ABSTRACT
Fish larvae and adults are morphologically distinct from each other and have different ecological requirements. Research on the dynamics of ichthyoplankton provides valuable information about the early stages of the life cycle of fish. This study analyzed the temporal and spatial distribution of the larval stages of three migratory species - Salminus brasiliensis (Cuvier, 1816), Prochilodus lineatus (Valenciennes, 1836) and Pseudoplatystoma corruscans (Agassiz, 1829) - to identify possible spawning areas in the last lotic remnant of the upper Paraná River, in the Brazilian territory. A total of 26 points were sampled monthly from October 2003 to November 2004. The PERMANOVA detected significant differences between the months only for S. brasiliensis (Pseudo-F = 3.88, p = 0.0021) and P. corruscans (Pseudo-F = 3.35, p = 0.004). Among the environmental variables, temperature was the most important and the Mantel test detected a significant correlation between temperature and the densities, only for P. lineatus (r = 0.09, p = 0.03). The species in our data chose for spawning tributaries that are not dammed, of which most are outside conservation areas. Therefore, recruitment, maintenance and possible actions aiming to recover the stocks of these species are dependent on the conservation of tributaries and floodplain lakes in the region, in addition to monitoring to prevent fishing during the reproductive period.
KEY WORDS
Ichthyoplankton; Prochilodus lineatus; Pseudoplatystoma corruscans; Salminus brasiliensis; Upper Paraná River
Fish colonize all types of habitats, which are subjected to various environmental conditions. They have various shapes, sizes and life cycles (Wootton 1991Wootton RJ (1991) Ecology of teleost fishes. London, Champman and Hall, 404p.). The life strategy of these organisms is associated with favorable conditions to the early development of eggs and larvae, highlighting sites and seasons with higher availability of food and shelter (Nakatani et al. 2001Nakatani K, Agostinho AA, Baumgartner G, Bialetzki A, Sanches PV, Makrakis MC, Pavanelli CS (2001) Ovos e larvas de peixe de água doce: Desenvolvimento e manual de identificação. Maringá, Eduem, 378p.). Nevertheless, larvae and adults are morphologically and behaviorally distinct, and thus have different ecological requirements, such as habitat and food (Sanches et al. 2001Sanches PV, Baumgartner G, Bialetzki A, Suiberto MR, Gomes FDC, Nakatani K, Barbosa NDC (2001) Caracterização do desenvolvimento inicial de Leporinus friderici (Osteichthyes; Anostomidae) da bacia do rio Paraná. Acta Scientiarum 23: 383-389. doi: 10.4025/actascibiolsci.v23i0.2693
https://doi.org/10.4025/actascibiolsci.v...
, Nakatani et al. 2001Nakatani K, Agostinho AA, Baumgartner G, Bialetzki A, Sanches PV, Makrakis MC, Pavanelli CS (2001) Ovos e larvas de peixe de água doce: Desenvolvimento e manual de identificação. Maringá, Eduem, 378p.).
Reproduction is one of the most important aspects of the biology of a species, since the maintenance of viable populations depends on its success(Suzuki & Agostinho 1997Suzuki HI, Agostinho AA (1997) Reprodução de peixes do reservatório de Segredo, p. 163-180. In: Agostinho AA, Gomes LC (Eds.). Reservatório de Segredo: bases ecológicas para o manejo. Maringá, Eduem, 387p.). Reproductive failure in consecutive years, mainly caused by changes in hydrological cycles (flow regulation and change in the timing of high flows) (Vazzoler 1996Vazzoler AEAM (1996) Biologia da reprodução de peixes teleósteos: teoria e prática. Maringá, Eduem, 196p., Agostinho et al. 2003 Agostinho AA , Gomes LC, Suzuki HI, Júlio Jr HF (2003) Migratory fish from the upper Paraná River basin, Brazil, p. 19-99. In: Carolsfeld J, Harvey B, Ross C, Baer A (Eds.). Migratory fishes of South America: biology, social importance and conservation status. Victoria, World Fisheries Trust, The World Bank and the International Development Research Center, 215p., 2004aAgostinho AA, Thomaz SM, Gomes LC (2004a) Threats for biodiversity in the floodplain of upper Paraná River: effects of hydrological regulation by dams. Ecohydrology and Hydrobiology 4(3): 267-280.), may result in depletion or even extinction of natural populations (Welcomme 1979Welcomme RL (1979) Fisheries ecology of floodplain rivers. London, Longman, 317p., Gomes & Agostinho 1997Gomes LC, Agostinho AA (1997) Influence of the flooding regime on the nutritional state and juvenile recruitment of the curimba, Prochilodus scrofa, Steindachner in upper Paraná River, Brazil. Fisheries Management and Ecology 4: 263-274., Suzuki & Agostinho 1997Suzuki HI, Agostinho AA (1997) Reprodução de peixes do reservatório de Segredo, p. 163-180. In: Agostinho AA, Gomes LC (Eds.). Reservatório de Segredo: bases ecológicas para o manejo. Maringá, Eduem, 387p., Ali & Wootton 1999Ali M, Wootton RJ (1999) Effect of variable food on reproductive performance breeding female three-spined sticklebacks. Journal of Fish Biology 55: 1040-1053. doi: 10.1111/j.1095-8649.1999.tb00739.x
https://doi.org/10.1111/j.1095-8649.1999...
, Agostinho et al. 2004bAgostinho AA, Gomes LC, Veríssimo S, Okada EK (2004b) Flood regime, dam regulation and fish in the Upper Paraná River: effects on assemblage attributes, reproduction and recruitment. Reviews in Fish Biology and Fisheries 14: 11-19.).
The Paraná River is intensely impacted by dams. A conservative count that takes into consideration only dams that are 10 m high above the water level (Agostinho et al. 1995Agostinho AA, Vazzoler AEAM, Thomaz SM (1995) The high River Paraná basin: limnological and ichthyological aspects, p. 59-103. In: Tundisi JG, Bicudo CEM, Matsumura-Tundisi T (Eds.). Limnology in Brazil. Rio de Janeiro, ABC/SBL, 376p., 2007Agostinho AA, Gomes LC, Pelicice FM (2007) Ecologia e manejo de recursos pesqueiros em reservatórios do Brasil. Maringá, Eduem, 501p.) totals 130 dams, transforming the river course into a cascade of dams.
The Sérgio Motta Dam, built in 1998 (known as Porto Primavera), has deeply affected the dynamics of the ichthyoplankton in the upper Paraná River floodplain (Sanches et al. 2006Sanches PV, Nakatani K, Bialetzki A, Baumgartner G, Gomes LC, Luiz EA (2006) Flow regulation dams affecting ichthyoplankton: the case of Porto Primavera dam; Paraná River; Brazil. River Research App 22: 555-565. doi: 10.1002/rra.922
https://doi.org/10.1002/rra.922...
), located immediately downstream from it. This study was conducted in this area (above the Porto Primavera Dam and the Itaipu Reservoir), which represents the lotic remnant of the Paraná River inside the Brazilian territory. Despite the fact that three conservation units (Ilha Grande National Park, State Park of the Várzeas of the Ivinhema River, and Environmental Protection Area of the Islands and Várzeas of the Paraná River) are present in the area,the local fish fauna is especially impacted by the dams (Agostinho et al. 2000Agostinho AA, Thomaz SM, Minte-Vera CV, Winemiller KO (2000) Biodiversity in the high Paraná River floodplain, p. 89-118. In: Gopal B, Junk WDJ, Davis JA (Eds.). Biodiversity in wetlands: assessment, function and conservation. Leiden, Backhuys Publishers, 310p., Sanches et al. 2006Sanches PV, Nakatani K, Bialetzki A, Baumgartner G, Gomes LC, Luiz EA (2006) Flow regulation dams affecting ichthyoplankton: the case of Porto Primavera dam; Paraná River; Brazil. River Research App 22: 555-565. doi: 10.1002/rra.922
https://doi.org/10.1002/rra.922...
), which directly impact water quality, spawning, and recruitment of populations (Gomes & Agostinho 1997Gomes LC, Agostinho AA (1997) Influence of the flooding regime on the nutritional state and juvenile recruitment of the curimba, Prochilodus scrofa, Steindachner in upper Paraná River, Brazil. Fisheries Management and Ecology 4: 263-274., Petry et al. 2003Petry AC, Agostinho AA, Gomes LC (2003) Fish assemblages of tropical floodplain lagoons: exploring the role of connectivity in a dry year. Neotropical Ichthyology 1(2): 111-119. doi: 10.1590/S1679-62252003000200005
https://doi.org/10.1590/S1679-6225200300...
, Agostinho et al. 2004bAgostinho AA, Gomes LC, Veríssimo S, Okada EK (2004b) Flood regime, dam regulation and fish in the Upper Paraná River: effects on assemblage attributes, reproduction and recruitment. Reviews in Fish Biology and Fisheries 14: 11-19.).
Species of great commercial value in professional fishing, such as Salminus brasiliensis (Cuvier, 1816), Prochilodus lineatus (Valenciennes, 1836) and Pseudoplatystoma corruscans (Agassiz, 1829), are among the most vulnerable to habitat changes in the Paraná River basin (Agostinho et al. 1997Agostinho AA, Júlio Jr HF, Gomes LC, Bini LM, Agostinho CS (1997) Composição, abundância e distribuição espaço-temporal da ictiofauna, p. 179-208. In: Vazzoler AEAM, Agostinho AA, Hahn NS (Eds.). A planície de inundação do alto rio Paraná: aspectos físicos; biológicos e socioeconômicos. Maringá, Eduem, 460p.). These species are migratory. Consequently, restrictions to water flow and changes in the seasonality of the hydrological cycle of the river, imposed by the dams, impair their reproduction, which depends on seasonal variations in river tide (Agostinho et al. 2003 Agostinho AA , Gomes LC, Suzuki HI, Júlio Jr HF (2003) Migratory fish from the upper Paraná River basin, Brazil, p. 19-99. In: Carolsfeld J, Harvey B, Ross C, Baer A (Eds.). Migratory fishes of South America: biology, social importance and conservation status. Victoria, World Fisheries Trust, The World Bank and the International Development Research Center, 215p.). According to the Red List of Threatened Fauna of the state of Paraná (Abilhoa & Duboc 2004Abilhoa V, Duboc LF (2004) Peixes, p. 581-682. In: Mikichs B, Bérnils RS (Eds.). Livro vermelho da fauna ameaçada no Estado do Paraná. Curitiba, Governo do Estado do Paraná, SEMA, IAP, 763p.), S. brasiliensis and P. corruscans are no longer abundant in the Paraná River basin. However, these species are still found in this stretch of the river where there are several tributaries, many of which are outside the conservation units. These tributaries are still not dammed, and can be used by migratory species as alternative migration routes, particularly for reproduction (Sanches et al. 2006Sanches PV, Nakatani K, Bialetzki A, Baumgartner G, Gomes LC, Luiz EA (2006) Flow regulation dams affecting ichthyoplankton: the case of Porto Primavera dam; Paraná River; Brazil. River Research App 22: 555-565. doi: 10.1002/rra.922
https://doi.org/10.1002/rra.922...
).
The identification of fish eggs and larvae, combined with data on their distribution provide considerable evidence about the spawning season, sites for reproduction, and natural breeding areas (Nakatani et al. 1997Nakatani K, Baumgartner G, Cavicchioli M (1997) Ecologia de ovos e larvas de peixes, p. 281-306. In: Vazzoler AEAM, Agostinho AA, Hahn NS (Eds.). A planície de inundação do alto rio Paraná: aspectos físicos; biológicos e socioeconômicos. Maringá, Eduem, 460p., Nakatani et al. 2001Nakatani K, Agostinho AA, Baumgartner G, Bialetzki A, Sanches PV, Makrakis MC, Pavanelli CS (2001) Ovos e larvas de peixe de água doce: Desenvolvimento e manual de identificação. Maringá, Eduem, 378p.). With this information, we can implement guidance and protection measures in these areas, preventing the capture of young individuals to maximize recruitment and maintenance of fish stocks (Baumgartner et al. 2004Baumgartner G, Nakatani K, Gomes LC, Bialetzki A, Sanches PV, Makrakis MC (2004) Identification of spawning sites and natural nurseries of fishes in the upper Paraná River, Brazil. Environmental Biology of Fishes 71: 115-125.).
This study aimed to evaluate the temporal and spatial distribution (indication of preferred areas and season for spawning) of larvae of three migratory species (S. brasiliensis, P. lineatus and P. corruscans) in the region of the upper Paraná River, located in the lotic remnant of the Paraná River in the Brazilian territory. We intended to answer the following questions: i) Do larval abundances vary in the temporal and spatial scales considered (temporal and spatial)?; ii) Does the abundance of larval stages (yolk-sac larvae, preflexion and flexion) differ between sampled stretches and the stretches chosen for spawning; and iii) is there any relationship between larval density and selected environmental variables (pH, dissolved oxygen, electric conductivity, water temperature, and river water level)?
MATERIAL AND METHODS
The study area is located in the Paraná River, in a 230 km stretch that divides the states of Paraná and Mato Grosso do Sul. It is delimited upstream by the Porto Primavera Dam (Sérgio Motta Hydroelectric Power Plant), and downstream by the Itaipu Reservoir. In this stretch there is still a river-floodplain system, which currently encompasses three important conservation units (Ilha Grande National Park, Ivinhema River State Park, and Environmental Protection Area of the Islands and 'Várzeas' of the Paraná River).
The area is mainly represented by the Ilha Grande archipelago, which divides the Paraná River into two channels, and is composed of a fluvial complex with about 157 islands of variable sizes. The presence of these islands transforms the river into an anastomosed channel, with backwater areas and secondary channels (Gogola et al. 2010Gogola TM, González LMA, Daga VS, Silva PRL, Sanches PV, Gubiani EA, Baumgartner G, Delariva RL (2010) Spatial and temporal distribution patterns of ichthyoplankton in a region affected by water regulation by dams. Neotropical Ichthyology 8: 341-349. doi: 10.1590/S1679-62252010000200013
https://doi.org/10.1590/S1679-6225201000...
). These characteristics promote great habitat heterogeneity, which in turn is associated with great biological diversity of terrestrial and aquatic organisms (Agostinho et al. 2000Agostinho AA, Thomaz SM, Minte-Vera CV, Winemiller KO (2000) Biodiversity in the high Paraná River floodplain, p. 89-118. In: Gopal B, Junk WDJ, Davis JA (Eds.). Biodiversity in wetlands: assessment, function and conservation. Leiden, Backhuys Publishers, 310p., 2004bAgostinho AA, Gomes LC, Veríssimo S, Okada EK (2004b) Flood regime, dam regulation and fish in the Upper Paraná River: effects on assemblage attributes, reproduction and recruitment. Reviews in Fish Biology and Fisheries 14: 11-19.).
Although under the influence of the dams, the stretch chosen for this research has several tributaries, such as the Piquiri and Ivaí rivers, located on the left (East) bank of the Paraná River (Paraná State), and Amambaí, Iguatemi and Ivinhema, on the right (West) bank (Mato Grosso do Sul State), all free of dams, with their flow characteristics exclusively dependent on the hydrological cycle (local rains). In addition to these tributaries, the region still has several floodplain lakes, with constant or intermittent connection with the Paraná River or secondary channels (Gogola et al. 2010Gogola TM, González LMA, Daga VS, Silva PRL, Sanches PV, Gubiani EA, Baumgartner G, Delariva RL (2010) Spatial and temporal distribution patterns of ichthyoplankton in a region affected by water regulation by dams. Neotropical Ichthyology 8: 341-349. doi: 10.1590/S1679-62252010000200013
https://doi.org/10.1590/S1679-6225201000...
).
The 26 sampling sites selected were distributed in the main channel of the Paraná River along the Ilha Grande National Park, tributaries (Ivinhema, Amambai, Piquiri and Iguatemi), marginal lakes (Saraiva Lake) and channels (Canal Ipoitã and Boca do Ipoitã), including the mouth of the major tributaries. For analysis, these sampling stations were categorized into three sections, according to the position of the main tributaries: i) Upper section, with sampling sites influenced by the rivers Ivinhema and Amambai (Boca do Ipoitã; Ipoitã Channel; Mouth of Ivinhema River; Channel of Paraná River; Bandeirantes Right Channel; Amambai; Bandeirantes Left Channel; Ilha Grande Pontal); ii) Middle section corresponding to sampling sites located in the main channel of the Paraná River (Triângulo; Porto Santo Antonio; Peruzzi; Paraná/Iguatemi; Alvarenga; Esmeralda; Três Coqueiros; São João), with only a few tributaries; iii) Lower section, with sampling sites influenced by the rivers Piquiri and Iguatemi (Iguatemi; Paraná/Saraiva; Saraiva Middle; Saraiva Channel; Ilha Grande Right Channel; Porto Cerâmica; Piquiri; Porto Terra Roxa; Ilha Grande Left Channel) (Fig. 1).

Location of the sampling stations (dark circles) 1: Bandeirantes Right Channel, 2: Amambai, 3: Triângulo, 4: Porto Santo Antonio, 5: Peruzzi, 6: Paraná/Iguatemi, 7: Iguatemi, 8: Paraná/Saraiva, 9: Saraiva Middle, 10: Saraiva Channel, 11: Ilha Grande Right Channel, 12: Bandeirantes Left Channel, 13: Ilha Grande Pontal, 14: Alvarenga, 15: Esmeralda, 16: Três Coqueiros, 17: São João, 18: Porto Luiz, 19 Porto Cerâmica, 20: Piquiri, 21: Porto Terra Roxa, 22: Ilha Grande Left Channel, 23: Paraná Channel, 24: Mouth of Ivinhema River, 25: Boca do Ipoitã, 26: Ipoitã Channel.
Samples were taken during the spawning season, from October 2003 to March 2004 (Vazzoler 1996Vazzoler AEAM (1996) Biologia da reprodução de peixes teleósteos: teoria e prática. Maringá, Eduem, 196p.). A cylindrical-conical plankton net (0.5 mm mesh size), with a flowmeter coupled to the mouth opening to obtain the volume of filtered water, was employed. In the sampling sites with lotic characteristics, the nets were submerged approximately 10 cm against the water current for 10 minutes. In the lentic sites, the depth and time used for sampling was the same as in lotic sampling, but the net was towed at a lower speed. We obtained one sample per collection point, which was fixed with formaldehyde solution (4%) buffered with calcium carbonate (CaCO3). During sampling we also measured the following environmental variables using digital portable equipments: pH, dissolved oxygen (mg.l-1), electric conductivity (µS.cm-1) and water temperature (°C). Fluviometric level data were supplied by Nupélia/Fluviometric Station of the Advanced Research Base in Porto Rico, Paraná State.
After sorting the larvae from the rest of the plankton, the larvae were identified according to Nakatani et al. (2001Nakatani K, Agostinho AA, Baumgartner G, Bialetzki A, Sanches PV, Makrakis MC, Pavanelli CS (2001) Ovos e larvas de peixe de água doce: Desenvolvimento e manual de identificação. Maringá, Eduem, 378p.) and classified into three categories according to Ahlstrom & Ball (1954Ahlstrom EH, Ball OP (1954) Description of eggs and larvae of jack mackerel (Trachurus symmetricus) and distribution and abundance of larvae in 1950 and 1951. Fishery Bulletin 56: 209-245.) and modified by Nakatani et al. (2001Nakatani K, Agostinho AA, Baumgartner G, Bialetzki A, Sanches PV, Makrakis MC, Pavanelli CS (2001) Ovos e larvas de peixe de água doce: Desenvolvimento e manual de identificação. Maringá, Eduem, 378p.): yolk-sac larval, preflexion and flexion stages. Larval abundance was standardized to 10 m3 of filtered water according to Tanaka (1973Tanaka S (1973) Stock assessment by means of ichthyoplankton surveys. FAO Fish Tech Paper 122: 33-51.), modified by Nakatani et al. (2001Nakatani K, Agostinho AA, Baumgartner G, Bialetzki A, Sanches PV, Makrakis MC, Pavanelli CS (2001) Ovos e larvas de peixe de água doce: Desenvolvimento e manual de identificação. Maringá, Eduem, 378p.). Specimens of P. corruscans, S. brasiliensis, and P. lineatus were deposited at Coleção Ictiológica da Universidade Estadual de Maringá, Paraná, Brazil (Nupélia vouchers numbers NUP 17332, NUP 1733, NUP 17330, respectively).
In order to test for significant differences between the means of larval abundance and considering the factors Month (Temporal variation: October, November, December, January, February and March) and Section (Spatial variation: Upper, Middle, and Lower) and their interaction, we used a distance-based analysis of variance, but the significance based on a permutation test, the PERMANOVA, employing the software PRIMER and PERMANOVA+. This software performs the permutational analysis of variance (Anderson 2001Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Austral Ecology 26: 32-46. doi: 10.1111/j.1442-9993.2001.01070.pp.x
https://doi.org/10.1111/j.1442-9993.2001...
, McArdle & Anderson 2001McArdle BH, Anderson MJ (2001) Fitting multivariate models to community data: a comment on distance based redundancy analysis. Ecology 82: 290-297. doi: 10.1890/0012-9658(2001)082[0290:FMMTCD]2.0.CO;2
https://doi.org/10.1890/0012-9658(2001)0...
) on the values of any distance measure, using permutation procedures to obtain the probability to reject the null hypothesis for the test (Pseudo - F). Whenever the PERMANOVA was significant, a pairwise test was applied a posteriori to identify which mean values of each level of the factor were different.
In this study, the matrix of original data (larval density) contained many zero values, which prevent calculating the similarity matrix (in the case using the Bray-Curtis similarity index). To minimize this problem, a constant value (1) was added to all the cases, thus allowing calculating the matrix. In this way, all empty cells were forced to have overall similarity (100%) (Clarke et al. 2006Clarke KR, Somerfield PJ, Chapman MG (2006) On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray-Curtis coefficient for denuded assemblages. Journal of Experimental Marine Biology and Ecology 330(1): 55-80. doi: 10.1016/j.jembe.2005.12.017
https://doi.org/10.1016/j.jembe.2005.12....
) and we sought for differences between the less similar samples, which presented greater abundance. For the evaluations of the possible spawning sites, only the graphs were inspected, given the low densities of larval stages (yolk-sac larvae, preflexion and flexion) when they were analyzed separately.
The association between the matrices of the environmental variables (one for each variable) and the matrices of larval density (one for each species) was evaluated through the Mantel tests. The metrics used to generate the matrices were: Bray-Curtis for the data of larval abundance (added the constant 1) and the Euclidean distance for the environmental variables. Since the sampling sites are distant from each other (most downstream and upstream most sites) they could represent very heterogeneous abiotic data, invalidating the use of the Mantel tests. To evaluate the homogeneity of the environmental variables among the sampling sites, a permutation test of multivariate homogeneity of group dispersions (Anderson 2006Anderson MJ (2006) Distance-based tests for homogeneity of multivariate dispersions. Biometrics 62: 245-253., Anderson et al. 2006Anderson MJ, Ellingsen KE, McArdle BH (2006) Multivariate dispersion as a measure of beta diversity. Ecology Letters 9(6): 683-693.) was carried out. This analysis was performed using the R software, package vegan (Oksanen et al. 2013Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O'Hara RB, Simpson GL, Henry M, Stevens H, Wagner H (2013) Vegan: Community Ecology Package. R package version 2.0-8. Available online at: Available online at: http://CRAN.R-project.org/package=vegan
[Accessed: 10/11/2014]
http://CRAN.R-project.org/package=vegan...
) by the functions betadisper using Euclidean distance to generate distance from centroids to each stretch followed by the permutest.betadisper function to test the mean distance from centroids among the stretches. In addition, scatter plots were built to explore possible patterns between the data of larval abundance and selected abiotic variables selected. The Mantel test was conducted using the PC-ORD version 4.01 (McCune & Mefford 1999McCune B, Mefford MJ (1999) PC-ORD. Multivariate analysis of ecological data. Gleneden Beach, MjM Software Design, Version 4.01.) and the graphs in the software 7.0 (StatSoft 2005Statsoft (2005) Electronic Statistic textbook. Available online at: Available online at: http://www.statsoft.com/textbook/stathome.html
[Accessed: 5/2/2011].
http://www.statsoft.com/textbook/stathom...
).
RESULTS
Variations in the water level of Paraná River
The floods in the studied stretch usually occur from October to March, coinciding with the greatest reproductive activity of fish, especially migratory species. However, higher values were observed at the river level in October, November, December and March. Lower values were verified (Fig. 2) in January, which corresponds to the peak of the reproductive activity.

Daily values of the level of Paraná River during the period from June 2003 to June 2004, recorded in the Field Station of the Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura (Nupélia) in Porto Rico, Paraná State.
With regard to environmental variables, during the study period the pH ranged from 5.9 to 8.0, electrical conductivity varied from 42.4 to 68.3 µS/cm, water temperature varied between 26.5 °C and 30.6°C and dissolved oxygen was between 2.2 and 9.2 mg/l (Fig. 3).

Values of the data of abiotic factors (pH, dissolved oxygen (mg.l-1), electric conductivity (µS.cm-1) and water temperature (°C), per sampling point, in the region of Ilha Grande National Park, in Paraná River.
Spatial and temporal variations in the abundance and identification of preferred section for spawning
During the study period, were captured a total of 169 larvae of long-distance migratory species. Among them, 53 were Salminus brasiliensis and its higher density was collected in section 20 (Piquiri), 40 larvae were Pseudoplatystoma corruscans and the highest density of this species was collected in section 24 (Mouth of Ivinhema River) and only 13 Prochilodus lineatus and its higher density was collected in section 11 (Ilha Grande Right Channel) (Table 1).
Relationship between the density of larvae Salminus brasiliensis, Pseudoplathystoma corruscans and Prochilodus lineattus and sampling stations.
These low values may be due to the low levels of the river during the reproductive period, mainly in January, 2004. High densities of S. brasiliensis were verified in October and November. In October, the highest mean value was registered in the Upper section, whereas in November, at the Lower section (Fig. 4). The mean values of larval density were generally low. However, they were lower from January to the end of the sampled period. The PERMANOVA detected significant differences between the months (Pseudo-F = 3.88, p = 0.0021). The mean values obtained in October and November were not different according to the pairwise test (p = 0.79), but they were different from the other months (p < 0.03).
Yolk-sac larvae (YL) were found in the three sections, with higher densities in the Lower stretch; preflexion larvae (PF) were only captured in the Middle and Upper stretches. Flexion larvae (FL) had the lowest densities, with no capture in the Lower section (Fig. 5). Thus, it can be stated that S. brasiliensis has reproduced in the vicinities of the sampling sites located in the Upper and Lower sections, due to the capture of larvae at the yolk-sac stage.

Spatial and temporal variation (4) in the mean density of Salminus brasiliensis and of development stages (5) (YL: yolk-sac, PF: preflexion, FL: flexion) in the region of Ilha Grande National Park, in Paraná River.
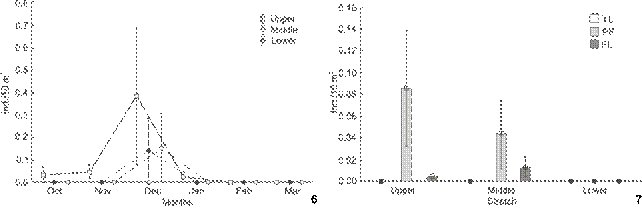
In the sampled stretches, higher captures of P. corruscans occurred in December (Fig. 6). However, the highest mean was registered in the Upper section sites. No record of this species was verified in February and March, which can be result from the low levels of the Paraná River in January 2005. The PERMANOVA also evidenced significant differences only between the months (Pseudo-F = 3.35, p = 0.004), and December differed from the others (pairwise test, p < 0.05). Pseudoplatys toma corruscans was not captured at yolk-sac larval stage. The stages of pre-flexion (PF) and flexion (FL) were only registered in the Upper and Middle sections (Fig. 7).

Spatial and temporal variation in the mean density of Pseudoplatystoma corruscans (6) and of development stages (7) (YL: yolk-sac, PF: preflexion, FL: flexion) in the region of Ilha Grande National Park, in Paraná River.
Prochilodus lineatus was captured in almost all studied months, but in low mean densities. The highest mean density of this fish was registered in the Lower stretch, followed by the Upper stretch, in October and November, respectively (Fig. 8). The PERMANOVA revealed no significant difference between the months and stretches (Pseudo-F < 0.75, p > 0.60). Yolk-sac larvae (YL) were found in the three sections of the stretch, with higher densities in the Upper and Middle ones. The pre-flexion stage larvae (PF) were captured only in the Middle and Lower sections, with the highest abundance in the latter section. Nevertheless, no larvae at flexion stage (FL) were captured (Fig. 9).

Spatial and temporal variation in the mean density of Prochilodus lineatus (8) and of development stages (9) (YL: yolk-sac, PF: preflexion, FL: flexion) in the region of Ilha Grande National Park, in Paraná River.
Relationship between larval abundance and environmental variables
The relationships between the densities of the selected migratory species and the environmental variables measured were not clear (Fig. 10). However, temperature was an important variable, mainly for S. brasiliensis, whose larvae were captured only when the temperature was below 28°C. This was shown by the lack of significance between the matrices (abundance and abiotic) obtained for this species (Mantel r = 0.08, p = 0.06). There was also no significant correlation between the matrices constructed for P. corruscans (r = -0.06, p = 0.09). However, for P. lineatus the correlation was significant (r = 0.09, p = 0.03), but the larval densities of this species were quite low. The distance to the centroid was 4.749 to the superior, 6.307 to the middle and 4.543 to the lower stretches and the mean distance comparing the stretches were 0.554 to the upper-middle, 0.912 to the superior-lower, 0.482 to the middle-lower comparisons. The test homogeneity of groups dispersions to environmental variables among the sampling sites are not significant (F = 0.3567, p = 0.713), validating the use of Mantel tests.

Relationship of environmental variables with densities of Salminus brasiliensis, Pseudoplatystoma corruscans and Prochilodus lineatus in the Upper Paraná River floodplain.
DISCUSSION
Studies performed in La Plata River basin have shown that the reproduction of migratory fish occurs from October to March (Baumgartner et al. 1997Baumgartner G, Nakatani K, Cavicchioli M, Baumgartner MST (1997) Some aspects of the ecology of fish larvae in the floodplain of the high Paraná River, Brazil. Revista Brasileira de Zoologia 14(3): 551-563. doi: 10.1590/S0101-81751997000300005
https://doi.org/10.1590/S0101-8175199700...
, 2004Baumgartner G, Nakatani K, Gomes LC, Bialetzki A, Sanches PV, Makrakis MC (2004) Identification of spawning sites and natural nurseries of fishes in the upper Paraná River, Brazil. Environmental Biology of Fishes 71: 115-125., Zaniboni-Filho & Schulz 2003Zaniboni-Filho E, Schulz UH (2003) Migratory fishes of the Uruguay river. In: Carolsfeld J, Harvey B, Baer A, Ross C (Orgs.), p. 135-168. Migratory fishes of the South América: biology social importance and conservation status. Victoria, World Fisheries Trust., Bialetzki et al. 2005Bialetzki A, Nakatani K, Sanches PV, Baumgartner G, Gomes LC (2005) Larval fish assemblage in the Baía River (Mato Grosso do Sul State, Brazil): temporal and spatial patterns. Environmental Biology of Fishes 73(1): 37-47., Reynalte-Tataje et al. 2008Reynalte-Tataje DA, Hermes-Silva S, Weiss LA, Zaniboni-Filho E (2008) Distribuição e abundância temporal do ictioplâncton no alto rio Uruguai, Brasil, p. 195-228. In: Zaniboni-Filho E, Nuñer APO (Eds.). Reservatório de Itá: estudos ambientais: desenvolvimento de tecnologias de cultivo e conservação da ictiofauna. Florianópolis, Editora UFSC, 319p., Tondato et al. 2010Tondato KK, Mateus LAF, Ziober SR (2010) Spatial and temporal distribution of fish larvae in marginal lagoons of Pantanal, Mato Grosso State, Brazil. Neotropical Ichthyology 8(1): 123-133. doi: 10.1590/S1679-62252010005000002
https://doi.org/10.1590/S1679-6225201000...
). In this study, larvae of the three species studied were captured in the same period, but the density of S. brasiliensis and P. corruscans decreased from February on. Likewise, considering the low levels of the Paraná River, the migratory fish species studied did spawn, strengthening the close relationship between their reproduction with the fluviometric level (Vazzoler 1996Vazzoler AEAM (1996) Biologia da reprodução de peixes teleósteos: teoria e prática. Maringá, Eduem, 196p.). These data also demonstrate the importance of the studied area for the migratory species, as there was an indication that at least two of them (S. brasiliensis and P. lineatus with record of yolk-sac larval stage) have spawning areas near the sampled sites.
The distribution pattern found for the larvae of the three species is consistent with the results of Nakatani et al. (1997Nakatani K, Baumgartner G, Cavicchioli M (1997) Ecologia de ovos e larvas de peixes, p. 281-306. In: Vazzoler AEAM, Agostinho AA, Hahn NS (Eds.). A planície de inundação do alto rio Paraná: aspectos físicos; biológicos e socioeconômicos. Maringá, Eduem, 460p.) and Agostinho et al. (2003 Agostinho AA , Gomes LC, Suzuki HI, Júlio Jr HF (2003) Migratory fish from the upper Paraná River basin, Brazil, p. 19-99. In: Carolsfeld J, Harvey B, Ross C, Baer A (Eds.). Migratory fishes of South America: biology, social importance and conservation status. Victoria, World Fisheries Trust, The World Bank and the International Development Research Center, 215p.). These authors described that during the spawning season, migratory species moved towards the headwaters of the tributaries, their pelagic eggs developed and hatched while drifting, and the larvae were conducted to the marginal aquatic environments, which are connected to the rivers during the flood season. Thus, the greater abundances of yolk-sac stage (YL) larvae of S. brasiliensis in the sampling sites of the Lower and Upper stretches are possibly the result of the influence of the rivers Iguatemi (tributary on the right bank) and Piquiri (tributary on the left bank); and from the rivers Ivinhema and Amambai (tributaries on the right bank) respectively, indicating that the species is using these environments as spawning areas. However, we cannot rule out the possibility that spawning had occurred in the main channel of the river as well, as observed for P. lineatus. Prochilodus lineatus larvae at the yolk-sac stage (YL) were captured in the sampling sites located in the main channel of Paraná River (Middle stretch). Although migratory species use the headwaters to spawn (Nakatani et al. 1997Nakatani K, Baumgartner G, Cavicchioli M (1997) Ecologia de ovos e larvas de peixes, p. 281-306. In: Vazzoler AEAM, Agostinho AA, Hahn NS (Eds.). A planície de inundação do alto rio Paraná: aspectos físicos; biológicos e socioeconômicos. Maringá, Eduem, 460p.), this species may also be spawning at this site (considered as a drift area), since it has high reproductive plasticity, as observed in hormone-induced spawning studies (Britski 1972Britski HA (1972) Peixes de água doce do Estado de São Paulo, p. 79-108. In: Branco SM (Org.). Poluição e piscicultura. São Paulo, Faculdade de Saúde Pública da USP, Instituto de Pesca - CPRN-AS, 216p., Godoy 1975Godoy MP (1975) Peixes do Brasil, Subordem Characoidei, Bacia do rio Mogi Guassu. Piracicaba, Editora Franciscana, 848p., Castro 1993Castro RMC (1993) Prochilodus britskii, a new species of Prochilodus fish (Ostariophysi; Characiformes) from the rio Apiacá, river Tapajós system, Mato Grosso, Brazil. Proceedings of the Biological Society of Washington 106: 57-62.).
The absence of P. corruscans larvae at the yolk-sac larval stage is not evidence that it does not spawn in the studied area. This is because a great number of pre-flexion larvae were registered in the Upper stretch, indicating that the species uses the vicinities of the area for spawning and uses the main channel of the river too (Godinho et al. 2007Godinho AL, Kynard B, Godinho HP (2007) Migration and spawning of female surubim (Pseudoplatystoma corruscans, Pimelodidae) in the São Francisco River, Brazil. Environmental Biology of Fishes 80: 421-433. doi: 10.1007./s10641-006-9141-1
https://doi.org/10.1007./s10641-006-9141...
). Its reproductive period is relatively short, covering the rainy months (November to January); the spawning is total or single (Bazzoli 2003Bazzoli N (2003) Parâmetros reprodutivos dos peixes de interesse comercial na região de Pirapora, p. 291-306. In: Godinho HP, Godinho AL (Eds.). Águas, peixes e pescadores do São Francisco das Minas Gerais. Belo Horizonte, Editora PUC Minas, CNPq/PADCT CIAMB III, 468p., Sato & Godinho 2003Sato Y, Godinho HP (2003) Migratory fishes of the São Francisco River, p 195-231. In: Carolsfeld J, Harvey B, Ross C, Baer A (Eds.). Migratory fishes of South America: Biology, Social importance and conservation status. Victoria, World Fisheries Trust, the World Bank and the International Development Research Center, 215p.).
Surveys carried out by Nakatani et al. (2004Nakatani K, Bialetzki A, Sanches PV (2004) Eggs and larvae of fishes in the upper Paraná River floodplain, p. 157-161. In: Agostinho AA, Rodrigues L, Gomes LC, Thomaz SM, Miranda LE (Eds.). Structure and functioning of the Paraná River and its floodplain. Maringá, Eduem, 275p.) demonstrate the importance of the tributaries for fish reproduction in the Upper Paraná River floodplain. These rivers are still not dammed, and their water level variations are exclusively controlled by their hydrological cycle (rain on the watersheds). Moreover, they present several marginal lakes and extensive flooding areas along their courses, providing favorable conditions for spawning and larval growth (Baumgartner et al. 2004Baumgartner G, Nakatani K, Gomes LC, Bialetzki A, Sanches PV, Makrakis MC (2004) Identification of spawning sites and natural nurseries of fishes in the upper Paraná River, Brazil. Environmental Biology of Fishes 71: 115-125., 2008Baumgartner G, Nakatani K, Gomes LC, Bialetzki A, Sanches PV, Makrakis MC (2008) Fish larvae from the upper Paraná River: do abiotic factors affect larval density? Neotropical Ichthyology 6(4): 551-558., Daga et al. 2009Daga VS, Gogola TM, Sanches PV, Baumgartner G, Baumgartner D, Piana PA, Gubian EA, Delariva RL (2009) Fish larvae assemblages in two floodplain lakes with different degrees of connection to the Paraná River, Brazil. Neotropical Ichthyology 7: 429-438. doi: 10.1590/S1679-62252009000300010
https://doi.org/10.1590/S1679-6225200900...
, Gogola et al. 2010Gogola TM, González LMA, Daga VS, Silva PRL, Sanches PV, Gubiani EA, Baumgartner G, Delariva RL (2010) Spatial and temporal distribution patterns of ichthyoplankton in a region affected by water regulation by dams. Neotropical Ichthyology 8: 341-349. doi: 10.1590/S1679-62252010000200013
https://doi.org/10.1590/S1679-6225201000...
). The results obtained in our analysis of distribution of developmental stages reinforce their importance. Some studies performed in Ivinhema River suggest that this is one of the main environments for fish reproduction in the Upper Paraná River (Baumgartner et al. 1997Baumgartner G, Nakatani K, Cavicchioli M, Baumgartner MST (1997) Some aspects of the ecology of fish larvae in the floodplain of the high Paraná River, Brazil. Revista Brasileira de Zoologia 14(3): 551-563. doi: 10.1590/S0101-81751997000300005
https://doi.org/10.1590/S0101-8175199700...
, Nakatani et al. 1997Nakatani K, Baumgartner G, Cavicchioli M (1997) Ecologia de ovos e larvas de peixes, p. 281-306. In: Vazzoler AEAM, Agostinho AA, Hahn NS (Eds.). A planície de inundação do alto rio Paraná: aspectos físicos; biológicos e socioeconômicos. Maringá, Eduem, 460p., Sanches et al. 2006Sanches PV, Nakatani K, Bialetzki A, Baumgartner G, Gomes LC, Luiz EA (2006) Flow regulation dams affecting ichthyoplankton: the case of Porto Primavera dam; Paraná River; Brazil. River Research App 22: 555-565. doi: 10.1002/rra.922
https://doi.org/10.1002/rra.922...
), probably due to the pristine characteristics of this river (part of this river is located in a State Park and most of it is a Protect Area), with intense riparian vegetation and extensive flooded areas (Souza Filho & Stevaux 1997Souza Filho EE, Stevaux JC (1997) Geologia e geomorfologia do complexo rio Baia, Curutuba, Ivinheima, p. 3-46. In: Vazzoler AEAM, Agostinho AA, Hahn NS (Eds.). A planície de inundação do alto rio Paraná: aspectos físicos; biológicos e socioeconômicos. Maringá, Eduem, 460p.).
The capture of S. brasiliensis larvae only occurred between October and December, at the three stages of development (yolk-sac larvae, pre-flexion and flexion). The record of flexion larvae suggests that the reproductive period of the species may have started in September, when an increase in the level of the river was registered (data not shown), leading to spawning. On the other hand, P. corruscans was captured from October to January, with higher larval density at the pre-flexion stage recorded in December, with occurrence also in October, which may indicate that this species, as well as S. brasiliensis, also started its reproductive period in September. Nakatani et al. (1997Nakatani K, Baumgartner G, Cavicchioli M (1997) Ecologia de ovos e larvas de peixes, p. 281-306. In: Vazzoler AEAM, Agostinho AA, Hahn NS (Eds.). A planície de inundação do alto rio Paraná: aspectos físicos; biológicos e socioeconômicos. Maringá, Eduem, 460p.) observed high numbers of eggs and larvae in September in the region of the Upper Paraná River floodplain. This occurrence deserves further attention, since the strategy to protect these species in the region consider the period between November and February as their main spawning season.
Unlike these species, P. lineatus presented the lowest larval density. This result is unexpected, since this species has greater reproductive activity when the values of flow and water level increase (Agostinho et al. 2004bAgostinho AA, Gomes LC, Veríssimo S, Okada EK (2004b) Flood regime, dam regulation and fish in the Upper Paraná River: effects on assemblage attributes, reproduction and recruitment. Reviews in Fish Biology and Fisheries 14: 11-19.). Again, the low values of the river level registered in January may have influenced these values, also indicating that P. lineatus starts reproducing later.
The study period, between October and March, is characterized by high values of temperature and precipitation that seem to be the main triggers for reproduction (Vazzoler 1996Vazzoler AEAM (1996) Biologia da reprodução de peixes teleósteos: teoria e prática. Maringá, Eduem, 196p.) and also appear to affect larval abundance in several habitats (Araújo-Lima & Oliveira 1998Araújo-Lima CARM, Oliveira EC (1998) Transport of larval fish in the Amazon. Journal of Fish Biology 53: 297-306. doi: 10.1111/j.1095-8649.1998.tb01033.x
https://doi.org/10.1111/j.1095-8649.1998...
). Studies conducted at the Upper Paraná River also identify these variables as determinants of the distribution of abundances of eggs and larvae of fish (Nakatani et al. 2001Nakatani K, Agostinho AA, Baumgartner G, Bialetzki A, Sanches PV, Makrakis MC, Pavanelli CS (2001) Ovos e larvas de peixe de água doce: Desenvolvimento e manual de identificação. Maringá, Eduem, 378p., Castro et al. 2002Castro RJ, Nakatani K, Bialetzki A, Sanches PV, Baumgartner G (2002) Temporal distribution and composition of the ichthyoplankton from Leopold´s Inlet on the upper Paraná River floodplain. Journal of Zoology 256(4): 437-443. doi: 10.1017/S095283690200047X
https://doi.org/10.1017/S095283690200047...
, Bialetzki et al. 2005Bialetzki A, Nakatani K, Sanches PV, Baumgartner G, Gomes LC (2005) Larval fish assemblage in the Baía River (Mato Grosso do Sul State, Brazil): temporal and spatial patterns. Environmental Biology of Fishes 73(1): 37-47., Sanches et al. 2006Sanches PV, Nakatani K, Bialetzki A, Baumgartner G, Gomes LC, Luiz EA (2006) Flow regulation dams affecting ichthyoplankton: the case of Porto Primavera dam; Paraná River; Brazil. River Research App 22: 555-565. doi: 10.1002/rra.922
https://doi.org/10.1002/rra.922...
, Baumgartner et al. 2008Baumgartner G, Nakatani K, Gomes LC, Bialetzki A, Sanches PV, Makrakis MC (2008) Fish larvae from the upper Paraná River: do abiotic factors affect larval density? Neotropical Ichthyology 6(4): 551-558., Reynalte-Tataje et al. 2011aReynalte-Tataje DA, Nakatani K, Fernandes R, Agostinho AA, Bialetzki A (2011a) Temporal distribution of ichthyoplankton in the Ivinhema River (Mato Grosso do Sul State/Brazil): influence of environmental variables. Neotropical Ichthyology 9(2): 427-436. doi: 10.1590/S1679-62252011005000017
https://doi.org/10.1590/S1679-6225201100...
). However, despite certain trends in fish communities, the differences among the reproductive strategies within a community enable the populations to respond differently to environmental variables at the regional level (Reynalte-Tataje et al. 2011bReynalte-Tataje DA, Agostinho AA, Bialetzki A, Hermes-Silva S, Fernandes R, Zaniboni-Filho E (2011b) Spatial and temporal variation of the ichthyoplankton in a subtropical river in Brazil. Environmental Biology of Fishes 94: 403-419. doi: 10.1007/s10641-011-9955-3
https://doi.org/10.1007/s10641-011-9955-...
). Although the three species studied herein have a limited biological plasticity (Agostinho et al. 1993Agostinho AA, Mendes VP, Suzuki HI, Canzi C (1993) Avaliação da atividade reprodutiva da comunidade de peixes dos primeiros quilômetros a jusante do reservatório de Itaipu. Revista Unimar 15: 175-189.), there was no significant influence of the environmental variables on the abundance of their larvae. However, an increase in photoperiod and temperature determine the start of gonad maturation in tropical fish, which is intensified during the reproductive period (Vazzoler 1996Vazzoler AEAM (1996) Biologia da reprodução de peixes teleósteos: teoria e prática. Maringá, Eduem, 196p.). This was observed in the present study, primarily for S. brasiliensis and P. corruscans, which had already been recorded as spawning in October. Nevertheless, the real importance of each of these variables for reproduction is not completely clear.
Temperature probably was a limiting factor for S. brasiliensis, since in general larvae of this species were registered in the sampled months with lower temperatures. Meanwhile, it is worth mentioning again that the Mantel test was not significant, pointing to the weak associations between the matrices of larval abundance and abiotic variables. In addition, it should be remembered that the levels of the river were low in the warm months (January and February). Indeed the predominance of larvae of S. brasiliensis and P. corruscans between October and December reveals that this species has reproductive activity in this period.
The low levels of the river recorded in January may result from the absence of rainfall or water retention by the upstream dams. The presence of dams creates irreversible impacts, mainly changing the natural pulses of flood and drought and their timing, which is detrimental to the spawning of species with reproductive cycles that depend on the rising of the fluviometric level (Agostinho et al. 2004aAgostinho AA, Thomaz SM, Gomes LC (2004a) Threats for biodiversity in the floodplain of upper Paraná River: effects of hydrological regulation by dams. Ecohydrology and Hydrobiology 4(3): 267-280., Sanches et al. 2006Sanches PV, Nakatani K, Bialetzki A, Baumgartner G, Gomes LC, Luiz EA (2006) Flow regulation dams affecting ichthyoplankton: the case of Porto Primavera dam; Paraná River; Brazil. River Research App 22: 555-565. doi: 10.1002/rra.922
https://doi.org/10.1002/rra.922...
). Nonetheless the impact would be worse if there were no tributaries, along with the Saraiva Lake, which are important for fish reproduction and growth (nursery) (Gogola et al. 2010Gogola TM, González LMA, Daga VS, Silva PRL, Sanches PV, Gubiani EA, Baumgartner G, Delariva RL (2010) Spatial and temporal distribution patterns of ichthyoplankton in a region affected by water regulation by dams. Neotropical Ichthyology 8: 341-349. doi: 10.1590/S1679-62252010000200013
https://doi.org/10.1590/S1679-6225201000...
). We should also point out that this study may have been hampered by low flows. However, periods like this tend to become more frequent with the predicted climate change.
Based on the results obtained in this study, we conclude that S. brasiliensis, P. corruscans and P. lineatus seek the dam-free tributaries for spawning. However, we cannot rule out the hypothesis that they also spawn in the lotic stretch of the main river. Godinho et al. (2007Godinho AL, Kynard B, Godinho HP (2007) Migration and spawning of female surubim (Pseudoplatystoma corruscans, Pimelodidae) in the São Francisco River, Brazil. Environmental Biology of Fishes 80: 421-433. doi: 10.1007./s10641-006-9141-1
https://doi.org/10.1007./s10641-006-9141...
) observed spawning sites of P. corruscans in the lotic stretch of the São Francisco River. The reproductive dynamics, recruitment, maintenance, and possible attempts to recover the stocks of these species are directly dependent on the preservation of tributaries, especially the Piquiri, Iguatemi and Amambaí rivers and lakes in the region, and also of a close monitoring to prevent fishing during the reproductive period.
ACKNOWLEDGEMENTS
We express our gratitude to the Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura (Nupélia), which made materials and equipments available to us; the Parque Tecnológico Itaipu (PTI) for granting a master scholarship to EBS, and to the Universidade Paranaense for funding the project. We also thank to the Consórcio Intermunicipal para Conservação do Remanescente do Rio Paraná e Áreas de Influência (Coripa) and to the Instituto Ambiental do Paraná (IAP), for their support in fieldwork. LCG and AB is Bolsista Produtividade of CNPq. The programs PIE/PELD (Process 520.026/98-5) and CNPq (Process 476162/04-1) financed the research in the Ivinhema River.
LITERATURE CITED
- Abilhoa V, Duboc LF (2004) Peixes, p. 581-682. In: Mikichs B, Bérnils RS (Eds.). Livro vermelho da fauna ameaçada no Estado do Paraná. Curitiba, Governo do Estado do Paraná, SEMA, IAP, 763p.
- Agostinho AA, Mendes VP, Suzuki HI, Canzi C (1993) Avaliação da atividade reprodutiva da comunidade de peixes dos primeiros quilômetros a jusante do reservatório de Itaipu. Revista Unimar 15: 175-189.
- Agostinho AA, Vazzoler AEAM, Thomaz SM (1995) The high River Paraná basin: limnological and ichthyological aspects, p. 59-103. In: Tundisi JG, Bicudo CEM, Matsumura-Tundisi T (Eds.). Limnology in Brazil. Rio de Janeiro, ABC/SBL, 376p.
- Agostinho AA, Júlio Jr HF, Gomes LC, Bini LM, Agostinho CS (1997) Composição, abundância e distribuição espaço-temporal da ictiofauna, p. 179-208. In: Vazzoler AEAM, Agostinho AA, Hahn NS (Eds.). A planície de inundação do alto rio Paraná: aspectos físicos; biológicos e socioeconômicos. Maringá, Eduem, 460p.
- Agostinho AA, Thomaz SM, Minte-Vera CV, Winemiller KO (2000) Biodiversity in the high Paraná River floodplain, p. 89-118. In: Gopal B, Junk WDJ, Davis JA (Eds.). Biodiversity in wetlands: assessment, function and conservation. Leiden, Backhuys Publishers, 310p.
- Agostinho AA , Gomes LC, Suzuki HI, Júlio Jr HF (2003) Migratory fish from the upper Paraná River basin, Brazil, p. 19-99. In: Carolsfeld J, Harvey B, Ross C, Baer A (Eds.). Migratory fishes of South America: biology, social importance and conservation status. Victoria, World Fisheries Trust, The World Bank and the International Development Research Center, 215p.
- Agostinho AA, Thomaz SM, Gomes LC (2004a) Threats for biodiversity in the floodplain of upper Paraná River: effects of hydrological regulation by dams. Ecohydrology and Hydrobiology 4(3): 267-280.
- Agostinho AA, Gomes LC, Veríssimo S, Okada EK (2004b) Flood regime, dam regulation and fish in the Upper Paraná River: effects on assemblage attributes, reproduction and recruitment. Reviews in Fish Biology and Fisheries 14: 11-19.
- Agostinho AA, Gomes LC, Pelicice FM (2007) Ecologia e manejo de recursos pesqueiros em reservatórios do Brasil. Maringá, Eduem, 501p.
- Ahlstrom EH, Ball OP (1954) Description of eggs and larvae of jack mackerel (Trachurus symmetricus) and distribution and abundance of larvae in 1950 and 1951. Fishery Bulletin 56: 209-245.
- Ali M, Wootton RJ (1999) Effect of variable food on reproductive performance breeding female three-spined sticklebacks. Journal of Fish Biology 55: 1040-1053. doi: 10.1111/j.1095-8649.1999.tb00739.x
» https://doi.org/10.1111/j.1095-8649.1999.tb00739.x - Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Austral Ecology 26: 32-46. doi: 10.1111/j.1442-9993.2001.01070.pp.x
» https://doi.org/10.1111/j.1442-9993.2001.01070.pp.x - Anderson MJ (2006) Distance-based tests for homogeneity of multivariate dispersions. Biometrics 62: 245-253.
- Anderson MJ, Ellingsen KE, McArdle BH (2006) Multivariate dispersion as a measure of beta diversity. Ecology Letters 9(6): 683-693.
- Araújo-Lima CARM, Oliveira EC (1998) Transport of larval fish in the Amazon. Journal of Fish Biology 53: 297-306. doi: 10.1111/j.1095-8649.1998.tb01033.x
» https://doi.org/10.1111/j.1095-8649.1998.tb01033.x - Baumgartner G, Nakatani K, Cavicchioli M, Baumgartner MST (1997) Some aspects of the ecology of fish larvae in the floodplain of the high Paraná River, Brazil. Revista Brasileira de Zoologia 14(3): 551-563. doi: 10.1590/S0101-81751997000300005
» https://doi.org/10.1590/S0101-81751997000300005 - Baumgartner G, Nakatani K, Gomes LC, Bialetzki A, Sanches PV, Makrakis MC (2004) Identification of spawning sites and natural nurseries of fishes in the upper Paraná River, Brazil. Environmental Biology of Fishes 71: 115-125.
- Baumgartner G, Nakatani K, Gomes LC, Bialetzki A, Sanches PV, Makrakis MC (2008) Fish larvae from the upper Paraná River: do abiotic factors affect larval density? Neotropical Ichthyology 6(4): 551-558.
- Bazzoli N (2003) Parâmetros reprodutivos dos peixes de interesse comercial na região de Pirapora, p. 291-306. In: Godinho HP, Godinho AL (Eds.). Águas, peixes e pescadores do São Francisco das Minas Gerais. Belo Horizonte, Editora PUC Minas, CNPq/PADCT CIAMB III, 468p.
- Bialetzki A, Nakatani K, Sanches PV, Baumgartner G, Gomes LC (2005) Larval fish assemblage in the Baía River (Mato Grosso do Sul State, Brazil): temporal and spatial patterns. Environmental Biology of Fishes 73(1): 37-47.
- Britski HA (1972) Peixes de água doce do Estado de São Paulo, p. 79-108. In: Branco SM (Org.). Poluição e piscicultura. São Paulo, Faculdade de Saúde Pública da USP, Instituto de Pesca - CPRN-AS, 216p.
- Castro RMC (1993) Prochilodus britskii, a new species of Prochilodus fish (Ostariophysi; Characiformes) from the rio Apiacá, river Tapajós system, Mato Grosso, Brazil. Proceedings of the Biological Society of Washington 106: 57-62.
- Castro RJ, Nakatani K, Bialetzki A, Sanches PV, Baumgartner G (2002) Temporal distribution and composition of the ichthyoplankton from Leopold´s Inlet on the upper Paraná River floodplain. Journal of Zoology 256(4): 437-443. doi: 10.1017/S095283690200047X
» https://doi.org/10.1017/S095283690200047X - Clarke KR, Somerfield PJ, Chapman MG (2006) On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray-Curtis coefficient for denuded assemblages. Journal of Experimental Marine Biology and Ecology 330(1): 55-80. doi: 10.1016/j.jembe.2005.12.017
» https://doi.org/10.1016/j.jembe.2005.12.017 - Daga VS, Gogola TM, Sanches PV, Baumgartner G, Baumgartner D, Piana PA, Gubian EA, Delariva RL (2009) Fish larvae assemblages in two floodplain lakes with different degrees of connection to the Paraná River, Brazil. Neotropical Ichthyology 7: 429-438. doi: 10.1590/S1679-62252009000300010
» https://doi.org/10.1590/S1679-62252009000300010 - Godinho AL, Kynard B, Godinho HP (2007) Migration and spawning of female surubim (Pseudoplatystoma corruscans, Pimelodidae) in the São Francisco River, Brazil. Environmental Biology of Fishes 80: 421-433. doi: 10.1007./s10641-006-9141-1
» https://doi.org/10.1007./s10641-006-9141-1 - Godoy MP (1975) Peixes do Brasil, Subordem Characoidei, Bacia do rio Mogi Guassu. Piracicaba, Editora Franciscana, 848p.
- Gomes LC, Agostinho AA (1997) Influence of the flooding regime on the nutritional state and juvenile recruitment of the curimba, Prochilodus scrofa, Steindachner in upper Paraná River, Brazil. Fisheries Management and Ecology 4: 263-274.
- Gogola TM, González LMA, Daga VS, Silva PRL, Sanches PV, Gubiani EA, Baumgartner G, Delariva RL (2010) Spatial and temporal distribution patterns of ichthyoplankton in a region affected by water regulation by dams. Neotropical Ichthyology 8: 341-349. doi: 10.1590/S1679-62252010000200013
» https://doi.org/10.1590/S1679-62252010000200013 - McArdle BH, Anderson MJ (2001) Fitting multivariate models to community data: a comment on distance based redundancy analysis. Ecology 82: 290-297. doi: 10.1890/0012-9658(2001)082[0290:FMMTCD]2.0.CO;2
» https://doi.org/10.1890/0012-9658(2001)082[0290:FMMTCD]2.0.CO;2 - McCune B, Mefford MJ (1999) PC-ORD. Multivariate analysis of ecological data. Gleneden Beach, MjM Software Design, Version 4.01.
- Nakatani K, Baumgartner G, Cavicchioli M (1997) Ecologia de ovos e larvas de peixes, p. 281-306. In: Vazzoler AEAM, Agostinho AA, Hahn NS (Eds.). A planície de inundação do alto rio Paraná: aspectos físicos; biológicos e socioeconômicos. Maringá, Eduem, 460p.
- Nakatani K, Agostinho AA, Baumgartner G, Bialetzki A, Sanches PV, Makrakis MC, Pavanelli CS (2001) Ovos e larvas de peixe de água doce: Desenvolvimento e manual de identificação. Maringá, Eduem, 378p.
- Nakatani K, Bialetzki A, Sanches PV (2004) Eggs and larvae of fishes in the upper Paraná River floodplain, p. 157-161. In: Agostinho AA, Rodrigues L, Gomes LC, Thomaz SM, Miranda LE (Eds.). Structure and functioning of the Paraná River and its floodplain. Maringá, Eduem, 275p.
- Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O'Hara RB, Simpson GL, Henry M, Stevens H, Wagner H (2013) Vegan: Community Ecology Package. R package version 2.0-8. Available online at: Available online at: http://CRAN.R-project.org/package=vegan [Accessed: 10/11/2014]
» http://CRAN.R-project.org/package=vegan - Petry AC, Agostinho AA, Gomes LC (2003) Fish assemblages of tropical floodplain lagoons: exploring the role of connectivity in a dry year. Neotropical Ichthyology 1(2): 111-119. doi: 10.1590/S1679-62252003000200005
» https://doi.org/10.1590/S1679-62252003000200005 - Reynalte-Tataje DA, Hermes-Silva S, Weiss LA, Zaniboni-Filho E (2008) Distribuição e abundância temporal do ictioplâncton no alto rio Uruguai, Brasil, p. 195-228. In: Zaniboni-Filho E, Nuñer APO (Eds.). Reservatório de Itá: estudos ambientais: desenvolvimento de tecnologias de cultivo e conservação da ictiofauna. Florianópolis, Editora UFSC, 319p.
- Reynalte-Tataje DA, Nakatani K, Fernandes R, Agostinho AA, Bialetzki A (2011a) Temporal distribution of ichthyoplankton in the Ivinhema River (Mato Grosso do Sul State/Brazil): influence of environmental variables. Neotropical Ichthyology 9(2): 427-436. doi: 10.1590/S1679-62252011005000017
» https://doi.org/10.1590/S1679-62252011005000017 - Reynalte-Tataje DA, Agostinho AA, Bialetzki A, Hermes-Silva S, Fernandes R, Zaniboni-Filho E (2011b) Spatial and temporal variation of the ichthyoplankton in a subtropical river in Brazil. Environmental Biology of Fishes 94: 403-419. doi: 10.1007/s10641-011-9955-3
» https://doi.org/10.1007/s10641-011-9955-3 - Sanches PV, Baumgartner G, Bialetzki A, Suiberto MR, Gomes FDC, Nakatani K, Barbosa NDC (2001) Caracterização do desenvolvimento inicial de Leporinus friderici (Osteichthyes; Anostomidae) da bacia do rio Paraná. Acta Scientiarum 23: 383-389. doi: 10.4025/actascibiolsci.v23i0.2693
» https://doi.org/10.4025/actascibiolsci.v23i0.2693 - Sanches PV, Nakatani K, Bialetzki A, Baumgartner G, Gomes LC, Luiz EA (2006) Flow regulation dams affecting ichthyoplankton: the case of Porto Primavera dam; Paraná River; Brazil. River Research App 22: 555-565. doi: 10.1002/rra.922
» https://doi.org/10.1002/rra.922 - Sato Y, Godinho HP (2003) Migratory fishes of the São Francisco River, p 195-231. In: Carolsfeld J, Harvey B, Ross C, Baer A (Eds.). Migratory fishes of South America: Biology, Social importance and conservation status. Victoria, World Fisheries Trust, the World Bank and the International Development Research Center, 215p.
- Souza Filho EE, Stevaux JC (1997) Geologia e geomorfologia do complexo rio Baia, Curutuba, Ivinheima, p. 3-46. In: Vazzoler AEAM, Agostinho AA, Hahn NS (Eds.). A planície de inundação do alto rio Paraná: aspectos físicos; biológicos e socioeconômicos. Maringá, Eduem, 460p.
- Statsoft (2005) Electronic Statistic textbook. Available online at: Available online at: http://www.statsoft.com/textbook/stathome.html [Accessed: 5/2/2011].
» http://www.statsoft.com/textbook/stathome.html - Suzuki HI, Agostinho AA (1997) Reprodução de peixes do reservatório de Segredo, p. 163-180. In: Agostinho AA, Gomes LC (Eds.). Reservatório de Segredo: bases ecológicas para o manejo. Maringá, Eduem, 387p.
- Tanaka S (1973) Stock assessment by means of ichthyoplankton surveys. FAO Fish Tech Paper 122: 33-51.
- Tondato KK, Mateus LAF, Ziober SR (2010) Spatial and temporal distribution of fish larvae in marginal lagoons of Pantanal, Mato Grosso State, Brazil. Neotropical Ichthyology 8(1): 123-133. doi: 10.1590/S1679-62252010005000002
» https://doi.org/10.1590/S1679-62252010005000002 - Vazzoler AEAM (1996) Biologia da reprodução de peixes teleósteos: teoria e prática. Maringá, Eduem, 196p.
- Welcomme RL (1979) Fisheries ecology of floodplain rivers. London, Longman, 317p.
- Wootton RJ (1991) Ecology of teleost fishes. London, Champman and Hall, 404p.
- Zaniboni-Filho E, Schulz UH (2003) Migratory fishes of the Uruguay river. In: Carolsfeld J, Harvey B, Baer A, Ross C (Orgs.), p. 135-168. Migratory fishes of the South América: biology social importance and conservation status. Victoria, World Fisheries Trust.
Publication Dates
-
Publication in this collection
Jul-Aug 2015
History
-
Received
21 Aug 2014 -
Reviewed
23 Apr 2014 -
Accepted
02 May 2015