Abstract
This study aimed to describe the species composition and abundance of the porcellanid crabs inhabiting the consolidated sublittoral off an island on the southeastern coast of Brazil. During a sampling program at Vitória Island (23º44'S, 45º01'W), collections were carried out monthly from February 2004 through January 2006. The crabs were hand-caught by SCUBA divers during the daytime. The greatest carapace width (CW) was measured, and the sex and developmental stage were identified for each crab. The specimens were grouped into five demographic categories: juvenile males, adult males, juvenile females, adult females, and ovigerous females. A total of 275 crabs belonging to three genera and six species were obtained. Petrolisthes amoenus (Guérin-Méneville, 1855) and Pachycheles riisei (Stimpson, 1858) were the most abundant species. Most of the demographic classes were represented for both species, except that no adult females of P. riisei were recorded. Among the porcellanids, Pachycheles monilifer (Dana, 1852) was the dominant species; it comprised more than 75% of the total individuals sampled, and showed the highest constancy index for the region. The results obtained in this study provided information regarding the faunal composition and the relative abundance of the species, which will be useful as a reference for other sublittoral consolidated-bottom regions along the southeast Brazilian coast.
Checklist; Porcellanidae; subtidal
ECOLOGY
Composition and abundance of porcellanid crabs (Crustacea: Decapoda: Anomura) from rocky bottoms off Vitória Island, southeast coast of Brazil
Douglas F. R. Alves; Samara de P. Barros-Alves; Valter J. Cobo* * Corresponding author: E-mail: vjcobo@unitau.br
Laboratório de Biologia Marinha, Instituto Básico de Biociências, Universidade de Taubaté.Avenida Tiradentes 500, Centro, 12030-180 Taubaté, SP, Brazil
ABSTRACT
This study aimed to describe the species composition and abundance of the porcellanid crabs inhabiting the consolidated sublittoral off an island on the southeastern coast of Brazil. During a sampling program at Vitória Island (23º44'S, 45º01'W), collections were carried out monthly from February 2004 through January 2006. The crabs were hand-caught by SCUBA divers during the daytime. The greatest carapace width (CW) was measured, and the sex and developmental stage were identified for each crab. The specimens were grouped into five demographic categories: juvenile males, adult males, juvenile females, adult females, and ovigerous females. A total of 275 crabs belonging to three genera and six species were obtained. Petrolisthes amoenus (Guérin-Méneville, 1855) and Pachycheles riisei (Stimpson, 1858) were the most abundant species. Most of the demographic classes were represented for both species, except that no adult females of P. riisei were recorded. Among the porcellanids, Pachycheles monilifer (Dana, 1852) was the dominant species; it comprised more than 75% of the total individuals sampled, and showed the highest constancy index for the region. The results obtained in this study provided information regarding the faunal composition and the relative abundance of the species, which will be useful as a reference for other sublittoral consolidated-bottom regions along the southeast Brazilian coast.
Key words: Checklist; Porcellanidae; subtidal
Porcellanidae crabs are distributed in tropical and subtropical regions through the western Atlantic Ocean, from North Carolina (USA) to Argentina. In Brazil, the family is represented by 23 species in seven genera, which are found along the entire Brazilian coast (Melo 1999). Porcellanids are mainly inhabitants of the coastal zone, and are commonly found in intertidal and sublittoral zones, particularly on hard substrata such as caves, excavations in coral reefs, under rocks, calcareous seaweed bottoms, worm tubes and sponge cavities (see Rodríguez et al. 2005).
Knowledge of the faunal composition on a regional scale can be used for several purposes, as follows: to provide basic information for comparative biodiversity studies; as a tool to recognize and to delimit protection areas; in the identification of potential anthropogenic impacts, through evaluation of the complexity of the local community; or to provide useful information to estimate the availability of living resources (Hendrickx & Harvey 1999).
Although information on the biology of crabs is still sparse, some studies have reported on important features of porcellanid populations in general (Micheletti-Flores & Negreiros-Fransozo 1999), community dynamics (Negreiros-Fransozo et al. 1997, Fransozo et al. 1998, Mantelatto & Souza-Carey 1998, Rivadeneira et al. 2010), community richness (Lira et al. 2001, Siddiqui & Kazmi 2003, Hernández-Ávila et al. 2007, Lira et al. 2007), geographic distributions of western Atlantic species (Werding et al. 2003, Rodríguez et al. 2005), and records of larvae in estuarine areas (Magris & Loureiro-Fernandes 2005).
According to Witman & Dayton (2001), although the number of experimental studies conducted in sublittoral habitats is increasing rapidly, the ecology of communities on subtidal rocky substrates is still at the descriptive phase relative to what is known for intertidal communities. However, basic patterns of community structure are known for many subtidal regions of the world.
For the southeastern Brazilian coast, research on decapods of the rocky sublittoral is still sparse (e.g. Mantelatto et al. 2004a, b, Alves et al. 2006), particularly at the community level (e.g. Hebling et al. 1994, Fransozo et al. 1998, Mantelatto & Fransozo 2000, Santos & Pires-Vanin 2004).
This study aimed to describe the species composition and abundance of the porcellanid crabs inhabiting the consolidated sublittoral off Vitória Island, on the northern coast of the state of São Paulo. We focused on the variability in the size of individuals, the distribution of demographic classes, and the abundance and constancy of species collected.
MATERIAL AND METHODS
The sampling program was carried out at Vitória Island (23º44'04"S, 45º01'35"W), southeastern coast of Brazil (Fig. 1). Collections were carried out monthly, from February 2004 through January 2006. The crabs were hand-caught by a SCUBA diver during daytime, on the rocky bottom, between rocks, crevices and cavities of sponges and corals such as Madracis decactis (Lyman, 1859) and Mussismilia hispida (Verrill, 1902). The sampling effort consisted of one hour per dive for each of the two divers, at depths of 5 to 20 m.
The specimens collected were transferred to the Marine Biology Laboratory at the University of Taubaté (UNITAU), where they were preserved in 70% ethanol and placed in labeled jars. The species were identified according to Melo (1999) and classified according to Martin & Davis (2001). The carapace width (CW) of each crab was measured with a vernier caliper (0.1 mm. The sex and developmental stage were identified based on the external morphology of the abdomen and its appendages. The crabs were grouped according to sex and developmental stage as follows: adult male (AM), juvenile male (JM), adult female (AF), juveline female (JF), ovigerous female (OF), indeterminate male (IM) and indeterminate female (IF). The adult stage was estimated based on the size of the smallest ovigerous female such as all crabs this size and up were identified as adults. For each species, the mean carapace width and its standard deviation were estimated. After the analyses, the specimens were deposited at the Carcinology Scientific Collection of the Marine Biology Laboratory (UNITAU), and at the Carcinology Collection of the Museu de Zoologia of the Universidade de São Paulo (MZUSP).
To evaluate the hierarchical importance of each species in the porcellanid community of the sublittoral consolidated bottoms around Vitória Island, the relative abundance (RAb) and the constancy index (C) of each species were estimated according to Dajoz (2005).
RESULTS
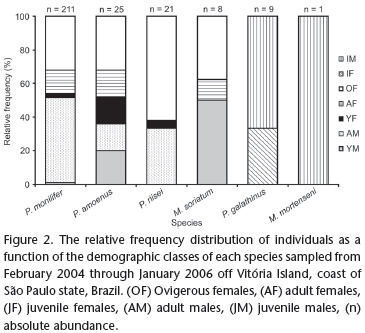
A total of 275 crabs, in three genera and six species, were obtained. The total number of individuals caught per species ranged from 211 for Pachycheles monilifer (Dana, 1852) to only one for Megalobrachium mortenseni Haig, 1962 (Tab. I). None of the species were recorded in all samples. Pachycheles monilifer, missing only in the sample from March 2004, had the greatest constancy index (0.958). By contrast, M. mortenseni was the least frequent in the samples (0.042) (Tab. I). Pachycheles monilifer showed the greatest relative abundance, comprising more than 75% of the individuals sampled; none of the remaining species accounted for 10% of the total sampled (Tab. I). Crab size ranged from 1.5 CW for Petrolisthes amoenus (Guérin-Méneville, 1855) (MZUSP-16713) to 10.3 mm CW for Petrolisthes galathinus (Bosc, 1802) (Tab. I).
Following P. monilifer, P. amoenus and Pachycheles riisei (Stimpson, 1858) were the most abundant over the entire sampling program. All demographic classes were recorded for these species, with the exception of P. riisei, for which no adult female was found. At least 60% of the specimens in the samples were adults (Fig. 2).
DISCUSSION
According to Melo (1999), 23 species of porcellanid crabs in seven genera are found in Brazilian waters; of these, 13 are known from the coast of the state of São Paulo. The number of species recorded in this study is about 26% of the total number of species recorded for the Brazilian coast, and 46% of the total richness recorded for the coast of São Paulo.
Comparing the species composition of the community we investigated with that of a soft-bottom benthic community in the same region reveals that these two kinds of bottom habitats harbor different porcellanid faunas. In the soft-bottom sublittoral of the northeastern coast of São Paulo, five species have been recorded: Megalobrachium roseum (Rathbun, 1900), Myniocerus angustus (Dana, 1852), Petrolisthes rosariensis Werding, 1978, Pisidia brasiliensis Haig in Rodrigues da Costa, 1968, and Porcellana sayana (Leach, 1820), which are associated with hermit crabs (Hebling et al. 1994, Negreiros-Fransozo et al. 1997, Fransozo et al. 1998).
The records above support the hypothesis that features of the substratum and the availability of food and refuges are important factors limiting the establishment of porcellanid crabs in the region. Abele (1974) stated that the substratum type is the major factor influencing the richness of decapod crustaceans in marine environments.
The species composition recorded in this study is similar to that reported for other types of hard substrates. For instance, Mantelatto & Souza-Carey (1998) found porcellanids in colonies of the bryozoan Schizoporella unicornis (Johnston, 1847). Micheletti-Flores & Negreiros-Fransozo (1999) reported porcellanids from sandy reefs of Phragmatopoma lapidosa Kinberg, 1867, of which only M. mortenseni was not associated with P. lapidosa.
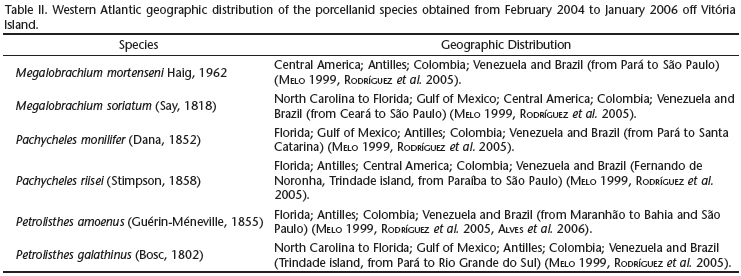
Among the species collected in this study, only P. monilifer is found on all kinds of substrata. Mantelatto & Souza-Carey (1998) also reported that this species was the most abundant crab found in bryozoan colonies, as found here in the Vitória Island region. However, S. unicornis colonies are common from the lower limit of the intertidal to depths of 20 m in the sublittoral (Ross & McCain 1976). Because colonies of S. unicornis provide environmental conditions that are very similar to rocky bottoms, they may help the establishment of P. monilifer, allowing this species to become abundant. Pachycheles monilifer is one of the most common porcellanid species in the sandy reefs of P. lapidosa in intertidal areas, suggesting that this crab may be more vulnerable to the fluctuations in abiotic factors that characterize these areas. According to Veloso & Melo (1993), Werding et al. (2003) and Rodríguez et al. (2005), P. monilifer has a wide geographical (see Tab. II) and ecological range on intertidal and sublittoral consolidated bottoms, revealing a greater ecological valence.
Another point that deserves attention is the occurrence of P. amoenus at the consolidated sublittoral off Vitória Island, because this species has only recently been reported from São Paulo (see Tab. II). The coast of the state of Bahia, northeastern Brazil, was believed to represent the southern limit of P. amoenus. This is because the Cabo Frio region, on the northeastern coast of the state of Rio de Janeiro, works as a "biological filter", which limits the southern distribution of thermophilic species to the coasts of Espírito Santo and northern Rio de Janeiro. The presence of P. amoenus on the coast of São Paulo means that it has advanced about 7º southwards, overcoming Cabo Frio's "biological filter". The wide gap between the previous and present southern limits of P. amoenus may be due to anthropogenic introduction (see Alves et al. 2006). However, an alternative explanation for this gap is that previous surveys in the region may have missed P. amoenus because individuals of this species are small and have cryptic behavior. The small size and cryptic behavior of P. amoenus may have prevented this species from being recorded earlier.
Petrolisthes amoenus was recorded as the second most abundant species during the studied period, with individuals in every demographic category. This suggests that it is well established in the region, and that it seems to be a successful competitor among the rocky-bottom benthic fauna. If P. amoenus is in fact an exotic species in the region, native species such as P. galathinus could be affected, mainly by competition for space on the hard substrata, which may be a sparse resource. Therefore, the presence of P. amoenus off Vitória Island and its impact must be further investigated, because the presence of a non-native species can result in alterations of the taxonomic composition of the local community (Tavares & Mendonça Jr 2004).
All species collected in this study can travel short distances by swimming, which seems to allow them to avoid predators (personal observation). Considering that all developmental stages of P. monilifer were found on rocky bottom, we suggest that this species does not exploit other habitats. These crabs most likely spend their entire post-larval phase in association with the hard sublittoral. This hypothesis cannot be extended to the other species we collected because of their low frequency in the samples, which affects the analysis of the sex ratios and the demographic distribution of classes.
The species composition and the abundance patterns of the Porcellanidae off Vitória Island are dominated by P. monilifer, the most important species in this community. Our results indicate some degree of habitat partitioning, in particular regarding the substrata, where different groups of species live on the soft and hard bottoms of the southeastern Brazilian coast. This situation suggests that programs aiming to monitor/preserve these crabs must take into account the types of substrata that they occupy. In general, our results add to knowledge of the regional biodiversity and can help to identify potential environmental impacts.
ACKNOWLEDGEMENTS
The authors are indebted Janet Reid for her constructive comments on early drafts of the manuscript, and help with the English language. We thank the "Omni Mare" Dive Center for providing facilities and other logistic assistance during the collections. All sampling in this study have been conducted in accordance to pertinent state and federal laws. V.J. Cobo is also grateful to the Universidade de Taubaté for financial support (Process # 251/2001-PRPPG). D.F.R. Alves is grateful to CNPq (Process # 134950/2007-0) for finantial support for his masters degree. Currently D.F.R. Alves and S.P. Barros-Alves, are associated to the Programa de Pós-Graduação em Zoologia of the Universidade Estadual Paulista (UNESP).
LITERATURE CITED
Submitted: 29.VII.2010; Accepted: 28.XI.2010.
Editorial responsibility: Paulo da C. Lana
- Abele, S.M. 1974. Species diversity of decapod crustaceans in marine habitats. Ecology 55: 156-161.
- Alves, D.F.R.; V.J. Cobo & G.A.S. Melo. 2006. Extension of the geographical distribution of some brachyuran and porcellanid decapods (Crustacea) to the coast of the State of São Paulo, Brazil. Revista Brasileira de Zoologia 23 (4): 280-1283.
- Dajoz, Z.R. 2005. Princípios de Ecologia. Porto Alegre, Editora Artmed, 7th ed.,520p.
- Fransozo, A.; F.L.M. Mantelatto; G. Bertini; L.C. Fernandez-Góes & J.M. Martinelli. 1998. Distribution and assemblages of anomuran crustacean in Ubatuba Bay, north coast of São Paulo State, Brazil. Acta Biológica Venezuelica 18 (4): 17-25.
- Hebling, N.J.; F.L.M. Mantelatto; M.L. Negreiros-Fransozo & A. Fransozo. 1994. Levantamento e distribuição de braquiúros e anomuros (Crustacea, Decapoda) dos sedimentos sublitorais da região da Ilha Anchieta, Ubatuba (SP). Boletim do Instituto de Pesca 21: 1-9.
- Hendrickx, M.E. & A.W. Harvey. 1999. Checklist of anomuran crabs (Crustacea: Decapoda) from the eastern tropical pacific. Belgian Journal of Zoology 129 (2): 363-389.
- Hernández-Ávila, I.; A. Gómez; C. Lira & L. Galindo. 2007. Benthic decapod crustaceans (Crustacea: Decapoda) of Cubagua Island, Venezuela. Zootaxa 1553: 33-45.
- Lira, C.; G. Hernández & J. Bolaños. 2001. Cangrejos porcelánidos (Decapoda: Anomura) de las islas orientales de Venezuela. I. El género Megalobrachium Stimpson, 1858, con dos adiciones a la carcinofauna Venezolana. Boletim Institut Oceanografic Venezuela 40 (1-2): 55-66.
- Lira, C.; G. Hernández; J. Bolaños; K. Graterol & M. Piñate C. 2007. Cangrejos porcelánidos (Decapoda: Anomura) de Las Islas Nororientales de Venezuela. II - El Género Pachycheles Stimpson, 1858. Boletim Institut Oceanografic Venezuela 46 (1): 37-50.
- Magris, R.A. & L. Loureiro-Fernandes. 2005. Levantamento preliminar das larvas de Porcellanidae (Decapoda, Anomura) do sistema estuarino dos rios Piraquê-Mirim, Aracruz, Espírito Santo. Brazilian Journal of Aquatic Sciences and Technology 9 (1): 49-54.
- Mantelatto, F.L.M. & A. Fransozo. 2000. Brachyuran community in Ubatuba Bay, northern coast of São Paulo State, Brazil. Journal of Shellfish Research 19 (2): 701-709.
- Mantelatto, F.L.M & M.M. Souza-Carey. 1998. Caranguejos anomuros (Crustacea, Decapoda) associados a Schizoporella unicornis (Bryozoa, Gymnolaemata) em Ubatuba (SP), Brasil. Anais do IV Simpósio de Ecossistemas Brasisleiros, Publicação ACIESP 2 (104): 200-206.
- Mantelatto, F.L.M.; F.C.R. Faria; R.B. Garcia & G.A.S. Melo. 2004a. Majoid crabs community (Crustacea: Decapoda) from infralittoral rocky/sandy bottom of Anchieta Island, Ubatuba. Brazilian Archives of Biology and Technology 47 (2): 273-279.
- Mantelatto, F.L.M.; R. Biagi; F.C.R. Faria; A.L. Meireles & G.A.S. Melo. 2004b. Checklist on brachyuran fauna (Decapoda) from infralittoral rocky/sandy bottom of Anchieta Island, São Paulo State, Brazil. Nauplius 12 (2): 135-142.
- Martin, J.W. & G.E. Davis. 2001. An updated classification of the recent Crustacea. Los Angeles, Natural History Museum of Los Angeles County, Science Series, 124p.
- Melo, G.A.S. 1999. Manual de Identificação dos Crustacea Decapoda do litoral brasileiro: Anomura; Thalassinidea; Palinuridea e Astacidea. São Paulo, Plêiade/Fapesp, 551p.
- Micheletti-Flores, C.V. & M.L. Negreiros-Fransozo. 1999. Porcellanid crabs (Crustacea, Decapoda) inhabiting sand reefs built by Phragmatopoma lapidosa (Polychaeta, Sabellariidae) at Paranapuã Beach, São Vicente, SP, Brazil. Revista Brasileira de Biologia 59 (1): 63-73.
- Negreiros-Fransozo, M.L.; A. Fransozo; F.L.M. Mantelatto; M.A.A. Pinheiro & S. Santos. 1997. Anomuran species (Crustacea, Decapoda) and their ecological distribution at Fortaleza Bay sublittoral, Ubatuba, São Paulo, Brazil. Iheringia, Série Zoologia (83): 187-194.
- Rivadeneira, M.M.; P. Hernáez; J.A. Baeza; S. Boltaña; M. Cifuentes; C. Correa; A. Cuevas; E. del Valle; I. Hinojosa; N. Ulrich; N. Valdivia; N. Vasquez; A. Zander & M. Thiel. 2010. Testing the abundant-centre hypothesis using intertidal porcelain crabs along the Chilean coast: linking abundance and life-history variation. Journal of Biogeography 37 (3): 486-498.
- Rodríguez, I.T.; G. Hernández & D.L. Felder. 2005. Review of the Western Atlantic Porcellanidae (Crustacea: Decapoda: Anomura) with new records, Systematic observations, and comments on Biogeography. Caribbean Journal of Science 41 (3): 544-582.
- Ross, J.R.P. & K.W. McCain. 1976. Schizoporella unicornis (Ectoprocta) in coastal waters of northwestern United States and Canada. Northwest Science 50 (3): 160-171.
- Santos, M.F.L. & A.M.S. Pires-Vanin. 2004. Structure and dynamics of the macrobenthic communities of Ubatuba bay, southeastern brazilian coast. Brazilian Journal of Oceanography 63 (1): 15-25.
- Siddiqui, F.A. & Q.B. Kazmi. 2003. A checklist of marine anomurans (Crustacea: Decapoda) of Pakistan, northern Arabian Sea. Memories Museum Victoria 60 (1): 87-89.
- Tavares, M. & J.B. Mendonça Jr. 2004. Introdução de Crustáceos Decápodes exóticos no Brasil: uma roleta ecológica, p. 59-76. In: J.S.V. Silva & R.C.C.L. Souza (Eds). Água de lastro e bioinvasão. Rio de Janeiro, Interciência, 224p.
- Veloso, V.G. & G.A.S. Melo. 1993. Taxonomia e distribuição da família Porcellanidae (Crustacea, Decapoda, Anomura) no litoral brasileiro. Iheringia, Série Zoologia, 75: 171-186.
- Werding, B.; A. Hiller & R. Lemaitre. 2003. Geographic and depth distributional patterns of western Atlantic Porcellanidae (Crustacea: Decapoda: Anomura), with an updated list of species. Memories Museum Victoria 60 (1): 79-85.
- Witman, J.D. & P.K. Dayton. 2001. Rocky subtidal communities, p. 339-366. In: M.D. Bertness; S.D. Gaine & M.E. Hay (Eds). Marine community ecology. Massachusetts, Sinauer Associates, 550p.
Publication Dates
-
Publication in this collection
27 May 2011 -
Date of issue
Apr 2011
History
-
Accepted
28 Nov 2010 -
Received
29 July 2010