid:27086915.
» http://dx.doi.org/10.18632/oncotarget.8689 - Wang W, Craig ZR, Basavarajappa MS, Hafner KS, Flaws JA. Mono-(2-ethylhexyl) phthalate induces oxidative stress and inhibits growth of mouse ovarian antral follicles. Biol Reprod. 2012;87(6):152. http://dx.doi.org/10.1095/biolreprod.112.102467 PMid:23077170.
» http://dx.doi.org/10.1095/biolreprod.112.102467 - Wei Y, Schatten H, Sun Q-Y. Environmental epigenetic inheritance through gametes and implications for human reproduction. Hum Reprod Update. 2015;21(2):194-208. http://dx.doi.org/10.1093/humupd/dmu061 PMid:25416302.
» http://dx.doi.org/10.1093/humupd/dmu061 - Wei Y, Yang CR, Wei YP, Zhao ZA, Hou Y, Schatten H, Sun QY. Paternally induced transgenerational inheritance of susceptibility to diabetes in mammals. Proc Natl Acad Sci USA. 2014;111(5):1873-8. http://dx.doi.org/10.1073/pnas.1321195111 PMid:24449870.
» http://dx.doi.org/10.1073/pnas.1321195111 - Williams JH, Phillips TD, Jolly PE, Stiles JK, Jolly CM, Aggarwal D. Human aflatoxicosis in developing countries: a review of toxicology, exposure, potential health consequences, and interventions. Am J Clin Nutr. 2004;80(5):1106-22. http://dx.doi.org/10.1093/ajcn/80.5.1106 PMid:15531656.
» http://dx.doi.org/10.1093/ajcn/80.5.1106 - Wirbisky-Hershberger SE, Sanchez OF, Horzmann KA, Thanki D, Yuan C, Freeman JL. Atrazine exposure decreases the activity of DNMTs, global DNA methylation levels, and dnmt expression. Food Chem Toxicol. 2017;109(Pt 1):727-34. http://dx.doi.org/10.1016/j.fct.2017.08.041 PMid:28859886.
» http://dx.doi.org/10.1016/j.fct.2017.08.041 - Wiser A, Sachar S, Ghetler Y, Shulman A, Breitbart H. Assessment of sperm hyperactivated motility and acrosome reaction can discriminate the use of spermatozoa for conventional in vitro fertilisation or intracytoplasmic sperm injection: preliminary results. Andrologia. 2014;46(3):313-5. http://dx.doi.org/10.1111/and.12068 PMid:23356546.
» http://dx.doi.org/10.1111/and.12068 - Yagi Y, Nakamura Y, Tomita I, Tsuchikawa K, Shimoi N. Teratogenic potential of di- and mono-(2-ethylhexyl)phthalate in mice. J Environ Pathol Toxicol. 1980;4(2-3):533-44. PMid:7462917.
- Yuan S, Wu B, Yu Z, Fang J, Liang N, Zhou M, Huang C, Peng X. The mitochondrial and endoplasmic reticulum pathways involved in the apoptosis of bursa of Fabricius cells in broilers exposed to dietary aflatoxin B1. Oncotarget. 2016;7(40):65295-306. http://dx.doi.org/10.18632/oncotarget.11321 PMid:27542244.
» http://dx.doi.org/10.18632/oncotarget.11321 - Zama AM, Bhurke A, Uzumcu M. Effects of Endocrine-disrupting Chemicals on Female Reproductive Health. Open Biotechnol J. 2016;10(1):54-75. http://dx.doi.org/10.2174/1874070701610010054
» http://dx.doi.org/10.2174/1874070701610010054 - Zamkowska D, Karwacka A, Jurewicz J, Radwan M. Environmental exposure to non-persistent endocrine disrupting chemicals and semen quality: an overview of the current epidemiological evidence. Int J Occup Med Environ Health. 2018;31(4):377-414. http://dx.doi.org/10.13075/ijomeh.1896.01195 PMid:30160090.
» http://dx.doi.org/10.13075/ijomeh.1896.01195 - Zhang T, Li L, Qin X-S, Zhou Y, Zhang X-F, Wang L-Q, De Felici M, Chen H, Qin GQ, Shen W. Di-(2-ethylhexyl) phthalate and bisphenol A exposure impairs mouse primordial follicle assembly in vitro. Environ Mol Mutagen. 2014;55(4):343-53. http://dx.doi.org/10.1002/em.21847 PMid:24458533.
» http://dx.doi.org/10.1002/em.21847 - Zhang X-F, Zhang L-J, Li L, Feng Y-N, Chen B, Ma J-M, Huynh E, Shi QH, De Felici M, Shen W. Diethylhexyl phthalate exposure impairs follicular development and affects oocyte maturation in the mouse. Environ Mol Mutagen. 2013;54(5):354-61. http://dx.doi.org/10.1002/em.21776 PMid:23625783.
» http://dx.doi.org/10.1002/em.21776 - Zhang Y, Wang T, Lan M, Zang XW, Li YL, Cui XS, Kim NH, Sun SC. Melatonin protects oocytes from MEHP exposure-induced meiosis defects in porcine. Biol Reprod. 2018;98(3):286-98. http://doi.org//10.1093/biolre/iox185
» http://doi.org//10.1093/biolre/iox185
Publication Dates
-
Publication in this collection
10 Aug 2020 -
Date of issue
2020
History
-
Received
21 Apr 2020 -
Accepted
23 June 2020



