Abstract
Two new species of Thaumastus (Thaumastus) from the state of Minas Gerais, southeastern Brazil, are described. They are diagnosed and characterized by the morphology of the shell and soft parts and compared with Brazilian species of the subgenus Thaumastus s.s. Martens, 1860. Thaumastus (T.) parvus sp. nov. is similar to T. (T.) baixoguanduensis Pena, Coelho & Salgado, 1996 but can be distinguished by the smaller size, smaller number of plates in the jaw, different number of follicle groups in the ovotestis and form of the fertilization complex. Thaumastus (T.) caetensis sp. nov. is similar to T. (T.) largillierti (Philippi, 1845) but differs by the width and the contour of the parietal side of the shell aperture. T. (T.) caetensis is also closer to Thaumastus (T.) baixoguanduensis but can be distinguished by the smaller dimensions (height, width and number of protoconch whorls), the lack of a transversal light band on the body whorl, the jaw with smaller number of plates, and the radula with 35 teeth. In the soft parts, this new species differs also in the number of follicle gatherings in ovotestis, fertilization complex with globose shape, and penian retractor muscle terminally and laterally attached to flagellum
Mollusca; morphology; Orthalicoidea; taxonomy
TAXONOMY AND NOMENCLATURE
Two new species of Thaumastus (Gastropoda: Pulmonata: Orthalicidae: Bulimulinae) from the state of Minas Gerais, Brazil
Meire Silva PenaI,1 1 Corresponding author. E-mail: meirepena@yahoo.com.br ; Norma Campos SalgadoII; Arnaldo C. dos Santos CoelhoII
IDepartamento de Ciências Biológicas, Pontifícia Universidade Católica de Minas Gerais. Avenida Dom José Gaspar 500, Coração Eucarístico, 30535-616 Belo Horizonte, MG, Brazil
IIDepartamento de Invertebrados, Museu Nacional, Universidade Federal do Rio de Janeiro. Quinta da Boa Vista, São Cristóvão, 20940-040 Rio de Janeiro, RJ, Brazil
ABSTRACT
Two new species of Thaumastus (Thaumastus) from the state of Minas Gerais, southeastern Brazil, are described. They are diagnosed and characterized by the morphology of the shell and soft parts and compared with Brazilian species of the subgenus Thaumastus s.s. Martens, 1860. Thaumastus (T.) parvus sp. nov. is similar to T. (T.) baixoguanduensis Pena, Coelho & Salgado, 1996 but can be distinguished by the smaller size, smaller number of plates in the jaw, different number of follicle groups in the ovotestis and form of the fertilization complex. Thaumastus (T.) caetensis sp. nov. is similar to T. (T.) largillierti (Philippi, 1845) but differs by the width and the contour of the parietal side of the shell aperture. T. (T.) caetensis is also closer to Thaumastus (T.) baixoguanduensis but can be distinguished by the smaller dimensions (height, width and number of protoconch whorls), the lack of a transversal light band on the body whorl, the jaw with smaller number of plates, and the radula with 35 teeth. In the soft parts, this new species differs also in the number of follicle gatherings in ovotestis, fertilization complex with globose shape, and penian retractor muscle terminally and laterally attached to flagellum.
Key words: Mollusca; morphology; Orthalicoidea; taxonomy.
This article is part of a large project on Brazilian Orthalicoidea (formerly Bulimuloidea according to BOUCHET & ROCROI 2005). The project expands the characterization of the species and knowledge of the genera and species included in this superfamily.
The specimens herein studied were collected from 1994 to 2001, in the state of Minas Gerais. After consulting published descriptions and comparing their morphology with other species of Thaumastus, we concluded that this material represented two new species.
Currently, five subgenera are recognized in Thaumastus, besides Thaumastus s.s., T. (Scholvienia), T. (Kara), T. (Quechua), and T. (Atahualpa) described by STREBEL (1910), and T. (Thaumastiella) described by WEYRAUCH (1956). BREURE (1979) stated that members of the subgenus Thaumastus are the only ones known from Brazil.
The Brazilian species of Thaumastus were characterized by JURBERG et al. (1979, 1988), BARROS et al. (1991), SALGADO & COELHO (1995), SALGADO et al. (1995), PENA et al. (1996, 2005). The intention of our research on species of Thaumastus is to re-characterize its subgenera as well as to provide a basis for a future phylogenetic analysis of the genus.
MATERIAL AND METHODS
The material studied consists of shells and specimens collected in the Serra do Curral in the municipalities of Belo Horizonte and Nova Lima, and in the Serra da Piedade, municipality of Caeté, state of Minas Gerais. It is housed in the mollusc collections of the Museu Nacional, Universidade Federal do Rio de Janeiro (MNRJ), Museu de Zoologia, Universidade de São Paulo (MZSP), Muséum National d'Histoire Naturelle (MNHN), and Zoologisches Museum der Universität Hamburg (ZMH).
All identifications were made by comparisons with type material and adult shells housed in the following museums: Academy of Natural Sciences of Philadelphia (ANSP), Forschungsinstitut und Naturmuseum Senckenberg (SMF), Frankfurt am Main, and Museum für Naturkunde der Humboldt-Universität (MNHU), Berlin. Original descriptions and illustrations were also studied.
The morphology of the shell and the reproductive system were examined in accordance with methods used by JURBERG et al. (1988) and SALGADO et al. (1995). The terminology used in the description of the protoconch follows that proposed by STREBEL (1910). Shells were measured with a Kanon Caliper rule:total height (h), width (w), height and width of aperture (ha, wa). The number of whorls was determined according to PARODIZ (1951) and the colors, as codified by KÜPPERS (1979). Radulas and jaws were prepared following JURBERG's (1964) technique. Shells and soft parts were studied and illustrated using a Wild M5 stereoscopic microscope and a Wild M20 compound microscope equipped with a drawing tube. Photos of shells were taken using a Canon F1 digital camera with a Canon macro lens 50. Scanning electron photomicrographs were made with a Zeiss DMS 950.
TAXONOMY
Thaumastus Martens, 1860
Type-species: Bulimus hartwegi Pfeiffer, 1846, by original designation.
Type-locality: Loja, Ecuador.
Thaumastus (Thaumastus) parvus, sp. nov.Figs 1-10
Diagnosis. Shell fragile, short. Dimensions: h: 35-42 mm; w: 14.4-18.7 mm; ha: 15.8-21.55 mm; wa: 9.15-12.05 mm; 5  -6¼ whorls slightly convex; aperture oval-elongated with very acuminate angle formed by outer lip and parietal margin; outline rounded in basal region. Jaw with 12-13 irregular plates and radula with 39 lateral teeth per half row. Reproductive system: seven follicle groups in ovotestis; deferent duct emerging above bifurcation of bursa duct and free oviduct and then penetrating under proximal phallus muscular sheath; ample phallus, retractor muscle attached to lateral and terminal flagellum areas.
-6¼ whorls slightly convex; aperture oval-elongated with very acuminate angle formed by outer lip and parietal margin; outline rounded in basal region. Jaw with 12-13 irregular plates and radula with 39 lateral teeth per half row. Reproductive system: seven follicle groups in ovotestis; deferent duct emerging above bifurcation of bursa duct and free oviduct and then penetrating under proximal phallus muscular sheath; ample phallus, retractor muscle attached to lateral and terminal flagellum areas.
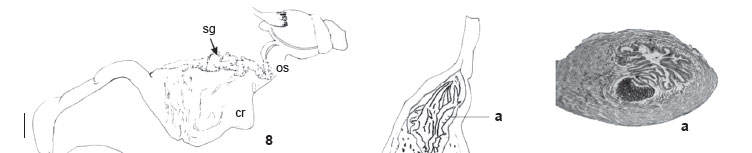
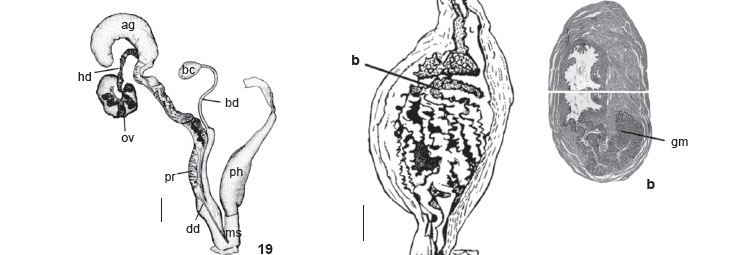
Description. Shell (Figs 1 and 2) short, fragile, oval-elongated, yellowish-brown (KÜPPERS: N40A70M30) with narrow and wide axial reddish-brown (N60A90M60) bands, 5-6¼ whorls, apex truncated. Protoconch (Fig. 3) with narrow, slightly lowered nucleus, fine oblique striae, evident suture, 2½-2  whorls. Teleoconch with thin, glossy, adhered periostracum, body whorl convex, suture slightly marked, and oblique columellar callus. Dimensions (57 shells of type-series): h: 35-42.8 mm; w: 14.4-18.7 mm; ha: 15.8-21.55 mm; wa: 9.15-12.05 mm. Mantle border (Fig. 4) with thick outer lobe (ol) and regular outline; inner lobe (il) restricted to area around pneumostome (pn) circula. Pallial complex (Fig. 5) translucent, slightly pigmented near pericardium; grayish-brown kidney (ki) with pericardium (pe) coupled with one of its sides; evident venation along pulmonary vein (pv) and secondary ureter; secondary ureter (su) running externally and alongside rectum. Digestive system: jaw (Fig. 6) with 12-13 irregular plates (central region ones narrower than lateral ones, sparsely pigmented with reddishbrown). Radula (Fig. 7) with 39 lateral teeth per half row; central tooth with trapezium-like base, rounded median and lateral cusps, partially symmetrical. One lateral tooth, beginning from L1, one cusp becomes reduced whereas other becomes tapered. (Cusps reduced from L24 on, leaving only triangle-like base behind.) Digestive tube (Fig. 8) with short anterior esophagus (os), crop (cr) significantly dilated and pleated with dorsal salivary glands (sg); posterior esophagus narrow, tubular, with length similar to crop length; stomach (st) C-shaped and tubular with slender and translucid walls. Reproductive system (Fig. 9): ovotestis (ov) with seven follicle groups; pleated hermaphrodite duct (hd), more pigmented near lobed albumen gland (ag); fertilization complex (fc) elongated at free extremity and dilated and bending over fertilization sac near hermaphrodite duct; uterus long with few pleats; prostate (pt) quite evident, white and slightly yellow, with some lateral folds; bursa duct (bd) long with constriction that reduces its diameter slightly before reaching halfway through its course; bursa copulatrix (bc) ovate; deferent duct (dd) emerging slightly above bifurcation of bursa duct and short, free oviduct, running adhered to it, bending to enter under phallus muscular sheath (ms), following course of developed robust phallus (ph) and then entering near extremities of long flagellum (fl) with external epiphallus (ep); phallus retractor muscle (rm) with lateral and terminal attachments. Histologically and microanatomically (Fig. 10 a, b, c), this complex of diverse areas consists of longitudinal muscular-epithelium folds (pilasters), glandular masses, muscular layers and different position of deferent duct lumen. These areas include: flagellum-epiphallus (limit) (Fig. 10a) with unilateral group of longitudinal high pilasters and glandular mass reducing lumen; epiphallus (Fig. 10b) with low pilasters and central lumen; phallus (Fig. 10c) with four high pilasters reducing central lumen.
whorls. Teleoconch with thin, glossy, adhered periostracum, body whorl convex, suture slightly marked, and oblique columellar callus. Dimensions (57 shells of type-series): h: 35-42.8 mm; w: 14.4-18.7 mm; ha: 15.8-21.55 mm; wa: 9.15-12.05 mm. Mantle border (Fig. 4) with thick outer lobe (ol) and regular outline; inner lobe (il) restricted to area around pneumostome (pn) circula. Pallial complex (Fig. 5) translucent, slightly pigmented near pericardium; grayish-brown kidney (ki) with pericardium (pe) coupled with one of its sides; evident venation along pulmonary vein (pv) and secondary ureter; secondary ureter (su) running externally and alongside rectum. Digestive system: jaw (Fig. 6) with 12-13 irregular plates (central region ones narrower than lateral ones, sparsely pigmented with reddishbrown). Radula (Fig. 7) with 39 lateral teeth per half row; central tooth with trapezium-like base, rounded median and lateral cusps, partially symmetrical. One lateral tooth, beginning from L1, one cusp becomes reduced whereas other becomes tapered. (Cusps reduced from L24 on, leaving only triangle-like base behind.) Digestive tube (Fig. 8) with short anterior esophagus (os), crop (cr) significantly dilated and pleated with dorsal salivary glands (sg); posterior esophagus narrow, tubular, with length similar to crop length; stomach (st) C-shaped and tubular with slender and translucid walls. Reproductive system (Fig. 9): ovotestis (ov) with seven follicle groups; pleated hermaphrodite duct (hd), more pigmented near lobed albumen gland (ag); fertilization complex (fc) elongated at free extremity and dilated and bending over fertilization sac near hermaphrodite duct; uterus long with few pleats; prostate (pt) quite evident, white and slightly yellow, with some lateral folds; bursa duct (bd) long with constriction that reduces its diameter slightly before reaching halfway through its course; bursa copulatrix (bc) ovate; deferent duct (dd) emerging slightly above bifurcation of bursa duct and short, free oviduct, running adhered to it, bending to enter under phallus muscular sheath (ms), following course of developed robust phallus (ph) and then entering near extremities of long flagellum (fl) with external epiphallus (ep); phallus retractor muscle (rm) with lateral and terminal attachments. Histologically and microanatomically (Fig. 10 a, b, c), this complex of diverse areas consists of longitudinal muscular-epithelium folds (pilasters), glandular masses, muscular layers and different position of deferent duct lumen. These areas include: flagellum-epiphallus (limit) (Fig. 10a) with unilateral group of longitudinal high pilasters and glandular mass reducing lumen; epiphallus (Fig. 10b) with low pilasters and central lumen; phallus (Fig. 10c) with four high pilasters reducing central lumen.
Holotype. BRAZIL, Minas Gerais: Belo Horizonte, Serra do Curral (19º57'15.00"S; 43º55'21.77"W), Mangabeiras Park, MNRJ 8107 (h: 42.8 mm; w: 16.2 mm; ha: 18.9 mm; wa: 11.4 mm), M.S. Pena, R.A.F. Cajá & L.M.A. Azevedo leg., I/1998. Paratypes: from the same locality as the holotype, MNRJ 8106, 4 shells (2 juv.), 1 specimen, 1 protoconch (on stub), 2 jaws, 4 soft parts, 1 roof of pallial cavity, 1 reproductive system, penian complex (21 histological slides), ovotestis (7 histological slides), M.S. Pena, L.M.A. Azevedo & R.A.F. Cajá leg., I/1998; MNRJ 8108, 32 shells (8 juv.); M.S. Pena, L.M.A. Azevedo & R.A.F. Cajá leg., I/1998; ANSP (unnumbered), 1 shell, M.S. Pena, L.M.A. Azevedo & R.A.F. Cajá leg., I/1998; MZSP 39387, 1 shell; ZMH 2960, 3 shells; MNRJ 8103, 8 shells (3 juv.), 1 roof of pallial cavity, 1 reproductive system, I. R.R. Dutra leg., 1994; MNRJ 8104, 2 shells; MNRJ 8105, 9 shells (1 juv.), M.S. Pena leg., XI/1996. Municipality of Nova Lima, Belo Horizonte Peak; MNHN (unnumbered), 1 shell, M.S. Pena leg. Municipality of Nova Lima, Belo Horizonte Peak; MNRJ 9810, 3 shells, 3 jaws, 3 soft parts, R. Soares & R.R. Dutra leg., XI/ 1994. Municipality of Belo Horizonte, Rola Moça Park; MNRJ 8109, 2 shells, M.S. Pena & L.M.A. Azevedo leg., I/1998; MNRJ 8102, 3 roofs of pallial cavities, 3 reproductive systems, 3 buccal masses, I.R.R. Dutra leg., XI/1994; MNRJ 9811, 4 specimens, M.S. Pena leg., XII/1997.
Etymology: From the Latin parvus, short, small, referring to the smallest known Brazilian Thaumastus species.
Remarks. The following diagnostic characters allowed us to include T. (T.) parvus sp. nov. in the genus and subgenus Thaumastus: shell elongated-ovate, light to dark brown with axial streaks, light spiral band, convex whorls; protoconch with fine axial striae groups that are closely set, suture evident, aperture subovate, peristome slightly expanded. And also, the reproductive system consisting of sub-cylindrical bursa duct with globose bursa copulatrix at its distal end. The deferent duct running adhered to free oviduct and bending to enter underneath the proximal sheath of penian complex. Penis changing into epiphallus without any external differentiation. The shells in T. (T.) parvus sp. nov. and T. (T.) baixoguanduensis are similar because of their elongated shape, pattern of striae in the protoconch, color, and shape of the aperture. The number of teeth in the radula is also similar. The reproductive systems share a similar shape of the bursa copulatrix, constriction in the median region of the bursa duct, deferent duct running under the phallus muscular sheath, as well as lateral and terminal attachment of the phallus retractor muscle. Nonetheless, T. (T.) baixoguanduensis differs from the new species in having larger dimensions (h: 50-56.10 mm, w: 23.02-25.9 mm), prominent nucleus in the protoconch, crenulated suture in the body whorl, thick peristome, developed columellar callus with rectilinear contour, as well as slight depletion and more plates in the jaw (14). The reproductive system consists of a different number of follicle groups (6) in the ovotestis, a round free extremity of the fertilization complex, a deferent duct emerging at the same level as the bifurcation of the bursa duct, free oviduct, and a long arched phallus with irregular diameter.
Thaumastus (Thaumastus) caetensis sp. nov. Figs 11-21
Diagnosis. Shell thick. Dimensions: h: 47-56.4 mm; w: 17-22 mm; ha: 18.5-23.5 mm; wa: 10.55-14.9 mm; protoconch with straight nucleus and 2½-2¾ whorls; outer lip projected forward. Jaw with 12 plates and radula with 35 lateral teeth on half row. Reproductive system: ovotestis with five follicle groups; fertilization complex distended in proximal portion to the hermaphrodite canal; bursa copulatrix duct with constriction on its middle portion that reduces to  its initial diameter; deferent duct penetrating under penis sheath.
its initial diameter; deferent duct penetrating under penis sheath.
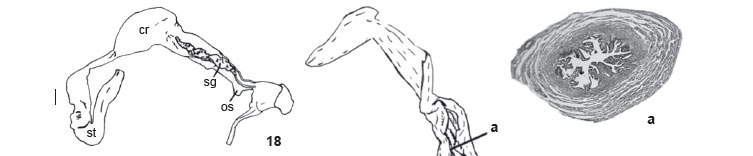
Description. Shell (Figs 11-12): Thick, oval-lengthened, 6½ to 7 whorls, apex truncated; yellowish-brown (N60A80M50) with wide and narrow axial reddish-brown (N60A90M60) bands irregularly alternated; protoconch (Fig. 13) with oblique striae slightly undulated and straight nucleus, and 2½-2¾ whorls. Teleoconch with axial thickened lines; thin, shiny and adhered periostracum; convex body whorl with slightly evident suture; aperture oval-lengthened, smaller than half of total shell height, roundly and large at base and sharp at apex; thin and white peristome; columellar callus straight and inclined; outer lip projected over parietal region in about of diameter of body whorls. Dimensions (27 shells of type-series): h: 47-56.4 mm; w: 17-22 mm; ha: 21.45-24.45 mm; wa: 10.55-14.9 mm; number of whorls: 6½. Mantle border (Fig. 14) with well-developed outer lobe (ol) and forming pneumostome (pn) on a split surface from which emerges the short inner lobe (il), shaped like a small fold. Pallial complex with roof of pallial cavity (Fig. 15) translucent, with evident vascularization and pulmonary vein bifurcated in anterior region; gray kidney, almost triangular; evident secondary ureter. Pericardium size corresponding to size of renal face where it is located. Digestive system: jaw (Fig. 16) formed by 12 irregular juxtaposed plates with variable pigmentation on border. Radula (Fig. 17) with 35 lateral teeth on a half row. Central tooth with trapezium-like base, a rounded median cusp and two symmetric and sharplypointed outer cusps. On lateral teeth, beginning from L1 the symmetry is gradually substituted by a strong reduction of one outer cusp and sharpening of the other. Digestive tube (Fig. 18): esophagus relatively long and posteriorly inserted to the buccal bulb with laterodorsal salivary glands. Crop with dorsal dilation at final portion of salivary glands. Posterior esophagus narrower in proximal crop and wider near tubular stomach. Internally, digestive tube with straight, shallow folds in dorsal region at beginning of crop, with accentuated bordering in cuticle belonging to narrowest region of posterior esophagus. Reproductive system (Fig. 19): ovotestis formed by lengthened follicles gathered on five pink groups of variable size; hermaphrodite duct very folded, varying in diameter, darkbrown pigmented; fertilization complex (Fig. 20) embedded into the concave face of albumen gland with free extremity and proximal portion of hermaphrodite duct distended; albumen gland with translucent or white lobules wrinkled in some parts; uterus white and pleated; prostrate globose with lateral pleating in some parts; oviduct origin slightly upon the bursa copulatrix duct bifurcation. Bursa duct with a regular diameter in the first half, then with unilateral constriction reducing it in the last half; bursa copulatrix globose. Deferent duct emerges from end of prostate, slightly upon oviduct bifurcation, follows adhered until the penial complex, where it folds and enter under the phallus muscular sheath and flagellum. Phallus more distended in region starting from muscular sheath; retractor muscle with terminal attachment to flagellum. Internally, the penian complex (Fig. 21 a, b, c) has its interior aperture (lumen) narrowed by many longitudinal wrinkles, short and distended at apical portion of penian complex and taller and folded at medium portion. Penian complex regions can be recognized in transversal histological cuts by the flagellum (Fig. 21a) with a narrow lumen partially filled by muscular pilasters. Epiphallus (Fig. 21b) with glandular area strongly developed and covering the deferent duct in the lumen side. On phallus region (Fig. 21c) the lumen is densely filled by many wrinkled pilasters. Vagina proximal portion with low pilasters limiting straight lumen and surrounded by one muscular tunic that involves the deferent duct.
Holotype: BRAZIL, Minas Gerais: Caeté (19º49'19"S; 43º40' 46"W), Serra da Piedade, MNRJ 8097 (h = 56.4 mm; w = 21.5 mm; ha = 23.5 mm; wa = 14.9 mm), M.S. Pena & R.R. Dutra leg., XI/1994. Paratypes: from the same locality of the holotype, MNRJ 8098, 3 shells, 1 roof of pallial cavity; 1 mantle border; 3 reproductive systems, M.S. Pena & I.R.R. Dutra leg., XI/1994; ANSP (unnumbered), 1 shell, M.S. Pena & I.R.R. Dutra leg., XI/1994; MNRJ 8099, 2 shells, 5 jaws, 2 roofs of pallial cavity, 2 reproductive systems, 4 ovotestis (4 histological slides), M.S. Pena, L.M.A. Azevedo & R.A.F. Cajá leg., I/1998; MNRJ 8100, 1 shell, M.S. Pena, L.M.A. Azevedo & R.A.F. Cajá leg., III/1998; MNRJ 8101, I specimen without buccal mass, 1 radula (on stub), M.S. Pena, L.M.A. Azevedo & R.A.F. Cajá leg., III/1999; MNRJ HSL7810, 1 shell, J. Evangelista leg., 6/VI/1960; MNRJ 3115, 3 shells, P.E. Vanzolini leg., II/1961; MNRJ 9806, 12 specimens (1 juv.); 1 soft part without penian complex; 1 soft part dissected; 2 jaws, 1 roof of pallial cavity, 1 reproductive system; 1 protoconch (on stub); penian complex (24 histological slides), M.S. Pena leg., XII/2001; MZSP 39388, 1 shell; ZMH 2966, 1 shell, M.S. Pena, L.M. Azevedo & R. Cajá leg., I/ 1998; MNHN (unnumbered), 2 shells, M.S. Pena, L.M. Azevedo & R.A.F. Cajá leg., I/1998.
Etymology. Name from Caeté municipality, type locality of the described material, where the Serra da Piedade mountain is located.
Remarks. Thaumastus (Thaumastus) caetensis sp. nov. and T. (T.) largillierti (Philippi, 1845) are similar in the number of whorls, the presence of slender peristomes, the color pattern, and the elongate outline. Thaumastus (T.) largillierti differs from the new species in having 23 mm of width, peristome with rectilinear contour at the parietal side, and larger thickness of the columellar callus. Thaumastus (T.) caetensis sp. n. is also closer to T. (T.) baixoguanduensis by presenting a yellowish brown color and reddish-brown streaks, oblique striae, and outer lip projected over the parietal region; in the soft parts, the two species show the same pathway of the deferent duct under the phallus muscular sheath, as well as the globular shape of bursa copulatrix and terminal insertion of the retractor muscle of phallus on flagellum. However, T. baixoguanduensis differs from the new species by the following features: smaller shell (maximum 52.5 mm); larger width (up to 25 mm); suture well-margined; columellar callus with internal outline flat and surface depressed; protoconch with higher number of whorls (3 ) and a transversal light band on the body whorl; jaw with 14 plates; greater number of teeth on the radula (36-39); 6 follicle gatherings in ovotestis; fertilization complex with globose free extremity; penian retractor muscle terminally and laterally attached to flagellum.
) and a transversal light band on the body whorl; jaw with 14 plates; greater number of teeth on the radula (36-39); 6 follicle gatherings in ovotestis; fertilization complex with globose free extremity; penian retractor muscle terminally and laterally attached to flagellum.
ACKNOWLEDGEMENTS
We are grateful to Júlio C. Monteiro (Malacologia, Museu Nacional/UFRJ) for drawings of the roof of pallial cavities and reproductive systems and to Paulo M. Costa for taking shell photos.
LITERATURE CITED
Submitted: 13.X.2010; Accepted: 24.IV.2011.
Editorial responsibility: Gabriel L.F. Mejdalani
- BARROS, H.M; A.C.S. COELHO & N.C. SALGADO. 1991. Superfamília Bulimuloidea do Brasil. Bulimulidae: Thaumastus (Thaumastus) magnificus (Grateloup, 1839) (Mollusca, Gastropoda, Pulmonata). Boletim do Museu Nacional, Zoologia 348:1-18.
- BOUCHET, P. & J.P. ROCROI. 2005. Classification and nomenclator of gastropod families. Malacologia 47(1-2):1-397.
- BREURE, A.S.H. 1979. Systematics, phylogeny and zoogeography of Bulimulinae (Mollusca, Gastropoda, Pulmonata) II. Zoologische Verhandelingen 168:3-215.
- JURBERG, P. 1964. Sobre Auris bilabiata melanostoma (Moricand, 1838) (Gastropoda, Pulmonata, Bulimulidae). Memórias do Instituto Oswaldo Cruz 62:81-94.
- JURBERG, P.; L.A.L. GOMES; H.M. BARROS & A.C.S. COELHO. 1979. Superfamília Bulimuloidea do Brasil. Bulimulidae: Thaumastus (Thaumastus) achilles (Pfeiffer, 1852) (Mollusca, Gastropoda, Pulmonata). Anais do V Encontro dos Malacologistas Brasileiros. In: Publicações Avulsas da Fundação Zoobotânica 4:21-32.
- JURBERG, P.; H.M. BARROS; L.A.L. GOMES & A.C.S. COELHO. 1988. Superfamília Bulimuloidea do Brasil. Bulimulidae: Thaumastus (Thaumastus) taunaisii (Férussac, 1822), com dados biológicos e aspectos comportamentais (Mollusca, Gastropoda, Pulmonata). Boletim do Museu Nacional, Zoologia 347:1-40.
- KÜPPERS, H. 1979. Atlas de los colores Barcelona, Blume Editorial, 161p.
- MARTENS, E. 1860. In: J.C. ALBERS & E. MARTENS. Die Helicee nach natürlicher Verwandtschaft systematisch geordnet. Leipzig, W. Engelmann, 2nd ed. XVIII+359p.
- PARODIZ, J.J. 1955. Métodos de Conquiliometria. Physis: Revista de la Asociación Argentina de Ciéncias Naturales 58(20):241-248.
- PENA, M.S.; A.C.S. COELHO & N.C. SALGADO. 1996. Bulimuloidea do Brasil. Bulimulidae: Duas novas espécies de Thaumastus Martens in Albers, 1860 (Mollusca, Gastropoda, Pulmonata). Boletim do Museu Nacional, Zoologia 368:1-12.
- PENA, M.S.; N.C. SALGADO & A.C.S. COELHO. 2005. New species of Thaumastus from Lagoa Santa region, Minas Gerais state, Brazil (Mollusca, Gastropoda, Pulmonata, Bulimuloidea). Iheringia, Série Zoologia 95(1):21-24.
- PFEIFFER, L. 1846-1847. Bulimus. In: PHILIPPI, R.A. 1845-1847. Abbildungen und Beschreibungen neuer oder wenig bekannter Conchylien. Cassel, Theodor Fischer, vol. 2, p. 111-115.
- PHILIPPI, R.A. 1845. Abbildungen und Beschreibungen neuer oder wenig bekannter Conchylien. Cassel, Theodor Fischer, 164p.
- SALGADO, N.C. & A.C.S. COELHO. 1995. Macroanatomia, microanatomia e histologia do sistema digestivo de Thaumastus (Thaumastus) taunaisii (Férussac, 1822), T. (T.) magnificus (Grateloup, 1839) e T. (T.) achilles (Pfeiffer, 1852). Boletim do Museu Nacional, Zoologia 362:1-19.
- SALGADO, N.C.; A.C.S. COELHO & H.M. BARROS. 1995. Macroanatomia, microanatomia e histologia do sistema reprodutor de Thaumastus (Thaumastus) taunaisii (Férussac, 1822), T. (T.) magnificus (Grateloup, 1839) e T. (T.) achilles (Pfeiffer, 1852) (Mollusca, Gastropoda, Bulimulidae). Boletim do Museu Nacional, Zoologia 358:1-47.
- STREBEL, H. 1910. Conchologische Mitteilungen aus dem Naturhistorischen Museum in Hamburg. Abhandlungen aus dem Gebiete der Naturwissenschaften 19:1-35.
- WEYRAUCH, W. 1956. The genus Naesiotus, with descriptions of new species and notes on other Peruvian Bulimulidae. Proceedings of the Academy of Natural Sciences of Philadelphia 158:1-8.
Publication Dates
-
Publication in this collection
16 Sept 2011 -
Date of issue
Aug 2011
History
-
Accepted
24 Apr 2011 -
Received
13 Oct 2010