Abstract
Bee fauna and associated flora from a grassland site in Brazil, surveyed 40 and 20 years ago, were newly surveyed with comparable methodology to evaluate changes in the bee fauna of this site, considering that human population and urbanization has exponentially increased in the last 40 years. In general, bee species richness has declined in 22%, as well as their abundance. Some of the previously abundant species are now absent, including Bombus bellicosus Smith, 1879, Gaesischia fulgurans (Holmberg, 1903) and Thectochlora basiatra (Strand, 1910). No particular trend of differential decrease among either taxonomic or functional groups was observed, except for a minor increase in the proportion of oligolectic species and a 50% reduction in the number of large species. The first two surveys were more similar to each other in species richness per bee genus, while the two most recent grouped together based on measures of anthropogenic impact. Furthermore, the number of plant species visited by bees increased, with a pronounced increase in ruderal and exotic species. Crop cultivation, competition with honeybees and climate changes may all be related to bee decline. Nevertheless, the effects of urbanization, in particular intense land occupation and few preserved natural areas can be pointed as the main causes of species decline. Due to continuing increase in human population, increased erosion in diversity is expected. Habitat protection is an additional challenge to bee conservation in the region, with no local conservation units set aside for grasslands. State and municipal agencies should urgently consider the establishment of reserves for the few remaining patches of natural grasslands.
Apidae; Apoidea; habitat fragmentation; pollinators; species richness; urban environments
CONSERVATION
Changes in wild bee fauna of a grassland in Brazil reveal negative effects associated with growing urbanization during the last 40 years
Aline C. MartinsI; Rodrigo B. GonçalvesII; Gabriel A. R. MeloI, * * Corresponding author. E-mail: garmelo@ufpr.br
ILaboratório de Biologia Comparada de Hymenoptera, Departamento de Zoologia, Universidade Federal do Paraná. Caixa Postal 19020, 81531-980 Curitiba, PR, Brazil
IIUniversidade Federal do Paraná. Campus Palotina, 85950-000 Palotina, PR, Brazil
ABSTRACT
Bee fauna and associated flora from a grassland site in Brazil, surveyed 40 and 20 years ago, were newly surveyed with comparable methodology to evaluate changes in the bee fauna of this site, considering that human population and urbanization has exponentially increased in the last 40 years. In general, bee species richness has declined in 22%, as well as their abundance. Some of the previously abundant species are now absent, including Bombus bellicosus Smith, 1879, Gaesischia fulgurans (Holmberg, 1903) and Thectochlora basiatra (Strand, 1910). No particular trend of differential decrease among either taxonomic or functional groups was observed, except for a minor increase in the proportion of oligolectic species and a 50% reduction in the number of large species. The first two surveys were more similar to each other in species richness per bee genus, while the two most recent grouped together based on measures of anthropogenic impact. Furthermore, the number of plant species visited by bees increased, with a pronounced increase in ruderal and exotic species. Crop cultivation, competition with honeybees and climate changes may all be related to bee decline. Nevertheless, the effects of urbanization, in particular intense land occupation and few preserved natural areas can be pointed as the main causes of species decline. Due to continuing increase in human population, increased erosion in diversity is expected. Habitat protection is an additional challenge to bee conservation in the region, with no local conservation units set aside for grasslands. State and municipal agencies should urgently consider the establishment of reserves for the few remaining patches of natural grasslands.
Key words: Apidae; Apoidea; habitat fragmentation; pollinators; species richness; urban environments.
Bees play a fundamental role as pollinators of a large portion of plant species, affecting directly the maintenance and structure of biological communities. The association between bees and flowering plants represents the most important instance of mutualism in the Hymenoptera (LASALLE & GAULD 1993), and the disappearance of bees would be followed by the disappearance of many plant species (BIESMEIJER et al. 2006). Flowers are the main nutritional source for bees and, along with nesting materials, comprise their main habitat requirement (WESTRICH 1996). Thus, pollination is one of the most important ecological interactions for maintaining biodiversity, and one of the most economically valuable ecosystem services (KEVAN & PHILLIPS 2001, KLEIN et al. 2007, GALLAI et al. 2009). The maintenance of bee diversity along seasonal and annual cycles is essential for the efficient pollination of several crops (KLEIN et al. 2003, KREMEN et al. 2002).
Impacts caused by habitat loss and other anthropogenic changes on bee assemblages has been investigated through two basic approachs: 1) by comparisons between areas with varying degrees of human impact during the same time interval (e.g., ZANETTE et al. 2005, CANE et al. 2006), or 2) by comparisons of changes in a specific area through time (e.g., MARLIN & LABERGE 2001, FRANKIE et al. 2009, KEARNS & OLIVERAS 2009a,b, BOMMARCO et al. 2012). These two types of monitoring, together with the study of plants that attract a large number of species or the use of trap-nests, were pointed out as essential for detection and control of the decrease in the number of pollinator populations (ALLEN-WARDEL et al. 1998, FRANKIE et al. 2002).
In the Neotropical region, the loss of habitat - due to deforestation or agricultural development - and the introduction of exotic species comprise the main threats to the bee fauna (FREITAS et al. 2009). However, well-documented examples of species decline are surprisingly uncommon (FREITAS et al. 2009) and only a single case of local extinction has been reported in Brazil (MARTINS & MELO 2009). The high diversity of the Brazilian bee fauna, with about 1,700 species (MELO 2007), is endangered due to a continuous process of urbanization and agricultural development. Here we provide the first documentation of negative impacts of anthropogenic development on the wild bee fauna in South America, based on three surveys conducted over the last 40 years in southern Brazil.
The first survey was conducted in the 1960s in a pioneer study of the bee fauna and associated flora in a grassland site in São José dos Pinhais (Paraná) (SAKAGAMI et al. 1967). This study was repeated two decades later and, although the number of species had remained the same, the number of genera and overall abundance both declined, dramatically for some species (BORTOLI & LAROCA 1990). A great increase in urbanization occurred around the study site between 1960 and 1980 and, in association with the increasing use of agricultural chemicals in the region, were pointed out as the main causes of the marked decline in bee abundance between the two surveys (BORTOLI & LAROCA 1990). Urbanization engenders serious consequences for the maintenance of bee diversity mainly due to the reduction of resources, including nesting sites and flower (food resource) abundance (LAROCA et al. 1982, ZANETTE et al. 2005, CANE 2005, FRANKIE et al. 2009).
To evaluate the current bee fauna and associated flora in this study area, we performed a one-year survey using the same collecting methodology after approximately 40 years since the first study. We identify the major changes in the richness and abundance of the wild bee fauna and flora, focusing on the main trends detected over the last 40 years of increasing urbanization and its impact on taxonomic and functional groups of wild bee fauna.
MATERIAL AND METHODS
The area of these surveys (SAKAGAMI et al. 1967, BORTOLI & LAROCA 1990) is a natural vegetation remnant, within the area of the Afonso Pena International Airport (25°32'S, 49°10'W), in São José dos Pinhais, Paraná, Brazil. The region comprises grassland (in the higher regions) and edaphic fields (in the lower regions subject to water overflow), and woodlands close to the rivers and patches of Araucaria angustifolia (Bertol.) Kunze forests (KLEIN & HATSCHBACH 1962). The climate is the Cfb type (Köppen), always humid, warm-mild pluvial, with more than five occurrences of frost each year.
The first survey in this area is hereafter referred to as S1 (SAKAGAMI et al. 1967), the second as S2 (BORTOLI & LAROCA 1990) and this study as S3. Significant urbanization occurred since S1 and S2, where the natural and agricultural areas were replaced by pavement or buildings (Figs 1-3). In addition to the urban matrix, the region around the study site is occupied by soybean fields within the airport area, and smaller plots of agricultural lands in the surroundings. Consequently our study site constitutes a patch of natural vegetation surrounded by urban or agricultural areas and is a likely refuge for the wild bee fauna.
Data on average temperature and precipitation in the study months during 2004 and 2005 were compiled from National Institute of Meteorology (INMET) database and compared to the normal climate records (1961-1990) for the region of São José dos Pinhais, from the Agronomical Institute of Paraná (IAPAR).
The collecting methodology of all three surveys consisted of sampling with entomological nets any bee visiting all flowering plants, in a predefined area during one year. In S3 the sampling took place at the warmest hours of the day, from 09:00-15:00 h, which includes the peak activity for most bee species (WILLIAMS et al. 2001), at two sites separated by about 2 km and comprising a total area of about 30,000 m2 (Fig. 3). Collecting at each site was carried out by two collectors (ACM and RBG), once a month at each site, from August 2004 to July 2005. The collected specimens were combined into a single sample in our analysis, as in the previous studies. The total sampling hours varies among studies and was more similar between S1 (n = 134 hours) and S2 (n = 150 hours) performed by one collector, while in this study, 117 hours were performed by each of the two collectors during the sampling period (Table I).
In S3, numbers of Trigona spinipes (Fabricius, 1793) and Apis mellifera Linnaeus, 1758 were quantified in the field, since they are the most abundant species and are easily recognizable. Other bees were collected and further identified by RBG, GARM, and Danúncia Urban. Bee higher level classification follows MELO & GONÇALVES (2005). Bee vouchers were stored in the Coleção Entomológica Padre Jesus Santiago Moure (DZUP, Universidade Federal do Paraná) under accession numbers 23689 to 27388. Plant vouchers were deposited in the Botany Department herbarium (UPCB) of the Universidade Federal do Paraná, under accession numbers 58072 to 58179.
In addition to the original publications by SAKAGAMI et al. (1967) and BORTOLI & LAROCA (1990), data on the bee fauna of the 1960s were analyzed by SAKAGAMI & LAROCA (1971) and by S. Laroca in his unpublished master dissertation. We used these latter studies to complement some data not mentioned in the former studies. Taxonomic classification of the previous species lists was updated based on MOURE et al. (2007). Bee vouchers of S1 were not kept together in DZUP and the vouchers of S2 were not made available by the authors. For these reasons, we could not compare morphospecies among the three surveys and so most analyses were at the genus level, a solution adopted in other long-term studies (e.g., KEARNS & OLIVERAS 2009a). In S1 data of abundance were only provided for the most common species, which also limited our comparative analyses.
Bee species were placed in functional groups according to biological features most related with bee ecological habitat requirements (WESTRICH 1996) and body size measured by intertegular distance (ITD) (CANE 1987). The defined groups of traits and respective states were: 1) Nest location (ground; pre-existing cavities); 2) Nesting behavior (solitary; eusocial; cleptoparasite); 3) Bee flower specificity (polylectic, generalist; olygolectic, specialist); and 4) Body size of females (ITD in mm) (1: 0.8-1.39; 2: 1.4-1.89; 3: 1.9-2.29; 4: 2.3-2.9; 5: > 3.0). Measurements of ITD for the bee genera were based on the species collected at the study site. The biological information was based mostly on MICHENER (2007). The biology of some genera is poorly known and, in those cases, traits were deduced from external morphological features, personal observations (GARM) or based on other closely related genera.
To establish a framework of anthropization in the area, we compiled data from the three sampling periods on 1) human population (BREMAEKER 2000, IPPUC 2006); 2) percentage of urban area; and 3) percentage of ruderal/exotic plant species.
Aerial photographs from 1963 and 1980 (Paraná's Environment State Agency - SEMA) and satellite image from 2004 (Urban Development Institute of São José dos Pinhais - IDU) were used to calculate the degree of urbanization in the three periods (Figs 1-3). The images were georeferenced and urban areas were delimited and their size calculated using the software ENVI 4.5 (ITT 2008). We considered as "urban" all areas presenting a regular matrix of buildings and roads, and the remaining non-urban areas were native grass fields, agricultural sites or non-paved roads.
To estimate the percentage of ruderal and exotic species we listed all plant species visited by the bee fauna in the three surveys and classified them by their original habitat and distribution: 1) native to the grasslands of southern Brazil, 2) native to Araucaria forests, and 3) widely spread ruderal native species or ruderal exotic species (non-native to the Brazilian flora). For plant sorting purposes, we were helped by botanists working in the herbariums of the Universidade Federal do Paraná (UPCB) and the Museu Botânico Municipal, Curitiba (MBM), familiar with Paraná's local flora.
The historical data from the previous surveys limited our options for analysis due to the lack of abundance information for most species in S1 and due to the impossibility of comparisons between morphospecies from the three surveys. Thus, for the most abundant species, we used Kato's method (KATO et al. 1952, LAROCA 1995), also employed by BORTOLI & LAROCA (1990) to compare the 1960s and 1980s faunas. This method consists of calculating the limits of relative abundance in percentage of dominant species, which are those presenting the lower confidence limit higher than the upper confidence limit for absent species based on the first survey (S1) (LAROCA 1995). The limits of relative abundance are calculated as follows: upper limit = [(n1.fo)/(n2 + n1.fo)].100, where {n1 = 2(K + 1); n2 = 2(N - K + 1)} and lower limit = [1 - (n1.fo)/(n2 + n1.fo)].100, where {n1 = 2(N - K + 1); n2 = 2(K + 1)}, where "N" is the total amount of individuals collected, "K" is the number of individuals in each group and "fo" is obtained from the table of critical values of the F-distribution, at the 5% level of significance with n1 and n2 degrees of freedom.
The Bray-Curtis similarity index was used to compare the three surveys in relation to number of species per bee genus, as well as for degree of anthropization (see previous section), using the Vegan package version 1.8-8 (OKSANEN et al. 2007) implemented in the software R 2.5.0 (R DEVELOPMENT CORE TEAM 2007).
RESULTS
Current bee fauna and associated flora
The current assemblage of bees at the study site comprises 131 species, 130 of which are native, plus the exotic species Apis mellifera (Table I, Appendix 1). Seventy six (59%) were identified to species, while the other 53 were classified to morphospecies (Appendix 1). A total of 3,061 individuals were sampled, 1681 (59%) were A. mellifera, and 1,380 native species, with Trigona spinipes (n = 173) being the most abundant, followed by Augochlora iphigenia Holmberg, 1886 (118), Ceratina (Rhysoceratina) sp. 1 (84), Paroxystoglossa jocasta (Schrottky, 1910) (80), Ceratina asuncionis Strand, 1910 (69), Augochloropsis cleopatra (Schrottky, 1902) (55), A. iris (Schrottky, 1902) (54), Pseudagapostemon pruinosus Moure & Sakagami, 1984 and Bombus pauloensis Friese, 1913 (49) (Appendix 1). Among the species collected, 99 (77%) are represented by fewer than 10 individuals, 20 (15%) by 11 to 49 individuals and eight (6%) by 50 or more individuals (Appendix 1).
A small portion of the 1,380 specimens, 73 individuals, were collected while they flew over the vegetation, and not on flowers. The remaining species where collected on 95 species of plants in 29 families (Appendix 2). Considering the flora visited by bees, Asteraceae was the most species-rich family, with 38 species, followed by Verbenaceae, with six species. Additionally, Convolvulaceae, Iridaceae, Lamiaceae, Rubiaceae, and Solanaceae stand out with four species each and the other families were represented by fewer than four species (Appendix 2).
Asteraceae had the largest number of bee visits followed by the next most visited families, in decreasing order of number of visits: Brassicaceae, Rosaceae, Malvaceae, Lamiaceae, Verbenaceae, and Apiaceae (Appendix 2). The families showing the largest richness in visiting bee species are: Asteraceae (101 bee species), Apiaceae (22), Brassicaceae (21), Rosaceae (20), Malvaceae (19), Iridaceae (15), Verbenaceae (16), and Solanaceae (14) (Appendix 2). Some plant species stood out among those receiving a large number of visiting individuals: Senecio oleosus Vell., Taraxacum officinale Weber, Baccharis myriocephala DC., and Raphanus raphanistrum L. These four species, together with Stevia veronicae DC., are also those on which the largest richness of bee species were collected (Appendix 2).
Changes in the bee fauna and their associated flora
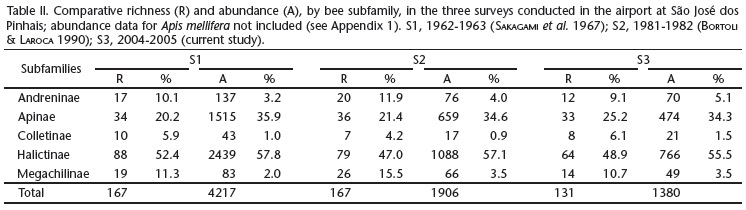
Species richness declined to the present survey, from 167 (S1 and S2) to 131, as well as a substantial decrease in number of individuals sampled, from 4217 (S1) to 1380 (S3) (Table I). S1 had a total of 167 species and 45 genera; S2, the same species richness, but only 37 genera; and S3 a total of 131 species and 39 genera (Appendix 3). In relative terms, taking into consideration the different sampling effort between the surveys, the reduction observed in S3 in both richness and abundance is much stronger, with the species capture rate in S3 reducing to less than 50% of that in S1 and to less than 20% in number of individuals collected per hour (Table I). Since the number of bee species remained unchanged in the interval of 20 years between S1 and S2 (Table I, Appendix 3), this suggests that events of population reduction precede the reduction in species richness, which was detected only in the current study.
The reduction in bee richness and abundance detected in S3 likely represents a long-term pattern and not the result of population fluctuation due to atypical climatic conditions prevalent during the sampling period, since the monthly climate variables (average temperature and precipitation) of the study months followed the averages expected for the region (data not shown; see Methods above).
Considering only the taxa identified to species, of a total of 113 species in the two previous surveys, 20 were absent in the present study (Appendix 4). Since there are no data on relative abundance for most of the species collected in S1 (Appendix 4), it is not possible to infer whether these 20 species are rare and consequently their absence in S3 would be simply a consequence of their rarity. However, some species cited as highly abundant in S1 or S2 are now absent, such as Bombus bellicosus Smith, 1879, Gaesischia fulgurans (Holmberg, 1903) and Thectochlora basiatra (Strand, 1910), and are apparently examples of local extinction (Fig. 4). Also, noticeable cases of population fluctuations were found, such as for Trigona spinipes, Ceratina asuncionis and Bombus pauloensis (Appendix 4, Fig. 4), as well as an increase in abundance of other species, including Augochlora iphigenia, A. amphitrite (Schrottky, 1910), Augochloropsis iris and Bombus morio Skorikov, 1922 (Appendix 4, Fig. 4).
Despite the decrease in richness and overall abundance, the relative proportions among the taxonomic groups are somewhat maintained, suggesting that the decrease in both richness and abundance are not taxonomic trends (Table II). Also, no particular trend is detected among the three surveys when comparing the variation in species diversity within functional groups (Table III). The proportion of species within each functional group seems to stay more or less constant across time, except perhaps for the small relative decrease in polylectic species and a corresponding relative increase in oligolectic species, in the current survey (Table III). Female body size also maintained a similar proportion among the surveys, except for the group of large bees that had a decrease of about 50% from S1 to S3 (category 5 in Table III). Interestingly, the group of large bees (e.g., bumblebees and Anthophora) comprised only four species in S3 compared to 10 in S1 (Table III).
A total of 1681 honeybees were sampled in S3 (Appendix 1). Honeybee abundance was not quantified in the studies of SAKAGAMI et al. (1967) and BORTOLI & LAROCA (1990) and so its recent relative abundance cannot be compared with that of the previous studies. Approximate estimates in SAKAGAMI & LAROCA (1971) suggest (table 1 of that study) that honeybees in S1 were more abundant than all wild bees combined in 26 of the 36 collecting days.
Framework of anthropization
The human population in the municipality of São José dos Pinhais is ten times larger in S3 than it was in S1 (Table IV). At the time of S2 the human population had increased approximately three times, from 20,111 to 70,634 inhabitants in the two decades since S1, and in the 2000s population size was more than 200 thousand inhabitants (Table IV).
The portion occupied by urban areas around the study site grew considerably from the 1960s to 2000s (Figs 1-3). In the 1960s, the urban matrix comprised only ~5% of the total area, increasing to 32% in the 1980s and to 56% in the 2000s (Table IV). The remaining area was considered as non-urban and included crop and pasture fields, patches of native grasslands, small forest fragments, as well as unpaved roads.
The number of plant species visited by bees increased considerably from 67 species in the first survey to 92 species in S3 (Fig. 5). In reference to habitat preference of the plants, the number of species associated with grasslands and forest habitats has remained similar over all studies. On the other hand, the number of ruderal native and exotic plant species has tripled, from 10 (15%) in the first survey to 34 (37%) in this study (Table IV, Fig. 5).
When the three factors analyzed above were taken together as a measure of the degree of anthropization the two most recent surveys (S2 and S3) are more similar to each other, based on Bray-Curtis similarity (BCiS2-S3 = 0.51), than either is with S1 (BCiS1-S3 = 0.18, BCiS1-S2 = 0.44). In terms of bees (species per genus), S1 and S2 are more similar to each other (BCiS1-S2 = 0.79) than to S3 (BCiS1-S3 = 0.74, BCiS2-S3 = 0.67).
DISCUSSION
The clear decline in both species richness and abundance over the past 40 years suggests that large impacts of urbanization may be common. While the current bee fauna is similar to that found in more preserved grassland sites in southern Brazil (GONÇALVES et al. 2009), some species have disappeared (e.g., Bombus bellicosus, Thectochlora basiatra and Gaesischia fulgurans) yet all of the major bee groups and genera seem to have been affected similarly during the intervening period.
Previous studies indicated a differential decrease of functional bee groups subject to habitat loss, such as oligolectic species (BIESMEIJER et al. 2006, CANE et al. 2006). Contrary to what was expected, we found a slight increase in the overall percentage of oligolectic species in S3, and a concomitant decrease in the proportion of polylectic species. This observation should be treated cautiously because current understanding of flower specialization in these species is poor. In addition to polylectic species, large bee species declined in numbers by 50%, while no changes were detected for the other body size categories. Larger bees can forage greater distances than small bees (GREENLEAF et al. 2007), but success in resource acquisition is certainly affected by the quality of the matrix in which they forage, and which here has been strongly impoverished by intense urbanization. Cleptoparasitic bees are also susceptible to habitat disturbance and fragmentation (BORTOLI & LAROCA 1990, LAROCA & ORTH 2002, CALVILLO et al. 2010), but here we found no evidence of decline in this group. These different responses of functional categories among distinct habitats are influenced not only by the disturbance per se, but also by climate, land use history and landscape structure that may come as a consequence of disturbance (MORETTI et al. 2009).
Globally, mostly bee declines have been attributed to habitat fragmentation and loss, agrochemicals, pathogens, alien species, climate change and their interactions (POTTS et al. 2010 and references therein). We propose that the main causes of the accentuated declines that we observed in the wild bee fauna include: 1) habitat loss due to increased human occupation, 2) changes in species composition of the available flower sources due to introduction of exotic plants, 3) global climate changes, and 4) competition with exotic honeybees.
Habitat loss, by urbanization and agricultural development, is considered to be the main cause of bee decline worldwide (FREITAS et al. 2009, POTTS et al. 2010, PATINY et al. 2009, WILLIAMS & OSBORNE 2009). Urbanization results in decreased availability of exposed soil for nests, ground nesting species in particular (CANE 2005, CANE et al. 2006, ZANETTE et al. 2005) as well as for the establishment of food plants within the bee's flying radius. In some urban areas in North America no changes in overall species richness (KEARNS & OLIVERAS 2009a) or variable responses (depending on the functional group) have been observed, such as the increase in richness of bee species that nest in preexisting cavities (CANE et al. 2006, TSCHARNTKE et al. 1998). This response shows that, for some species, urbanization is not always detrimental to all bees.
Areas of natural vegetation remnants in large cities may serve as refuges, depending upon the quality of the surrounding matrix for foraging (MCFREDERICK & LEBUHN 2006, STEFFAN-DEWENTER et al. 2002). Urbanization around our study site has been very intense, leaving almost nothing of the original habitats, as well as with extensive modification of the surrounding matrix (Figs 1-3) due to exponential human population growth (Table IV). Within urban areas, habitat type is a very important influence on species composition, with residential areas and urban peripheries richer in species than city centers (LOSOSOVÁ et al. 2011). Comparing our study site (an urban periphery) to a nearby urban center park (Passeio Público Park in the city of Curitiba), we find a similar trend in which the central city park has a much more impoverished wild bee fauna (TAURA & LAROCA 2001).
Agricultural development at the study site, especially soybeans that today occupies more than 200 ha within the airport area, is likely to have negatively impacted the bees as well. Pesticides along with other agrochemicals may cause impoverishment of bee faunas (e.g., KEARNS et al. 1998, KREMEN et al. 2002) and was previously noted as one of the factors causing the decrease in bee abundance in S2 (BORTOLI & LAROCA 1990). Yet, despite changes in the original vegetation cover, agriculture and cattle ranching may maintain larger portions of suitable habitats for nests and food plants for bees (MARLIN & LABERGE 2001), with less impact than that due to urban sprawl (CANE 2005).
Long-term changes at the study site include floral composition visited by bees, and an increase in the number of visited plants as a consequence of the increase in the number of ruderal and exotic species. In the 1960s, ruderal plants comprised 15% of those visited by bees, while now they comprise 37% (Table IV). Also, the number of introduced plant species increased in S2 in the 20-year interval between the first two surveys (BORTOLI & LAROCA 1990: fig. 19). Functional diversity in plants may often be more important than species richness (DÍAZ & CABIDO 2001) and thus a substantial increase in the number of exotic plant species is likely to influence the availability of adequate food sources.
In the last few decades, global climate change caused by humans has threatened the survival of several species around the world (WILLIAMS et al. 2007). Although not widely studied in insects, climate change is likely to be a critical factor that will influence changes in species distribution and local extinction events. Climate change has been suggested as the main cause for local extinction of Bombus bellicosus at our study site and elsewhere in the state of Paraná (MARTINS & MELO 2009). This bumblebee is associated with regions with mild climates in southern South America, and Paraná is the northern limit of its original distribution (MARTINS & MELO 2009). Other factors, however, are also important causes of bumblebee decline worldwide (WILLIAMS & OSBORNE 2009), such as changing land use patterns, introduction of parasites from commercial colonies (MEEUS et al. 2011) and changes in the yearly supply of flower resources (CARVELL et al. 2006). Differences in tongue length and diet breadth of bumblebees living in the same region have been implicated in their differential decline (BOMMARCO et al. 2012). Nonetheless, because there are no records of exotic bumblebees, particularly Bombus terrestris (Linnaeus, 1758), in Brazil and since other bumblebee species in the area have not been affected, the parasite hypothesis seems unlikely to be involved in the local extinction of B. bellicosus and so climate change remains as an important possibility.
Honeybees were reported from the study site since S1 (SAKAGAMI & LAROCA 1971) and were dominant in all three surveys. Potential and real impacts of Apis mellifera on the native bee fauna in the Neotropical region have not been conclusively resolved (e.g., PEDRO & CAMARGO 1991, WILMS & WIECHERS 1997, ROUBIK & WOLDA 2001). Honeybees as competitors is undeniable: populations are large, their foraging spectrum is vast and they remain active throughout the year (in tropical and subtropical regions), with extensive niche overlap with native species. This only implies competition for resources if their availability may be limiting (GOULSON 2003). Nevertheless, no studies have yet concluded that feral honeybees cause extinction of native bees (MORITZ et al. 2005) with a few exceptions, such as on islands (KATO et al. 1999, KATO & KAWAKITA 2004).
The long-term comparisons presented here document a steady decline in the diversity and abundance of the native bees at our study site and the negative effects of rapid urbanization taking place in most Brazilian cities. As the number of inhabitants is continually growing in the area - with already more than 250,000 in 2010 (IBGE 2010) - a continued impoverishment over time of the bee fauna is expected. The natural grasslands of southern Brazil are extremely endangered and poorly protected by few natural parks, especially in the state of Paraná (OVERBECK et al. 2009, VÉLEZ et al. 2009). Habitat protection is an additional challenge to bee conservation in the region, with no local conservation units set aside for grasslands. State and municipal agencies should urgently consider the establishment of reserves that include in addition to forest remnants, the few remaining patches of natural grasslands. Last but not least, to preserve these essential pollinators in the urban environments, Brazilian cities should include a larger portion of suitable places to wild bee fauna in their planning (e.g., city parks, squares and bee-friendly gardens).
ACKNOWLEDGEMENTS
We would like to thank Danúncia Urban for identification of the Anthidiini and Eucerini bees and Osmar S. Ribas, Gert Hatschbach and Olavo Guimarães (in memorian) for the plant identifications. We are also grateful to Gabriel A.R. de Paula for allowing us to use unpublished data from his Master's studies of ITD of female bees collected at the Afonso Pena Airport. We thank the Brazilian Company for Airport Infrastructure (INFRAERO) for permission to carry out field work at the Afonso Pena Airport; the Secretary of Environment of Paraná State (SEMA) and Institute for Urban Development (IDU) for providing the aerial photographs and satellite images. We would like to thank to Antonio J.C. Aguiar for help in field work and motivation. We also acknowledge the CNPq for scholarships to ACM and RBG and research scholarship to GARM. Thanks to James J. Roper for his help with the English.
LITERATURE CITED
Submitted: 10.VIII.2012; Accepted: 24.X.2012.
Editorial responsibility: Kleber Del Claro
Click to enlarge
Click to enlarge
Click to enlarge
Click to enlarge
- ALLEN-WARDELL, G.; P. BERNHARDT; R. BITNER; A. BURQUEZ; S. BUCHMANN; J. CANE; P.A. COX; V. DALTON; P. FEINSINGER; M. INGRAM; D. INOUYE; C.E. JONES; K. KENNEDY; P. KEVAN; H. KOOPOWITZ; R. MEDELLIN; S. MEDELLIN-MORALES; G.P. NABHAN; B. PAVLIK; V. TEPEDINO; P. TORCHIO & S. WALKER. 1998. The potential consequences of pollinator declines on the conservation of biodiversity and stability of food crop yields. Conservation Biology 12 (1): 8-17.
- BIESMEIJER, J.C.; S.P.M. ROBERTS; M. REEMER; R. OHLEMULLER; M. EDWARDS; T. PEETERS; A.P. SCHAFFERS; S.G. POTTS; R. KLEUKERS; C.D. THOMAS; J. SETTELE & W.E. KUNIN. 2006. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 313 (5785): 351-354.
- BOMMARCO, R.; O. LUNDIN; H.G. SMITH & M. RUNDLÖF. 2012. Drastic historic shifts in bumble-bee community composition in Sweden. Proceedings of the Royal Society B 22 (1727): 309-315.
- BORTOLI, C. & S. LAROCA. 1990. Estudo biocenótico em Apoidea (Hymenoptera) de uma área restrita em São Jos dos Pinhais (PR, Sul do Brasil), com notas comparativas. Dusenia 15: 1-112.
- BREMAEKER, F.E.J. 2000. Evolução demográfica dos municípios das regiões metropolitanas brasileiras, segundo a base territorial de 1997 (1960/1991). Curitiba, Instituto Brasileiro de Administração Municipal (IBAM), Séries estudos demográficos no. 19.
- CALVILLO, L.M.; V.M. RAMÍREZ; V. PARRA-TABLA & J. NAVARRO. 2010. Bee diversity in a fragmented landscape of the Mexican Neotropic. Journal of Insect Conservation 14 (4): 323-334.
- CANE, J.H. 1987. Estimation of bee size using intertegular span (Apoidea). Journal of the Kansas Entomological Society 60 (1): 145-147.
- CANE, J.H. 2005. Bees, pollination and the challenges of sprawl, p. 109-124. In: E.A. JOHNSON & M.W. KLEMENS (Eds). Nature in Fragments: the Legacy of Sprawl. New York, Columbia University Press.
- CANE, J.H.; R.L. MINCKLEY; L.J. KERVIN; T.H. ROULSTON & N.M. WILLIAMS. 2006. Complex responses within a desert bee guild (Hymenoptera: Apiformes) to urban habitat fragmentation. Ecological Applications 16 (2): 632-644.
- CARVELL, C.; D. ROY; S. SMART; R. PYWELL; C. PRESTON & D. GOULSON. 2006. Declines in forage availability for bumblebees at a national scale. Biological Conservation 132 (4): 481-489.
- DÍAZ, S. & M. CABIDO. 2001. Vive la différence: plant functional diversity matters to ecosystem processes. Trends in Ecology and Evolution 16 (11): 646-655.
- FRANKIE, G.W.; S.B. VINSON; R.W. THORP; M.A. RIZZARDI; M. TOMKINS; L.E. NEWSTROM-LLOYD. 2002. Monitoring: an essential tool in bee ecology and conservation, p. 187-198. In: P.G. KEVAN & V.L. IMPERATRIZ-FONSECA (Eds). Pollinating Bees: the Conservation Link Between Agriculture and Nature. Brasília, Ministério do Meio Ambiente.
- FRANKIE, G.W.; M. RIZZARDI; S.B. VINSON & T.L. GRISWOLD. 2009. Decline in bee diversity and abundance from 1972-2004 on a flowering leguminous tree, Andira inermis in Costa Rica at the interface of disturbed dry forest and the urban environment. Journal of the Kansas Entomological Society 82 (1): 1-20.
- FREITAS, B.M.; V.L. IMPERATRIZ-FONSECA; L. M. MEDINA; A.M.P. KLEINERT; L. GALETTO; G. NATES-PARRA & J.J.G. QUEZADA-EUÁN. 2009. Diversity, threats and conservation of native bees in the Neotropics. Apidologie 40 (3): 332-346.
- GALLAI, N.; J. SALLES; J. SETTELE & B.E. VAISSIÈRE. 2009. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecological Economics 68 (3): 810-821.
- GONÇALVES, R.B.; G.A.R. MELO & A.J.C. AGUIAR. 2009. A assembléia de abelhas (Hymenoptera, Apidae) de uma área restrita de campos naturais do Parque Estadual de Vila Velha, Paran e comparações com áreas de campos e cerrado. Papéis Avulsos de Zoologia 49 (14): 163-181.
- GOULSON, D. 2003. Effects of introduced bees on native ecosystems. Annual Review of Ecology and Systematics 34: 1-26.
- GREENLEAF, S.S.; N.M. WILLIAMS; R. WINFREE & C. KREMEN. 2007. Bee foraging ranges and their relationship to body size. Oecologia 153 (3): 589-96.
- IBGE. 2010. Censo 2010. Instituto Brasileiro de Geografia e Estatística. Available online at: http://www.ibge.gov.br/censo2010/resultados_do_censo2010.php [Accessed: 18/II/2011]
- IPPUC. 2006. População e taxa de crescimento anual dos municípios da região metropolitana de Curitiba, Paran, Brasil - 1970 - 2005. Instituto de Pesquisa e Planejamento Urbano de Curitiba. Available online at: http://www.ippuc.org.br [Accessed: 06/VIII/2006]
- ITT. 2008. ENVI 4.5. Environment for visualizing images. Visual Information Solutions (ITT). Boulder.
- KATO, M. & A. KAWAKITA. 2004. Plant-pollinator interactions in New Caledonia influenced by introduced honey bees. American Journal of Botany 91 (11): 1814-1827.
- KATO, M.; T. MATSUTA & Z. YAMASHITA.1952. Associative ecology of insects found in the paddy field cultivated by various planting forms. Science Reports Tohoku University, IV (Biology) 19: 291-301.
- KATO, M.; A. SHIBATA; T. YASUI & H. NAGAMASU. 1999. Impact of introduced honeybees, Apis mellifera, upon native bee communities in the Bonin (Ogasawara) Islands. Research in Population Ecology 41 (2): 217-228.
- KEARNS, C.A.; D.W. INOUYE & N.M. WASER. 1998. Endangered mutualisms: the conservation of plant-pollinator interactions. Annual Review of Ecology and Systematics 29: 83-112.
- KEARNS, C.A. & D.M. OLIVERAS. 2009a. Environmental factors affecting bee diversity in urban and remote grassland plots in Boulder, Colorado. Journal of Insect Conservation 13 (6): 655-665.
- KEARNS, C.A. & D.M. OLIVERAS. 2009b. Boulder County bees are revisited: a resampling of Boulder Colorado bees a century later. Journal of Insect Conservation 13 (6): 603-613.
- KEVAN, P.G. & T.P. PHILLIPS. 2001. The economic impact of pollinator declines: an approach to assessing the consequences. Conservation Ecology 15 (1): 8.Available online at: http://www.consecol.org/vol5/iss1/art8/ [Accessed: 18/II/2011]
- KLEIN, A.M.; I. STEFFAN-DEWENTER & T. TSCHARNTKE. 2003. Fruit set of highland coffee increases with the diversity of pollinating bees. Proceedings of Royal Society B 270 (1518): 955-961.
- KLEIN A.M.; B.E. VAISSIÈRE; J.H. CANE; I. STEFFAN-DEWENTER; S.A. CUNNINGHAM; C. KREMEN & T. TSCHARNTKE. 2007. Importance of pollinators in changing landscapes for world crops. Proceedings of Royal Society B 274 (1608): 303-313.
- KLEIN, R.M. & G. HATSCHBACH. 1962. Fitofisionomia e notas sobre a vegetação para acompanhar a planta fitogeográfica do município de Curitiba e arredores. Boletim da Universidade do Paraná, Geografia Física 4: 1-30.
- KREMEN, C.; N. WILLIAMS & R.W. THORP. 2002. Crop pollination from native bees at risk from agricultural intensification. Proceedings of National Academy of Sciences 99 (26): 16812-16816.
- LAROCA, S. 1995. Ecologia: princípios e métodos. Petrópolis, Editora Vozes.
- LAROCA, S. & I. ORTH. 2002. Melissocoenology: historical perspective, method of sampling, and recommendations to the Program of Conservation and Sustainable Use of Pollinators, with Emphasis on Bees (ONU), p. 217-225. In: P.G. KEVAN & V.L. IMPERATRIZ-FONSECA (Eds). Pollinating Bees: the Conservation Link Between Agriculture and Nature. Brasília, Ministério do Meio Ambiente.
- LAROCA, S.; J.R. CURE-HAKIM & C. BORTOLI. 1982. A associação das abelhas silvestres (Hymenoptera, Apoidea) de uma área restrita no interior da cidade de Curitiba (Brasil): uma abordagem biocenótica. Dusenia 13 (3): 93-117.
- LASALLE, J. & I.D. GAULD. 1993. Hymenoptera: their diversity, and their impact on the diversity of other organisms, p. 1-26. In: J. LASALLE & I.D. GAULD (Eds). Hymenoptera and Biodiversity. Wallingford, The Natural History Museum, C.A.B. International.
- LOSOSOVÁ Z.; M. HORSÁK; M. CHYTRÝ; T. ÈEJKA; J. DANIHELKA; K. FAJMON; O. HÁJEK. 2011. Diversity of Central European urban biota: effects of human-made habitat types on plants and land snails. Journal of Biogeography 38 (6): 1152-1163.
- MARLIN, J.C. & W.E. LABERGE. 2001. The native bee fauna of Carlinville, Illinois, revisited after 75 years: a case for persistence. Conservation Ecology 15 (1): 9. Available online at: http://www.consecol.org/vol5/iss1/art9/ [Accessed: 18/II/2011]
- MARTINS, A.C. & G.A.R. MELO. 2009. Has the bumblebee Bombus bellicosus gone extinct in the northern portion of its distribution range? Journal of Insect Conservation 14 (2): 207-210.
- MCFREDERICK, Q.S. & G. LEBUHN G. 2006. Are urban parks refuges for bumble bees Bombus spp. (Hymenoptera, Apidae)? Biological Conservation 129 (3): 372-382.
- MEEUS, I.; M.J.F. BROWN; D.C. DE GRAAF & G. SMAGGHE. 2011. Effects of invasive parasites on bumblebee declines. Conservation Biology 25 (4): 662-71.
- MELO, G.A.R. 2007. Introductory Remarks, p. V-XI. In: J.S. MOURE, D. URBAN & G.A.R. MELO (Eds). Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region. Curitiba, Sociedade Brasileira de Entomologia.
- MELO, G.A.R. & R.B. GONÇALVES. 2005. Higher-level bee classifications (Hymenoptera, Apoidea, Apidae sensu lato). Revista Brasileira de Zoologia 22 (1): 153-159.
- MICHENER, C.D. 2007. The Bees of the World. 2nd Ed. Baltimore, Johns Hopkins University Press.
- MORETTI, M.; F.D. BELLO; S.P. ROBERTS & S.G. POTTS. 2009. Taxonomical vs functional responses of bee communities to fire in two contrasting climatic regions. Journal of Animal Ecology 78 (1): 98-108.
- MORITZ, R.F.A.; S. HÄRTEL & P. NEUMANN. 2005. Global invasions of the western honeybee (Apis mellifera) and the consequences for biodiversity. Ecoscience 12 (3): 289-301.
- MOURE, J.S.; D. URBAN & G.A.R. MELO. 2007. Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region. Curitiba, Sociedade Brasileira de Entomologia.
- OKSANEN, J.; R. KINDT; P. LEGENDRE; B. O'HARA & M.H.H. STEVENS. 2007. Vegan: Community Ecology Package. R package version 2.12-1. Available online at: http://cran.r-project.org [Accessed: 17/II/2011]
- OVERBECK, G.E.; S.C. MÜLLER; A. FIDELIS; J. PFADENHAUER; V.P. PILLAR; C.C. BLANCO; I.I. BOLDRINI; R. BOTH & E.D. FONSECA. 2009. Os Campos Sulinos: um bioma negligenciado, p. 26-41. In: V.P. PILLAR; S.C. MÜLLER; Z.M.S. CASTILHOS & A.V.A. JACQUES (Eds). Campos Sulinos: Conservação e Uso Sustentável da Biodiversidade. Brasília, Ministério do Meio Ambiente.
- PATINY, S.; P. RASMONT & D. MICHEZ. 2009. A survey and review of the status of wild bees in the West-Palaearctic region. Apidologie 40 (3): 313-331.
- POTTS, S.G.; J.C. BIESMEIJER; C. KREMEN; P. NEUMANN; O. SCHWEIGER &W.E. KUNIN. 2010. Global pollinator declines: trends, impacts and drivers. Trends in Ecology and Evolution 25 (6): 345-53.
- PEDRO, S.M.R. & J.M.F. CAMARGO. 1991. Interactions on floral resources between the Africanized honey bee Apis mellifera L. and the native bee community (Hymenoptera: Apoidea) in a natural "cerrado" ecosystem in southeast Brazil. Apidologie 22 (4): 397-415.
- R DEVELOPMENT CORE TEAM. 2010. R: A language and environment for statistical computing. R foundation for Statistical Computing. Available online at: http://cran.r-project.org [Accessed: 17/II/2011]
- ROUBIK, D.W. & H. WOLDA. 2001. Do competing honey bees matter? Dynamics and abundance of native bees before and after honey bee invasion. Population Ecology 43 (1): 53-62.
- SAKAGAMI, S.F. & L. LAROCA. 1971. Relative abundance, phenology and flower visits of apid bees in eastern Paraná, southern Brazil (Hymenoptera, Apidae). Kontyû 39 (3): 217-230.
- SAKAGAMI, S.F.; S. LAROCA & J.S. MOURE. 1967. Wild bee biocenotics in So Jos dos Pinhais (PR), South Brazil, Preliminary Report. Journal of the Faculty of Science Hokkaido University, Series VI, Zoology 16: 253-291.
- STEFFAN-DEWENTER, I.; U. MÜNZENBERG; C. BÜRGER; C. THIES & T. TSCHARNTKE. 2002. Scale dependent effects of landscape context on three pollinator guilds. Ecology 83 (5): 1421-1432.
- TAURA, H.M. & S. LAROCA. 2001. A associação das abelhas silvestres de um biótopo urbano de Curitiba (Brasil), com comparações espaço-temporais: abundância relativa, fenologia, diversidade e exploração de recursos (Hymenoptera, Apoidea). Acta Biologica Paranaense 30 (1-4): 35-137.
- TSCHARNTKE, T.; A. GATHMANN & I. STEFFAN-DEWENTER I. 1998. Bioindication using trap-nesting bees and wasps and their natural enemies: community structure and interactions. Journal of Applied Ecology 35 (5): 708-719.
- VÉLEZ, E.; L. CHOMENKO; W. SCHAFFER & M. MADEIRA. 2009. Um panorama sobre as iniciativas de conservação dos Campos Sulinos, p. 356-379. In: V.P. PILLAR; S.C. MÜLLER; Z.M.S. CASTILHOS & A.V.A. JACQUES (Eds). Campos Sulinos: Conservação e Uso Sustentvel da Biodiversidade. Brasília, Ministério do Meio Ambiente.
- WESTRICH, P. 1996. Habitat requirements of central European bees and the problems of partial habitats, p. 1-16. In: A. MATHESON; S.L. BUCHMANN; C. O'TOOLE; P. WESTRICH & I. WILLIANS (Eds). The Conservation of Bees. New York, Academic Press.
- WILLIAMS, J.W.; S.T. JACKSON & J.E. KUTZBACH. 2007. Projected distributions of novel and disappearing climates by 2100 AD. Proceedings of National Academy of Sciences 104 (14): 5738-5742.
- WILLIAMS, P.H. & J.L. OSBORNE. 2009. Bumblebee vulnerability and conservation world-wide. Apidologie 40 (3): 367-387.
- WILLIAMS, N. M.; R. L. MINCKLEY & F.A. SILVEIRA. 2001. Variation in native bee faunas and its implications for detecting community changes. Conservation Ecology 5 (1): 7 Avalaible online at: http://www.consecol.org/vol5/iss1/art7/ [Accessed: 18/II/2011]
- WILMS, W. & B. WIECHERS. 1997. Floral resource partitioning between native Melipona bees and the introduced Africanized honey bee in the Brazilian Atlantic rain forest. Apidologie 28 (6): 339-355.
- ZANETTE, L.R.S.; R.P. MARTINS & S.P. RIBEIRO. 2005. Effects of urbanization on neotropical wasp and bee assemblages in a Brazilian metropolis. Landscape and Urban Planning 71 (2-4): 105-121.




Publication Dates
-
Publication in this collection
09 May 2013 -
Date of issue
Apr 2013
History
-
Received
10 Aug 2012 -
Accepted
24 Oct 2012