Abstract
The Cerrado biome is located in the central region of Brazil and consists mainly of savanna vegetation. In this study we assessed the richness of tiger moths (Arctiidae) of the Brazilian Cerrado. Specifically, we 1) assessed species richness in one-degree cells in the biome, 2) identified areas where these moths are poorly known, and 3) tested if similarities in species composition are related to geographical distance in the relatively well-sampled areas. We obtained the data mainly from specimens deposited in museums, but we also included additional information from the literature. We compiled 2,321 records belonging to 723 species. Specimens were recorded in 108 localities distributed in 67 one-degree cells. Species occurring exclusively in one or two one-degree cells represented 64% of the total number of species. Sample effort was not uniform in the biome, as there were very few records in the northern region of the Cerrado. The best-sampled one-degree cell had 239 species. Species assemblages were structured in space with a clear trend of localities near one another presenting more similarities in faunal composition than distant localities. This distance decay in similarity was slightly more pronounced along the longitudinal than along the latitudinal distances. We conclude that the Cerrado still remains poorly inventoried for tiger moths, particularly in its northern portion, where many unrecorded species may be found in the future. Despite of this limited knowledge, the best-sampled region indicates that richness of tiger moths in the Cerrado is comparable to the species-rich forest biomes in the Neotropical region.
Arctiinae; distance decay in similarity; Lithosiinae; museum data; sample effort
ECOLOGY
Richness of tiger moths (Lepidoptera: Arctiidae) in the Brazilian Cerrado: how much do we know?
Viviane G. FerroI, * * Corresponding author. E-mail: vgferro@yahoo.com ; Adriano S. MeloI; Ivone R. DinizII
IDepartamento de Ecologia, Instituto de Ciências Biológicas, Universidade Federal de Goiás. Caixa Postal 131, 74001-970 Goiânia, GO, Brazil
IIDepartamento de Zoologia, Instituto de Ciências Biológicas, Universidade de Brasília. 70910-900 Brasília, DF, Brazil
ABSTRACT
The Cerrado biome is located in the central region of Brazil and consists mainly of savanna vegetation. In this study we assessed the richness of tiger moths (Arctiidae) of the Brazilian Cerrado. Specifically, we 1) assessed species richness in one-degree cells in the biome, 2) identified areas where these moths are poorly known, and 3) tested if similarities in species composition are related to geographical distance in the relatively well-sampled areas. We obtained the data mainly from specimens deposited in museums, but we also included additional information from the literature. We compiled 2,321 records belonging to 723 species. Specimens were recorded in 108 localities distributed in 67 one-degree cells. Species occurring exclusively in one or two one-degree cells represented 64% of the total number of species. Sample effort was not uniform in the biome, as there were very few records in the northern region of the Cerrado. The best-sampled one-degree cell had 239 species. Species assemblages were structured in space with a clear trend of localities near one another presenting more similarities in faunal composition than distant localities. This distance decay in similarity was slightly more pronounced along the longitudinal than along the latitudinal distances. We conclude that the Cerrado still remains poorly inventoried for tiger moths, particularly in its northern portion, where many unrecorded species may be found in the future. Despite of this limited knowledge, the best-sampled region indicates that richness of tiger moths in the Cerrado is comparable to the species-rich forest biomes in the Neotropical region.
Key words: Arctiinae; distance decay in similarity; Lithosiinae; museum data; sample effort.
The Cerrado occupies an area of 2 million km2 and is the second largest biome of South America, second only to the Amazon forest (RATTER et al. 1997). The bulk of the Cerrado is located on the Brazilian central plateau (hereafter, designed by core region), with small and isolated areas in southern, northern, and northeastern Brazil, as well as northwestern Paraguay and eastern Bolivia (BRIDGEWATER et al. 2004). The Cerrado is part of a xeric vegetation corridor that includes the Caatinga biome, in northeastern Brazil, and the Chaco, in Argentina, Paraguay, and Bolivia (OLIVEIRA-FILHO & RATTER 1995). This dry corridor runs between two important moist forest regions in tropical South America: the Amazon forest in the northwest, and the Atlantic forest in the east and south.
The biological richness of the Cerrado is high for most taxonomic groups documented to date. The flora of this biome, for instance, is the richest of all Neotropical savannas, including 951 wood species (BRIDGEWATER et al. 2004). Moreover, there are records of 856 species of birds (SILVA & SANTOS 2005), 161 species of mammals, 150 species of amphibians, and 120 species of reptiles (MYERS et al. 2000). DIAS (1996) estimated that 90,000 insect species occur in the Cerrado, including approximately 35% of the bees, 23% of the termites, and 13% of butterflies known to occur in the neotropics (CAVALCANTI & JOLY 2002). As for moths, CAMARGO & BECKER (1999) estimated that 12% of the saturniids of the New World probably occur in the Cerrado.
MYERS et al. (2000) estimated that 80% of the Cerrado has been subjected to some type of human alteration, and considered this biome as a world hotspot. A recent survey showed that more than half of the Cerrado has been cleared or transformed for human uses in the past 35 years (KLINK & MACHADO 2005). At least 112 species of Cerrado animals are threatened with extinction (FUNDAÇÃO BIODIVERSITAS 2008), and only 2.2% of the Cerrado area is legally protected. Moreover, approximately 20% of endemic and threatened species occur outside the parks and reserves of this region (KLINK & MACHADO 2005).
Species of the order Lepidoptera, which includes butterflies and moths, have commonly been employed in studies addressing conservation of disturbed habitats because they quickly respond to environmental changes (e.g., DEVRIES et al. 1997, KITCHING et al. 2000). Additionally, the order is a species-rich group whose representatives are usually locally abundant, accessible to standardized sampling and, at least for some families, the taxonomy is relatively well known (DEVRIES et al. 1997, FERRO & DINIZ 2008). Moths of the families Arctiidae, Saturniidae, and Sphingidae, in particular, are commonly used as indicators in ecosystem monitoring programs (HILTY & MERENLENDER 2000).
The arctiids, commonly known as tiger moths, include almost 11,000 species (SCOBLE 1995). A total of 5,931 described species was recorded in the Neotropical region (WATSON & GOODGER 1986, HEPPNER 1991) and BROWN JR & FREITAS (1999) estimated that there are nearly 2,000 species in Brazil. According to JACOBSON & WELLER (2002), the family contains three subfamilies: Arctiinae, Lithosiinae, and Syntominae. The Arctiinae are small-to-medium sized moths (SCOBLE 1995) that occur world-wide, but are specially rich in the neotropics, comprising 73% of the 6,523 described species (HEPPNER 1991). The Lithosiinae are cosmopolitan small-sized moths, but only 34% of the 3,445 species occur in the neotropics (HEPPNER 1991, SCOBLE 1995). The Syntominae are small-to-medium sized moths (SCOBLE 1995) and include 977 species, none in the New World (HEPPNER 1991).
In the present study we assessed Arctiidae richness in the Brazilian Cerrado biome for the first time. Specifically, we 1) map and describe the species-richness in the biome, as currently known, 2) highlight areas where these moths are poorly known, and 3) investigate the spatial relationship in similarities among the 14 best-sampled regions.
MATERIAL AND METHODS
The Cerrado region, located in altitudes ranging from 300 to 900 m, has a high diversity of soils, geomorphology, climate, and vegetation types (DURIGAN et al. 2003, SILVA et al. 2006). Nearly 95% of the climate in this biome is classified as Aw, according to the Köppen system, with an annual temperature average between 18 and 28°C, and precipitation ranging from 600 to 2,200 mm per year. Moreover, the climate is highly seasonal, so that 80% of the biome has five to seven months of dry season during the winter (DIAS 1996). The soils in 90% of the region are nutrient-poor, highly acidic, and aluminum-rich latosols and quartzsols. The flora is very heterogeneous, presenting a gradient that ranges from evergreen forest with closed canopy (gallery forest) to a total absence of woody vegetation ("campo limpo") (DURIGAN et al. 2003, SILVA et al. 2006).
Most of our data on the the geographic distribution of the Cerrado tiger moths were obtained from specimens deposited in Brazilian museums, plus a few records from the literature. We visited the 10 most important arctiid collections in the country: Coleção Becker, Coleção Entomológica da Universidade de Brasília, Coleção Entomológica do Instituto Nacional de Pesquisas da Amazônia, Coleção Entomológica Padre Jesus Santiago Moure da Universidade Federal do Paraná, Fundação Instituto Oswaldo Cruz, Museu Paraense Emilio Goeldi, Museu Entomológico Ceslau Biezanko da Universidade Federal de Pelotas, Museu de Zoologia da Universidade Estadual de Campinas Adão José Cardoso, Museu de Zoologia da Universidade de São Paulo, and Museu Nacional da Universida-de Federal do Rio de Janeiro. The individuals were identified using information available in the literature (PIÑAS RUBIO et al. 2000, PIÑAS RUBIO & MANZANO 2003, WATSON & GOODGER 1986) and by comparison with digital images of identified specimens in the Coleção Becker. This is the best collection of arctiid of the Cerrado and most of its specimens were identified by comparison with types found in museums. Additionally, BENDIB & MINET (1999) was used to identify the Lithosiinae tribes.
Our data set contained only species that could be identified with certainty by using the adult external morphology (head and leg characters, wing venation, and coloration). We did not consider individuals with doubtful provenance. Individuals of the same species collected on multiple dates at the same locality were considered as a single record. Most of the museum specimens did not contain information on the geographic coordinates of the sampled site. In such cases, we used the coordinates of the nearest city where the specimen was collected, obtained from the "splink" information system available at the Reference Center on Environmental Information (http://splink.cria.org.br/geoloc). We also obtained data on species records from literature (FERREIRA et al. 1995, FERRO & DINIZ 2007a), but did not include records when information about the sampling sites was innacurate and a few taxonomical and ecological publications dealing with the fauna of eucalyptus plantations.
The Cerrado biome boundary was defined according to the coordinates available from the INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA (2008). We used a grid containing one-degree latitude/longitude cells to map tiger moth richness in the core region of the Brazilian Cerrado. Most species were recorded in one or two localities only, precluding the use of minimum convex polygons of occurrence to estimate species richness in cells.
We selected the 14 best-sampled cells (i.e., those containing more than 61 species). These 14 cells represent a compromise between the minimum sampling effort in the cells and the maximum number of cells to evaluate resemblance among regions. We used the ordination and Mantel tests to evaluate whether similarities in species composition were related to spatial distance. We calculated a dissimilarity matrix using the Simpson beta-diversity index (KOLEFF et al. 2003a), which is related to Jaccard and Sorensen indices, but has less sensitivity to sample size (KOLEFF et al. 2003a, BASELGA 2007). In our study, the 14 best-sampled cells differed greatly in species richness and this was the main reason for selecting the Simpson beta-diversity index. We also used the dissimilarity matrix to obtain the ordination of the 14 cells on two axes using a Non-Metric Multidimensional Scaling (NMDS).
Additionally, we employed Mantel tests to assess whether dissimilarities in species composition among cells were related to: 1) geographical distances, 2) differences in latitude, and 3) differences in longitude among cells. We used the same dissimilarity matrix used in the NMDS ordination. The Mantel tests were done using the Spearman coefficient of correlation and 1,000 permutations to assess statistical significance. The choice for the Spearman coefficient was based on previous work indicating the non-linearity of the distance decay relationship (NEKOLA & WHITE 1999). We wrote routines and used functions available in the packages 'MASS' (VENABLES & RIPLEY 2002) and 'vegan' (OKSANEN et al. 2008) of the R environment (R DEVELOPMENT CORE TEAM 2007) to data analyses.
RESULTS
About 30% of the specimens examined were not included in our data set due to doubtful provenance or difficulties in sorting among morphospecies (hereafter treated as 'species'). A total of 16 species could not be identified to genus, and 14 could not be identified to species. A portion of this unidentified material likely represents new species or genera. The remaining specimens might correspond to species already described but for which we were not able to identify. The data set included a total of 2,321 records belonging to 723 species, and to 256 genera. Out of the 723 species, 621 belonged to the Arctiinae (86%). Although all six Arctiinae tribes occur in the Cerrado region, the Phaegopterini and Ctenuchini were dominant, comprising 53.5% of the total arctiid species (Tab. I). The remaining 102 species were Lithosiinae, and belonged mostly to the tribes Cisthenini and Lithosiini (Tab. I).
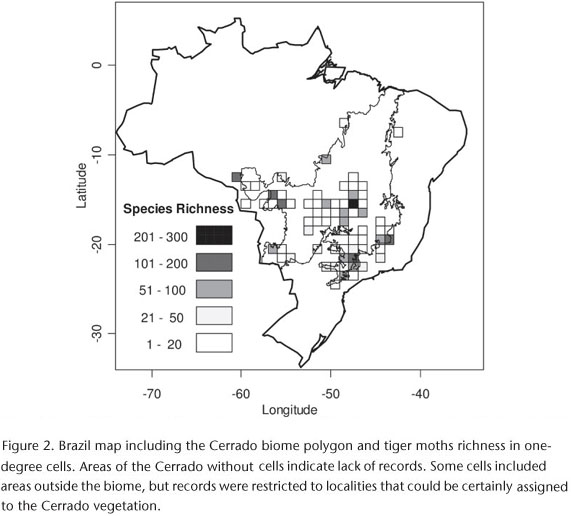
Species were recorded in 108 localities of the Cerrado distributed in 67 one-degree cells. The number of species sampled exclusively in one and two cells was 315 and 146, respectively. Collectively, they represented 64% of all species (Fig. 1). Only eight species were recorded in more than 15 cells (Fig. 1).
Our sampling effort was not homogeneously distributed in the biome (Fig. 2). For instance, we do not have records for vast regions of the northern part of the Cerrado. Most records were concentrated in the southeastern portion (Fig. 2), which is the most populated region and is close to the largest Brazilian museums and universities. The cell containing the most records (261) and species (239) included Brasília (capital of Brazil), in the central part of the biome.
The ordination of the 14 best-sampled cells containing more than 61 species revealed that nearby areas harbor similar sets of species (Fig. 3). Also, a gradient was found in the first axis, with western and northern cells scored in the right side of the diagram (Fig. 3). Consistent with the NMDS ordination, the Mantel tests revealed that dissimilarities in species composition were strongly correlated with geographical distances (r = 0.71, p < 0.001). The main component contributing to this strong correlation was longitude (r = 0.56, p < 0.001), although the correlation with latitude was also relatively high (r = 0.44, p < 0.001).
DISCUSSION
The richness of arctiid recorded in this study for the Cerrado represents 36% of the 2,000 species estimated to occur in Brazil according to BROWN JR & FREITAS (1999), and 12% of the 5,931 Neotropical species (HEPPNER 1991). Similarly, CAMARGO & BECKER (1999) observed that Saturniidae moths in the Cerrado represented 42% of the Brazilian and 12% of the New World fauna. Despite the similarity found between our data and CAMARGO & BECKER'S (1999), it is not yet clear whether the relative proportions of Brazilian and Neotropical arctiid and saturniid moths occurring in the Cerrado can be generalized to other lepidopterans. There is no published information on other moth families and our data indicate that many arctiid species may be found in future inventories of the northern Cerrado.
The sampling effort was not homogenously distributed in the Cerrado area. Instead, museum records were concentrated in the southern part of the biome, and there were several areas in the biome with no or few species records, especially in the northeastern region. For instance, there were no records in the Cerrado regions of the states of Bahia and Maranhão. In the states of Piauí and Tocantins there were records from only one and three sites, respectively. The southern portion of the Cerrado, which includes most of the arctiid records, also includes regions that are poorly inventoried. For instance, there are only a few records in Cerrado areas of the state of Mato Grosso do Sul, and in the southeastern portion of the biome (borders of São Paulo and Paraná states). These poorly inventoried regions are in the most disturbed part of the biome. In the state of São Paulo, for example, the vegetation has been reduced from 33,929 km2 to 2,379 km2 in the last 30 years (DURIGAN et al. 2003). The northeastern region of the biome, for which only a few records are available, includes the lesser-disturbed areas of Cerrado. However, this area is currently threatened by the expansion of the agricultural frontier and subsequent development of infrastructures (CAVALCANTI & JOLY 2002). Future collections of arctiid in the northern portion of the Cerrado should increase the total species richness in the biome. Additionally, these inventories may allow better estimates of species ranges and evaluation of endemism in the biome.
The cell containing most species and records was located in the central portion of the Cerrado (Brasília) and included 33% of all species recorded in the entire biome. BRIDGEWATER et al. (2004) found similar results for wood plants and reported that 25% of all Cerrado plants were recorded in 13 sites around Brasília. The region of Brasília also harbors the highest known richness of Mimosa L. legumes (SIMON & PROENÇA 2000) and anurans (DINIZ-FILHO et al. 2007) in the Cerrado. This repeated pattern of highest richness around Brasília is probably due to concentrated sampling effort in this region, where the largest and oldest university (Universidade de Brasília) of the core region of the Cerrado is located. Tiger moth species richness in the Brasília region (239 species) is only slightly lower than that observed in well-inventoried Neotropical moist sites. FERRO & DINIZ (2007b), for instance, compiled about 4,800 museum records accumulated during decades of arctiid (mostly Arctiinae) sampling at Estação Biológica de Boracéia, a southeastern Brazilian Atlantic Rain Forest site, and found 256 species. Moreover, ZIKÁN & ZIKÁN (1968) listed 306 currently valid arctiid species in a high-altitude site in the Atlantic forest, and HILT & FIEDLER (2005) found 287 species in an inventory of a 2 km2 area of montane cloud forest in Ecuador. Together, these values indicate that local species richness in the Cerrado is similar to those in moist forests. This result does not imply, however, that total (regional) species richness in the Cerrado is similar to values observed in other biomes. Too few localities were well inventoried in the Cerrado and many areas remain mostly unsampled, precluding a reliable estimation of total diversity.
The multivariate analyses of species composition in the 14 best-sampled cells revealed that those cells closest to one another share more similar sets of species than those localities that are farther appart, a pattern that had been previously reported for other groups of organisms, such as plants and parasite communities of vertebrate hosts (NEKOLA & WHITE 1999, POULIN 2003). This trend, known as distance decay of similarity, can be caused by differences in environmental conditions or by the dispersal restrictions of certain organisms (NEKOLA & WHITE 1999, BECK & KHEN 2007). Unfortunately, the limited number of cells used in our analyses preclude our ability to draw conclusions on the role played by these two factors in our data. However, the results of our study, carried out in an area encompassing 17 degrees of longitude and 13 degrees of latitude, are consistent with the expectation that distance decay patterns are most likely to be revealed by large-scale studies (NEKOLA & WHITE 1999).
Given the existence of latitudinal gradients in species richness and environmental conditions (e.g., HAWKINS et al. 2003), one might expect that the slope of the distance decay relationship would be steepest along latitudinal distances (see KOLEFF et al. 2003b). Instead, we observed steepest decreasing similarity along longitudinal distances and this gradient deserves further investigation in other taxonomic groups. RATTER et al. (2003), for instance, identified five floristic regions within the core region of the Cerrado arranged both along the latitudinal and longitudinal gradients. In the east portion of the biome they identified three provinces (namelly North & Northeastern, Central & Southeastern, and Southern) and, in the western portion, they identified two provinces (namelly Central & Western and Far Western Mesotrophic). Thus, the spatial gradient observed in our arctiid data should be a consequence of differences in host-plant composition between the floristic regions in the Cerrado (RATTER et al. 2003).
How much do we know about Arctiidae richness in the Cerrado? The biome includes an expressive proportion of the richness in Brazil and in the Neotropical region. The identification of many regions without records and the high proportion of rare species indicate that many more species should be found in further inventories. The local richness of the most intensively sampling region (Brasília) was similar to values from intensively sampled sites of moist forests in the Neotropical region. Lastly, we found that the resemblance in species composition among the best-sampled cells followed a pattern of distance decay in similarity, particularly along the longitudinal gradient. The importance of this longitudinal gradient deserves further studies to test, for instance, whether arctiid assemblages in the Cerrado at large spatial scales is determined by the distribution of their host plants.
ACKNOWLEDGEMENTS
We would like to give special thanks to Vitor O. Becker for his friendly hospitality on several occasions, providing us with access to his moth collection and library, as well as training the first author in identifying the Arctiidae. We are also greatly indebted to museum curators for allowing VGF's access to the collections and for logistic support. Additionally we would like to thank the Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA), Instituto Florestal de São Paulo (IF-SP), and Agência Ambiental de Goiás for allowing us to gather samples in protected areas and providing logistic support. The first author received a student fellowship from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (proc. 141715/2005-7). The second author received research grants (476256/2004-6, 476304/2007-5) and a research fellowship (302482/2008-3) from CNPq, and a research grant from International Foundation for Science (A/4107-1). The third author received research grants from CNPq (303992/2004-2; 472211/2003-0).
LITERATURE CITED
Submitted: 12.XI.2009; Accepted: 27.VIII.2010.
Editorial responsibility: Glauco Machado
- BASELGA, A. 2007. Disentangling distance decay of similarity from richness gradients: response to Soininen et al. 2007. Ecography 30 (6): 838-841.
- BECK, J. & C.V. KHEN. 2007. Beta-diversity of geometrid moths from northern Borneo: effects of habitat, time and space. Journal of Animal Ecology 76 (2): 230-237.
- BENDIB, A. & J. MINET. 1999. Lithosiinae main lineages and their possible interrrelationships. I. Definition of new or resurrected tribes (Lepidoptera: Arctiidae). Annals de la Societé Entomologique de France 35 (3-4): 241-263.
- BRIDGEWATER, S.; J.A. RATTER & J.F. RIBEIRO. 2004. Biogeographic patterns, b-diversity and dominance in the cerrado biome of Brazil. Biodiversity and Conservation 13 (12): 2295-2318.
- BROWN JR, K.S. & A.V.L. FREITAS. 1999. Lepidoptera, p. 225-243. In: C.R.F. BRANDÃO & E.M. CANCELLO (Eds.). Biodiversidade do Estado de São Paulo: síntese do conhecimento ao final do século XX - Invertebrados terrestres. São Paulo, FAPESP, 279p.
- CAMARGO, A.J.A. & V.O. BECKER. 1999. Saturniidae (Lepidoptera) from the Brazilian Cerrado: composition and biogeographic relationships. Biotropica 31 (4): 696-705.
- CAVALCANTI, R.B. & C.A. JOLY. 2002. Biodiversity and conservation priorities in the Cerrado region, p. 351-367. In: P.S. OLIVEIRA & R.J. MARQUIS (Eds). The Cerrados of Brazil: Ecology and natural history of a neotropical savanna. New York, Columbia University Press, 398p.
- DEVRIES, P.J.; D. MURRAY & R. LANDE. 1997. Species diversity in vertical, horizontal, and temporal dimensions of a fruit-feeding butterfly community in an Ecuadorian rainforest. Biological Journal of the Linnean Society 62 (3): 343-364.
- DIAS, B.F.S. 1996. Cerrados: uma caracterização, p. 11-25. In: B.F.S. DIAS (Ed.). Alternativas de Desenvolvimento dos Cerrados: Manejo e Conservação dos Recursos Naturais Renováveis. Brasília, Funatura, 97p.
- DINIZ-FILHO, J.A.F.; L.M. BINI; M.P. PINTO; T.F.L.V.B. RANGEL; P. CARVALHO; S.L. VIEIRA & R.P. BASTOS. 2007. Conservation biogeography of anurans in Brazilian Cerrado. Biodiversity and Conservation 16 (4): 997-1008.
- DURIGAN, G.; M.F. SIQUEIRA; G.A.D.C. FRANCO; S. BRIDGEWATER & J.A. RATTER. 2003. The vegetation of priority areas for Cerrado conservation in São Paulo state, Brazil. Edinburgh Journal of Botany 60 (2): 217:241.
- FERREIRA, P.S.F.; A.S. PAULA & D.S. MARTINS. 1995. Análise faunística de Lepidoptera Arctiidae em área de reserva natural remanescente de floresta tropical em Viçosa, Minas Gerais. Anais da Sociedade Entomológica do Brasil 24 (1): 123-133.
- FERRO, V.G. & I.R. DINIZ. 2007a. Composição de espécies de Arctiidae (Lepidoptera) em áreas de Cerrado. Revista Brasileira de Zoologia 24 (3): 635-646.
- FERRO, V.G. & I.R. DINIZ. 2007b. Arctiidae (Insecta: Lepidoptera) da Estação Biológica de Boracéia (Salesópolis, São Paulo, Brasil). Biota Neotropica 7 (3). Avaliable online at: http://www.biotaneotropica.org.br/v7n3/pt/fullpaper?bn03107032007+pt [Accessed: 5.XI.2009]
- FERRO, V.G. & I.R. DINIZ. 2008. Species biological attributes affecting the description date of tiger moths (Arctiidae) in the Brazilian Cerrado. Diversity and Distributions 14 (3): 472-482.
- FUNDAÇÃO BIODIVERSITAS. 2008. Lista da fauna brasileira ameaçada de extinção. Avaliable online at: http://www.biodiversitas.org.br/f_ameaca/ [Accessed: 5.XI.2009]
- HAWKINS, B.A.; E.E. PORTER & J.A.F. DINIZ-FILHO. 2003. Productivity and history as predictors of the latitudinal diversity gradient of terrestrial birds. Ecology 84 (6): 1608-1623.
- HEPPNER, J.B. 1991. Faunal regions and the diversity of Lepidoptera. Tropical Lepidoptera 2 (Suppl. 1): 1-85.
- HILT, N. & K. FIEDLER. 2005. Diversity and composition of Arctiidae moth ensembles along a sucessional gradient in the Ecuadorian Andes. Diversity and Distributions 11 (5): 387-398.
- HILTY, J. & A. MERENLENDER. 2000. Faunal indicator taxa selection for monitoring ecosystem health. Biological Conservation 92 (2): 185-197.
- INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. 2008. Mapa de Biomas. Available online at: http://mapas.ibge.gov.br/biomas2/viewer.htm [Accessed: 5.XI.2009]
- JACOBSON, N.L. & S.J. WELLER. 2002. A cladistic study of the Arctiidae (Lepidoptera) by using characters of immatures and adults. Lanham, Entomological Society of America, Thomas Say Publication in Entomology: Monographs, 98p.
- KITCHING, R.L.; A.G. ORR; L. THALIB; H. MITCHELL; M.S. HOPKINS & A.W. GRAHAM. 2000. Moths assemblages as indicators of environmental quality in remnants of upland Australian rain forest. Journal of Applied Ecology 37 (2): 284-297.
- KLINK, C.A. & R.B. MACHADO. 2005. Conservation of the Brazilian Cerrado. Conservation Biology 19 (3): 707-713.
- KOLEFF, P.; K.J. GASTON & J.J. LENNON. 2003a. Measuring beta diversity for presence-absence data. Journal of Animal Ecology 72 (3): 367-382.
- KOLEFF, P.; J.J. LENNON & K.J. GASTON. 2003b. Are there latitudinal gradients in species turnover? Global Ecology and Biogeography 12 (6): 483-498.
- MYERS, N.; R.A. MITTERMEIER; C.G. MITTERMEIER; G.A.B. FONSECA & J. KENT. 2000. Biodiversity hotspots for conservation priorities. Nature 403 (6772): 853-858.
- NEKOLA, F.C. & P.S. WHITE. 1999. The distance decay of similarity in biogeography and ecology. Journal of Biogeography 26 (4): 867-878.
- OKSANEN, J.; R. KINDT; P. LEGENDRE; R. O'HARA; G.L. SIMPSON & M.H.H. STEVENS. 2008. Vegan: Community Ecology Package. R package version 1.13-0. Available online at: http://vegan.r-forge.r-project.org [Accessed: 5.XI.2009]
- OLIVEIRA-FILHO, A.T. & J.A. RATTER. 1995. A study of the origin of central Brazilian forests by the analysis of plant species distribution patterns. Edinburgh Journal of Botany 52 (2): 141-194.
- PIÑAS RUBIO, F.S.J.; S. RAAB-GREEN; G. ONORE & I. MANZANO. 2000. Mariposas del Ecuador. Family Arctiidae. Quito, Pontificia Universidad Católica del Ecuador, vol. 20, 32+84p.
- PIÑAS RUBIO, F.S.J. & I. MANZANO. 2003. Mariposas del Ecuador. Arctiidae, subfamilia Ctenuchinae. Quito, Compañia de Jesús, vol. 21b, 95p.
- POULIN, R. 2003. The decay of similarity with geographical distance in parasite communities of vertebrate hosts. Journal of Biogeography 30 (10): 1609-1615.
- R DEVELOPMENT CORE TEAM. 2007. R: A language and environment for statistical computing. Vienna, R Foundation for Statistical Computing. Available online at: http://www.R-project.org [Accessed: 5.XI.2009]
- RATTER, J.A.; J.F. RIBEIRO & S. BRIDGEWATER. 1997. The Brazilian cerrado vegetation and threats to its biodiversity. Annals of Botany 80 (3): 223-230.
- RATTER, J.A.; S. BRIDGEWATER & J.F. RIBEIRO. 2003. Analysis of the floristic composition of the Brazilian cerrado vegetation III: Comparison of the woody vegetation of 376 areas. Edinburgh Journal of Botany 60 (1): 57-109.
- SCOBLE, M.J. 1995. The Lepidoptera: form, function and diversity. New York, Oxford University Press, 404p.
- SILVA, J.F.; M.R. FARIÑAS; J.M. FELFILI & C.A. KLINK. 2006. Spatial heterogeneity, land use and conservation in the cerrado region of Brazil. Journal of Biogeography 33 (3): 536-548.
- SILVA, J.M.C. & M.P.D. SANTOS. 2005. A importância relativa dos processos biogeográficos na formação da avifauna do Cerrado e de outros biomas brasileiros, p. 219-233. In: A. SCARIOT; J.C. SOUZA-SILVA & J.M. FELFILI (Eds). Cerrado: ecologia, biodiversida-de e conservação. Brasília, Ministério do Meio Ambiente, 439p.
- SIMON, M.F. & C. PROENÇA. 2000. Phytogeographic patterns of Mimosa (Mimosoideae, Leguminosae) in the Cerrado biome of Brazil: an indicator genus of high-altitude centers of endemism? Biological Conservation 96 (3): 279-296.
- VENABLES, W.N. & B.D. RIPLEY. 2002. Modern Applied Statistics with S. New York, Springer, 495p.
- ZIKÁN, J.F. & W. ZIKÁN. 1968. Inseto-fauna da Itatiaia e da Mantiqueira 3: Lepidoptera. Pesquisa Agropecuária Brasileira 3 (1): 45-109.
- WATSON, A. & D.T. GOODGER. 1986. Catalogue of the neotropical tiger-moths. Ocasional Papers on Systematic Entomology, British Museum (National History) 1 (1): 1-71.
Publication Dates
-
Publication in this collection
23 Nov 2010 -
Date of issue
Oct 2010
History
-
Received
12 Nov 2009 -
Accepted
27 Aug 2010