Abstract
The present study analyzed qualitatively and quantitatively the temporal and spatial changes in the fish assemblages present in seabob shrimp distribution sites in two important fishing areas of the Santa Catarina coast. Monthly bottom trawls, conducted between March 2007 and February 2008 in Balneário Barra do Sul and Penha, yielded a total of 38,608 specimens belonging to 54 species, of which 35 occurred in both regions. The Sciaenidae was the most represented family in both number of species and abundance of individuals. The dominant species were Stellifer rastrifer (Jordan, 1889), S. brasiliensis (Schultz, 1945), Paralonchurus brasiliensis (Steindachner, 1875), Isopisthus parvipinnis (Cuvier, 1830), Trichiurus lepturus (Linnaeus, 1758), and Pellona harroweri (Fowler, 1917). The ichthyofaunal structure showed a spatiotemporal dependence with significant differences in March 2007 and January and February 2008 between Balneário Barra do Sul and Penha. The main differences were attributed to dominant species, which were responsible for significantly altering the abundance of the assemblages. The species richness was similar in both sites, whereas the diversity and equitability indices showed monthly fluctuation patterns that differed between Balneário Barra do Sul and Penha. The Shannon index differed in August, September and November 2007 and January and February 2008, with an H' ranging from 0.74 to 1.98 influenced by dominant species, whereas significant variations were observed for the Pielou index in November 2007 and February 2008. The Berger-Parker index demonstrated a clear dominance pattern where the monthly numeric contribution of a single species ranged from approximately 30 to 80%. Although the results showed differences between fishing grounds 25% of the time, we conclude that fish assemblies captured as accompanying fauna in artisanal seabob shrimp fisheries showed statistically similar characteristics for most of the year, although Balneário Barra do Sul and Penha are 35 km away from each other.
Artisanal fishing; assemblage structure; bycatch; demersal fish
AQUACULTURE AND FISHERIES
Spatiotemporal variations of the ichthyofaunal structure accompanying the seabob shrimp, Xiphopenaeus kroyeri (Crustacea: Penaeidae), fishery in important fishery areas of the Santa Catarina shore, Brazil
Jurandir J. Bernardes JúniorI* * Corresponding author. E-mail: jurandirjbjr@gmail.com ; Jorge L. Rodrigues FilhoII; Joaquim O. BrancoIII; José R. VeraniII
IPrograma de Pós-Graduação em Aquicultura, Centro de Ciências Agrárias, Departamento de Aquicultura, Universidade Federal de Santa Catarina. Rodovia Admar Gonzaga 1346, 88034-001 Florianópolis, SC, Brazil
IIDepartamento de Hidrobiologia, Universidade Federal de São Carlos. Rodovia Washington Luis, km 235, 13565-905 São Carlos, SP, Brazil
IIILaboratório de Biologia, Centro de Ciências Tecnológicas da Terra e do Mar, Universidade do Vale do Itajaí. Rua Uruguai 458, Caixa Postal 360, 88302-202 Itajaí, SC, Brazil
ABSTRACT
The present study analyzed qualitatively and quantitatively the temporal and spatial changes in the fish assemblages present in seabob shrimp distribution sites in two important fishing areas of the Santa Catarina coast. Monthly bottom trawls, conducted between March 2007 and February 2008 in Balneário Barra do Sul and Penha, yielded a total of 38,608 specimens belonging to 54 species, of which 35 occurred in both regions. The Sciaenidae was the most represented family in both number of species and abundance of individuals. The dominant species were Stellifer rastrifer (Jordan, 1889), S. brasiliensis (Schultz, 1945), Paralonchurus brasiliensis (Steindachner, 1875), Isopisthus parvipinnis (Cuvier, 1830), Trichiurus lepturus (Linnaeus, 1758), and Pellona harroweri (Fowler, 1917). The ichthyofaunal structure showed a spatiotemporal dependence with significant differences in March 2007 and January and February 2008 between Balneário Barra do Sul and Penha. The main differences were attributed to dominant species, which were responsible for significantly altering the abundance of the assemblages. The species richness was similar in both sites, whereas the diversity and equitability indices showed monthly fluctuation patterns that differed between Balneário Barra do Sul and Penha. The Shannon index differed in August, September and November 2007 and January and February 2008, with an H' ranging from 0.74 to 1.98 influenced by dominant species, whereas significant variations were observed for the Pielou index in November 2007 and February 2008. The Berger-Parker index demonstrated a clear dominance pattern where the monthly numeric contribution of a single species ranged from approximately 30 to 80%. Although the results showed differences between fishing grounds 25% of the time, we conclude that fish assemblies captured as accompanying fauna in artisanal seabob shrimp fisheries showed statistically similar characteristics for most of the year, although Balneário Barra do Sul and Penha are 35 km away from each other.
Key words: Artisanal fishing; assemblage structure; bycatch; demersal fish
Trawling for shrimp is an efficient method to catch target species, but it also accidentally captures a great variety of organisms called accompanying fauna or bycatch (Alverson et al. 1994). There is a consensus that shrimp trawling is responsible for the highest rates of incidental catches of demersal organisms (Andrew & Pepperell 1992, Kelleher 2005, Gillett 2008). In tropical regions, the accompanying fauna associated with penaeid shrimp fisheries represents approximately 68% of the collected biomass (Kelleher 2005), and fish comprise over 25% of this biomass (Haimovici & Mendonça 1996).
Artisanal fleets contribute a significant share to the total crustacean production in the shallow waters of the Brazilian continental shelf. For instance, in 2004 alone, they caught approximately seven thousand tons of biomass (MMA/IBAMA 2005). It is estimated that, on average, for every kilogram of shrimp, between 1 and 12 kilograms of fish are incidentally caught (Ruffino & Castello 1992, Vianna & Almeida 2005, Branco & Verani 2006b, Souza et al. 2008). In most regions where shrimp trawling is practiced, a tiny portion of the bycatch is consumed (fish of low commercial value), and the remainder is discarded (Alverson et al. 1994, Ambrose et al. 2005, Bail & Branco 2007).
Understanding the impact of shrimp trawling on marine resources is fundamental for maintaining the sustainability and balance of coastal ecosystems and the trophic chain of the exploited areas (Pitcher & Hart 1982, Ricklefs & Miller 2000, Dayton et al. 2002). In the last decades, the composition, diversity and spatiotemporal variations of ichthyofauna have been the focus of a number of studies along the Brazilian coast (Coelho et al. 1986, Paiva-Filho & Schmiegelow 1986, Ruffino & Castello 1992, Santos et al. 1998, Santos 2000, Tischer & Santos 2001, Graça Lopes et al. 2002, Branco & Verani 2006a, b, Gomes & Chaves 2006, Schwarz Jr et al. 2007), but only a few studies have compared among fishing grounds (Chaves et al. 2003, Souza et al. 2008).
With a 531 km shoreline spanning 337 localities where artisanal fisheries and 17 fishermen communities occur, the state of Santa Catarina is the greatest producer of seabob shrimp, Xiphopenaeus kroyeri Heller, 1862, of the southern region (PROZEE 2006). This activity represents an important socioeconomic contribution to the north-central coast, especially in the municipalities of Penha and Balneário Barra do Sul, which are approximately 35 km from each other.
Therefore, due to the socioeconomic and ecological relevance of shrimp trawling, this study aimed to qualitatively and quantitatively assess the spatial and temporal variations in the fish community captured as accompanying fauna in seabob shrimp fishery at two important fishing areas of the Santa Catarina coast.
MATERIAL AND METHODS
Collections were conducted monthly from March 2007 to February 2008 in the Balneário Barra do Sul region (municipalities of Balneário Barra do Sul and São Francisco do Sul, 26º23'-26º28'S and 48º30'-48º35'W) and in the Armação do Itapocoroy region (municipality of Penha, 26º40'-26º47'S and 48º36'-48º38'W) (Fig. 1). Balneário Barra do Sul and Penha are located on the north central coast of the state of Santa Catarina, a region characterized by a mesothermal humid climate, and rainfall and average annual temperature of 1,690 mm and 20.2ºC, respectively (Araújo et al. 2006b). Close to the coastal region, predominate the Coastal Water mass which has salinity below 34 and temperature ranging from 19 to 28ºC in winter and summer, respectively (Schettini et al. 1999). For the majority of the year, winds are predominantly from the northeast, changing periodically to southerly winds with advances of the polar frontal systems, which are more frequent and intense in winter (Schettini et al. 1999, Araújo et al. 2006b). The fishing grounds are composed predominantly of sandy facies varying from very fine to medium sand (Corrêa et al. 1996, Abreu et al. 2006), but considerably wide layers of fine sediments (silt and clay) also occur parallel to the coast (Horn Filho 2003).
Each month in the morning, we conducted three fishing hauls (samples) at each of the two regions mentioned above. Sampling took place at the traditional seabob shrimp fishing areas, for one hour, at depths ranging from 5 to 25 m. In each region, three fishing hauls (samples) were performed in the traditional seabob shrimp fishing areas for one hour each during the morning period, at depths from 5 to 25 m. Although it is mentioned in the literature that depth can influence species composition (Chaves et al. 2003, Souza et al. 2008), no differences were observed in the assemblies caught between 5 and 25 meters in previous studies conducted in the Penha region (Bail & Branco 2003, Branco & Verani 2006a); therefore, we adopted trawls among these isobaths to compose representative samples of the regions. A double-rig shrimp vessel equipped with two bottom trawls with 3 cm mesh in the sleeve and 2 cm mesh in the trawl bag, was used for trawling, conducted at an average speed of two knots (Branco 2005). The contents of the nets were packed in plastic bags, tagged and kept in a styrofoam cold box with ice. Fish were identified at the species level (Figueiredo 1977, 1980, Soares 1978, Figueiredo & Menezes 1978, Menezes & Figueiredo 1980, 1985), and the abundance of each species was recorded.
Each species was classified into one of three categories according to its frequency of occurrence in samples, as follows: constant (present in samples between 9 to 12 months), accessory (6 to 8 months) or incidental (1 to 5 months) (Severino-Rodrigues et al. 2002).
For characterization of the fish assemblages, sample-based rarefaction curves scaled to the number of individuals (Colwell et al. 2004) were constructed and the Shannon diversity (H') index, Pielou equitability (J') index (Magurran 2004) and CPUE (fish per hour) were calculated. These indices and CPUE were evaluated for normality by a Kolmogorov-Smirnov test and for homogeneity of variances by the Bartlett test and monthly compared between Balneário Barra do Sul and Penha through a bifactorial analysis of variance (ANOVA) at a significance level of 0.05 (Zar, 1996, Underwood 1997). The CPUE data were fourth root-transformed to meet the assumptions of analysis. When an interaction effect was significant, Tukey's test was performed at a significance level of 0.05 (Zar 1996). Concomitantly, the Berger-Parker index (d) was used to measure the temporal dominance in each region (Magurran 2004).
To visualize similarity patterns between samples, non-metric Multidimensional Scaling (nMDS) of similarities was carried out using a proximity analysis (Clarke & Warwick 1994) where the Bray-Curtis index was applied to the abundance matrix transformed by the fourth root. From this analysis, a conformity index (Stress) was obtained, determining the degree of proximity of the graphic representation and of real data. Stress values under 0.20 allow for the potential use of two MDS dimensions. However, values above the limit of this interval reduce confidence in interpreting the details of the graphic representation, suggesting superposition of clusters as a supplement to the analysis (Clarke & Warwick 1994). The superposition of clusters, calculated from the Bray-Curtis similarity index and grouped by the UPGMA method, was used at the cut-off level of 60% as an alternative to confirm the proximity patterns of samples.
A space-time differences hypothesis of the ichthyofaunal structure was tested through a non-parametric multivariate analysis of variance (PERMANOVA) at a 0.05 significance level (Anderson 2001, 2005). For this purpose, the design consisted of two orthogonal factors, "Sites" (two levels, random) and "Months" (twelve levels, random and crossed with Sites), as well as the interaction between them. In this analysis, the Bray-Curtis index was calculated from the ichthyofauna abundance matrix transformed by the fourth root for the purpose of increasing the relative contribution of rare species (Clarke et al. 2006). For all tests, a subset of 9,999 permutations was used. When statistical differences were observed, an a posteriori pair-wise multivariate t-test was applied (Anderson 2005). The specific contribution of each fish species to the monthly dissimilarity between sites was evaluated by the Similarity Percentage test (SIMPER) (Clarke & Warwick 1994).
RESULTS
A total of 21,482 fish were collected in Balneário Barra do Sul (BS) belonging to 46 species, 40 genera and 21 families, whereas in Penha (P), 17,126 specimens were collected belonging to 43 species, 38 genera and 24 families (Tab. I). A total of 38,608 fish were sampled belonging to 54 species, of which 35 were common to the two regions.
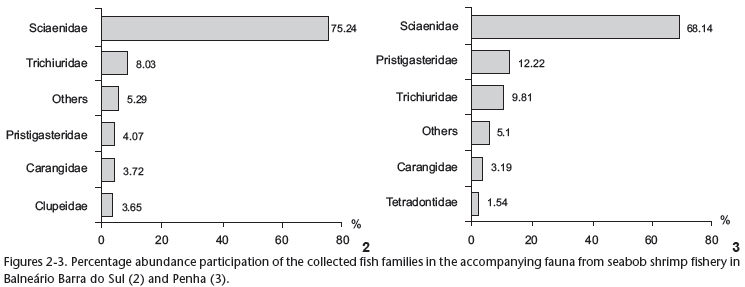
Sciaenidae, Trichiuridae, Carangidae and Pristigasteridae represented 91.06% of the abundance in the collections in Balneário Barra do Sul (Fig. 2), and 93.36% in Penha (Fig. 3). The number of different species collected was greatest within Sciaenidae (13 spp. in BS and 11 in P) followed by Carangidae (5 spp. in BS and 4 in P) and Tetraodontidae (3 spp. in each site).
According to the categories of occurrence, the species that were considered constant were also more abundant (86% in BS and 64% in P), with the occurrence of nine species in Balneário Barra do Sul belonging to the Sciaenidae, Pristigasteridae, Carangidae and Trichiuridae families and seven species in Penha belonging to the Sciaenidae, Pristigasteridae, Carangidae and Synoglossidae families (Tab. I). Approximately 60% of the species collected (30 spp. in BS and 27 spp. in P) were classified as incidental, and represented the lowest contributions to the total number of specimens. In the accessory category, seven species were present in Balneário Barra do Sul and nine in Penha (Tab. I), representing 5 and 30% of the species abundance, respectively.
The multivariate analysis (PERMANOVA) of the ichthyofaunal structure showed significant differences between sites and months, and a significant interaction between these factors, indicating that the temporal variation was distinct between Balneário Barra do Sul and Penha (Tab. II). The a posteriori pair-wise test showed that the species composition and abundance of individuals differed significantly in March 2007, and January and February 2008.
The dispersion pattern of samples in the nMDS plot showed the spatiotemporal dependence of ichthyofaunal structure (Fig. 4) and, despite the stress value being moderately high (0.17), enabled the classification of the data into three groups: group I consisted of samples collected in Balneário Barra do Sul and Penha, and was defined based on the high relative abundance values of Isopisthus parvipinnis (Cuvier, 1830), Pellona harroweri (Fowler, 1917), Selene setapinnis (Mitchill, 1815), Stellifer rastrifer (Jordan, 1889), Stellifer brasiliensis (Schultz, 1945) and Trichiurus lepturus (Linnaeus, 1758), and moderate relative abundance values of Lagocephalus laevigatus (Linnaeus, 1766) and Larimus breviceps (Cuvier, 1830); group II associated the months of August, September and October (P), and September 2007 (BS), and was characterized by the important numeric contribution of Urophycis brasiliensis (Günther, 1880) and intermediate numbers of Paralonchurus brasiliensis (Steindachner, 1875) and T. lepturus; group III, despite the close proximity between samples, represented the collections that occurred in Penha, highlighted by Stellifer, Lycengraulis grossidens Agassiz (1829), and L. breviceps and intermediate abundance values of I. parvipinnis, P. harroweri and T. lepturus.
The months of November 2007, February 2008 (P) and March 2007 (BS) were analyzed separately due to particularities in the composition and abundance of their species assemblies when compared to the other groups. In Penha, high relative abundances were recorded for Chirocentrodon bleekerianus (Poey, 1867) and Ctenosciaena gracilicirrhus (Metzelar, 1919) in the sampling conducted in November, whereas in February, Stellifer stellifer (Bloch, 1790), L. grossidens and Menticirrhus americanus (Linnaeus, 1758) were the taxa with highest relative abundances. In Balneário Barra do Sul, the increased relative abundance of L. laevigatus and P. brasiliensis differentiated March from the other months.
The dominant species in the ichthyofauna were Stellifer rastrifer, S. brasiliensis, Paralonchurus brasiliensis, Isopisthus parvipinnis, Trichiurus lepturus and Pellona harroweri. The percentage of species contribution to the dissimilarity (SIMPER) between the Sites and Months interaction is represented in Tab. III. Across the entire sampling period, the average dissimilarity between sites varied from 61.36% (November) to 93.23% (March), with three to seven species contributing over 80% of the differences in the structure of the assemblages. The analysis indicated a larger contribution of scianids, in which the same species occupied different ranks.
The monthly fluctuation of CPUE differed between Sites, as is evident by the significant interaction of factors in ANOVA (Tab. IV), in which the contrast of means by Tukey's test attributed these differences to the months of May, October and December 2007 (Fig. 5). In Balneário Barra do Sul the high incidence of S. rastrifer and S. brasiliensis in samples contributed to the differences observed in May (83.2% of capture) and October (67.2%), whereas in December, P. brasiliensis, S. brasiliensis and C. bleekerianus were responsible (75.9%) for the higher CPUE in Penha.

Comparing the rarefaction curves, the expected species richness was slightly higher in Balneário Barra do Sul (above 500 individuals), however, this difference was not significant due to an overlap of confidence intervals (Fig. 6). The Shannon diversity index (H') showed a significant interaction between space and time scales (Tab. IV). The resulting variation from the interaction between factors had a significant effect on the monthly fluctuation pattern between the Sites, in particular in August, September and November 2007, and January and February 2008, with H' ranging from 0.74 to 1.98 (Fig. 7). The lowest average H', which was observed in the month of February in Balneário Barra do Sul, was due to the dominance of I. parvipinis, whereas in Penha, the low number of species and the high abundance of P. brasiliensis contributed to the pronounced differences that were calculated for August, September and November, and S. rastrifer influenced the low average H' observed in January (Fig. 7).

The equitability index (J') fluctuated from moderate to high, with values between 0.51 and 0.83 in Balneário Barra do Sul and between 0.40 and 0.78 in Penha (Fig. 8). Although the averages were statistically similar for the Sites and Months factors, the variance analysis showed a space-time dependence, in which the distribution of the species caught differed significantly between the fishing regions in November 2007 and February 2008 (Tab. IV and Fig. 8).
The relative importance of the most abundant species, as expressed by the Berger-Parker index (d), followed a different pattern of variation between the regions with an abrupt inversion between June and August 2007 averages and, subsequently in Penha, between November 2007 and February 2008 (Fig. 9). In Balneário Barra do Sul, the index fluctuated between 0.33 (August 2007) and 0.59 (February 2008) and in Penha from 0.34 (June 2007) to 0.81 (November 2007) (Fig. 9).
DISCUSSION
The amount of collected fish in Balneário Barra do Sul and Penha reinforce the observation made in several previous studies that, despite its efficiency in capturing target species, shrimp trawl fishing is not very selective, and results in the collection of a great diversity of accompanying organisms (Alverson et al. 1994, Bakun et al. 2005, Kelleher 2005, Gillett 2008).
However, the consistently low selectivity of this bottom trawl gear does not result in similar composition and abundance of the fish that are caught in different fishing areas, and disparities in the number of species present in shrimp fisheries are frequent in the literature (Coelho et al. 1986, Paiva-Filho & Schmiegelow 1986, Ruffino & Castello 1992, Chaves et al. 2003, Gomes & Chaves 2006, Santos et al. 2008).
The high number of incidental species showed the frequent displacement in the study areas where their occurrence was associated with low abundance. Although species richness in our samples was high, nine and seven species were numerically dominant in Balneário Barra do Sul and Penha, respectively, which is a common characteristic observed in demersal fish associations (Ansari et al. 1995, Chaves et al. 2003, Branco & Verani 2006a).
Sciaenidae is among the world's most economically important fish families, representing up to 54 species in the coastal and estuarine waters of the Brazilian shoreline (Menezes et al. 2003, Nelson 2006). The fishes in this family are commonly found in shallow continental shelf over sand or mud substrates (Menezes & Figueiredo 1980), and most are demersal and dominate the accompanying fauna in artisanal seabob shrimp fisheries (Coelho et al. 1986, Paiva-Filho & Schmiegelow 1986, Giannini & Paiva-Filho 1990, Ruffino & Castello 1992, Santos et al. 1998, Hostim-Silva et al. 2002, Chaves et al. 2003, Vianna & Almeida 2005, Branco & Verani 2006a, b, Gomes & Chaves 2006, Schwarz Jr et al. 2007, Souza et al. 2008). In southeastern and southern regions, the most abundant species captured as accompanying fauna are P. brasiliensis, C. gracilicirrhus, Micropogonias furnieri (Desmarest, 1823), M. americanus, L. breviceps, S. brasiliensis, S. rastrifer, Cynoscion jamaicensis (Vaillant & Bocourt, 1883) and I. parvipinnis (Coelho et al. 1986, Paiva-Filho & Schmiegelow 1986, Muto et al. 2000, Branco & Verani 2006a, b, Schwarz Jr et al. 2007, Souza et al. 2008). Therefore, as observed along the Brazilian coast, this family, represented by 13 species and accounting for over 60% of the total abundance, was predominant in terms of number of species and abundance in the Balneário Barra do Sul and Penha regions.
Studies on the impacts of trawl fisheries on benthic and demersal communities suggest that the community structure and functioning can vary on different temporal and spatial scales (Gillett 2008, Hougues & Swain 2008). The results of PERMANOVA showed a significant spatiotemporal dependence, indicating that the interaction between the factors was expressively influenced by the community dynamics, in which fluctuations in the composition and abundance might not be explained by a single source of variation.
The similarity analysis demonstrated that, in March 2007, the ichthyofaunal composition differed between sites (93.23% dissimilarity). This discrepancy was due to the presence of C. bleekerianus, S. rastrifer, L. breviceps and Oligoplites saurus (Bloch & Schneider, 1801) in Balneário Barra do Sul, and by the presence of T. lepturus and the relatively higher average abundance of P. brasiliensis and I. parvipinnis in Penha. In January and February 2008, by contrast, the demersal fish assemblages showed a stronger dominance pattern, and most of the dissimilarities observed between Balneário Barra do Sul and Penha were due to differences in the relative abundance of S. rastrifer, S. brasiliensis and I. parvipinnis. Establish the main factors that affect the organization of local communities is a challenge for a couple of reasons. First, it is impossible to measure all of the variables at play. Second, ascertaining the complex interrelations among species living in an environment that changes over time and space is prohibitive (Muto et al. 2000). Depth, temperature, salinity and nature of the bottom sediments have been cited as the most important abiotic factors influencing the structure of soft bottom fish communities (Pires-Vanin et al. 1993, Giannini & Paiva-Filho 1995, Lowe-McConnell 1999, Barletta et al. 2005. Schwarz Jr. et al. 2007).
According to Haimovici et al. (1996) the water temperature affected the abundance of demersal bony fish on the Rio Grande shelf (southern Brazil), while species composition was associated with depth. For Muto et al. (2000) the spatial organization exhibited by the ichthyofauna along a depth gradient in the São Sebastião system (southeastern Brazil) were closely related to the distribution of the water masses which are characterized by different combinations of salinities and temperatures. Even so, it is believed that the type of substrate will have influences on both prey and demersal fish distribution, as most species live in a close association with the bottom (Araújo et al. 2006a).
Balneário Barra do Sul and Penha showed similar trends on CPUE fluctuation except in May, October and December 2007, when these trends were influenced by contributions of dominant species, particularly S. rastrifer, S. brasiliensis, and P. brasiliensis. The higher abundance of S. rastrifer and S. brasiliensis in Balneário Barra do Sul, and their low abundance or absence in Penha during October (spring) and the inversion of the CPUE in December, including the presence of P. brasiliensis, may be related to reproductive aggregation. The latter, is common within scianids (Muniz & Chaves 2008) and results in large numbers of individuals becoming vulnerable to the trawls at once. This observation may be corroborated by the observed noted pulse spawning period in the south between October and December for main species of the Stellifer genus (Chaves & Vendel 1997, Almeida & Branco 2002) and during spring and summer for P. brasiliensis (Souza & Chaves. 2007, Robert et al. 2007).
On the São Paulo coast, Muto et al. (2000) showed a significant correlation between water temperature and depth in Sciaenidae distribution, with the greatest abundance of P. brasiliensis and S. rastrifer occurring in warmer (25ºC) and shallow (15-25 m) waters. In Santos bay, Giannini & Paiva-Filho (1990) reported that S. rastrifer performs seasonal migration, remaining in the outer portion of the bay during the autumn and migrating to the inner portion during the winter, probably due to higher temperatures in shallow waters.
Although our methodology did not allow us to assess the influence of depth on the fish composition, we know that in our data trawls occurred in areas traditionally used by the artisanal fleet with a depth ranging from 5 to 25 m. Considering this information, it is possible that the differences between sites observed in May, when the water temperature is dropping, may be related to the displacement of dominant species among deeper and shallower areas, diverging from what was observed in previous studies (Bail & Branco 2003, Branco & Verani 2006a) for the Penha region.
In regard to CPUE, it is important to call attention to the peak in July (winter) with an important capture of T. lepturus (34.37% in BS and 38.38% in P of monthly capture). The numerical dominance of this species in winter may be related to the presence of juveniles in the shallow continental shelf due to higher food availability (Bittar et al. 2008). Among the food items, the occurrence of the fishes from Sciaenidae, Clupeidae and Engraulidae, and the shrimps Xiphopenaeus kroyeri, Pleoticus muelleri (Bate, 1888), and Artemesia longinaris (Bate, 1888) (Bittar et al. 2008) were both recorded in the study areas (Branco & Verani 2006a, b).
The fluctuation pattern of the Shannon diversity index across months was distinct in the two fishing regions. According to Krebs (1999), the Shannon measure increases with the number of species and gives more weight to rare species. In August, Balneário Barra do Sul showed the highest species diversity and evenness throughout the year. The disparate richness between Balneario Barra do Sul (21) and Penha (12) due to the presence of infrequent species, especially from the families Engraulidae, Clupeidae and Stromateidae, and the high participation of P. brasiliensis in Penha (64.42%) were mainly responsible for the differences in the Shannon diversity between fishing regions in August. Other observations were even more sensitive to rare species, as the contributions of P. brasiliensis in September (71.15%) and November (82.06%) and S. rastrifer (71.97%) in January and I. parvipinis in February (62.72%) reflected significant variation in the Shannon index between the sites.
During the year, the diversity index ranged between 1.04 and 1.98 in Balneário Barra do Sul and between 0.74 and 1.80 in Penha. Greenwood (1975) argued that there is a gradual increase in species diversity with decreasing latitude. Also, according to Rosenzweig (1995), the highest richness values are generally obtained in the tropical zone. In a study by Tischer & Santos (2001), the ichthyofauna accompanying a shrimp fishery in the mouth of the São Francisco River (northeastern Brazil) showed an average diversity of 3.20. On the São Paulo coast (southeastern Brazil), Paiva-Filho & Schmiegelow (1986) recorded values between 1.64 and 3.07 for Santos Bay, whereas, in São Sebastião, Muto et al. (2000) observed variation between 1.93 and 2.67. In the state of Santa Catarina (southern Brazil), the diversity ranged between 1.11 and 1.97 in seven years of sampling in the Penha region (Branco & Verani 2006a) and between 1.98 and 2.50 in five years of sampling in the Itajai-Açú river estuary (Branco et al. 2009). Apparently, the species diversity estimated by the Shannon index decreased with increasing latitude; however, other sources of variation appear to be more important to explain such observations. Differences in diversity values between these studies may be related to the size of the sampled area, seasonality, fishing effort, fishing gear used and degree of anthropogenic influence in the areas.
As observed for diversity, Pielou's evenness index showed the spatiotemporal interaction differing significantly between fish regions in November 2007 and February 2008 due to a high incidence of P. brasiliensis (Penha) and I. parvipinis (Balneário Barra do Sul), respectively. However, the index ranged from moderate to high with averages above 0.5, suggesting an acceptable homogeneity in the distribution of specimens among the species (Magurran 2004). Pielou evenness is not very sensitive to variations in abundance of rare taxa (Beisel et al. 2003). Even in the communities that were studied, in which the number of infrequent species caught was high, the fact that S. rastrifer, S. brasiliensis, P. brasiliensis, I. parvipinnis, T. lepturus and P. harroweri were relatively abundant throughout the study period, and alternated as the most abundant species in the samples, strongly influenced the increase in evenness average recorded for the collected ichthyofauna in Balneário Barra do Sul and in Penha. The Berger-Parker index clearly showed the dominance pattern, where the monthly numeric contribution of a single species ranged from approximately 30 to 80%.
It is important to highlight that the differences in ichthyofaunal structure between fishing grounds were observed 25% of the time (three months), probably due to changes in environmental factors as explained previously. On the other hand, these results enabled us to conclude that the fish assemblages captured as accompanying fauna in artisanal seabob shrimp fisheries was statistically similar in both sites throughout most of the year, even though Balneário Barra do Sul are 35 km away from each other.
ACKNOWLEDGMENTS
Thanks to our friends at the Biology Laboratory of the Universidade do Vale do Itajaí for their extraordinary contribution to the collecting and processing of samples, to our friend Jean Berná Paim for making the map of the study areas and to anonymous referees for constructive comments on the manuscript. The present study was performed with the support of the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), processes 501112/2007-3 and 305476/2006-8 and partially supported the work of J.O. Branco and J.R. Verani.
LITERATURE CITED
Submitted: 13.VII.2010; Accepted: 28.IX.2010.
Editorial responsibility: Cassiano Monteiro Neto
- Abreu, J.G.N.; J.T. Menezes & F.D. Rosa. 2006. Morfologia submarina e sedimentologia da Armação do Itapocoroy, Penha, SC, p. 37-46. In: J.O. Branco & A.W.C. Marenzi (Orgs). Bases ecológicas para um desenvolvimento sustentável: estudos de caso em Penha, SC. Itajaí, Editora da UNIVALI, 292p.
- Almeida, L.R. & J.O. Branco. 2002. Aspectos biológicos de Stellifer stellifer na pesca artesanal do camarão sete-barbas, Armação do Itapocoroy, Penha, Santa Catarina, Brasil. Revista Brasileira de Zoologia 19 (2): 601-610.
- Alverson, D.L.; M.H. Freeberg; J.G. Pope & S.A. Murawski. 1994. A global assessment of fisheries bycatch and discards. Roma, FAO, FAO Fisheries Technical Paper 339, 233p.
- Ambrose, E.E.; B.B. Solarim & A.B. Williams. 2005. Assessment of fish by-catch species from costal artisanal shrimp beam trawl fisheries in Nigeria. Fisheries Research 71: 123-132.
- Anderson, M.J. 2001. A new method for non-parametric multivariate analysis of variance. Austral Ecology 26:32-46.
- Anderson, M.J. 2005. PERMANOVA: a fortran Computer Program for Permutational Multivariate Analysis of Variance. Auckland, Department of Statistics, University of Auckland, 24p.
- Andrew, N.L. & J.G. Pepperell. 1992. The by-catch of shrimp trawl fisheries, p. 527-565. In: M. Barnes; A.D. Ansell & R.N. Gibson (Eds). Oceanography and Marine Biology Annual Review.London, UCL Press, vol 30, 640p.
- Ansari, Z.A.; A. Chatterfi; B.S. Ingole; R.A. Sreepada; C.U. Rivonkar & A.H. Parulekar. 1995. Community structure seasonal variation of on inshore demersal fish community at Goa, west coast of India. Estuarine Coastal and Shelf Science 41: 593-610.
- Araújo, F.G.; F.J.D. Guimarães & M.R.D. Costa. 2006a. Environmental influences on distribution of four Sciaenidae species (Actinopterygii, Perciformes) in a tropical bay at Southeastern Brazil. Revista Brasileira de Zoologia 23 (2): 497-508.
- Araújo, S.A.; H. Haymussi; F.H. Reis & F.E. Silva. 2006b. Caracterização climatológica do município de Penha, SC, p. 11-28. In: J.O. Branco & A.W.C. Marenzi (Orgs). Bases ecológicas para um desenvolvimento sustentável: estudos de caso em Penha, SC. Itajaí, Editora da UNIVALI, 292p.
- Bail, G.C. & J.O. Branco. 2003. Ocorrência, abundância e diversidade da ictiofauna na pesca do camarão sete-barbas, na Região de Penha, SC. Notas Técnicas da FACIMAR 7: 73-82.
- Bail, G.C. & J.O. Branco. 2007. Pesca artesanal do camarão-sete-barbas: uma caracterização sócio-econômica na Penha, SC. Brazilian Journal of Aquatic Science and Technology 11 (2): 25-32.
- Bakun, A.; S. Heileman; K.H. Kock; P. Mace; W. Perrin; K. Stergiou; U. R. Sumaila; M. Vierros; K. Freire & Y. Sadovy. 2005. Marine Fisheries Systems, p. 477-511. In: R. Watson & A. H. Zakri (Orgs). Ecosystems and Human Well-Being: Current State and Trends Washington, Island Press, vol. 1, 901p.
- Barletta, M.; A. Barletta-Bergan; U. Saint-Paul & G. Hubold. 2005. The role of salinity in structuring the fish assemblages in a tropical estuary. Journal of Fish Biology 66: 45-72.
- Beisel, J.N.; P. Usseglio-Polatera; V. Bachmann & J.C. Moreteau. 2003. A Comparative Analysis of Evenness Index Sensitivity. International Review of Hydrobiology 88: 3-15.
- Bittar, V.T.; B.F. Castello & A.P.M.D. Beneditto. 2008. Hábito alimentar do peixe-espada adulto, Trichiurus lepturus, na costa norte do Rio de Janeiro, sudeste do Brasil. Biotemas 21 (2): 83-90.
- Branco, J.O. 2005. Biologia e pesca do camarão-sete-barbas Xiphopenaeus kroyeri (Heller) (Crustacea, Penaeidae), na Armação do Itapocoroy, Penha, SC. Revista Brasileira de Zoologia 22 (4): 1050-1062.
- Branco, J.O. & J.R. Verani. 2006a. Análise quali-quantitativa da ictiofauna acompanhante na pesca do camarão-sete-barbas, na Armação do Itapocoroy, Penha, Santa Catarina. Revista Brasileira de Zoologia 23 (2): 381-391.
- Branco, J.O. & J.R. Verani. 2006b. Pesca do camarão sete-barbas e sua fauna acompanhante, na Armação do Itapocoroy, Penha, SC, p. 153-170. In: J. O. Branco & A. W. C. Marenzi (Orgs). Bases ecológicas para um desenvolvimento sustentável: estudos de caso em Penha, SC. Itajaí, Editora da UNIVALI, 292p.
- Branco, J.O.; F. Freitas Júnior; J.R.Verani & M. Hostim-Silva. 2009. Ictiofauna do Saco da Fazenda, Itajaí, SC, p. 207-226. In: J.O. Branco; M. J. Lunardon-Branco & V. R. Bellotto (Eds). Estuário do Rio Itajaí-Açú, Santa Catarina: caracterização ambiental e alterações antrópicas. Itajaí, Editora UNIVALI, 312p.
- Chaves, P.T.C. & A.L. Vendel. 1997. Reprodução de Stellifer rastrifer (Jordan) (Teleostei, Sciaenidae) na Baía de Guaratuba, Paraná, Brasil. Revista Brasileira de Zoologia 14 (1): 81-89.
- Chaves, P.T.C; G. Cova-Grando & C. Calluf. 2003. Demersal ichthyofauna in a continental shelf region on the south coast of Brazil exposed to shrimp trawl fisheries. Acta Biológica Paranaense 32 (1-4): 69-82.
- Corrêa, I.C.S.; L.R. Martins; J.M.M. Ketzer; A.R.D. Elias & R. Martins. 1996. Evolução sedimentológica e paleogeográfica da plataforma continental sul e sudeste do Brasil. Pesquisas 9: 56-61.
- Clarke, K.R. & R.M. Warwick. 1994. Change in marine communities: an approach to statistical analysis and interpretation. Plymouth, Plymouth Marine Laboratory, 144p.
- Clarke, K.R.; M.G. Chapman; P.J. Somerfield & H.R. Needham. 2006. Dispersion-based weighting of species counts in assemblage analyses. Marine Ecology Progress Series 320: 11-27.
- Coelho, J.A.P.; A. Puzzi; R. Graça-Lopes; E.S. Rodrigues & O. Preto Jr. 1986. Análise da rejeição de peixes na pesca artesanal dirigida ao camarão sete-barbas (Xiphopenaeus kroyeri) no litoral do Estado de São Paulo. Boletim do Instituto de Pesca 13 (2): 51-61.
- Colwell, R.K.; C.X. Mao & J. Chang. 2004. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 85 (10): 2717-2727.
- Dayton, P.K.; S. Thrush & F. Coleman. 2002. Ecological Effects of Fishing in Marine Ecosystems of the United States. Arlington, Pew Oceans Commission, 44p.
- Figueiredo, J.L. 1977. Manual de peixes marinhos do Sudeste do Brasil. I. Introdução. Cações, raias e quimeras. São Paulo, Museu de Zoologia, Universidade de São Paulo, 104p.
- Figueiredo, J.L. 1980. Manual de peixes marinhos do sudeste do Brasil. III. Teleostei (2). São Paulo, Museu de Zoologia, Universidade de São Paulo, 90p.
- Figueiredo, J.L. & n. Menezes. 1978. Manual de peixes marinhos do Sudeste do Brasil. II. Teleostei (1). São Paulo, Museu de Zoologia, Universidade de São Paulo, 110p.
- Giannini, R. & A.M. Paiva-Filho. 1990. Os Sciaenidae (Teleostei: Perciformes) da Baía de Santos (SP), Brasil. Boletim do Instituto Oceanográfico 38 (1): 69-86.
- Giannini, R. & A.M. Paiva-Filho. 1995. Distribuição temporal, espacial e bioecologia do cangoá, Stellifer brasiliensis (Teleostei: Sciaenidae), na Baía de Santos, São Paulo, Brasil. Arquivos Ciência do Mar 29 (1-2): 5-15.
- Gillett. R. 2008. Global study of shrimp fisheries. Roma, FAO, FAO Fisheries Technical Paper 475, 331p.
- Gomes, I.D. & P.T. Chaves. 2006. Ictiofauna integrante da pesca de arrasto camaroeiro no litoral sul do Estado do Paraná, Brasil. Bioikos 20 (1): 9-13.
- Graça-Lopes, R.; A.R.G. Tomás; S.L.S Tutui; E. Severino Rodrigues & A. Puzzi. 2002. Fauna acompanhante da pesca camaroeira no litoral do estado de São Paulo, Brasil. Boletim do Instituto de Pesca 28 (2): 173-188.
- Greenwood, P.H. 1975. A history of fishes. New York, Jonh Wiley & Sons, 467p.
- Haimovici, M. & J.T. Mendonça. 1996. Descartes da fauna acompanhante na pesca de arrasto de tangones dirigida a linguados e camarões na plataforma continental do sul do Brasil. Atlântica 18: 161-177.
- Haimovici, M.; A.S. Martins & P.C. Vieira. 1996. Distribuição e abundância de peixes teleósteos demersais sobre a plataforma continental do sul do Brasil. Revista Brasileira de Biologia 56 (1): 27-50.
- Horn Filho, N.O. 2003. Setorização da Província Costeira de Santa Catarina em base aos aspectos geológicos, geomorfológicos e geográficos. Geosul 18 (35): 71-98.
- Hostim-Silva, M.; M.J.D. Vicente; V. Figna & J.P. Andrade. 2002. Ictiofauna do rio Itajaí-Açú, Santa Catarina, Brasil. Notas Técnicas da FACIMAR 6: 127-135.
- Hougues, P.B. & D.P. Swain. 2008. Impacts of environmental change and direct and indirect harvesting effects on the dynamics of a marine fish community. Canadian Journal of Fisheries and Aquatic Sciences 65: 2088-2104.
- Kelleher, K. 2005. Discards in the world's marine fisheries - An update. Roma, FAO, FAO Fisheries Technical Paper 470, 131p.
- Krebs, C.J. 1999. Ecological methodology. Menlo Park, Benjamin/Cummings, 2nd ed., 620p.
- Lowe-Mcconnell, R.H. 1999. Estudos ecológicos de comunidades de peixes tropicais. São Paulo, Editora da Universidade de São Paulo, 534p.
- Magurran, A.E. 2004. Measuring biological diversity. Oxford, Blackwell Publishing, 256p.
- Menezes, N. & J.L. Figueiredo. 1980. Manual de peixes marinhos do Sudeste do Brasil. IV. Teleostei (3). São Paulo, Museu de Zoologia, Universidade de São Paulo, 96p.
- Menezes, N. & J.L. Figueiredo. 1985. Manual de peixes marinhos do Sudeste do Brasil. V. Teleostei (4). São Paulo, Museu de Zoologia, Universidade de São Paulo, 105p.
- Menezes, N.A.; P.A. Buckup; J.L. Figueiredo & R.L. Moura. 2003. Catálogo das espécies de peixes marinhos do Brasil. São Paulo, Museu de Zoologia, Universidade de São Paulo, 160p.
- MMA/IBAMA. 2005. Estatística da pesca 2004: Brasil, grandes regiões e unidades da federação. Brasília, MMA, Ibama, 98p.
- Muniz, E.R. & P.T.C. Chaves. 2008. Condição reprodutiva da betara preta, Menticirrhus americanus (Teleostei, Sciaenidae), na pesca realizada no litoral norte de Santa Catarina, Brasil. Acta Scientiarum Biological Sciences 30 (4): 339-344.
- Muto, E.Y; L.H. Soares & C.L.D.B. Rossi-Wongtschowski. 2000. Demersal fish assemblages off São Sebastião, southeastern Brazil: structure and environmental conditioning factors (summer 1994). Revista Brasileira de Oceanografia 48 (1): 9-27.
- Nelson, J.S. 2006. Fishes of the world. Hoboken, John Wiley, 4th ed., 601p.
- Paiva-Filho, A.M. & J.M.M. Schmiegelow. 1986. Estudo sobre a ictiofauna acompanhante da pesca do camarão sete-barbas (Xiphopenaeus kroyeri) nas proximidades da Baía de Santos - SP. I. Aspectos quantitativos. Boletim do Instituto Oceanográfico 34: 79-85.
- Pires-Vanin, A.M.S.; B.C.L.D. Rossi-Wongtschowski; E. Aidar; H.S.L. Mesquita; L.S.H. Soares & M. Katsuragawa. 1993. Estrutura e função do ecossistema de plataforma continental do Atlântico Sul brasileiro: síntese dos resultados. Publicação Especial do Instituto Oceanográfico 10: 217-231.
- Pitcher, T.J. & P.J.B. Hart. 1982. Fisheries ecology. London, Croom Helm, 414p.
- PROZEE. 2006. Relatório final do projeto de monitoramento da atividade pesqueira no litoral do Brasil - Projeto ESTATPESCA. Brasília, Convênio SEAP/IBAMA/PROZEE Nº 109/2004, Fundação de Amparo à Pesquisa de Recursos Vivos na Zona Econômica Exclusiva, 328p.
- Robert, M.D.C.; M.A. Michels-Souza & P.T. Chaves. 2007. Biologia de Paralonchurus brasiliensis (Steindachner)(Teleostei, Sciaenidae) no litoral sul do Estado do Paraná, Brasil. Revista Brasileira de Zoologia 24 (1): 191-198.
- Rosenzweig, M.L. 1995. Species diversity in space and time. Cambridge, Cambridge University Press, 436p.
- Ruffino, M.L. & J.P. Castello. 1992 [1993]. Alterações na ictiofauna acompanhante da pesca do camarão barba-ruça (Artemesia longinaris) nas imediações da Barra de Rio Grande, Rio Grande do Sul - Brasil. Nerítica 7 (1-2): 43-55.
- Santos, M.C.F. 2000. Diversidade ecológica da ictiofauna acompanhante nas pescarias de camarões em Tamandaré (Pernambuco-Brasil). Boletim Técnico-Cientifico do CEPENE 8 (1): 7-26.
- Santos, M.C.F.; A.E.T.S. Freitas & M. Mendes da Silva. 1998. Composição da ictiofauna acompanhante da pesca de camarão em Tamandaré/PE e Pontal do Peba/AL. Boletim Técnico-Cientifico do CEPENE 6 (1): 1-17.
- Santos, M.C.F.; L. Almeida & C.G.M. da Silva. 2008. Avaliação quali-quantitativa da ictiofauna acompanhante na pesca do caramão sete-barbas, Xiphopenaeus kroyeri (Heller, 1862) no município de Caravelas (Bahia - Brasil). Boletim Técnico-Cientifico do CEPENE 16 (1): 99-107.
- Schettini, C.A.F.; J.L.B. Carvalho & E.C. Truccolo. 1999. Aspectos hidrodinâmicos da enseada da Armação do Itapocoroy, SC. Notas Técnicas da FACIMAR 3: 99-109.
- Schwarz Jr, R.; A.C.N.P. Franco; H.L. Spach; C. Santos; H.A. Pichler & G.M.L.N. Queiroz. 2007. Variação da estrutura espacial da ictiofauna demersal capturada com rede de arrasto de porta na Baía dos Pinheiros, PR. Boletim do Instituto de Pesca 33 (2): 157-169.
- Severino-Rodrigues, E.; D.S.F. Guerra & R. Graça-Lopes. 2002. Carcinofauna acompanhante da pesca dirigida ao camarão sete-barbas (Xiphopenaeus kroyeri) desembarcado na praia do Perequê, Estado de São Paulo, Brasil. Boletim do Instituto de Pesca 28 (1): 33-48.
- Soares, L.H. 1978. Estudo dos Bothidae, Cynoglossidae e Soleidae capturados nos bancos de camarão, no Estado do Rio Grande do Norte - Brasil. Boletim do Departamento de Oceanografia e Limnologia do Centro de Biociências 6: 16-27.
- Souza, L.M. & P.T. Chaves. 2007. Atividade reprodutiva de peixes (Teleostei) e o defeso da pesca de arrasto no litoral norte de Santa Catarina, Brasil. Revista Brasileira de Zoologia 24 (4): 1113-1121.
- Souza, U.P.; R.C.D. Costa; I.A. Martins & A. Fransozo. 2008. Associações entre as biomassas de peixes Sciaenidae (Teleostei: Perciformes) e de camarões Penaeoidea (Decapoda: Dendrobranchiata) no litoral norte do Estado de São Paulo. Biota Neotropica 8 (1): 83-92.
- Tischer, M. & M.C.F. Santos. 2001. Algumas considerações sobre a ictiofauna acompanhante da pesca de camarões na foz do rio São Francisco (Alagoas/Sergipe-Brasil). Boletim Técnico-Cientifico do CEPENE 9 (1): 155-165.
- Underwood, A.J. 1997. Experiments in ecology: Their logical design and interpretation using analysis of variance. Cambridge, Cambridge University Press, 150p.
- Vianna, M. & T. Almeida. 2005. Bony fish bycatch in the Southern Brazil pink shrimp (Farfantepenaeus brasiliensis and F. paulensis) fishery. Brazilian Archives of Biology and Technology 48 (4): 611-623.
- Zar, J.H. 1996. Biostatistical analysis. New Jersey, Prentice Hall, 121p.
Publication Dates
-
Publication in this collection
27 May 2011 -
Date of issue
Apr 2011
History
-
Received
13 July 2010 -
Accepted
28 Sept 2010