Abstract
It is time for an expansion and enrichment of evolutionary theory. The "back to the future" proposal contained herein is based on three postulates: 1) Neo-Darwinism is too impoverished for this task; 2) its predecessor, Darwinism, contained the necessary breadth of vision and metaphor to be the basis for an inclusive and unifying theory of biology; and 3) the necessary framework for this new stage in the evolution of evolutionary theory is largely in place. We make our case through the use of a number of metaphorical dualisms designed to help focus discussions toward a more cooperative and productive approach to the study of living systems. Along the way, we suggest a number of self-induced paradoxes in neo-Darwinian accounts of evolution that are resolved by our perspective.
Darwinism; nature of the organism; nature of the conditions; extended synthesis; metaphors; time, sequency; simultaneity; thermodynamics; metabolism; space; information; function; history; complexity; cohesion; compensatory changes; evolutionary transitions; cooperation; division of labor; ecological hierarchy; genealogical hierarchy; self-organization
INVITED REVIEW
Children of time: the extended synthesis and major metaphors of evolution
Daniel R. BrooksI, * * Corresponding author. E-mail: dan.brooks@utoronto.ca ; Salvatore J. AgostaII
ISchool of Biological Sciences, University of Nebraska-Lincoln, Lincoln NE 68588-0514, USA
IICenter for Environmental Studies and Department of Biology, Virginia Commonwealth University, Richmond VA 23284-3050, USA
ABSTRACT
It is time for an expansion and enrichment of evolutionary theory. The "back to the future" proposal contained herein is based on three postulates: 1) Neo-Darwinism is too impoverished for this task; 2) its predecessor, Darwinism, contained the necessary breadth of vision and metaphor to be the basis for an inclusive and unifying theory of biology; and 3) the necessary framework for this new stage in the evolution of evolutionary theory is largely in place. We make our case through the use of a number of metaphorical dualisms designed to help focus discussions toward a more cooperative and productive approach to the study of living systems. Along the way, we suggest a number of self-induced paradoxes in neo-Darwinian accounts of evolution that are resolved by our perspective.
Key words: Darwinism; nature of the organism; nature of the conditions; extended synthesis; metaphors; time, sequency; simultaneity; thermodynamics; metabolism; space; information; function; history; complexity; cohesion; compensatory changes; evolutionary transitions; cooperation; division of labor; ecological hierarchy; genealogical hierarchy; self-organization.
Modern science labors under a self-imposed duality. One part is highly conservative. A working framework is accepted as true, or at least provisionally true, and research programs elaborate ways in which that framework is useful. All such pursuits have shown that our frameworks are either false or incomplete. This necessitates the second part of the duality, finding alternatives that are less false or less incomplete. Reflecting this duality, scientists use two kinds of language: nomenclature (labels) and metaphor. Scientists use nomenclature to reduce ambiguity within a theoretical framework assumed to be fundamentally true. If a North American and a European call a bird a "robin," they are referring to two distantly related species. If, however, they both say Turdus migratorius, there is no confusion.
Many scientists mistrust metaphors because they allow too many possible meanings, introducing unwanted ambiguity. Scientific change, however, is creative, and that requires metaphorical language. By extending existing nomenclature to accommodate new concepts and empirical findings, metaphors give people a reason to learn new ideas and the means to learn them. And finally, metaphors present natural truths using language that allows understanding by non-specialists. Darwin masterfully linked everyday experience and knowledge with technical observations through metaphors. The widespread acceptance of Darwinism, however, shifted the language of evolution from metaphor to nomenclature. Today's normative framework for evolutionary biology is neo-Darwinism, or the Synthetic Theory of Evolution. ELDREDGE (1985, 1995) proposed the heterodox view that neo-Darwinism is a narrowing of Darwinism that neglects much of Darwin's panoramic vision. BROOKS (2010, 2011a,b,c) detailed a number of important differences between neo-Darwinism and Darwinism. At present, there is considerable discussion among evolutionary biologists about the need for an "Extended Synthesis" (e.g., PIGLIUCCI & MÜLLER 2010a; see contributions in PIGLIUCCI & MÜLLER 2010b) but there is no consensus on what shape it will take.
The diversity of viewpoints about evolution parallels the diversity of life. In this review, we will be provocative in an attempt to make readers feel ambivalent about their own views of evolution. We do not believe that theoretical advances will emerge from ecclesiastical dialogues among different entrenched viewpoints in which each side expects the other to convert at some point, until an eventual winner emerges. Rather, we think it will require biologists to learn how they think about evolution by learning how others see their views (FISCH & BENBAJI 2011). Metaphor is essential for this.
Fortunately, metaphors cannot disappear-forgotten or set aside, they can always be recovered and reexamined in light of new information or challenges to an existing framework. So, we will return to the metaphors elaborated by Darwin and neglected during the development of the New Synthesis. Then we will discuss briefly the notion that Herbert Spencer's influence initiated neo-Darwinism. Next, we show how Darwin's metaphors provide a common link among some late 20th century reactions against the "hardened synthesis." We then present new metaphors aimed toward allowing researchers to connect Darwin's view of evolution, proposals from the 1980s and 1990s, and calls for an Extended Synthesis.
BACK TO THE FUTURE: DARWINISM AS THE FIRST COMPLEXITY THEORY
The belief that neo-Darwinism is a refinement of Darwinism begins with the final sentence of the sixth chapter of the Origin of Species:
"Hence in fact the law of the Conditions of Existence is the higher law; as it includes, through the inheritance of former adaptations, that of Unity of Type." - Darwin, 1872: 195
Many consider the law of the Conditions of Existence to be natural selection, but BROOKS (2010, 2011a) recently suggested this alternative
"... there are two factors: namely, the nature of the organism and the nature of the conditions. The former seems to be much more the important; for nearly similar variations sometimes arise under, as far as we can judge, dissimilar conditions; and, on the other hand, dissimilar variations arise under conditions which appear to be nearly uniform." - Darwin, 1872: 32
Darwin believed that organisms produced offspring similar but not identical to each other; transmitted those similarities and differences to their offspring; and acted in their own behalf. Mostly importantly, those capacities held regardless of the Nature of the Conditions. This is Darwin's Necessary Misfit (BROOKS & HOBERG 2007, BROOKS 2010, 2011a). Without substantial autonomy from the surroundings, there could be no reproductive overrun, hence no struggle for survival, thus no natural selection. Natural selection was the outcome of conflicts created a priori by the conditions of existence. It was a consequence of the higher law. The final paragraph of the sixth chapter of the Origin supplies the essential context
"It is generally acknowledged that all organic beings have been formed on two great laws [our bold] - unity of type and the conditions of existence...On my theory, unity of type is explained by unity of descent. The expression [our bold] of conditions of existence...is fully embraced by the principle of natural selection..."Hence in fact the law [our bold] of the Conditions of Existence is the higher law; as it includes, through the inheritance of former adaptations, that of Unity of Type." - Darwin, 1872: 194-195.
The higher law encompasses all interactions between the nature of the organism and the nature of the conditions. The "law of the conditions of existence" is metaphorical, whereas the "nature of the conditions" and "expression of the conditions of existence" refer to the circumstances in which conflicts of interest between organisms take place (MAYNARD SMITH & SZATHMÀRY 1995).
Darwin lacked mechanisms of inheritance and ontogeny, yet understood that organisms were genealogically and developmentally cohesive. It was in the nature of the organism to produce offspring that were all highly similar to each other and their parents and other ancestors. He also postulated that reproduction produced variation without regard for environmental conditions, and therefore it was in the nature of the organism to produce offspring in numbers far exceeding the resources available for their support. This cannot happen in a Panglossian (Lamarckian) world, so there must be constraints on responses to the surroundings. Darwin resolved this conundrum by postulating that the nature of the organism created those constraints. And yet, they are not absolute. All reproducing organisms have positive Darwinian fitness, but some are fitter than others in their particular environments, where they predominate numerically over their merely adequate relatives. However selection-challenged, those relatives survive and play decisive evolutionary roles. When the conditions change, the fittest in the old environment might not survive at all, whereas some of the merely adequate might flourish. Natural selection was thus an emergent property of the inevitable conflict created by the conditions of existence and also a metaphor for the ways in which such conflicts are resolved, setting the stage for resolution of conflicts yet to come.
Darwin believed evolution was an outcome of interactions between two classes of phenomena, each following their own rules yet spatially and temporally entwined. Furthermore, the two classes of phenomena were not co-equal, the more important inevitably creating conflict, the "lesser" leading to resolutions of those conflicts. In today's lexicon, Darwin's theory was clearly a theory of complex adaptive systems (JUARRERO 2002), but in his day, good theories were simple. Perhaps understanding how heterodox his views were, Darwin proposed two rich metaphors to help visualize his theory of the fundamental complexity of evolution.
"As buds give rise by growth to fresh buds, and these, if vigorous, branch out and overtop on all sides many a feebler branch, so by generation I believe it has been with the great Tree of Life, which fills with its dead and broken branches the crust of the earth, and covers the surface with its ever-branching and beautiful ramifications." - Darwin, 1872: 132
The Tree of Life metaphor is a symbol of a major part of the evolutionary process. Living systems are capable of acting in their own behalf, but more importantly, they regularly take the initiative, using what they have inherited. Metaphorically, the present is the state in which biological systems create their own futures based on their own pasts. Organisms carry so much of their history with them that most explanations for their appearance and function stem from their past-this is the focus of historical ecology (BROOKS & MCLENNAN 2002). Particular origins in space and time play integral roles in explaining the properties of organisms and the species they form, and how they interact with their surroundings, including other species. In Europe, "sycamore" is a maple (Acer pseudoplatanus) and "plane tree" (Platanus orientalis) is what North Americans call "sycamore" (Platanus occidentalis). Darwin's metaphor of natural classification being a phylogeny enables us to understand why North American sycamores and European plane trees resemble each other so closely, why their ecologies are so similar, why they hybridize so readily. Darwin's phylogenetic tree metaphor contrasted with a progressive view of diversity embodied in the Scala Naturae, underscoring the notion of evolution as one of selective accumulation of diversity rather than selective replacement.
"It is interesting to contemplate a tangled bank, clothed with many plants of many kinds, with birds singing on the bushes, with various insects flitting about, and with worms crawling through the damp earth, and to reflect that these elaborately constructed forms, so different from each other, and dependent upon each other in so complex a manner, have all been produced by laws acting around us. These laws, taken in the largest sense, being Growth with reproduction; Inheritance which is almost implied by reproduction; Variability from the indirect and direct action of the conditions of life, and from use and disuse; a Ratio of Increase so high as to lead to a Struggle for Life, and as a consequence to Natural Selection, entailing Divergence of Character and the Extinction of less improved forms. Thus, from the war of nature, from famine and death, the most exalted object which we are capable of conceiving, namely, the production of the higher animals, directly follows. There is grandeur in this view of life, with its several powers, having been originally breathed by the Creator into a few forms or into one; and that, whilst this planet has gone circling on according to the fixed law of gravity, from so simple a beginning endless forms most beautiful and most wonderful have been, and are being evolved." - Darwin 1872: 459
The "Tangled Bank" statement evokes selective accumulation of diversity producing complex ecosystems. It also reinforces Darwin's view that natural selection is an emergent property.
NEO-DARWINISM: THE TRIUMPH OF SPENCER OVER DARWIN
Bowler (1983) wrote an influential text about the "eclipse of Darwinism" and its end, which he linked to the rise of the Modern Synthesis. We are concerned with the origin of the eclipse. STOCKING (1982) noted that by 1871, anthropologists and sociologists considered "evolution" synonymous with progressive historical sequences. A sociologist of that period, Herbert Spencer, interpreted natural selection as a phenomenon in which the fittest out-competed their rivals. As "survival of the fittest" gained popularity, North American Lamarckian E.D. Cope raised the question (COPE 1887): "If evolution is survival of the fittest, what explains the origin of the fittest?"
Darwinians would have dismissed Cope's arguments, saying evolution was not survival of the fittest but survival of the adequate. "Spencerians," however, responded by linking "fitness" and "progress" and thus "adaptation" and "progress" (later "optimality"). In this way, natural selection could be construed as a creative and progressive process. We, therefore, trace the origin of neo-Darwinism to the Spencerian Heresy in the last quarter of the 19th century. An effective proponent of Spencer's view was August Weismann, and some Darwinians coined the derogatory term "neo-Darwinism" to refer to his views (HULL 1988).
Spencer's views were popular enough that Darwin tried to reinterpret Survival of the Fittest in metaphorical rather than nomenclatural terms. He was unsuccessful. GEORGE GAYLORD SIMPSON (1944), for example, characterized natural selection as a sculptor. With his metaphor of formless blocks of marble, Simpson exposed the fundamental difference between Darwinism and neo-Darwinism with respect to the nature of the organism. Darwinians would have characterized lineages of inheritance as the sculptors and natural selection as more an art critic than an artist.
For Darwin, inheritance introduced historical contingency into evolutionary explanations. This perspective was consistent with a Humean interpretation of history as a series of causal events that may influence the future without assuming it to be pre-determined; a perspective highlighted by Darwin's phylogenetic tree metaphor. Neo-Darwinism fit well with a more Hegelian view that history is a passive record of the emergence of inevitable events
"...biology is no longer simply a branch of history. It is now a science." - Morgan, 1932
Having eliminated the nature of the organism by making selection creative, neo-Darwinists killed time by eliminating phylogeny as explanatory.
To underscore our perspective, we present the following statements about evolution expressed by self-described neo-Darwinians, preceded by Darwin's views in the 6th edition of the Origin:
1) Evolution is the interplay of the nature of the organism and the nature of the conditions, the nature of the organism being far more important became evolution is adaptation by random variation to changing environments;
2) Phylogeny is a critical part of causal explanations became phylogeny is a passive record of past selection events (more recently, analogous to the error term in an ANOVA model);
3) Ecology is played out on an evolutionary stage became evolution is played out on an ecological stage;
4) Evolutionary outcomes are generally gradual, because evolution is the net outcome of organisms responding to many different aspects of the nature of the conditions (so many selection vectors) became evolution is inherently gradual;
5) Fitness space is "sloppy" (AGOSTA & KLEMENS 2008); most evolutionary dynamics are the result of organisms with non-zero fitness wandering through fitness space became fitness space is highly optimized with fuzzy boundaries, and organisms do not change fitness space without eliminating a less fit occupant;
6) High levels of variation are expected; all variants with non-zero fitness in the environments in which they are born survive became high levels of variation are not expected; the variant with the highest fitness replaces all others (if there is variation, all variants have the same fitness);
7) The conservative nature of inheritance and insensitivity of reproduction to the nature of the conditions produce more organisms needing the same resources than there are resources available became limited environmental resources create conflicts;
8) Survival of the adequate became survival of the fittest;
9) Survival (non-zero fitness) is paramount became optimality (maximum fitness) is paramount.
10) Necessity is the nature of the organism (material inheritance) and chance is the nature of the conditions became chance is the nature of the organism ("random variation") and necessity is the nature of the conditions.
11) Conflict is resolved by accommodation (division of labor sensu MAYNARD SMITH & SZATHMÀRY 1995) became old conflict is replaced by new conflict.
The "hardening of the synthesis" led to a simple view of evolution: function follows the nature of the conditions and form follows function, blurring the distinction between Darwinian and Lamarckian explanations (ELDREDGE 1985, 1995).
ENDING THE ECLIPSE: THE MAJOR METAPHORS OF EVOLUTION
Darwin's metaphors allow us to understand how Darwinism and neo-Darwinism have been conflated. They also clarify connections between Darwinism and some major critiques of neo-Darwinism in the 1980s and 1990s, such as The Hierarchy View (ELDREDGE 1985, 1995): Living systems are simultaneously part of an informational hierarchy of replicators (the Genealogical Hierarchy - Nature of the Organism) and an energetic hierarchy of interactors (the Ecological Hierarchy - Nature of the Conditions). The Information View (BROOKS & WILEY 1986, 1988): irreversibility in biological systems results from entropic increases in biological information, constrained by intrinsic (inheritance system informational dynamics - Nature of the Organism) and extrinsic (selection - Nature of the Conditions) properties. The Evolutionary Transitions View (MAYNARD SMITH & SZATHMÀRY 1995): evolutionary transitions increase the efficiency of storing and transmitting information (Nature of the Organism), thereby enhancing organism/environment interactions (Nature of the Conditions). All of these proposals are radical in the literal sense that they return to the roots of Darwinism.
MCSHEA & BRANDON (2011) and CALCOTT & STERELNY (2011) attempted to subsume these and related critiques of the 1980s and 1990s within neo-Darwinism. We believe they failed because neo-Darwinism is not rich enough metaphorically to accommodate the full diversity of life and life functions. We suggest three classes of metaphors (time, space and complexity) we believe can frame productive discussions about the future of evolutionary theory.
TIME
Everything that happens has a material cost. Physics treats this reality as an accounting system, where transactions can be measured in terms of the transformation of energy (most easily seen as heat loss) or of the movements of particles in the system affected by the transformation of energy. No matter the particular manifestation of these transformations, there is always a net cost, called "entropy." So, in every spontaneously occurring causal activity, entropy increases. This accounting can only be done retrospectively, using the temporal record produced by each causal event. Early thermodynamics limited the usable energy account of a system. When the usable energy inside a system was exhausted "equilibrium" had been reached, the system having achieved maximum entropy. Just as the Darwinian reality had inspired the unsavory vision of "Nature red in tooth and claw," this view of thermodynamics inspired the terrifying vision of the "Heat death of the universe." Ludwig von Boltzmann linked these views in a public lecture in Leipzig in 1905, proclaiming that the 19th century had been Darwin's century, and that Darwin's theory was the first step to a statistical mechanical theory of biology.
LOTKA (1913, 1925) characterized biological systems as metabolic systems, maintaining themselves in highly organized states by exchanging matter and energy with their surroundings. He suggested that the inevitable structural decay that accompanies such transactions could be delayed, although not reversed, by the system's accumulation of energy from outside to do work within the system. Organisms undergo heat-generating transformations, involving a net loss of energy from the system, and conservative transformations, changing free energy into states that can be stored and utilized in subsequent transformations. All conservative transformations in biological systems are coupled with heat generating transformations, but the reverse is not true; maintaining structure is expensive (BROOKS et al. 1989, BROOKS & MCLENNAN 1990, MAURER & BROOKS 1991).
For closed thermodynamic systems, once the matter inside the system is dispersed maximally, given the boundaries of its container, equilibrium is reached and the bank account is empty. All work ceases. Equilibrium systems show no duality in energy use, clearly inadequate for understanding biological systems. Open systems are those for which new energy and matter can flow through the system, allowing the system to continue to function so long as the flows continue. Total entropy changes (dS) (entropy production) are subdivided into two components, one accounting for exchanges between the system and its surroundings (deS: heat-generating transformations) and the other for production by processes internal to the system (diS: conservative transformations). Exchanges between organisms and their surroundings are accompanied by a great deal of waste dissipated into the surroundings; hence, deS is large compared with diS. Open systems, however, must produce entropy internally. Or, dS = deS + diS, diS > 0.
Internal production (diS), manifested as storage and transmission of information, is critically important in biological evolution, even though it represents a tiny portion of an organism's energy budget.
Biological systems maintain themselves in highly organized states far from thermodynamic equilibrium through causal engagement with the surroundings. That engagement must be mediated by a physical distinction between the "inside" and an "outside" of the organism, allowing for autonomous internal processes. This boundary is provided by cell membranes, which are not only physical barriers between the inside and outside of the organism but are also mechanisms for modulating the exchange of matter and energy between the organism and its surroundings.
Production Rules in biological systems govern the internal processes. Following ZOTIN & ZOTINA (1978), BROOKS & WILEY (1988) used the symbol Ψ to denote two classes of processes: 1) those involved in dissipation from the system (external dissipation: Ψa), mostly heat generated by production within the organism and lost to the surroundings, adding to the energy lost as a result of bringing matter and usable energy into the system from the surroundings, and 2) those involved in dissipation within the system (bound dissipation: Ψµ, all structure maintained within the organism). In biological systems, cm can be further subdivided into allocations for accumulating biomass (cµb) and allocations for accumulating information that can be passed on by inheritance (cµi). Brooks and WILEY (1988) suggested that diS could be viewed heuristically as diS = Ψaª + Ψµb + Ψµ1.
Heat-generating processes (deS plus Ψa), occur when energy and entropy flow in opposite directions, tending to move the system towards disordered states. Organisms mitigate these effects by "exporting" entropy to the surroundings; if all the heat generated by processes associated with bringing matter and energy into an organism stayed in the organism, it would rapidly die. Conservative transformations are characterized by energy and entropy flowing in the same direction, entropy production being retained within the system and tending to move the system towards more structured states. As entropy and energy flow through biological systems at different rates, structure accumulates at different levels of organization; furthermore, the structure at any given level is constrained by energy and entropy flows at other levels.
Organisms maintain themselves through time by exploiting "resource gradients" in the surroundings (ULANOWICZ 1997), determined by interactions between abiotic and biotic factors. Abiotic factors can be structured in part by ca; for example, metabolic processes are involved in the degradation of high-grade energy sources into lower grade forms of energy, including heat. Both the capture of incoming solar energy by biological systems, and the mass re-radiation of heat by these organisms affects the thermal profile of the earth. Additionally, the production of oxygen as a byproduct of photosynthesis and of carbon dioxide as a byproduct of aerobic metabolism affect the composition of the earth's atmosphere. Biotic factors are also subject to the influences of cµb + cµi. Metabolic production tends to move biological systems in the direction of minimizing energy gradients in the environment, to the extent permitted by the inherited capabilities of the organisms involved (ULANOWICZ 1997, BROOKS & MCLENNAN 2000). In other words, accumulated genetic information (cµb + cµi) constrains the patterns of energy flow (ca) within organisms and between organisms and their surroundings (including other organisms).
Biological systems produce entropy at different rates because energy stored by conservative transformations is degraded at different rates. At the lowest organizational levels, the shortest time intervals, and the smallest spatial scales, the greatest relative contribution to c is ca. For cellular or sub-cellular structures over short time intervals, physiological processes dominate explanations. Most entropy production is dissipated into metabolic heat loss. At intermediate levels of organization, space or time, the effects of cµb predominate. Most entropy production at this scale is dissipated into accumulation and maintenance of biomass. Finally, on the largest and longest scales, cµi predominates, and the patterns relevant to biological explanations are formed by accumulation and maintenance of genealogical diversity. From the perspective of the environment, such patterns of biodiversity tend to be organized with respect to energy gradients, whereas from the perspective of the genealogical system, biodiversity is organized with respect to sister-group relationships and patterns of geographical distribution that mirror geological evolution occurring on similar temporal and spatial scales.
We're all children of Time.
- Shevek, The Dispossessed (LeGuin, 1974: 385)
Systems showing irreversible behavior have a sense of time generated by the thermodynamic costs of their behavior. Biological systems make time in two ways, a dualism called the Shevek Effect (BROOKS 2010, 2011c). Exchanges of matter and energy between the system and surroundings (deS) generate cyclical time. Metabolic and other homeostatic mechanisms are examples. Shevek called this Simultaneity, because the endpoint of a complete cycle of irreversible processing of matter and energy is the starting point - beginning and end are the same. These cycles maintain living systems, allowing them to persist long enough to be able to change, but they do not create change by themselves. For such changes to occur, biological systems must also be able to make linear time, which Shevek called Sequency. Sequential time is made by thermodynamic production (diS); reproduction, inheritance, ontogeny, and speciation are examples.
Both simultaneity and sequency processes can be envisioned in terms of energy flows. More often, however, we express sequency processes in terms of the flow of information. The relationship between information and material phenomena, including thermodynamics, has had a long and turbulent history. Information theory has developed from two general perspectives, "communications theory" and "measurement theory". These perspectives overlap in proposing that information 1) is anything transmitted from a "source" through a "channel" to a "receiver" and 2) is an abstraction rather than a material part of the system. Neither of those conceptions is adequate for describing biological systems.
In communications theory, the amount of information sent from a source is calculated using a statistical entropy function. Errors in transmission can result from poor encoding at the source or from noise in the transmission channel. Meaningful information is that subset of information transmitted actually recorded by the receiver (there may or may not be a separate decoder). All of the processes affecting the transmission and reception of the information thus decrease the entropy of the message from its maximal value at the source. Physical entropies are expected to increase as a result of work done on the system, so either information transmission is not a physical process or the communications view of entropy is non-physical.
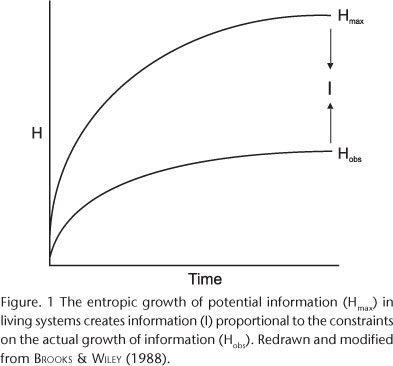
Measurement theory provides a second formalism. BRILLOUIN (1962) distinguished "free information,"a metaphor, and "bound information," referring to material properties of the system. Bound information is determined with respect to the "complexions" (microstates) of the system. Hence, it is also calculated using a statistical entropy function but, contrary to communications theory, is expected to exist only in systems for which there is a non-arbitrary microstate/macrostate distinction. Bound information is defined as: I = Hmax - Hobs; where Hmax refers to the totally relaxed state of the system (usually estimated by randomizing observed components of the system). Brillouin defined I as "negentropy", which is converted into bound information by measurement (measuring devices are receivers), so negentropy = information. Information points to, but is not a material part of, the system.
BROOKS & WILEY (1988) presumed there must be an additional conception of information because biological information is a communication system with a material basis. They used a general mathematical formalism summarizing changes in the number of parts, the number of kinds of parts, and the relative frequency of the different kinds of parts, key elements of biological complexity. This simple informational "entropy function" has four potentially illuminating formulations: 1) the "actual" entropy (Hobs) calculated on the basis of the observed distribution of components of the system (Information Content, or Expressed Information); 2) the "maximum possible" entropy (Hmax), estimated by calculating the entropy value for the components of the system at any given time if they were all randomized (Information Capacity); 3) an absolute difference (Hmax-Hobs) (Information or Macroscopic Information: GATLIN 1972, LANDSBERG 1984a,b); and 4) two conceptually related relative differences Hobs/Hmax and Id/Hmax (Id being Information Density) called Order (LANDSBERG 1984a,b) and Redundancy (GATLIN 1972), respectively. Simple heuristic simulations emulating biological processes associated with the storage and transmission of information (e.g., reproduction, ontogeny, and speciation) produced three generalities (for illustrations, see BROOKS & WILEY 1988): 1) Hobs is an increasing function of time; 2) Hobs is a concave function of time, as historical constraints retard the rate of entropy increases; and 3) the difference between Hmax and Hobs is an increasing function of time, permitting the growth of structure and organization (COLLIER & HOOKER 1999). Observation (3) points to an informational duality in the nature of the organism; entropy and information/organization/order/redundancy increase over time (Fig. 1). If Hmax is a function of the capacity, or potential, of a system and Hobs is a function of the expression of some of that potential, the difference between information capacity and information content is proportional to the constraints, inherent and extrinsic, on the system. For example, additive genetic variance could be construed as an indication of population-level entropy, while genetic correlations would be an indication of organizing principles constraining that variance.
COLLIER (1998, 2000) related information to the causal capacity of a system, its ability to impose distinctions on its surroundings. Collier proposed that physical (material) information systems occur as arrays, or multi-dimensional messages, in which the information has: 1) a physical basis, 2) an energetic cost, and 3) a real (non-arbitrary) macrostate/microstate distinction. Since the discovery of the chemical structure and function of DNA, there has been a material basis for biological information, satisfying (1) (COLLIER & HOOKER 1999, BROOKS et al. 1989, SMITH 1988, 1998, 2000). Requirement (2) is satisfied by showing that energy and information are interconvertible. Energy dissipated within the system as a result of work done on the system (heat-generating transformations, or ca of diS) is intropy (overhead of ULANOWICZ 1997), while energy converted into structure (conservative transformations, or cµ of diS) is enformation. Conservative processes within biological systems are coupled with heat-generating processes, so there is an energetic cost associated with the production and maintenance of biological information. Intropy and enformation are interconvertable (e.g., energy brought in from the surroundings can be converted into structure, e.g. glycogen, which can then be converted into heat).
Finally, macrostate/microstate distinctions are determined objectively by part/whole associations (COLLIER 1998, 2000). The number of accessible microstates is increased by the production of new components, either at a given level or through the opening up of new levels of organization. For example, auto-catalytic processes producing monomers make "monomer space" available. Some monomers have high chemical affinities for each other, and will spontaneously clump into dimers and polymers. Once polymers begin to form, "polymer space" becomes available to the evolving system. At this level, polymers are macrostates and monomer and dimer distributions are microstates. Causal interactions among polymers create new levels of organization in which polymer distributions are the microstates and new levels of organization are the macrostates, and so on. Each new functional level creates a hierarchy of increasing structural intricacy, manifested by increasing allocation of the entropy production in structure. Therefore, the allocation of diS to cµ might be nearly proportional to entropy increases due to the expansion of phase space resulting from the creation of new possible microstates. A protein coding unit could be a macrostate, while all the actual sequences that code for that protein would be its microstates; a locus could be a macrostate, and all alleles corresponding to that locus the microstates; phenotypes could be macrostates, and all genotypes corresponding to a given phenotype would be microstates (BROOKS & WILEY 1988).
Cohesive properties are also essential to understanding microstate/macrostate distinctions in biological systems. Ranging from molecular affinities to cell-cell adhesion to genetic compatibility, mate recognition, and genealogy, they provide resistance to fluctuations from lower levels, allowing macroscopic properties to emerge. Cohesion is thus analogous to inertia (we discuss cohesion below). This introduces an irreducible hierarchical structure to the evolution of living systems. The major transitions (MAYNARD SMITH & SZATHMÀRY 1995, 1999) are all associated with the emergence of new forms of cohesion, which permit information to be stored and transmitted more efficiently.
Interactions between Simultaneity and Sequency
We're just recycled history machines. - Jimmy Buffett
Theoretical studies in nonequilibrium thermodynamics (e.g., PRIGOGINE & WIAME 1946) and popular texts (e.g., SCHRÖDINGER 1945, BLUM 1968, PRIGOGINE 1980) in the aftermath of WWII laid the groundwork for a popular view about the thermodynamics of life. These physicists argued that life was so improbable it demanded a special explanation. They accepted the progressive nature of evolution, which they also considered contrary to the expectations of the Second Law. Prigogine and colleagues developed a model by which life could originate as an improbable event and evolve into increasing improbable states. Looking to flows between the system and its surroundings, deS, for insights, they discovered that near equilibrium, random fluctuations in the exchanges between the system and surroundings could theoretically produce states of lowered entropy. They surmised that if such fluctuations were, on rare occasions, "captured" in a stable state, they could move themselves farther and farther away from thermodynamic equilibrium, "feeding on negentropy" (SCHRÖDINGER 1945).
This view became so popular that when BRODA (1983) discussed Boltzmann's 1905 lecture, he changed "entropy" to "entropy [negentropy]" throughout the text. And yet, Boltzmann stated that life was a struggle for entropy. It seems Boltzmann was able to see the organism's perspective, focusing on the source rather than the fate of the matter and energy needed to sustain life. Living systems must find usable energy. Plants find this in the form of photons coming from the sun. The source of those photons is thermonuclear reactions in the sun. Being relatively low energy products of the sun's thermonuclear reactions "exported from the system to the surroundings," photons are part of the sun's entropy production. Photonic energy used by plants to build biomass is part of plant entropy production, so when herbivores eat, they are feeding on entropy.
In exchanging matter and energy with their surroundings, organisms degrade their surroundings more than themselves, maintaining a low-entropy state relative to their surroundings. This observation formed the basis for the principle of maximum entropy production (SWENSON 1989) and a view of self-organization (DEPEW & WEBER 1995). This may look like the Darwinian duality, but "self-organization" in this context means the tendency for the system to organize itself according to the nature of the conditions. The principle of maximum entropy production asserts that systems will utilize resources from the surroundings as rapidly as possible; those sequestering the maximum amount of energy fastest win, starving out their slower competitors. Both concepts ascribe causality (necessity) to the conditions rather than the organism (BROOKS 2011a,b). Absent from these formulations is thermodynamic production, diS (the nature of the organism).
It is more complete to recognize that biological systems maintain low-entropy states relative to their surroundings, but not relative to their own previous states. Entropy can, and does, increase through time - the inevitable structural decay that accompanies such transactions can only be delayed (LOTKA 1913, 1925). But the entropy increase that characterizes biological systems is far from maximum entropy production. ZOTIN & ZOTINA (1978) studied metabolic rate change during ontogeny. Early in ontogeny, organisms exhibit high metabolic rates, corresponding to something like maximum entropy production. Through time, however, this "immature" stage is always replaced by a "mature" or "steady state" phase, characterized by reduced metabolic rate. Finally, all organisms enter a "senescent" stage in which metabolic rate decreases further. This same dynamic occurs during ecosystem succession (ULANOWICZ 1997). Decreasing rates of entropy production are determined by interactions between the surroundings and the "sense of self" the organism inherits from its parent(s). The material information system of inheritance is thus critical in determining the ways in which the organism interacts with the surroundings to produce its actual lifespan. Evolutionary persistence is associated with decreasing rates of entropy production, not maximal entropy production.
BLUM (1968) introduced the metaphor of time as an arrow, being propelled into the future by power applied from "outside." But irreversibility requires only temporal asymmetry among the moving parts of an ensemble of components. Temporal asymmetry may occur as a result of some elements moving more slowly than others, interacting more often with fewer elements of the overall ensemble. This produces non-random, or historical correlations that slow parts of the system, intensifying temporal asymmetries (in cosmological models, gravity performs this function). Accumulated historical correlations produce irreversible system behavior regardless of the quantity or quality of novelty. Or, as Maynard SMITH & SZATHMÀRY (1995) stated, the conjunction of two or more events, each of which is improbable, is sufficient to make any evolutionary transition irreversible. The reduction in entropy production rate over time in biological systems is a direct manifestation of irreversibility resulting from the net accumulation of historical correlations. For neo-Darwinism, irreversible phenomena result from selection driving the system toward a better future. If at some point the next step to a better future requires retracing the past, selection ratchets that as well. Only time is inherently irreversible, and that is treated as free and epiphenomenal. But there is no free lunch in the universe- even time must be paid for. Our viewpoint accords with empirical phenomena, and also allows evolutionary biologists to accept irreversibility without embracing Spencerian progressivism. There is no distinction between ratchet irreversibility and temporal irreversibility - ratchet irreversibility is the establishment of temporal correlations strong enough to limit the options the system can explore.
A new biological framework must incorporate a naturalistic explanation for the origin of life, but cannot be solely a theory of the origin of life. The mechanisms that allowed life to originate must also explain its subsequent evolution, including the origin of the inherent constraints that allowed natural selection to emerge as a natural process. The first goal of biological theory, therefore, is explaining the origin of evolvable life (MAYNARD SMITH & SZATHMÀRY 1995, 1999, SZATHMÀRY 2006). The solution lies in recognizing that organisms are metabolic and information systems. The major transitions involve trade-offs between the need to exist, a metabolic problem, and the need to reproduce, an informational problem. Simultaneity is how we pay for evolution, it is not evolution itself. This is the reason metabolic-cycles-only theories of the origin of life fail (MAYNARD SMITH & SZATHMÀRY 1995). Evolvable life also requires sequency. Simultaneity phenomena in biology are much more costly than sequency phenomena, so it is far cheaper to evolve than to remain static. The transitions from non-life to proto-life to life involved division of labor into metabolic and informational functions, allowing more efficiency for each, but at the cost of a loss of independence through greater integration. Each of these transitions was irreversible, adding the sequency essential for evolutionary theory at little additional cost.
Temporal production opens the system and allows persistence and change. A useful visual metaphor for the interaction of simultaneity and sequency could be the symbol of the Great Circle of Life, called "helix" in Greek mythology (Fig. 2).
SPACE
The distinction between the nature of the organism and the nature of the conditions creates a metaphorical spatial duality. The ways in which organisms interact with themselves, the Genealogical Hierarchy, creates Information Space. The ways in which organisms interact with their surroundings, the Ecological Hierarchy, creates Function Space, a subset of which is Fitness Space.
Information Space
"The bond is simply inheritance, that cause which alone, so far as we positively know, produces organisms quite like each other..." - Darwin, 1872: 348
Information space encompasses all possible material inheritance systems (JABLONKA & LAMB 1995). The nature of the organism is to explore possible inheritances through reproduction and to expand information space through innovation. The realm of possible inheritances is Potential Information. The realm of actual inheritances is Realized Information.
Organisms are digital replicators, giving them virtually unlimited capacity for variation (MAYNARD SMITH & SZATHMÀRY 1995, 1999, SZATHMÀRY 2000). Consider a replicator comprising a string of DNA 1,000 bases long. If the string were a holistic replicator, it would function strictly as a single unit. In that case, the sequence contains a maximum of 2,000 bits of potential information. This has been used to suggest that there is not enough information in the DNA of a cell to specify an entire organism, and therefore information from the surroundings must be incorporated during development. This is consistent with the view that organisms organize themselves with respect to the surroundings - the organism's information system is cheaply produced because its only function is allowing the system to conform to the much larger amount of information in the surroundings.
If the string functions as a digital replicator, however, it could be read at multiple levels, from single bases to the entire sequence. Each of those readings would have a maximum information capacity of 2,000 bits. If each reading of a given sequence is equivalent to all others, and if these readings are not interactive, the total possible readings would have a maximum information capacity of 2,002,000 bits. If the bases are interactive, then these self-interactions will constrain the total information capacity. The total amount of information that could be expressed at any one point in time is also constrained by the fact that bases (a similar argument holds for genes, tissues, and organisms) are causally linked, so accessing some information will limit expression of other, producing an upper bound on the amount of information potential/capacity that could be expressed at any one time. Accessing the same system in different ways through time permits the same information to be additive, since at each point in time it is a different 2,000 bits being expressed. Despite these inherent constraints, there is abundant potential information - in fact, much of that potential must be dissipated in order to distill out an organized organism. Ontogeny is that distillation process - it's an energy-efficient algorithm for converting digital information into analog output using matter and energy from the surroundings to accomplish the task, paid for by the dissipation of potential information.
FRAUTSCHI (1982, 1988) and LANDSBERG (1984a,b) contrasted two classes of processes that generate entropy. The first is equilibration of temperatures between system and surroundings. Biological systems exhibit this kind of entropic behavior through simultaneity processes that result in the dissipation of energy (heat loss) to their surroundings. The second is expansion of the realm of possibilities (the phase space) in which the system resides, increasing its number of accessible microstates (possible configurations). System organization increases so long as equilibration (equiprobable distribution of the system over its microstates) occurs at a slower rate than the expansion of the phase space, allowing a lag between the increase in realized entropy or expressed information (Hobs) and the increase in the maximum possible entropy or potential information (Hmax) (Fig. 1), which is a linear function of the logarithm of the number of states or size of the phase space. So long as the phase space expands faster than the system can fill it up, increasing entropy can be accompanied by the emergence of organized structure. In cosmology, this argument explains the spontaneous and irreversible emergence of stars, solar systems, galaxies, and other organized structures, in which fundamental forces linking material bodies, like gravity, slow down the entropic diffusion of matter in the universe to such an extent that organized structures emerge as a result of increasing entropy. A similar phenomenon accompanies the dissipation of energy from the sun into space, where some of it is captured by photosynthetic life. Sequency processes produce these effects in living systems.
Mutations and higher order genotypic and epigenetic innovations expand the informational phase space, while inheritance systems (JABLONKA & LAMB 1995, MAYNARD SMITH & SZATHMÀRY 1995, 1999, SZATHMÀRY 2000) play roles analogous to fundamental forces like gravity (BROOKS & WILEY 1988). Evolutionary transitions are produced by innovations that increase the efficiency of storing and transmitting information. The occupied information space increases through time but the total amount of information space grows even more (Fig. 1); realized evolutionary innovations always produce new potential innovations.
Function Space
Organisms impose themselves on their surroundings, creating the space in which biological functions occur. Within this space, each organism creates a "fitness valley" analogous to gravity wells created by bodies in space/time. The portion of function space in which organisms could have non-zero fitness is Fundamental Fitness Space. The portion of fundamental fitness space accessed is Realized Fitness Space.
The same Evolutionary Lagload (genealogical conservatism: Maynard Smith, 1976) that produces the difference between realized and potential information prevents the genealogical system from being distributed at maximum density in all places at all times, so there will always be unoccupied fitness space. Realized fitness space may grow, but lagload keeps it a subset of fundamental fitness space. Evolutionary innovations may increase both fundamental and realized fitness space over time. When an organism dies, its fitness valley disappears. If one organism is replaced by another with the same requirements (virtually guaranteed by conservative reproduction), the fitness valley may appear to persist but has no independent existence. The apparent persistence results from historical conservatism of the nature of the organism-to the extent that niches are real, they are products of the nature of the organism, not the nature of the conditions. Organisms thus (re)constitute niches; they do not construct them, nor are the surroundings inherently organized into niches.
Interactions Between Information and Function Space
The origin of evolvable life created the spatial dualism. Each aspect of the dualism grows on its own and both interact in ways that mutually constrain, but do not reduce, each other's growth. The result is evolvable space. Standard neo-Darwinian accounts describe well the microscopic dynamics within evolvable space, but leave two other issues unresolved. The first of these is macroscopic dynamics and the second is the emergence of meaning in life (not to be confused with the meaning of life), which relates directly to issues of cooperation in evolution, and which we discuss more fully below.
MACROEVOLUTIONARY DYNAMICS: SLOPPY FITNESS AND SLOSHING BUCKETS
Darwin's dualistic view of the nature of the organism is apparent in his using adaptation to mean both "function" (nomenclature) and "evolutionary response to environmental change" (metaphor). Organisms need functions sufficient for survival and reproduction in the environment of the moment. During periods of relative environmental stasis, populations accumulate variations on those adaptations conferring non-zero fitness. Organisms whose adaptations are particularly well suited to those local conditions dominate numerically at any given time. Equally critical for Darwin was that all variants with non-zero fitness survive to some degree; if a single best-adapted variant replaced all others in a species, that species would go extinct with the next environmental change. A subset of "adaptations as functions" became "adaptations as evolutionary process." Darwin made this a cornerstone of his theory because organisms were often found in situations in which the same adaptation had non-zero fitness in different environments (e.g., deciduousness in trees originated as an adaptation associated with drought but also functions well as an adaptation associated with cold winters: WANNTORP 1983), and that organisms found in the same surroundings often exhibit many different adaptations for dealing with those surroundings
"...for nearly similar variations sometimes arise under, as far as we can judge, dissimilar conditions; and, on the other hand, dissimilar variations arise under conditions which appear to be nearly uniform." - Darwin, 1872: 32
During periods of environmental stasis, Darwinism is "survival of the adequate with the fittest dominating numerically," while during periods of environmental change, Darwinism is "survival of the adequate with the fittest going extinct, replaced by a variant that had lower fitness in the previous environment." This is the Gambler's Ruin: no matter how fit you are at any given place and time, you can still go extinct if conditions change. Just ask the non-avian dinosaurs.
The smaller the proportion of fitness space occupied, the "sloppier" is the actual "fit" (AGOSTA & KLEMENS 2008). This "sloppiness" should increase over evolutionary time as the total information/fitness space grows, increasing the diffe-rence between what is possible and what is realized at any given time. Sloppy fitness space allows room for creativity and innovation because a lot of non-zero fitness space is always potentially available, however, this does not rule out the possibility of local tightly optimized adaptations. Furthermore, sloppy fitness space allows the genealogical hierarchy to operate with a high degree of autonomy from the ecological hierarchy without sacrificing adaptability. The ability to move from surroundings that are deteriorating with respect to fitness is more important in determining survival than how well adapted the system is to any particular part of fitness space. That adaptability (or resilience) is a function of how many historical alternatives you maintain in your collective genome (only successful adaptations get carried forward, so retained history is history of past success), and of how many ways - old and new - that information can be used to respond to changing conditions.
The adaptability and resiliency afforded by sloppy fitness space provides many opportunities for Ecological Fitting between organisms and their environment (AGOSTA & KLEMENS 2008, AGOSTA et al. 2010). The term "ecological fitting" was coined by Janzen (1985) to describe the situation in which organisms use traits (information) they already possess to form novel trophic interactions with other species or, more generally, to expand into new habitats, use new resources or respond to rapidly changing conditions. The essential point (Janzen 1980, 1985) is that this "fitting" can structure ecological communities (i.e., the interactions within and between the genealogical hierarchy and the surroundings at any given point in time and space) in ways that appear intimately "(co)evolved" to the naïve biologist, when in fact most species and their traits evolved elsewhere and were co-opted (MAYNARD SMITH & SZATHMÀRY 1995, MCLENNAN 2008) to function in the novel condition. Sloppy fitness space, as a result of evolutionary lagload (genealogical conservatism) and the overall developmental and phenotypic plasticity of genomes (WEST-EBERHARD 2003), is what underlies the opportunities for ecological fitting (AGOSTA & KLEMENS 2008). The sloppier the fitness space, the less specialization and adaptation are inherent evolutionary "dead-ends" (AGOSTA et al. 2010), and the more potential for organisms to do things their ancestors did and, more fortuitously, to do truly innovative things in response to changing conditions. Because what is realized is always a subset of what is possible (Hmax > Hobs), even highly "specialist" organisms like many parasites and plant-feeding insects can maintain the ability (information) to become "generalists", allowing them to shift rapidly to novel resources in ecological time without the time delays associated with the evolution of novel capabilities (BROOKS & MCLENNAN 2002, AGOSTA 2006, BROOKS et al. 2006a,b, AGOSTA & KLEMENS 2008, 2009, AGOSTA et al. 2010).
AGOSTA & KLEMENS (2008) visualized ecological fitting as a result of the interaction between sloppy fitness space (nature of the organism) and the environment (nature of the conditions). Consider a bivariate world whose dimensions define all possible combinations of two resources, X and Y (Fig. 3). At a given point in space or time, the local or realized conditions are defined by the subset of X and Y that exist. For an evolutionary unit (e.g., genome, species, macroevolutionary lineage), its fundamental fitness space is all combinations of X and Y for which positive fitness can be achieved. Realized fitness space is that portion of fundamental fitness space accessed to meet the realized conditions. The extent to which fitness space is "adapted" to any particular set of realized conditions will be a function of time and selection pressures, but evolutionary lagload ensures the fitness space will always remain in large part a function of retained history (genealogical conservatism). The mismatch between fundamental fitness space and realized fitness space at any given point in time or space is proportional to how "sloppy" the fitness space is. From large-scale environmental perturbations invoked in the Sloshing Bucket model (see below) to small-scale perturbations like local extinction of a host species, it is this "sloppiness" that provides the essential adaptability and resiliency of life to respond temporally to rapidly changing conditions (Fig. 3a) and to disperse and explore new options across the physical landscape (Fig. 3b). Following disturbance or dispersal and depending on patterns of reproductive isolation, wandering through sloppy fitness space both facilitates evolutionary diversification by exposing organisms to novel conditions (Fig. 3a) and promotes evolutionary stasis if there are many ecologically fit populations experiencing contradictory selection pressures and significant gene flow (Fig. 3b).
While evolutionary lagload provides organisms with sloppy fitness (adaptability and resiliency), it also places severe constraints on the ways in which and the extent to which evolvin-g lineages can respond to conditions in their present environments, and thus their ability to produce "optimal" offspring to "match" the current condition. This implies that there will be parts of fitness space where reproductive overrun creates conflicts from which there is no escape. These will be the foci of intense selection and even extinction. The major transitions are associated with innovations in the genealogical system that increase the efficiency of storing and transmitting information (MAYNARD SMITH & SZATHMÀRY 1995). Large-scale and novel innovations in the genealogical system should produce large-scale and novel interactions between information space and fitness space. This, in turn, should result in novel forms of selection. BROOKS & WILEY (1988) used the term "irreversible" synonymously with "transition" and proposed that all evolutionary innovations were characterized by the coupled evolution of novel forms of cohesion and selection. And ELDREDGE'S (1995) view emphasizes a hierarchy of selection processes emerging from the (co)evolution of the genealogical and ecological hierarchies.
Evolutionary lagload raises the possibility that there can be environmental changes of such a great magnitude or rapid rate that species information systems cannot cope, and become extinct. ELDREDGE (2003) used the metaphor of a sloshing bucket for discussions of the macroevolutionary dynamics of environment-driven extinction and subsequent evolutionary renewal. In cases of great environmental perturbation (the bucket is sloshed so much that some of the contents are lost), portions of information space are eliminated, not just restricted in fitness space. As well, the nature of the conditions may also change. Both phenomena create new opportunities for the species that escape extinction.
Environmentally caused mass extinctions catalyze new speciation events. Extinctions in one area open up geographical territory for colonization by survivors, which initiates various forms of allopatric speciation. This alone, however, only increases the number of species-it does not necessarily increase evolutionary diversity. Fitness space becomes sloppier as a result of the reduction of occupied information space, meaning that the range of non-zero fitness options for the survivors changes and grows. And that means increased opportunities for co-option of preexisting adaptations (MCLENNAN 2008). Competitive Release occurs when one species that had been precluding another from part of its potential fitness space goes extinct. If the environment changes in such a way that a species may occupy more of its potential fitness space, it might appear that a new adaptation had evolved
Bucket-sloshing events also help establish the conditions under which novel adaptations may arise, even if they do not initiate them. But most diversification is due to the rapid deployment of previously realized and potential information in previously inaccessible parts of fitness space, known as taxon pulses, turnover pulses, and taxon cycles. This is the source of the punctuated aspect of the recovery following mass extinctions. As well, the more rapid the deployment of old and new information in fitness space, the more rapid the accumulation of historical correlations, slowing the process of diversification. This pattern has been interpreted by neo-Darwinians as an indication that the surroundings are constructed in such away that only a certain number of species can be accommodated and as that number of species is approached, the rate of diversification slows. But the nature of species with respect to how their member organisms interact with their surroundings is an emergent property of the genealogical system, not the surroundings. Every new species that evolves represents potential new habitat for other species to come. This is the way evolvable life builds the 'window of vitality' (ULANOWICZ 1997) we call the biosphere.
MEANING IN LIFE
Ultimate meaning in biology is self-meaning; thus, whenever there is a conflict between the organism and the environment, the organism takes precedence, with a resulting loss of fitness. Ultimate explanation in biology thus is inheritance, lightly to heavily filtered by selection.
Non-living systems exist, living systems behave. Organisms are complex adaptive systems operating through self-referential features, such as feedback loops and other homeostatic mechanisms, which provide information (signals) about their internal state. Organisms do not signal themselves intentionally; the signaling process emerges from the kind of physical systems they are. Signaling mechanisms are of two types, (1) those in which the organism signals itself about itself and (2) those in which the organism signals itself about its surroundings (BROOKS & MCLENNAN 1997).
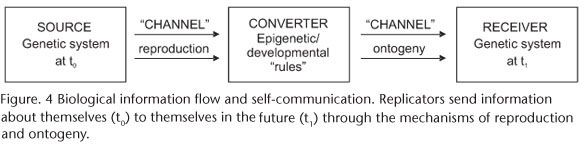
Many information theorists define information to be meaningful only if a receiver has converted it. A standard neo-Darwinian perspective (GATLIN 1972) construes genealogical replicators as the source of biological information (chance) and the environment in which they live as the receiver (necessity), the nature of the conditions determining what is meaningful. Our perspective is that the nature of the organism determines what is meaningful. Replicators are both source and receiver (Fig. 4), transmitting information from themselves at (t0) to themselves in the future (tn) (CSANYI 1989, KAMPIS 1991, 1998, BROOKS & MCLENNAN 1997, COLLIER & HOOKER 1999). This perspective has parallels in self-correcting computer programs and self-correcting capacities of DNA. Information is meaningful because it is organized in such a way that an organism develops and the information system is transmitted to the next generation.
Organisms possess many self-signaling mechanisms. These enhance the functional integration of organisms, a key to their persistence as coherent entities through time, as well as their vital connections with their surroundings. Cell, molecular, and developmental biologists routinely describe molecules, cells, and tissues as signaling each other during reproduction and ontogeny. Some internal signaling systems produce effects in the organism that are apparent to other organisms in the environment. Such signals are by-products of internal-state signaling not produced intentionally as signals sent to specific organisms. They exist in the absence of any external receiver because they the signaler is the receiver. Predictable responses by other organisms may be an emergent property of these effects. Just as the effects of internal signaling in the sender may have external manifestations, the ability to receive a particular signal, and the meaning imputed to it, are properties of the receiving organism's internal signaling system, and do not originate intentionally. Organisms cannot evolve mechanisms for receiving a particular signal from their surroundings prior to the origin of that signal.
There are three classes of possible outcomes when internal signaling is perceived by other organisms. First, the effect may evoke no response by the observer. In practice it may be difficult to distinguish a case in which the organism perceives a signal and rejects the information (does not respond) from a case in which the organism does not perceive the signal (GERHARDT et al. 1994, MCLENNAN & RYAN 1999). An antelope will evoke a hunting response in hungry lions, little response from lions that have just fed, and no response from a daisy. Second, the effect may produce aggregating responses by the receiver that are positive to both sender and receiver (e.g., they represent potential mates), or positive to one but not the other (one may represent food to the other). And third, the effect may produce avoidance responses by the receiver.
The sender is unintentionally broadcasting and observers in the environment are not intentionally receiving a particular signal. Regardless of intention, perceiving the signals produces an additional level of selection, one in which receivers become agents of selection. The fate of the organism's unintentional signal is determined by a complex balance of costs and benefits associated with the three response categories listed above. So long as total benefits outweigh the total costs to the sender, the signal will persist through continued reproduction, having a net cohesive influence on its genealogical system. Organisms are simultaneously in the environment and part of the environment (MAYNARD SMITH 1976), so it is only a small step to move from the emergence of mutual signaling systems - produced unintentionally - to a transitional dynamic in which another level of cost-benefit considerations emerges, one involving enhanced cohesion and functional integration accompanied by more constraints through mutual inter-dependence.
Organisms are both signals and signal-bearers, because some of their attributes may be perceived by other organisms in their surroundings. If external signals are by-products of internal signaling, then the effects of such 'internal conversations' may have more than one meaning to observers in the environment. The evolutionary fate of signal bearers will be an outcome of the ways in which other organisms perceive the signal bearer. In addition, if the signals that selection can act upon are only a subset of all the internal signaling, which selection can affect only indirectly, we must assume that various selection pressures, operating on different levels of biological organization, may themselves conflict to some extent. Consider an organism that changes color when it eats a particular type of high-energy food. The change in color is an unintentional by-product of an internal signal of well being (more energy coming into the organism's metabolic system) that may also make the organism highly visible to predators. Selection will favor enhanced efficiency in obtaining high-energy food and minimize external manifestations of that success. This is the source of Darwin's view that most evolutionary change will be incremental - the process is not inherently gradualistic, but there are so many conflicting selection pressures on organisms in genealogical associations (populations, species) that the net outcome is most likely to be gradual. As much as ideas of episodic, or punctuated, evolutionary change may have been seen as contrary to neo-Darwinian thought, they are in no way outside the framework of Darwinism.
Organisms appear to know where they are going in the intersection of information and function space because inheritance rarely strays far from where it has been. Organisms appear to know where they are going with respect to interactions with their surroundings because intention and meaning are decoupled and meaning is both contextual and contingent. That is, by de-coupling intention and meaning, equating them with the nature of the organism and the nature of the conditions, respectively, BROOKS & MCLENNAN (1997) provided a material basis for selection that exorcises the specter of teleology from evolutionary explanations.
COMPLEXITY
The dualities of space and time interact with each other internally, but space and time also interact on a higher level. This produces the complex system of evolving life that embodies yet another metaphorical duality.
Cohesion
"...homologous parts tend to cohere." - Darwin, 1872: 158
MAYNARD SMITH & SZATHMÀRY (1995) suggested that the conjunction of two or more events, each of which is improbable, is sufficient to make evolutionary transitions irreversible. History, in the form of conservative inheritance, maintains improbable events long enough to increase the chances that two or more of them will become causally intertwined. A good example is self-reflexive nucleotide copying. The more faithful the replication, the higher the degree of historical correlation, the more stable the product and the slower the rate of evolutionary change, but the greater the chance for accumulated rare events to meet.
The cohesive nature of historical correlations also makes evolution affordable. History lowers the cost of innovation, because innovations are modifications of pre-existing information. History lowers the cost of ecological specialization, because specialists on widespread resources have many options (BROOKS & MCLENNAN 2002, AGOSTA et al. 2010). It also lowers the cost of "adaptability," a synonym for retained history of what worked in the past. Finally, it lowers the cost of community organization if colonizers bring traits that allow them to coexist with residents with little or no competition (BROOKS & MCLENNAN 2002).
In addition to historical cohesion, all major transitions are marked by the origin of particular traits that enhanced informational cohesion (BROOKS & WILEY 1988) and the efficiency of storing and transmitting information specifying production rules (MAYNARD SMITH & SZATHMÀRY 1995).
Finally, organisms and their environments have significant causal relationships - as we have noted, selection processes are the cost of autonomous production of information. The nature of the organism and the nature of the conditions thus always produce complementary types of cohesion and selection. The transition from proto-life to life, the emergence of organisms replicating according to rules largely insensitive to the environment, created environmental selection. Likewise, the origin of sex produced complex species, which are more cohesive informationally than asexual lineages (BROOKS & WILEY 1988), but also produced sexual selection. Selection makes organisms environmentally cohesive. Organisms impose themselves on their surroundings, producing environmental selection proportional to the mismatch between the nature of the organism and the nature of the conditions. This causal interaction produces mutual information resulting from the exchanges between organisms and their environments, constrained by the inherited capabilities of the organisms involved (COLLIER 1998, 2000). Mutual information thus determines the sloppiness of fitness space. This leads to a slightly modified version of the Red Queen Hypothesis: the more evolution there is, the more space there is for evolution to occur; more than enough, for example, for real creativity and innovation to emerge, so we need not turn selection into a creator and innovator. That capacity is already part of the nature of the organism.
Compensatory Changes
The evolutionary interaction of information space and fitness space produces prodigious amounts of evolutionary potential as both "spaces" expand over time. Space and time, each with their own internal duality, interact in ways that limit each other yet allow realized biological diversity to grow. At the same time, the relative autonomy of the nature of the organism means that aspects of space and time associated with the nature of the organism will expand faster than aspects of space and time associated with the nature of the conditions. This inevitably produces density-dependent effects that result in compensatory changes (BROOKS & WILEY 1988).
A well-known example of compensatory changes is deformation of tubular structures into hexagons when closely packed. This occurs widely in living systems, from microfiber bundles in muscle tissues to the cells in beehives. But compensatory changes can also be more complex. Spea bombifrons is a species of spadefoot toad. Fossorial creatures, males and females emerge from underground burrows to mate and lay eggs in temporary pools created by massive rainstorms. The larvae are herbivorous initially. If the temporary pool evaporates too quickly, increasing larval density, they will forsake their vegetarian lifestyle for aggressive cannibalism. With each sibling consumed, the successful cannibals ingest a thyroid gland, which increases the concentration of thyroxin in its body. The effect of the increased thyroxin "density" is accelerated rates of metamorphosis that allow the larvae to escape the trap of an evaporating pool.
Interaction between Cohesion and Compensatory Changes
We believe the evolutionary transitions perspective best shows how cohesion and compensatory changes interact to produce the panoply of evolutionary dynamics. MAYNARD SMITH & SZATHMÀRY'S (1995) framework relies heavily on cooperation and division of labor. We believe that cooperation and division of labor are equivalent to cohesion and compensatory change, respectively. All transitions involve the evolution of new forms of cohesion that enhance self-stability through conflict resolution by means of compensatory changes that MAYNARD SMITH AND SZATHMÀRY called division of labor. Each transition augments the autonomy of the nature of the organism, which produces, as a compensatory change, a new form of selection complementing the new level of organization produced by the new cohesive property.
Subsequent modifications of the original formulation by Maynard Smith and Szathmàry enhance the inclusive nature of the transitional theory and resolve major conundra created by neo-Darwinism.
QUELLER (1997) observed that Maynard Smith and Szathmàry's major transitions were of two classes, fraternal and egalitarian, based on the source of the building blocks used to produce evolutionary innovations. Fraternal transitions arise when the building blocks have a common origin. A good example would be cell differentiation in the ontogeny of multi-cellular organisms. Egalitarian transitions, by contrast, involve elements having independent origins. Good examples of egalitarian transitions are the evolution of eukaryotic cells containing mitochondria and chloroplasts. Egalitarian transitions are examples of group selection, so Queller's perspective tells us that Maynard Smith and Szathmàry's framework, with its emphasis on cooperation, eliminates the conflict about group selection created by neo-Darwinism. Rather than argue about group versus kin selection in normative (nomenclatural) terms, we can now acknowledge that when the costs of cooperation are lower than the benefits, cooperation will be favored, regardless of the genealogical origins of the cooperating elements. As well, if intention is not meaning (BROOKS & MCLENNAN 1997), the concept of maximal selfishness, initiated by Spencer and refined by DAWKINS (1976), is exposed as something that is overcome in evolution rather than something that drives the process. This shifts our focus to questions of how cooperation occurs and what are its outcomes.
MAYNARD SMITH & SZATHMÀRY (1995, 1999) distinguished two classes based on limitations overcome in accomplishing transitions. Selection-limited transitions are those for which it is relatively easy to access a part of information space, but evolutionary success requires special conditions. Variation (information)-limited transitions are those for which it is difficult to access a particular part of information space but does not require special conditions for it to become entrenched evolutionarily. The latter are ruled out normatively by neo-Darwinism as examples of absolute fitness. But variation-limited transitions are not immune from selection, they simply do not require special conditions initially. And if selection is only called upon to select, not to create, this renders moot the conflict about relative and absolute fitness within neo-Darwinism.
Finally, BROOKS & MCLENNAN (2002) proposed three classes of transitions based on their macroevolutionary effects. These classes of transitions have much in common with Szathmàry's distinctions. Brooks and McLennan took the position that all sequency (irreversible) events are evolutionary transitions. The most common are those that are not difficult to achieve and do not require special conditions. Brooks and McLennan referred to these as "taking advantage of the rules of the game". They occur often but exert historical influences only on microevolutionary scales.
Transitions that Brooks and McLennan referred to as "changing the rules of the game" are either difficult to achieve but require no special conditions (variation-limited transitions), or are easy to achieve but require special conditions (selection-limited transitions). These transitions occur infrequently but consistently throughout the evolution of life and exert historical effects on macroevolutionary time scales. The evolution of herbivory in metazoans is a good example of a variation-limited rule transition. Three obstacles must be overcome in order to produce a true metazoan herbivore. First, a cellulose-digesting microbe that could live in the intestine of the metazoan must be found in the accessible environment of the metazoan. Second, the metazoan must provide the microbe a suitable living place in the digestive tract. And third, the metazoan must have a mechanism for transmitting the microbes to subsequent generations. Only if all three of these elements are combined can a metazoan become a true herbivore. This explains why herbivory has originated only a few times. At the same time, whenever herbivory evolves, it seems to have been selectively favored in any environment where plants live, explaining the great success of the few groups in which herbivory has evolved. An example of a variation-limited transition might be the repeated evolution of mammals with white fur, apparently restricted to boreal habitats where their color is cryptic. All speciation events are examples of this class of transitions, and may be either variation- or selection-limited (BROOKS & MCLENNAN 2002).
Finally, there are the transitions that Brooks and McLennan referred to as "changing the game": they are both difficult to achieve and require special conditions. These transitions are extremely rare but exert great historical influence on the subsequent evolution of biodiversity and the biosphere. They include the major transitions of MAYNARD SMITH & SZATHMÀRY (1995).
Brooks and McLennan's distinctions resolve two additional problematic elements of neo-Darwinism. They restore Darwinian historicity by placing evolutionary transitions in a narrative flow, ordering the sequence of origins of the various elements of each, arguably the most important contribution of the phylogenetics revolution. In addition, they provide a means of assessing the relative influence of each transition without placing a value on them.
These three enhancements of the transitional view of evolution define the dimensions of the intersection of information space and function space, i.e. evolvable space. Evolva-ble space comprises the products of evolutionary transitions that may be selection-limited or variation-limited and which may have short- to long-term temporal persistence (Fig. 5).
CONCLUSIONS
We hope this exercise will be helpful in framing discussions about the new framework in evolutionary theory, whatever we call it. A unifying and inclusive theory of biology must embrace the complexity of Darwin's Tree of Life and Tangled Bank. It must take us easily from the origin of evolvable life to today's diversity, from biology to chemistry and physics. And back again. It must account for the origins of biological phenomena, rather than just their subsequent maintenance.
That framework now largely exists. Increasing the autonomy of information flow enhances self-stability, which creates various forms of selection, which enhance mutual stability between life and its surroundings. The more evolution occurs, the more evolution is possible. The Meta-Game of Life is, "Persist as long as possible by integrating information flow and functional engagement with the surroundings." The strategy for accomplishing the meta-game is, "Increasing the efficiency of the information flow (the nature of the organism) enhances self-stability, which creates various forms of selection, which enhance mutual stability between the system and its surroundings (the nature of the conditions)."
Even more simply, paraphrasing MAYNARD SMITH & SZATHMÀRY (1995), "Because life originated as evolvable life, so long as the information flows, adaptation will take care of itself."
ACKNOWLEDGMENTS
We express our thanks to the editors of Zoologia for inviting us to contribute to this celebration. We also express special thanks to Misthu Banrjee, John Collier, Niles Eldredge, Alicia Juarrero, Cy Finnegan, Menachem Fisch, Jack Maze, Deborah McLennan, Jonathan Smith, and Eors Szathmary for their cooperative sharing of insights and criticisms that helped make this contribution possible.
LITERATURE CITED
AGOSTA, S.J. 2006. On ecological fitting, plant-insect associations, herbivore host shifts, and host plant selection. Oikos 114: 556-565.
AGOSTA, S.J. & J.A. KLEMENS. 2008. Ecological fitting by pheno-typically flexible genotypes: implications for species associa-tions, community assembly and evolution. Ecology Letters 11: 1123-1134.
AGOSTA, S.J. & J.A. KLEMENS. 2009. Resource specialization in a phytophagous insect: no evidence for genetically based performance tradeoffs across hosts in the field or laboratory. Journal of Evolutionary Biology 22: 907-912.
AGOSTA, S.J.; N. JANZ & D.R. BROOKS. 2010. How specialists can be generalists: resolving the "parasite paradox" and implications for emerging infectious disease. Zoologia 27: 151-162.
BLUM, H.F. 1968. Time's arrow and evolution. Princeton, Princeton University Press, 3rd ed.
BOWLER, P.J. 1983. The eclipse of Darwinism. Baltimore, John Hopkins University Press.
BRILLOUIN, L. 1962. Science and information theory. New York, Academic Press, 2nd ed.
BRODA, E. 1983. DARWIN AND BOLTZMANN, p. 61-70. In: E. GEISSLER & W. SCHELER (Eds). Darwin today: the 8thKühlungsborn colloquium on philosophical and ethical problems of biosciences. Abhandlungen der Akademien der Wissens-chaf-ten der DDR. Berlin, Akademie.
BROOKS, D.R. 2010. The mastodon in the room: how Darwinian is neo-Darwinism? Studies in History and Philosophy of Biological and Biomedical Sciences 42: 82-88.
BROOKS, D.R. 2011a. The extended synthesis: something old, something new. Evolution: Education and Outreach 4: 3-7.
BROOKS, D.R. 2011b. The extended synthesis: the law of the conditions of existence. Evolution: Education and Outreach 4: 254-261.
BROOKS, D.R. 2011c. The major transitions of evolution: visualizing the extended synthesis. Evolution: Education and Outreach 4: 446-452.
BROOKS, D.R. & E.P. HOBERG. 2007. Darwin's necessary misfit and the sloshing bucket: the evolutionary biology of emerging infectious diseases. Evolution: Education & Outreach 1: 2-9.
BROOKS, D.R. & D.A. MCLENNAN. 1990. Searching for a general theory of biological evolution. Journal of Ideas 1: 35-46.
BROOKS, D.R. & D.A. MCLENNAN. 1997. Biological signals as material phenomena. Rev. pensee d'aujord d'hui 25: 118-127. [in Japanese]
BROOKS, D.R. & D.A. MCLENNAN. 2000. The nature of the organism and the emergence of selection processes and biological signals, p. 185-218. In: E. TABORSKY (Ed.). Semiotics, evolution, energy. Aachen, Shaker Verlag.
BROOKS, D.R. & D.A. MCLENNAN. 2002. The nature of diversity: an evolutionary voyage of discovery. Chicago, University of Chicago Press.
BROOKS, D.R. & E.O. WILEY. 1986. Evolution as entropy: toward a unified theory of biology. Chicago, University of Chicago Press.
BROOKS, D.R. & E.O. WILEY. 1988. Evolution as entropy: toward a unified theory of biology. Chicago, University of Chicago Press, 2nd ed.
BROOKS, D.R.; J. COLLIER, B.A. MAURER, J.D.H. SMITH, & E.O. WILEY. 1989. Entropy and information in evolving biological systems. Biology and Philosophy 4: 407-432.
BROOKS, D.R.; D.A. MCLENNAN; V. LEON-REGAGNON & E. HOBERG. 2006a. Phylogeny, ecological fitting and lung flukes: helping solve the problem of emerging infectious diseases. Revista Mexicana de Biodiversidad 77: 225-233.
BROOKS, D.R.; D.A. MCLENNAN; V. LEON-REGAGNON & D. ZELMER. 2006b. Ecological fitting as a determinant of parasite community structure. Ecology 87: S67-S85.
CALCOTT, B. & K. STERELNY. 2011. The major transitions in evolution revisited. Boston, MIT Press.
COLLIER, J. 1998. Information increase in biological systems: How does adaptation fit? p. 129-140 In: G. VAN DE VIJVER, S.N. SALTHE & M. DELPOS (Eds). Evolutionary systems: biological and epistemological perspectives on selection and self-organization. Dordrecht, Kluwer Academic Publisher.
COLLIER, J. 2000. The dynamical basis of information and the origins of semiosis, p. 111-138. In: E. TABORSKY (Ed.). Semiotics, evolution, energy. Aachen, Germany, Shaker Verlag.
COLLIER, J. & C. HOOKER. 1999. Complexly organised dynamical systems. Open Systems and Informtion Dynamics 6: 241-302.
COPE, E.D. 1887. Origin of the fittest: essays on evolution. New York, D. Appleton and Company.
CSANYI, V. 1989. Evolutionary systems and society: a general theory. Durham, Duke University Press.
DAWKINS, R. 1976. The selfish gene. Oxford, Oxford University Press.
DARWIN, C. 1872. The origin of species. London, John Murray, 6th ed.
DEPEW, D. & B. WEBER. 1995. Darwinism evolving. Cambridge, Bradford Books.
ELDREDGE, N. 1985. Unfinished synthesis. New York, Columbia University Press.
ELDREDGE, N. 1995. Reinventing darwin: the great debate at the high table of evolutionary theory. NewYork, John Wiley and Sons.
ELDREDGE, N. 2003. The sloshing bucket: how the physical realm controls evolution, p. 3-32. In: J. CRUCTHFIELD & P. SHUSTER (Eds). Evolutionary dynamics: exploring the interplay of selection, accident, neutrality, and function. SFI Studies in the Sciences of Complexity Series. New York, Oxford University Press.
FISCH, M. & Y. BENBAJI. 2011. The view from within: normativity and the limits of self-criticism. South Bend, University of Notre Dame Press.
FRAUTSCHI, S. 1982. Entropy in an expanding universe. Science 217: 593-599.
FRAUTSCHI, S. 1988. Entropy in an expanding universe, p. 11-22. In: B. WEBER; D.J. DEPEW & J.D. SMITH (Eds). Entropy, information and evolution: new perspectives on physical and biological evolution. Cambridge, MIT Press.
GATLIN, L.L. 1972. Information theory and the living system. New York, Columbia University Press.
GERHARDT, H.C.; M.L. DYSON, S.D. TANNER & C.G. MURPHY. 1994. Female treefrogs do not avoid heterospecific calls as they approach conspecific calls: implications for mechanisms of mate choice. Animal Behavior 47: 1323-1332.
HULL, D.L. 1988. Science as a process: an evolutionary account of the social and conceptual development of science. Chicago, University of Chicago Press.
JABLONKA, E. & M.J. LAMB. 1995. Epigenetic inheritance and evolution. Oxford, Oxford University Press.
JANZEN, D.H. 1980. When is it coevolution? Evolution 34: 611-612.
JANZEN, D.H. 1985. On ecological fitting. Oikos 45: 308-310.
JUARRERO, A. 2002. Dynamics in action: intentional behavior as a complex system. Boston, MIT Press.
KAMPIS, G. 1991. Self-modifying systems in biology and cognitive science: a new framework for dynamics, information and complexity. Oxford, Pergamon.
KAMPIS, G. 1998. Evolution as its own cause and effect, p. 255-265. In: G. VAN DE VIJVER; S.N. SALTHE & M. DELPOS (Eds). Evolutionary systems: biological and epistemological perspectives on selection and self-organization. Dordrecht, Kluwer Academic.
LANDSBERG, P.T. 1984a. Is equilibrium always an entropy maximum? Journal of Statistical Physics 35: 159-169.
LANDSBERG, P.T. 1984b. Can entropy and "order" increase together? Physics Letters 102A: 171-173.
LEGUIN, U. 1974. The dispossessed. New York, Harper Collins.
LOTKA, A.J. 1913. Evolution from the standpoint of physics, the principle of the persistence of stable forms. Scientifc American, Supplement 75: 345-6, 354, 379.
LOTKA, A.J. 1925. Elements of physical biology. Baltimore, Williams and Wilkins.
MAURER, B.A. & D.R. BROOKS. 1991. Energy flow and entropy production in biological systems. Journal of Ideas 2: 48-53.
MAYNARD SMITH, J. 1976. What determines the rate of evolution? American Naturalist 110: 331-338.
MAYNARD SMITH, J. & E. SZATHMÀRY. 1995. The major transitions in evolution. Oxford, W.H. Freeman.
MAYNARD SMITH, J. & E. SZATHMÀRY. 1999. The origins of life. Oxford, Oxford University Press.
MCLENNAN, D.A. 2008. The concept of co-option: why evolution often looks miraculous. Evolution: Education and Outreach 1: 246-258.
MCLENNAN, D.A. & M. J. RYAN. 1999. Interspecific recognition and discrimination based upon olfactory cues in northern swordtails. Evolution 53: 880-888.
MCSHEA, D.W. & R.N. BRANDON. 2011. Biology's first law: the tendency for diversity and complexity to increase in evolutionary systems. Chicago, University of Chicago Press.
MORGAN, T.H. 1932. The scientific basis of evolution. New York, W.W. Norton & Company.
PIGLIUCCI, M. & G.B. MÜLLER. 2010a. Elements of an extended evolutionary synthesis. In: M. PIGLIUCCI & G.B. MÜLLER (Eds). Evolution - the extended synthesis. Cambridge, MIT Press.
PIGLIUCCI, M. & G.B. MÜLLER. 2010b. Evolution - the extended synthesis. Cambridge, MIT Press.
PRIGOGINE, I. 1980. From being to becoming. San Francisco, W. H. Freeman.
PRIGOGINE, I. & J.M. WIAME. 1946. Biologie et thermodynamique des phénomènes irréversìbles. Experientia 2: 451-453.
QUELLER, D.C. 1997. Cooperators since life began. Quarterly Review of Biology 72: 184-188.
SCHRÖDINGER, E. 1945. What is life? Cambridge, Cambridge University Press.
SIMPSON, G.G. 1944. Tempo and mode in evolution. New York, Columbia University Press.
SMITH, J.D.H. 1988. A class of mathematical models for evolution and hierarchical information theory. Institute for Mathematics and its Applications Preprint Series 396: 1-13.
SMITH, J.D.H. 1998. Canonical ensembles, competing species, and the arrow of time, p. 141-154. In: G. VAN DE VIJVER; S.N. SALTHE & M. DELPOS (Eds). Evolutionary systems: biological and epistemological perspectives on selection and self-organization. Dordrecht, Kluwer Academic Publishers.
SMITH, J.D.H. 2000. On the evolution of semiotic capacity, p. 283-309. In: E. TABORSKY (Ed.). Semiotics, evolution, energy. Aachen, Germany, Shaker Verlag.
STOCKING JR, G.W. 1982. Race, culture and evolution: essays in the history of anthropology. Chicago, University of Chicago Press.
SWENSON, R. 1989. Emergent attractors and the law of maximum entropy production: foundations to a theory of general evolution. Systems Research 6: 187-197.
SZATHMÀRY, E. 2000. The evolution of replicators. Philosophical Transactions of the Royal Society of London, Series B 355: 1669-1676.
SZATHMÀRY, E. 2006. Evolution. Darwin for all seasons. Science 313: 306-307.
ULANOWICZ, RE. 1997. Ecology: the ascendent perspective. New York, Columbia University Press.
WANNTORP, H.-E. 1983. Historical constraints in adaptation theory: traits and non-traits. Oikos 41: 157-160.
WEST-EBERHARD, M.J. 2003. Developmental plasticity and evolution. New York, Oxford University Press.
ZOTIN, A.I. & R.S. ZOTINA. 1978. Experimental basis for qualitative phenomenological theory of development, p. 61-84. In: I. LAMPRECHT & A.I. ZOTIN (Eds.). Thermodynamics of biological processes. Berlin, de Gruyter.
Submitted: 10.VII.2012
Accepted: 20.IX.2012.
Editorial responsibility: Walter A.P. Boeger
- AGOSTA, S.J. 2006. On ecological fitting, plant-insect associations, herbivore host shifts, and host plant selection. Oikos 114: 556-565.
- AGOSTA, S.J. & J.A. KLEMENS. 2008. Ecological fitting by pheno-typically flexible genotypes: implications for species associa-tions, community assembly and evolution. Ecology Letters 11: 1123-1134.
- AGOSTA, S.J. & J.A. KLEMENS. 2009. Resource specialization in a phytophagous insect: no evidence for genetically based performance tradeoffs across hosts in the field or laboratory. Journal of Evolutionary Biology 22: 907-912.
- AGOSTA, S.J.; N. JANZ & D.R. BROOKS. 2010. How specialists can be generalists: resolving the "parasite paradox" and implications for emerging infectious disease. Zoologia 27: 151-162.
- BLUM, H.F. 1968. Time's arrow and evolution. Princeton, Princeton University Press, 3rd ed.
- BOWLER, P.J. 1983. The eclipse of Darwinism. Baltimore, John Hopkins University Press.
- BRILLOUIN, L. 1962. Science and information theory. New York, Academic Press, 2nd ed.
- BRODA, E. 1983. DARWIN AND BOLTZMANN, p. 61-70. In: E. GEISSLER & W. SCHELER (Eds). Darwin today: the 8th Kühlungsborn colloquium on philosophical and ethical problems of biosciences. Abhandlungen der Akademien der Wissens-chaf-ten der DDR. Berlin, Akademie.
- BROOKS, D.R. 2010. The mastodon in the room: how Darwinian is neo-Darwinism? Studies in History and Philosophy of Biological and Biomedical Sciences 42: 82-88.
- BROOKS, D.R. 2011a. The extended synthesis: something old, something new. Evolution: Education and Outreach 4: 3-7.
- BROOKS, D.R. 2011b. The extended synthesis: the law of the conditions of existence. Evolution: Education and Outreach 4: 254-261.
- BROOKS, D.R. 2011c. The major transitions of evolution: visualizing the extended synthesis. Evolution: Education and Outreach 4: 446-452.
- BROOKS, D.R. & E.P. HOBERG. 2007. Darwin's necessary misfit and the sloshing bucket: the evolutionary biology of emerging infectious diseases. Evolution: Education & Outreach 1: 2-9.
- BROOKS, D.R. & D.A. MCLENNAN. 1990. Searching for a general theory of biological evolution. Journal of Ideas 1: 35-46.
- BROOKS, D.R. & D.A. MCLENNAN. 1997. Biological signals as material phenomena. Rev. pensee d'aujord d'hui 25: 118-127. [in Japanese]
- BROOKS, D.R. & D.A. MCLENNAN. 2000. The nature of the organism and the emergence of selection processes and biological signals, p. 185-218. In: E. TABORSKY (Ed.). Semiotics, evolution, energy. Aachen, Shaker Verlag.
- BROOKS, D.R. & D.A. MCLENNAN. 2002. The nature of diversity: an evolutionary voyage of discovery. Chicago, University of Chicago Press.
- BROOKS, D.R. & E.O. WILEY. 1986. Evolution as entropy: toward a unified theory of biology. Chicago, University of Chicago Press.
- BROOKS, D.R. & E.O. WILEY. 1988. Evolution as entropy: toward a unified theory of biology. Chicago, University of Chicago Press, 2nd ed.
- BROOKS, D.R.; J. COLLIER, B.A. MAURER, J.D.H. SMITH, & E.O. WILEY. 1989. Entropy and information in evolving biological systems. Biology and Philosophy 4: 407-432.
- BROOKS, D.R.; D.A. MCLENNAN; V. LEON-REGAGNON & E. HOBERG. 2006a. Phylogeny, ecological fitting and lung flukes: helping solve the problem of emerging infectious diseases. Revista Mexicana de Biodiversidad 77: 225-233.
- BROOKS, D.R.; D.A. MCLENNAN; V. LEON-REGAGNON & D. ZELMER. 2006b. Ecological fitting as a determinant of parasite community structure. Ecology 87: S67-S85.
- CALCOTT, B. & K. STERELNY. 2011. The major transitions in evolution revisited. Boston, MIT Press.
- COLLIER, J. 1998. Information increase in biological systems: How does adaptation fit? p. 129-140 In: G. VAN DE VIJVER, S.N. SALTHE & M. DELPOS (Eds). Evolutionary systems: biological and epistemological perspectives on selection and self-organization. Dordrecht, Kluwer Academic Publisher.
- COLLIER, J. 2000. The dynamical basis of information and the origins of semiosis, p. 111-138. In: E. TABORSKY (Ed.). Semiotics, evolution, energy. Aachen, Germany, Shaker Verlag.
- COLLIER, J. & C. HOOKER. 1999. Complexly organised dynamical systems. Open Systems and Informtion Dynamics 6: 241-302.
- COPE, E.D. 1887. Origin of the fittest: essays on evolution. New York, D. Appleton and Company.
- CSANYI, V. 1989. Evolutionary systems and society: a general theory. Durham, Duke University Press.
- DAWKINS, R. 1976. The selfish gene. Oxford, Oxford University Press.
- DARWIN, C. 1872. The origin of species London, John Murray, 6th ed.
- DEPEW, D. & B. WEBER. 1995. Darwinism evolving. Cambridge, Bradford Books.
- ELDREDGE, N. 1985. Unfinished synthesis. New York, Columbia University Press.
- ELDREDGE, N. 1995. Reinventing darwin: the great debate at the high table of evolutionary theory. NewYork, John Wiley and Sons.
- ELDREDGE, N. 2003. The sloshing bucket: how the physical realm controls evolution, p. 3-32. In: J. CRUCTHFIELD & P. SHUSTER (Eds). Evolutionary dynamics: exploring the interplay of selection, accident, neutrality, and function. SFI Studies in the Sciences of Complexity Series. New York, Oxford University Press.
- FISCH, M. & Y. BENBAJI. 2011. The view from within: normativity and the limits of self-criticism. South Bend, University of Notre Dame Press.
- FRAUTSCHI, S. 1982. Entropy in an expanding universe. Science 217: 593-599.
- FRAUTSCHI, S. 1988. Entropy in an expanding universe, p. 11-22. In: B. WEBER; D.J. DEPEW & J.D. SMITH (Eds). Entropy, information and evolution: new perspectives on physical and biological evolution. Cambridge, MIT Press.
- GATLIN, L.L. 1972. Information theory and the living system. New York, Columbia University Press.
- GERHARDT, H.C.; M.L. DYSON, S.D. TANNER & C.G. MURPHY. 1994. Female treefrogs do not avoid heterospecific calls as they approach conspecific calls: implications for mechanisms of mate choice. Animal Behavior 47: 1323-1332.
- HULL, D.L. 1988. Science as a process: an evolutionary account of the social and conceptual development of science. Chicago, University of Chicago Press.
- JABLONKA, E. & M.J. LAMB. 1995. Epigenetic inheritance and evolution. Oxford, Oxford University Press.
- JANZEN, D.H. 1980. When is it coevolution? Evolution 34: 611-612.
- JANZEN, D.H. 1985. On ecological fitting. Oikos 45: 308-310.
- JUARRERO, A. 2002. Dynamics in action: intentional behavior as a complex system. Boston, MIT Press.
- KAMPIS, G. 1991. Self-modifying systems in biology and cognitive science: a new framework for dynamics, information and complexity. Oxford, Pergamon.
- KAMPIS, G. 1998. Evolution as its own cause and effect, p. 255-265. In: G. VAN DE VIJVER; S.N. SALTHE & M. DELPOS (Eds). Evolutionary systems: biological and epistemological perspectives on selection and self-organization. Dordrecht, Kluwer Academic.
- LANDSBERG, P.T. 1984a. Is equilibrium always an entropy maximum? Journal of Statistical Physics 35: 159-169.
- LANDSBERG, P.T. 1984b. Can entropy and "order" increase together? Physics Letters 102A: 171-173.
- LEGUIN, U. 1974. The dispossessed. New York, Harper Collins.
- LOTKA, A.J. 1913. Evolution from the standpoint of physics, the principle of the persistence of stable forms. Scientifc American, Supplement 75: 345-6, 354, 379.
- LOTKA, A.J. 1925. Elements of physical biology. Baltimore, Williams and Wilkins.
- MAURER, B.A. & D.R. BROOKS. 1991. Energy flow and entropy production in biological systems. Journal of Ideas 2: 48-53.
- MAYNARD SMITH, J. 1976. What determines the rate of evolution? American Naturalist 110: 331-338.
- MAYNARD SMITH, J. & E. SZATHMÀRY. 1995. The major transitions in evolution. Oxford, W.H. Freeman.
- MAYNARD SMITH, J. & E. SZATHMÀRY. 1999. The origins of life. Oxford, Oxford University Press.
- MCLENNAN, D.A. 2008. The concept of co-option: why evolution often looks miraculous. Evolution: Education and Outreach 1: 246-258.
- MCLENNAN, D.A. & M. J. RYAN. 1999. Interspecific recognition and discrimination based upon olfactory cues in northern swordtails. Evolution 53: 880-888.
- MCSHEA, D.W. & R.N. BRANDON. 2011. Biology's first law: the tendency for diversity and complexity to increase in evolutionary systems. Chicago, University of Chicago Press.
- MORGAN, T.H. 1932. The scientific basis of evolution. New York, W.W. Norton & Company.
- PIGLIUCCI, M. & G.B. MÜLLER. 2010a. Elements of an extended evolutionary synthesis. In: M. PIGLIUCCI & G.B. MÜLLER (Eds). Evolution - the extended synthesis. Cambridge, MIT Press.
- PIGLIUCCI, M. & G.B. MÜLLER. 2010b. Evolution - the extended synthesis. Cambridge, MIT Press.
- PRIGOGINE, I. 1980. From being to becoming. San Francisco, W. H. Freeman.
- PRIGOGINE, I. & J.M. WIAME. 1946. Biologie et thermodynamique des phénomènes irréversìbles. Experientia 2: 451-453.
- QUELLER, D.C. 1997. Cooperators since life began. Quarterly Review of Biology 72: 184-188.
- SCHRÖDINGER, E. 1945. What is life? Cambridge, Cambridge University Press.
- SIMPSON, G.G. 1944. Tempo and mode in evolution. New York, Columbia University Press.
- SMITH, J.D.H. 1988. A class of mathematical models for evolution and hierarchical information theory. Institute for Mathematics and its Applications Preprint Series 396: 1-13.
- SMITH, J.D.H. 1998. Canonical ensembles, competing species, and the arrow of time, p. 141-154. In: G. VAN DE VIJVER; S.N. SALTHE & M. DELPOS (Eds). Evolutionary systems: biological and epistemological perspectives on selection and self-organization. Dordrecht, Kluwer Academic Publishers.
- SMITH, J.D.H. 2000. On the evolution of semiotic capacity, p. 283-309. In: E. TABORSKY (Ed.). Semiotics, evolution, energy. Aachen, Germany, Shaker Verlag.
- STOCKING JR, G.W. 1982. Race, culture and evolution: essays in the history of anthropology. Chicago, University of Chicago Press.
- SWENSON, R. 1989. Emergent attractors and the law of maximum entropy production: foundations to a theory of general evolution. Systems Research 6: 187-197.
- SZATHMÀRY, E. 2000. The evolution of replicators. Philosophical Transactions of the Royal Society of London, Series B 355: 1669-1676.
- SZATHMÀRY, E. 2006. Evolution. Darwin for all seasons. Science 313: 306-307.
- ULANOWICZ, RE. 1997. Ecology: the ascendent perspective. New York, Columbia University Press.
- ZOTIN, A.I. & R.S. ZOTINA. 1978. Experimental basis for qualitative phenomenological theory of development, p. 61-84. In: I. LAMPRECHT & A.I. ZOTIN (Eds.). Thermodynamics of biological processes. Berlin, de Gruyter.
Publication Dates
-
Publication in this collection
21 Jan 2013 -
Date of issue
Dec 2012