Abstract
In this study we evaluated how anuran species were distributed in riparian habitats along two large rivers. Sampling was carried out between January and March 2012 in the municipality of Cachoeiras de Macacu, state of Rio de Janeiro. We delimited 20 plots along each river, ten in portions inside the forest of the Reserva Ecológica de Guapiaçu (REGUA), and with comparatively greater amount of forest cover, and ten outside REGUA, with comparatively lesser forest cover surrounding the rivers. We recorded 70 individuals from 14 frog species in the Manoel Alexandre River and 63 individuals from 15 frog species in the Guapiaçu River. The most abundant species in both rivers was Cycloramphus brasiliensis (Steindachner, 1864), and it was more abundant in sections with greater amount of forest cover. This information, coupled with the occurrence of species that are more adapted to open and more disturbed habitats in river sections that harbor lesser riparian vegetation, help to explain differences in amphibian species composition between river sections with greater and lesser forest cover. The results of our study highlight the importance of preserving riparian vegetation associated with rivers in the Atlantic Forest for the conservation of amphibians.
Amphibian conservation; aquatic environments; connectivity; frog species; riparian forests
Land-use change represents a serious threat to amphibians, and poses a challenge for their
conservation (Hof et al. 2011Hof C, Araújo MB, Jetz W, Rahbek C (2011) Additive threats from
pathogens, climate and land-use change for global amphibian diversity. Nature 480:
516-519. doi: 10.1038/nature10650
https://doi.org/10.1038/nature10650...
). Local amphibian
communities can be structured by several factors, some of which are affected by land use.
Examples are the size of forest fragments (Bell &
Donnelly 2006Bell KE, Donnelly MA (2006) Influence of forest fragmentation on
community structure of frogs and lizards in northeastern Costa Rica. Conservation
Biology20(6): 1750-1760. doi: 10.1111/j.1523-1739.2006.00522.x
https://doi.org/10.1111/j.1523-1739.2006...
), the presence of suitable reproductive sites (Zimmerman & Bierregaard 1986Zimmerman BL, Bierregaard RO (1986) Relevance of the equilibrium theory
of island biogeography and species-area relations to conservation with a case from
Amazonia. Journal of Biogeography 13(2): 133-143. doi:
10.2307/2844988
https://doi.org/10.2307/2844988...
), and the presence of
riparian vegetation (Parris & McCarthy 1999Parris KM, McCarthy MA (1999) What influences the structure of frog
assemblages at forest streams? Australian Journal of Ecology 24(5): 495-502. doi:
10.1046/j.1442-9993.1999.00989.x
https://doi.org/10.1046/j.1442-9993.1999...
).
Riparian habitats can be important for maintaining the structural and physicochemical
characteristics of aquatic environments, hence they are important factors determining
community composition in these habitats (Gomi et al.
2006Gomi T, Sidle RC, Noguchi S, Negishi JN, Nik AR, Sasaki S (2006)
Sediment and wood accumulations in humid tropical headwater streams: Effects of
logging and riparian buffers. Forest Ecology and Management 224(1-2): 166-175. doi:
10.1016/j.foreco.2005.12.016
https://doi.org/10.1016/j.foreco.2005.12...
). In fact, anuran assemblages are frequently structured along a continuum of
riparian habitats (e.g., Keller et al. 2009Keller A, Rödel M-O, Linsenmair E, Grafe TU (2009) The importance of
environmental heterogeneity for species diversity and assemblage structure in Bornean
stream frogs. Journal of Animal Ecology 78: 305-314. doi:
10.1111/j.1365-2656.2008.01457.x
https://doi.org/10.1111/j.1365-2656.2008...
, Rodríguez-Mendoza & Pineda 2010Rodríguez-Mendoza C, Pineda E (2010) Importance of riparian remnants for
frog species diversity in a highly fragmented rainforest. Biology Letters 6: 781-784.
doi: 10.1098/rsbl.2010.0334
https://doi.org/10.1098/rsbl.2010.0334...
), which can be
crucial for vulnerable amphibian species in tropical forest areas. Besides, riparian
habitats provide connectivity between forest patches and facilitate gene flow between
amphibian populations (Richards-Zawacki 2009Richards-Zawacki CL (2009) Effects of slope and riparian habitat
connectivity on gene flow in an endangered Panamanian frog, Atelopus varius.
Diversity and Ditributions 15: 796-806. doi:
10.1111/j.1472-4642.2009.00582.x
https://doi.org/10.1111/j.1472-4642.2009...
). Since
many amphibian species depend on these environments for reproduction (Haddad & Prado 2005Haddad CFB, Prado CPA (2005) Reproductive modes in frogs and their
unexpected diversity in the Atlantic Forest of Brazil. BioScience 55(3):
207-217.), and some of them live all their life cycles
closely associated with water bodies that are near riparian vegetation, preservation of the
latter may be critical to the viability of several species (Ficetola et al. 2009Ficetola GF, Padoa-Schioppa E, Bernardi F (2009) Influence of landscape
elements in riparian buffers on the conservation of semiaquatic amphibians.
Conservation Biology23(1): 114-123. doi:
10.1111/j.1523-1739.2008.01081.x
https://doi.org/10.1111/j.1523-1739.2008...
, Todd et al.
2009Todd BD, Luhring TM, Rothermel BB, Gibbons W (2009) Effects of forest
removal on amphibian migrations: implications for habitat and landscape connectivity.
Journal of Applied Ecology 46: 554-561. doi:
10.1111/j.1365-2664.2009.01645.x
https://doi.org/10.1111/j.1365-2664.2009...
).
The Atlantic Forest has high biodiversity and endemism, being among the five most
threatened biomes in the world (Mittermeier et al.
2005Mittermeier RA, Gil RP, Hoffman M, Pilgrim J, Brooks T, Mittermeier CG,
Lamoreux J, Fonseca GAB (2005) Hotspots revisited: earth's biologically richest and
most endangered terrestrial ecoregions. Washington, DC, Conservation International,
392p.). However, despite the fact that human interference has reduced the Atlantic
Forest to about 12% of its original area (Ribeiro et al.
2009Ribeiro MC, Metzger JP, Martensen AC, Ponzoni FJ, Hirota MM (2009) The
Brazilian Atlantic Forest: how much is left, and how is the remaining forest
distributed? Implications for conservation. Biological Conservation 142: 1144-1156.
doi:10.1016/j.biocon.2009.02.021
https://doi.org/10.1016/j.biocon.2009.02...
), it still harbors more than 480 amphibian species. Most of these species
have an aquatic larval phase (Becker et al. 2007Becker CG, Fonseca CR, Haddad CFB, Batista RF, Prado PI (2007) Habitat
split and the global decline of amphibians. Science 318(5857): 1775-1777. doi:
10.1126/science.1149374
https://doi.org/10.1126/science.1149374...
).
Since the mid 2010s there has been an intense discussion in Brazil regarding a review of
the current Brazilian Forest Act (Metzger et al.
2010Metzger JP, Lewinsohn TM, Joly CA, Verdade LM, Martinelli LA, Rodrigues
RR (2010) Brazilian Law: full speed in reverse? Science329: 276-277.). Among the most controversial points of this review is the proposed
reduction of the minimum width of riparian vegetation in each property, which can result in
serious consequences for the persistence of many amphibian species. This situation is
worsened by the fact that even under the current legislation the protection area of
riparian vegetation has not been enforced (Becker et al.
2007Becker CG, Fonseca CR, Haddad CFB, Batista RF, Prado PI (2007) Habitat
split and the global decline of amphibians. Science 318(5857): 1775-1777. doi:
10.1126/science.1149374
https://doi.org/10.1126/science.1149374...
). In this context, it is urgent to gather field data to evaluate the
importance of riparian vegetation for amphibian species and local communities. It is still
unknown how riparian forest loss affects anuran communities in Atlantic Forest rivers. The
aim of this study was to compare anuran species composition in riparian habitats of two
rivers with different amounts of forest cover (greater and lesser forest cover). We
expected that community composition would differ between plots with greater and lesser
riparian forest cover: those with more deforested plots were expected to harbor more
generalist species than plots with greater forest cover, which were expected to be mostly
occupied by anuran species that are strictly adapted to forests.
MATERIAL AND METHODS
We conducted this study between January and March 2012 in the Reserva Ecológica de
Guapiaçu, REGUA (22°24'S, 42°44'W), in the municipality of Cachoeiras de Macacu, state
of Rio de Janeiro, Brazil. REGUA is a private reserve comprising about 7,600 ha of
Atlantic forest, most of which is continuous with the large forest patch of the Serra
dos Órgãos mountain range (Almeida-Gomes & Rocha
2014Almeida-Gomes M, Siqueira CC, Borges-Júnior VNT, Vrcibradic D, Fusinatto
LA, Rocha CFD (2014b) Herpetofauna of the Reserva Ecológica de Guapiaçu (REGUA) and
its surrounding areas, in the state of Rio de Janeiro, Brazil. Biota Neotropica
14(3): 1-15. doi: 10.1590/1676-0603007813
https://doi.org/10.1590/1676-0603007813...
). Sampling was carried out in two rivers, Manoel Alexandre and Guapiaçu
(Figs. 1-3). Both rivers have a gradient of riparian forest cover: the portions of rivers
inside REGUA have greater forest cover, which progressively reduces as the rivers leave
REGUA. Rivers outside REGUA have little riparian vegetation surrounded mostly by pasture
areas. A total of 20 plots, each 20 m in length were delimited along each river, 10
inside and 10 outside REGUA, totaling 400 m sampled in each river. Each plot ranged from
one margin of the river to the other, and was separated from the nearest plot by at
least 10 m. To limit altitudinal variation, the maximum difference in altitude between
upstream and downstream plots was approximately 100 m at the Manoel Alexandre River
(108-220 m), and 60 m at the Guapiaçu River (39-98 m).

Location of Reserva Ecológica de Guapiaçu (REGUA, black square) in the state of Rio de Janeiro.

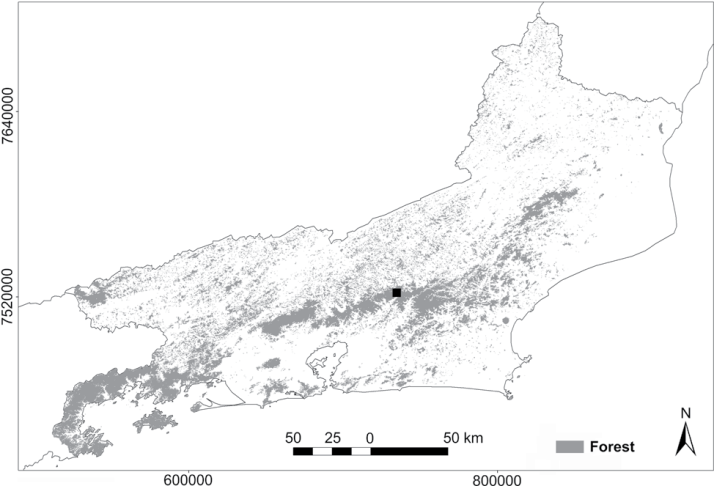
Study area indicating the location of plots along the Manoel Alexandre (2) and Guapiaçu (3) rivers, municipality of Cachoeiras de Macacu, state of Rio de Janeiro, Brazil. (squares) Plots with greater forest cover, (triangles) plots with lesser forest cover.
We conducted visual encounter surveys, VES (Crump &
Scott 1994Crump ML, Scott Jr NJ (1994) Visual encounter surveys, p. 84-92. In:
Heyer WR, Donnely MA, Roy WM, Hayek LC, Foster MS (Eds) Measuring and Monitoring
Biological Diversity: Standard Methods for Amphibians. Washington, DC, Smithsonian
Institution Press, 384p.) at nighttime (19:00 to 23:00 h) using headlamps. We conducted VES
within each plot up to five meters distance from the water and along the entire river
channel (Fig. 4), with a sampling effort of two
hours in each plot. The total sampling effort at each river was 40 hours of nocturnal
active search. Each individual was identified to species and then released at the same
point it was originally captured. To evaluate the ordination of plots regarding
amphibian species composition and abundance pooled for both rivers, we used a two-axis
Non-Metric Multidimensional Scaling, NMDS, on a matrix of Bray-Curtis dissimilarity
distances between plots (Legendre & Legendre
1998Legendre P, Legendre L (1998) Numerical Ecology. Amsterdam, Elsevier,
853p.). The two NMDS axes were used as dependent variables in a factorial
multivariate analysis of variance (MANOVA) using Wilks' λ to test if community
composition represented by the two NMDS axes differs with respect to the rivers,
riparian forest cover (lesser or greater), or an interaction of both (Báez et al. 2012Báez S, Collins SL, Pockman WT, Johnson JE, Small EE (2012) Effects of
experimental rainfall manipulations on Chihuahuan Desert grassland and shrubland
plant communities. Oecologia 172(4): 1117-1127. doi:
10.1007/s00442-012-2552-0
https://doi.org/10.1007/s00442-012-2552-...
). We conducted all analyzes in the R
environment (version 2.13.0, R Development Core Team 2011).

A river plot showing the total sampled area for anurans, including margins (gray) and river channel (white).
RESULTS
We recorded a total of 133 individuals from 22 anuran species at both rivers. The most abundant species were Cycloramphus brasiliensis (Steindachner, 1864) (33.8% of all individuals), Rhinella ornata (Spix, 1824) (16.5%), and Leptodactylus latrans (Steffen, 1815) (12.8%) (Table I). In the Manoel Alexandre River, 70 individuals were recorded from 14 frog species, ten species (N = 39) in plots with higher forest cover, six (60%) only in these portions of the river. In plots with lower forest cover, eight species were recorded (N = 31), four (50%) only in these portions of the river. The most abundant species in the Manoel Alexandre River was C. brasiliensis, 48.7% in greater forest cover plots, 32.2% in lesser forest cover plots. In the Guapiaçu River, 63 individuals were recorded from 15 frog species, eight species (N = 31) in plots with greater forest cover, four (50%) exclusively in these portions of the river. In plots with lesser forest cover, 11 species were recorded (N = 32), seven of them (63.6%) exclusively in these portions of the river. The most abundant species in greater cover plots of the Guapiaçu River was also C. brasiliensis (48.4%), but in lesser cover plots L. latrans was the most abundant (37.5%).
The ordination of amphibian species composition and abundance (Stress = 0.124) showed that most plots with comparatively greater forest cover clustered together (Fig. 5). Conversely, the plots with lesser forest cover were more scattered in the ordination biplot. The difference in species composition was nearly significant between rivers (MANOVA: Wilks' λ = 0.871, F1,36 = 2.589, p = 0.089), but differences were significant between plots with different forest cover (Wilks' λ = 0.619, F1,36 = 10.764, p < 0.001), and for the interaction between rivers and forest cover (Wilks' λ = 0.821, F1,36 = 3.811; p = 0.032).

Ordination of sampling plots along the axes of Non-metric Multidimensional Scaling (NMDS) of anuran species composition and abundances in two large rivers in the municipality of Cachoeiras de Macacu, state of Rio de Janeiro, Brazil. Black circles are plots with greater riparian forest cover, and white circles represent plots with lesser riparian forest cover.
DISCUSSION
The results indicate that riparian habitats are extremely important for amphibian
conservation since they maintain high species richness and abundance of frogs in spite
of the relatively small area occupied by riparian forests. These specific environments
may also provide more realistic assessments of extinction threats faced by
stream-dwelling amphibians in the Atlantic Forest and other biomes (Almeida-Gomes et al. 2014Almeida-Gomes M, Siqueira CC, Borges-Júnior VNT, Vrcibradic D, Fusinatto
LA, Rocha CFD (2014b) Herpetofauna of the Reserva Ecológica de Guapiaçu (REGUA) and
its surrounding areas, in the state of Rio de Janeiro, Brazil. Biota Neotropica
14(3): 1-15. doi: 10.1590/1676-0603007813
https://doi.org/10.1590/1676-0603007813...
a). In a 10-year survey in
the same region,(Almeida-Gomes et al. 2014Almeida-Gomes M, Siqueira CC, Borges-Júnior VNT, Vrcibradic D, Fusinatto
LA, Rocha CFD (2014b) Herpetofauna of the Reserva Ecológica de Guapiaçu (REGUA) and
its surrounding areas, in the state of Rio de Janeiro, Brazil. Biota Neotropica
14(3): 1-15. doi: 10.1590/1676-0603007813
https://doi.org/10.1590/1676-0603007813...
b), 73
amphibian species were registered, 10 of which are endemic to the state of Rio de
Janeiro. The present study was comparatively shorter, yet it found one third of the
species recorded in the region, including half of the endemic species recorded in that
previous study.
Our results also demonstrate that a reduction in riparian forest cover has an effect on
anuran species composition in both rivers. Riparian habitats can be used by amphibians
for foraging, dispersal and reproduction, and the habitat size required to preserve some
species is up to hundreds of meters (Semlitsch &
Bodie 2003Semlitsch RD, Bodie JR (2003) Biological criteria for buffer zones
around wetlands and riparian habitats for amphibians and reptiles. Conservation
Biology17(5): 1219-1228. doi: 10.1046/j.1523-1739.2003.02177.x
https://doi.org/10.1046/j.1523-1739.2003...
, Ficetola et al. 2009Ficetola GF, Padoa-Schioppa E, Bernardi F (2009) Influence of landscape
elements in riparian buffers on the conservation of semiaquatic amphibians.
Conservation Biology23(1): 114-123. doi:
10.1111/j.1523-1739.2008.01081.x
https://doi.org/10.1111/j.1523-1739.2008...
). The
removal of riparian vegetation can result in marked physical and structural changes in
the river and on its margins (Gomi et al. 2006Gomi T, Sidle RC, Noguchi S, Negishi JN, Nik AR, Sasaki S (2006)
Sediment and wood accumulations in humid tropical headwater streams: Effects of
logging and riparian buffers. Forest Ecology and Management 224(1-2): 166-175. doi:
10.1016/j.foreco.2005.12.016
https://doi.org/10.1016/j.foreco.2005.12...
),
and these changes generally cause a shift in amphibian species composition (Olson et al. 2007Olson DH, Anderson PD, Frissel CA, Welsh Jr HH, Bradford DF (2007)
Biodiversity management approaches for stream-riparian areas: Perspectives for
Pacific Northwest headwater forests, microclimates, and amphibians. Forest Ecology
and Management246: 81-107. doi:10.1016/j.foreco.2007.03.053
https://doi.org/10.1016/j.foreco.2007.03...
, Rodríguez-Mendoza & Pineda 2010Rodríguez-Mendoza C, Pineda E (2010) Importance of riparian remnants for
frog species diversity in a highly fragmented rainforest. Biology Letters 6: 781-784.
doi: 10.1098/rsbl.2010.0334
https://doi.org/10.1098/rsbl.2010.0334...
). The most abundant species in both
rivers studied, C. brasiliensis, is a stream-dwelling amphibian typical
of forested areas (Maia-Carneiro et al. 2012Maia-Carneiro T, Dorigo TA, Almeida-Gomes M, Van Sluys M, Rocha CFD
(2012) Feedings habitats, microhabitat use, and daily activity of Cycloramphus
brasiliensis (Anura: Cycloramphidae) from the Atlantic Rainforest, Brazil. Zoologia
29(3): 277-279. doi: 10.1590/S1984-46702012000300007
https://doi.org/10.1590/S1984-4670201200...
),
that is classified as Near Threatened (IUCN
2012IUCN (2012) Red list of Threatened Species. Version 2012.2. Available
online at: www.iucnredlist.org [Accessed: 30 January 2013]
www.iucnredlist.org...
). It was comparatively more abundant in the portions of the rivers that have
greater forest cover. On the other hand, some species that are typical of open areas
(natural or anthropogenic), for instance Scinax alter (Lutz, 1973),
were found only in the plots with lesser riparian vegetation. The riparian vegetation in
portions of the rivers outside the REGUA forest was mostly surrounded by pastures, which
allows the occupation of these areas by frog species adapted to disturbed habitats, such
as some hylids and leptodactylids (Almeida-Gomes et al.
2008Almeida-Gomes M, Vrcibradic D, Siqueira CC, Kiefer MC, Klaion T,
Almeida-Santos P, Nascimento D, Ariani CV, Borges-Júnior VNT, Freitas-Filho RF, Van
Sluys M, Rocha CFD (2008) Herpetofauna of an Atlantic Rainforest area (Morro São
João) in Rio de Janeiro State, Brazil. Anais da Academia Brasileira de Ciências
80(2): 291-300., 2010Almeida-Gomes M, Almeida-Santos M, Goyannes-Araújo P, Borges-Júnior VNT,
Vrcibradic D, Siqueira CC, Ariani CV, Dias AS, Souza VV, Pinto RR, Van Sluys M, Rocha
CFD (2010) Anurofauna of an Atlantic Rainforest fragment and its surroundings in
Northern Rio de Janeiro State, Brazil. Brazilian Journal of Biology 70(3):
871-877.).
The value of riparian forests for the conservation of biodiversity and ecosystem
services has already been recognized (Ricketts
2004Ricketts TH (2004) Tropical forest fragments enhance pollinator activity
in nearby coffee crops. Conservation Biology18(5): 1262-1271. doi:
10.1111/j.1523-1739.2004.00227.x
https://doi.org/10.1111/j.1523-1739.2004...
, Lees & Peres 2008Lees AC, Peres CA (2008) Conservation value of remnant riparian forest
corridors of varying quality for amazonian birds and mammals. Conservation Biology22:
439-449. doi: 10.1111/j.1523-1739.2007.00870.x
https://doi.org/10.1111/j.1523-1739.2007...
, Mendenhall et al. 2014Mendenhall CD, Frishkoff LO, Santos-Barrera G, Pacheco JS, Mesfun E,
Quijano FM, Ehrlich PR, Ceballos G, Daily GC, Pringle RM (2014) Countryside
biogeography of Neotropical reptiles and amphibians. Ecology 95: 856-870. doi:
10.1890/12-2017.1
https://doi.org/10.1890/12-2017.1...
), yet few studies have
ascertained the effect riparian forest removal on local communities. This study
demonstrates that riparian forests maintain a diverse community of amphibians that is
drastically different from the community next to riparian pastures. Species that are
more adapted to disturbed areas can benefit from forest reduction, which may cause the
changes in species composition observed in our analysis. Our results exemplify the
potential consequences of the proposed changes' the Brazilian forest code: population
reductions and local extinctions of frog species that depend on these riparian
environments.
ACKNOWLEDGMENTS
Mauricio Almeida-Gomes received Post-Doctoral scholarship from Programa Nacional de Pós-Doutorado (PNPD-CAPES). This study was supported by research grants from CNPq (processes 304791/2010-5 and 472287/2012-5) and FAPERJ through Cientistas do Nosso Estado Program (process E-26/102.404.2009 and E-26/102.765.2012) to C. F. D. Rocha and from CNPq (processes 307961/2011-7) and FAPERJ through Cientistas do Nosso Estado Program (process E-26/102.765/2012) to M.V. Vieira. We thank Nicholas J. Locke of the Reserva Ecológica de Guapiaçu (REGUA) for logistical support during our fieldwork in that area and all colleagues who helped us with data collection.
- Almeida-Gomes M, Rocha CFD (2014) Landscape connectivity may explain anuran species distribution in an Atlantic forest fragmented area. Landscape Ecology 29(1): 29-40. doi: 10.1007/s10980-013-9898-5
» https://doi.org/10.1007/s10980-013-9898-5 - Almeida-Gomes M, Vrcibradic D, Siqueira CC, Kiefer MC, Klaion T, Almeida-Santos P, Nascimento D, Ariani CV, Borges-Júnior VNT, Freitas-Filho RF, Van Sluys M, Rocha CFD (2008) Herpetofauna of an Atlantic Rainforest area (Morro São João) in Rio de Janeiro State, Brazil. Anais da Academia Brasileira de Ciências 80(2): 291-300.
- Almeida-Gomes M, Almeida-Santos M, Goyannes-Araújo P, Borges-Júnior VNT, Vrcibradic D, Siqueira CC, Ariani CV, Dias AS, Souza VV, Pinto RR, Van Sluys M, Rocha CFD (2010) Anurofauna of an Atlantic Rainforest fragment and its surroundings in Northern Rio de Janeiro State, Brazil. Brazilian Journal of Biology 70(3): 871-877.
- Almeida-Gomes M, Lorini ML, Rocha CFD, Vieira MV (2014a) Underestimation of extinction threat to stream-dwelling amphibians due to lack of consideration of narrow area of occupancy. Conservation Biology 28(2): 616-619. doi: 10.1111/cobi.12196
- Almeida-Gomes M, Siqueira CC, Borges-Júnior VNT, Vrcibradic D, Fusinatto LA, Rocha CFD (2014b) Herpetofauna of the Reserva Ecológica de Guapiaçu (REGUA) and its surrounding areas, in the state of Rio de Janeiro, Brazil. Biota Neotropica 14(3): 1-15. doi: 10.1590/1676-0603007813
» https://doi.org/10.1590/1676-0603007813 - Báez S, Collins SL, Pockman WT, Johnson JE, Small EE (2012) Effects of experimental rainfall manipulations on Chihuahuan Desert grassland and shrubland plant communities. Oecologia 172(4): 1117-1127. doi: 10.1007/s00442-012-2552-0
» https://doi.org/10.1007/s00442-012-2552-0 - Becker CG, Fonseca CR, Haddad CFB, Batista RF, Prado PI (2007) Habitat split and the global decline of amphibians. Science 318(5857): 1775-1777. doi: 10.1126/science.1149374
» https://doi.org/10.1126/science.1149374 - Bell KE, Donnelly MA (2006) Influence of forest fragmentation on community structure of frogs and lizards in northeastern Costa Rica. Conservation Biology20(6): 1750-1760. doi: 10.1111/j.1523-1739.2006.00522.x
» https://doi.org/10.1111/j.1523-1739.2006.00522.x - Crump ML, Scott Jr NJ (1994) Visual encounter surveys, p. 84-92. In: Heyer WR, Donnely MA, Roy WM, Hayek LC, Foster MS (Eds) Measuring and Monitoring Biological Diversity: Standard Methods for Amphibians. Washington, DC, Smithsonian Institution Press, 384p.
- Ficetola GF, Padoa-Schioppa E, Bernardi F (2009) Influence of landscape elements in riparian buffers on the conservation of semiaquatic amphibians. Conservation Biology23(1): 114-123. doi: 10.1111/j.1523-1739.2008.01081.x
» https://doi.org/10.1111/j.1523-1739.2008.01081.x - Gomi T, Sidle RC, Noguchi S, Negishi JN, Nik AR, Sasaki S (2006) Sediment and wood accumulations in humid tropical headwater streams: Effects of logging and riparian buffers. Forest Ecology and Management 224(1-2): 166-175. doi: 10.1016/j.foreco.2005.12.016
» https://doi.org/10.1016/j.foreco.2005.12.016 - Haddad CFB, Prado CPA (2005) Reproductive modes in frogs and their unexpected diversity in the Atlantic Forest of Brazil. BioScience 55(3): 207-217.
- Hof C, Araújo MB, Jetz W, Rahbek C (2011) Additive threats from pathogens, climate and land-use change for global amphibian diversity. Nature 480: 516-519. doi: 10.1038/nature10650
» https://doi.org/10.1038/nature10650 - IUCN (2012) Red list of Threatened Species. Version 2012.2. Available online at: www.iucnredlist.org [Accessed: 30 January 2013]
» www.iucnredlist.org - Keller A, Rödel M-O, Linsenmair E, Grafe TU (2009) The importance of environmental heterogeneity for species diversity and assemblage structure in Bornean stream frogs. Journal of Animal Ecology 78: 305-314. doi: 10.1111/j.1365-2656.2008.01457.x
» https://doi.org/10.1111/j.1365-2656.2008.01457.x - Lees AC, Peres CA (2008) Conservation value of remnant riparian forest corridors of varying quality for amazonian birds and mammals. Conservation Biology22: 439-449. doi: 10.1111/j.1523-1739.2007.00870.x
» https://doi.org/10.1111/j.1523-1739.2007.00870.x - Legendre P, Legendre L (1998) Numerical Ecology. Amsterdam, Elsevier, 853p.
- Maia-Carneiro T, Dorigo TA, Almeida-Gomes M, Van Sluys M, Rocha CFD (2012) Feedings habitats, microhabitat use, and daily activity of Cycloramphus brasiliensis (Anura: Cycloramphidae) from the Atlantic Rainforest, Brazil. Zoologia 29(3): 277-279. doi: 10.1590/S1984-46702012000300007
» https://doi.org/10.1590/S1984-46702012000300007 - Mendenhall CD, Frishkoff LO, Santos-Barrera G, Pacheco JS, Mesfun E, Quijano FM, Ehrlich PR, Ceballos G, Daily GC, Pringle RM (2014) Countryside biogeography of Neotropical reptiles and amphibians. Ecology 95: 856-870. doi: 10.1890/12-2017.1
» https://doi.org/10.1890/12-2017.1 - Metzger JP, Lewinsohn TM, Joly CA, Verdade LM, Martinelli LA, Rodrigues RR (2010) Brazilian Law: full speed in reverse? Science329: 276-277.
- Mittermeier RA, Gil RP, Hoffman M, Pilgrim J, Brooks T, Mittermeier CG, Lamoreux J, Fonseca GAB (2005) Hotspots revisited: earth's biologically richest and most endangered terrestrial ecoregions. Washington, DC, Conservation International, 392p.
- Olson DH, Anderson PD, Frissel CA, Welsh Jr HH, Bradford DF (2007) Biodiversity management approaches for stream-riparian areas: Perspectives for Pacific Northwest headwater forests, microclimates, and amphibians. Forest Ecology and Management246: 81-107. doi:10.1016/j.foreco.2007.03.053
» https://doi.org/10.1016/j.foreco.2007.03.053 - Parris KM, McCarthy MA (1999) What influences the structure of frog assemblages at forest streams? Australian Journal of Ecology 24(5): 495-502. doi: 10.1046/j.1442-9993.1999.00989.x
» https://doi.org/10.1046/j.1442-9993.1999.00989.x - Ribeiro MC, Metzger JP, Martensen AC, Ponzoni FJ, Hirota MM (2009) The Brazilian Atlantic Forest: how much is left, and how is the remaining forest distributed? Implications for conservation. Biological Conservation 142: 1144-1156. doi:10.1016/j.biocon.2009.02.021
» https://doi.org/10.1016/j.biocon.2009.02.021 - Richards-Zawacki CL (2009) Effects of slope and riparian habitat connectivity on gene flow in an endangered Panamanian frog, Atelopus varius. Diversity and Ditributions 15: 796-806. doi: 10.1111/j.1472-4642.2009.00582.x
» https://doi.org/10.1111/j.1472-4642.2009.00582.x - Ricketts TH (2004) Tropical forest fragments enhance pollinator activity in nearby coffee crops. Conservation Biology18(5): 1262-1271. doi: 10.1111/j.1523-1739.2004.00227.x
» https://doi.org/10.1111/j.1523-1739.2004.00227.x - Rodríguez-Mendoza C, Pineda E (2010) Importance of riparian remnants for frog species diversity in a highly fragmented rainforest. Biology Letters 6: 781-784. doi: 10.1098/rsbl.2010.0334
» https://doi.org/10.1098/rsbl.2010.0334 - Semlitsch RD, Bodie JR (2003) Biological criteria for buffer zones around wetlands and riparian habitats for amphibians and reptiles. Conservation Biology17(5): 1219-1228. doi: 10.1046/j.1523-1739.2003.02177.x
» https://doi.org/10.1046/j.1523-1739.2003.02177.x - Todd BD, Luhring TM, Rothermel BB, Gibbons W (2009) Effects of forest removal on amphibian migrations: implications for habitat and landscape connectivity. Journal of Applied Ecology 46: 554-561. doi: 10.1111/j.1365-2664.2009.01645.x
» https://doi.org/10.1111/j.1365-2664.2009.01645.x - Zimmerman BL, Bierregaard RO (1986) Relevance of the equilibrium theory of island biogeography and species-area relations to conservation with a case from Amazonia. Journal of Biogeography 13(2): 133-143. doi: 10.2307/2844988
» https://doi.org/10.2307/2844988
Publication Dates
-
Publication in this collection
Jan-Feb 2015
History
-
Received
30 May 2014 -
Reviewed
10 Aug 2014 -
Accepted
06 Dec 2014