Abstract
To assess the genetic potential of white corn for corn grits, 28 hybrids resulting from diallel crosses of eight parents, including parents and S1 generations, were evaluated. The combining abilities and levels of heterosis and inbreeding depression were estimated. The parents used were the main white maize hybrids of the Brazilian germplasm. The combinations HT 9332 x HT 392, IAC Nelore x HT 9332, IAC Nelore x HT 932, and IPR 119 x IAC Nelore are promising for reciprocal recurrent selection programs targeting higher grain yield. Hybrid IPR 127 was indicated as a source of industrial lines with favorable traits for dry-rolled corn production. Hybrid IAC Nelore had the lowest estimate of inbreeding depression and was recommended for the breeding of lines and participation in higher-yielding hybrids and composites.
Key words:
Zea mays L.; heterosis; grit production
Resumo
Com o objetivo de avaliar o potencial genético de milho branco para canjica, foram avaliados 28 híbridos resultantes de cruzamentos dialélicos entre oito genitores, incluindo os parentais e as gerações S1. Foram estimadas as capacidades combinatórias e os níveis de heterose e depressão endogâmica. Os genitores utilizados foram os principais híbridos de milho branco pertencentes ao germoplasma brasileiro. As combinações HT 9332 x HT 392, IAC Nelore x HT 9332, IAC Nelore x HT 932 e IPR 119 x IAC Nelore são promissoras para programas de seleção recorrente recíproca para maior rendimento de grãos. O híbrido IPR 127 foi indicado como fonte de linhagens com características industriais favoráveis ao canjicamento. O híbrido IAC Nelore apresentou a menor estimativa de depressão por endogamia e foi recomendado para a extração de linhagens e participação em híbridos e compostos mais produtivos.
Palavras-chave:
Zea mays L.; heterose; rendimento de canjica
INTRODUCTION
The transformation of maize in different derivatives allows the use of cereals as raw material for multiple applications in the food industry. Approximately 90 different products are derived from corn, among which grits, corn meal, hominy, oil, starch, amylose, amylopectin, zein, and fibers are worth mentioning for their economic importance. The use of degerminated and ground grain for the production of "fast-food" in the form of extruded foods, commonly known as "snacks", is a major application of corn in the food industry (Gonçalves et al. 2003Gonçalves RA, Santos JP, Tomé PHF, Pereira RGFA, Ascheri JLR and Abreu CMP (2003) Yield and chemical composition of corn cultivate in dry-milling and grit production. Ciência e Agrotecnologia 27: 643-650.).
The corn-processing companies that exploit the specialty market have only five white maize cultivars available for grit production (hybrids IPR 119 and IPR 127 and varieties AL Bianco, BR 451 and RS 21). In addition, no studies are available in the literature would provide a theoretical framework of the estimation of genetic parameters, especially for economically relevant traits, e.g., grit production.
The so-called "canjica" is the corn grain or part of it, with partial or total absence of germ resulting from a scraping process, degermination (Castro et al. 2009Castro MVL, Naves MMV, Oliveira JP and Oliveira Froes L (2009) Rendimento industrial e composição química de milho de alta qualidade proteica em relação a híbridos comerciais. Pesquisa Agropecuária Tropical 39: 233-242.). In this process, the grain is broken and divided into endosperm, germ and pericarp.
In Brazil, degermination is known as a semi-wet process, in which the corn grain is immersed in hot water and steam is injected. Thus, maize degermination without endosperm disintegration results in higher grit quality. The process can also occur without moistening the grain, using a corn degerminator.
After degermination, the corn grits can be grouped in size classes classified according to the abrasion intensity or for degermination, in subgroups (unpolished; polished), according to color classes (yellow, white or mixed) or for the levels of impurities. If the defect percentage exceeds the maximum tolerance limits, the grains should be classified as below standard, or disqualified when unfit for human or animal consumption.
The purpose of this study is to promote the use of maize as corn grits, by making new cultivars available on the market. The breeding program was initiated with the evaluation of the germplasm by diallel crosses among eight parents from adapted genotypes (commercial hybrids). The general and specific combining abilities were estimated by the methodology of Griffing (1956Griffing B (1956) Concept of general and specific combining ability in relation to diallel crossing systems. Australian Journal Biology Science 9: 463-493.), as well as heterosis parameters by the methodology of Gardner and Eberhart (1966Gardner CO and Eberhart SA (1966) Analysis and interpretation of the variety cross diallel and related populations. Biometrics 22: 439-452.). The magnitude of inbreeding depression of the parents was estimated by the methodology of Gardner (1965Gardner CO and Eberhart SA (1966) Analysis and interpretation of the variety cross diallel and related populations. Biometrics 22: 439-452.), and the mean of the possible sets of lines that can be developed from these parents  , as proposed by Vencovsky and Barriga (1992Vencovsky R and Barriga P (1992) Genética biométrica no fitomelhoramento. Revista Brasileira de Genética, Ribeirão Preto, 496p.).
, as proposed by Vencovsky and Barriga (1992Vencovsky R and Barriga P (1992) Genética biométrica no fitomelhoramento. Revista Brasileira de Genética, Ribeirão Preto, 496p.).
MATERIAL AND METHODS
The crosses to obtain the hybrid combinations were performed in the second growing season of 2009. The experimental evaluations of the parents, the hybrids and their selfings were performed on the Experimental Farm of Iguatemi - FEI (lat 23º 25' S, long 51º 57' W, alt 510 m asl), in Maringá (PR), in the growing seasons 2009/2010 and 2010/2011.
The experiments were designed as complete blocks with randomized treatments and three replications. We evaluated 44 treatments, of which 8 were parents; 28 hybrids obtained by crossing parents in a complete diallel scheme (excluding the reciprocals) and 8 genotypes belonging to the S1 generation derived from selfing of the parents used. The description of the eight genotypes used in this study is given in Table 1.
Each experimental unit consisted of two 15-m rows, spaced 0.90 m apart, and plants spaced 0.20 m apart, with a total assessed area of 27 m2. Fertilization and management practices were applied according to technical recommendations for corn (Galvão and Miranda 2004Galvão JCC and Miranda GV (2004) Tecnologias de produção de milho., Editora UFV, Viçosa 366p. ).
Grain yield data (YD, in t ha-1) were collected and standardized to 13% moisture. The percentages of grit yield (GY), small grit yield (SY) and bran residue (BY) were obtained according to the procedure proposed by Gonçalves (2003Gonçalves RA, Santos JP, Tomé PHF, Pereira RGFA, Ascheri JLR and Abreu CMP (2003) Yield and chemical composition of corn cultivate in dry-milling and grit production. Ciência e Agrotecnologia 27: 643-650.).
For grit yield, a clean sample of 10 kg corn grain per plot was prepared. Each sample was placed in a drum type gritting processor for a pre-set period of 22 seconds per kilogram of sample, at a rotation of 600 rpm of the knive blades of the degerminador. After abrasion, the resulting material was separated into grits and bran using a Tyler 5 sieve. The grits were manually extracted from the bran in a 4 mesh sieve (openings 5.54 mm, wire 21 to 0.81 mm). Soon after the sample had been passed through the gritting processor, the fractions grits (canjica, fraction corresponding to the endosperm), small grits and germ (embryo) were weighed.
From the measured traits, individual analysis of variance was performed with treatment partitioning as follows: parents, F1 hybrid combinations, S1 generations, and the contrasts F1 hybrids vs parents and parents + F1 hybrids vs S1 generations.
The analysis of variance for traits as affected by the environments was carried out considering the genotype and environment effects as fixed, by the mathematical model:

where:  is the observation of variable
is the observation of variable  in the k-th block of the i-th treatment in the j-th environment;
in the k-th block of the i-th treatment in the j-th environment;  is the general mean;
is the general mean;  is the effect of the i-th treatment with i = 1, 2,..., g;
is the effect of the i-th treatment with i = 1, 2,..., g;  is the effect of the j-th environment, with j = 1, 2,..., e;
is the effect of the j-th environment, with j = 1, 2,..., e;  is the interaction effect of the i-th treatment with the j-th environment;
is the interaction effect of the i-th treatment with the j-th environment;  is the effect of the k-th block within the j-th environment, with k = 1, 2,..., r; and
is the effect of the k-th block within the j-th environment, with k = 1, 2,..., r; and  is the random error associated with observation
is the random error associated with observation  .
.
The partitioning of treatment of the treatment x environment interaction was performed as follows: parents x environment, F1 hybrid combinations x environments, S1 generations x environments, and the contrasts ENT#091;F1 hybrid combinations vs parentsENT#093; x environment and ENT#091;parents + F1 hybrids vs S1 generationsENT#093; x environment. Means were compared by the Scott-Knott test at 5% probability.
Combined diallel analyses were performed using method 2 (parents and F1) of Griffing (1956Griffing B (1956) Concept of general and specific combining ability in relation to diallel crossing systems. Australian Journal Biology Science 9: 463-493.) and by the method of Gardner and Eberhart (1966Gardner CO and Eberhart SA (1966) Analysis and interpretation of the variety cross diallel and related populations. Biometrics 22: 439-452.), developed for the analysis of heterosis parameters, both adapted by Cruz et al. (2012Cruz CD, Regazzi AJ and Carneiro PCS (2012) Modelos biométricos aplicados ao melhoramento genético., Editora UFV, Viçosa 514p.). The methodology proposed by Gardner and Eberhart (1966) details the interpretation of the potential per se of the varieties and their hybrids, from the estimation of the variety effect and the partitioning of the total heterosis in mean heterosis, varietal and specific heterosis effects.
The inbreeding depression percentage of the parents was estimated by the method proposed by Gardner (1965Gardner CO (1965) Teoria de genética estadística aplicable as las medias de variedades, sus cruces y poblaciones afines Fitotecnica Latinoamericana 2: 11-22.) and the mean of the possible sets of lines that can be bred from these parents , according to Vencovsky and Barriga (1992Vencovsky R and Barriga P (1992) Genética biométrica no fitomelhoramento. Revista Brasileira de Genética, Ribeirão Preto, 496p.).
Genetic analysis was performed using the software package Genes (Cruz 2006Cruz CD (2006) Programa Genes: Biometria. Editora UFV, Viçosa, 382p.).
RESULTS AND DISCUSSION
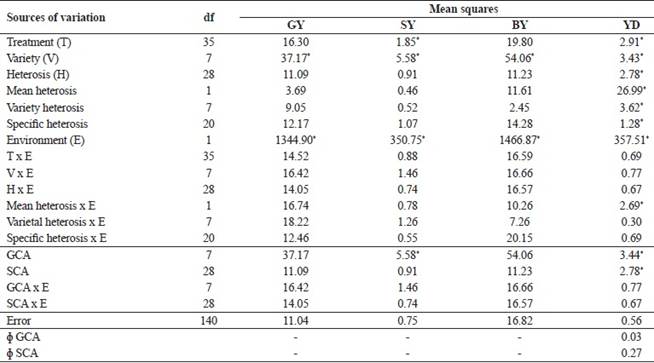
The parent effect was significant (p <0.05) for YD (Table 2), indicating phenotypic variability in yield between the parents. For the F1 hybrids, only SY and YD had significant effects. There was no significant interaction between parents and environments, nor between hybrids and environments. Thus, the genotype performance in crop cultivation did not differ, allowing the selection of the best genotypes based on the annual means.
The effect of S1 generations was not significant for the variables (Table 2), indicating no differential effect on the studied genotypes for these traits. However, for GY and SY, the significance of the interaction between S1 generations and environments showed a different behavior of genotypes in the two seasons.
The absence of significant contrast between parent and F1 combinations (Table 2) indicated that the mean hybrid behavior was not different from the mean behavior of the parent, raising doubts about the feasibility of breeding modern hybrids with high performance in the desirable industrial traits. However, it was noted that the significance of interaction between parents versus F1 and environments for YD indicated that the difference in yield performance between parents and F1 combinations may vary according to the growing season.
Resende and Duarte (2007Resende MDV and Duarte JB (2007) Precisão e controle de qualidade em experimentos de avaliação de cultivares. Pesquisa Agropecuária Tropical37: 182-194.) used selective accuracy ( ) as a parameter to assess the quality of cultivar evaluation experiments. According to the same authors, the estimated values of selective accuracy for GY and YD could be classified in the category of high experimental precision, while the estimated values for SY and BY fall into the category of moderate accuracy (Table 2).
) as a parameter to assess the quality of cultivar evaluation experiments. According to the same authors, the estimated values of selective accuracy for GY and YD could be classified in the category of high experimental precision, while the estimated values for SY and BY fall into the category of moderate accuracy (Table 2).
In the S1 generation in both years, the mean YD of the genotypes was lowest (Table 3), due to the manifestation of the effects of deleterious alleles caused by inbreeding. A hybrid combination that stood out from the others for GY in the second year was IAC Nelore x HT 932, whereas in the first year there were no outstanding combinations.
The absence of heterotic significance for the traits GY, SY and BY (Table 4) allowed the conclusion that the dominance effects were not relevant for the gene expression of these traits. However for YD, the participation of these effects in the trait expression was evident.
The effect of varieties was significant for all traits (Table 4), allowing the conclusion that additive effects were important in trait expression. The interaction of the variety with environment effect was non-significant for all traits, indicating that the effect can be evaluated based on the annual means.
The significance of variety heterosis and specific heterosis for YD only (Table 4) indicated that the parents did not differ in their respective mean gene frequencies and had the same dispersion of gene frequencies for GY, SY and BY. Moreover, the significance of specific heterosis effects on YD indicated high frequencies of gene loci with dominance of the parents.
The parents IPR 127 and HT 932 were mainly responsible for the increase in GY, based on their estimates of variety effect ( ) (Table 5), resulting in negative estimates for SY and BY, which is desirable. Thus, both parents can be recommended to increase the corn gritting ability.
) (Table 5), resulting in negative estimates for SY and BY, which is desirable. Thus, both parents can be recommended to increase the corn gritting ability.
The YD data indicated the possibility of selection of superior parents, both for breeding of lines (based on ) as well as for crosses, exploiting the heterotic effect of the parent ( ). We recommend the parents HT 5392, IAC Nelore, HT 9332, and HD 332 to breed lines with high YD (Table 5). If the target is the development of hybrid from these parents, the genotypes IPR 127, HT 392 and IAC Nelore become the most recommendable, in view of their positive values. However, considering simultaneous gains in canjica and grain yield, no parents could be identified that would meet this condition perfectly for line breeding. For crosses and hybrids from the parents, based on , parent IAC Nelore can be recommended, due to the high variety heterosis.
). We recommend the parents HT 5392, IAC Nelore, HT 9332, and HD 332 to breed lines with high YD (Table 5). If the target is the development of hybrid from these parents, the genotypes IPR 127, HT 392 and IAC Nelore become the most recommendable, in view of their positive values. However, considering simultaneous gains in canjica and grain yield, no parents could be identified that would meet this condition perfectly for line breeding. For crosses and hybrids from the parents, based on , parent IAC Nelore can be recommended, due to the high variety heterosis.
 for grit yield (GY), small grit yield (SY), bran yield (BY) and grain yield data (YD) and the varietal heterosis effect
for grit yield (GY), small grit yield (SY), bran yield (BY) and grain yield data (YD) and the varietal heterosis effect  for grain yield (YD), and the general combining ability (
for grain yield (YD), and the general combining ability (The results of the combined diallel analysis according to the Griffing's Method 2, model I (fixed effects) (1956Griffing B (1956) Concept of general and specific combining ability in relation to diallel crossing systems. Australian Journal Biology Science 9: 463-493.) are in Table 4, showing that the GCA effects were only significant for SY and YD (p <0.05), while the SCA effects were significant only for YD. No statistical significance was detected for any interactions between GCA and SCA estimates and the environmental effect (growing seasons). Therefore, the analysis of significant estimates was carried out on the basis of biannual means. The quadratic components indicated a predominance of non-additive over additive effects for YD.
A longer period of time is probably required to efficiently define the predominance of gene effects on the inheritance of the variables GY, SY and BY. Thus, a greater number of years of experimentation would be necessary, although these traits are potentially highly correlated with YD. One possible improvement strategy would be to breed superior lines for subsequent crosses and analysis of the resulting hybrids, so that the lines should be derived from hybrids with parents with high , of which at least one has a relatively high GCA. In addition, it is important that these hybrids have a high and large deviation from the mean expected for this trait.
The parents with estimates of positive effects of GCA ( ) for YD were HD 332, HT 9332, HT 5392, and IAC Nelore (Table 5). The latter two seem the most suitable for intrapopulational breeding programs, for having a higher concentration of alleles with additive effects (Cruz et al. 2012Cruz CD, Regazzi AJ and Carneiro PCS (2012) Modelos biométricos aplicados ao melhoramento genético., Editora UFV, Viçosa 514p.).
) for YD were HD 332, HT 9332, HT 5392, and IAC Nelore (Table 5). The latter two seem the most suitable for intrapopulational breeding programs, for having a higher concentration of alleles with additive effects (Cruz et al. 2012Cruz CD, Regazzi AJ and Carneiro PCS (2012) Modelos biométricos aplicados ao melhoramento genético., Editora UFV, Viçosa 514p.).
According to Cruz et al. (2012Cruz CD, Regazzi AJ and Carneiro PCS (2012) Modelos biométricos aplicados ao melhoramento genético., Editora UFV, Viçosa 514p.), the most promising hybrid combinations must be selected by the most favorable  estimates for the trait in question, but with at least one parent that has superior effects of GCA. Thus, the hybrid combinations HT 9332 x HT 392 (0.278), IAC Nelore x HT 9332 (0.315), IAC Nelore x HT 932 (0.323), and IPR 119 x IAC Nelore (0.573) are the most recommended for reciprocal recurrent selection programs targeting higher YD, in which one parent A acts as the tester of a population B, so that genotypes with a significant genetic complementation of each other are selected.
estimates for the trait in question, but with at least one parent that has superior effects of GCA. Thus, the hybrid combinations HT 9332 x HT 392 (0.278), IAC Nelore x HT 9332 (0.315), IAC Nelore x HT 932 (0.323), and IPR 119 x IAC Nelore (0.573) are the most recommended for reciprocal recurrent selection programs targeting higher YD, in which one parent A acts as the tester of a population B, so that genotypes with a significant genetic complementation of each other are selected.
For trait SY, parents with negative estimates were HT 932, IAC Nelore and IPR 127 (Table 5), recommended for a reduction in SY in intrapopulational breeding programs.
In the mean, all estimates of inbreeding depression (ID) for YD and BY were positive. This indicated that the means tend to decrease over the selfing generations, which is expected due to the expression of effects of deleterious alleles caused by inbreeding.
With regard to YD, the annual means of the percentage of ID in each parent ranged from 40.87 (IAC Nelore) to 62.29% (IPR 127) (Table 6). The differences between the ID values of the parents were relatively modest, most likely because the genotypes studied had already undergone one or more selection cycles.
Estimates of inbreeding depression (ID), in percentage, and the potential of hybrids as sources of inbred lines
for the genotypes used in diallel crosses, in each growing season, for grit yield (GY) and small grit yield (SY), and the mean of both growing seasons for bran yield (BY) and grain yield (YD)
The lowest ID value for YD was found for the intervarietal hybrid IAC Nelore (Table 6). Due to its genetic structure, this genotype has greater yield stability, increasing expectations of the possibility of breeding of lines with high frequency of favorable alleles. For YD, the results of also show that higher-yielding lines could be bred from IAC Nelore, due to the higher frequency of favorable alleles. The negative estimates of may result from estimation errors contained in or from limitations of the genetic model, insofar as it disregards epistatic effects (Vencovsky and Barriga 1992Vencovsky R and Barriga P (1992) Genética biométrica no fitomelhoramento. Revista Brasileira de Genética, Ribeirão Preto, 496p.). To obtain results with positive values, it was inferred that ID should not exceed 50% after only one selfing.
With the estimates of of the first year, the parents most recommended for breeding of lines with high GY were IPR 127 and HT 932. In the second year, IPR 119 and HT 932 stood out. For trait SY, the parents most recommended for breeding of lines with low SY in the first year were HT 932 and HT 5392, and in the second year, IPR 119 and HT 392. The lines with lowest BY would be bred from the parents IPR 127, IAC Nelore and HT 932.
Although predominance of additive effects for the studied industrial traits and of non- additive effects for YD seems likely, further studies will be required to check the possibility of increased corn gritting ability, especially with regard to its relationship with YD over several years.
REFERENCES
- Castro MVL, Naves MMV, Oliveira JP and Oliveira Froes L (2009) Rendimento industrial e composição química de milho de alta qualidade proteica em relação a híbridos comerciais. Pesquisa Agropecuária Tropical 39: 233-242.
- Cruz CD (2006) Programa Genes: Biometria. Editora UFV, Viçosa, 382p.
- Cruz CD, Regazzi AJ and Carneiro PCS (2012) Modelos biométricos aplicados ao melhoramento genético., Editora UFV, Viçosa 514p.
- Galvão JCC and Miranda GV (2004) Tecnologias de produção de milho., Editora UFV, Viçosa 366p.
- Gardner CO (1965) Teoria de genética estadística aplicable as las medias de variedades, sus cruces y poblaciones afines Fitotecnica Latinoamericana 2: 11-22.
- Gardner CO and Eberhart SA (1966) Analysis and interpretation of the variety cross diallel and related populations. Biometrics 22: 439-452.
- Gonçalves RA, Santos JP, Tomé PHF, Pereira RGFA, Ascheri JLR and Abreu CMP (2003) Yield and chemical composition of corn cultivate in dry-milling and grit production. Ciência e Agrotecnologia 27: 643-650.
- Griffing B (1956) Concept of general and specific combining ability in relation to diallel crossing systems. Australian Journal Biology Science 9: 463-493.
- Resende MDV and Duarte JB (2007) Precisão e controle de qualidade em experimentos de avaliação de cultivares. Pesquisa Agropecuária Tropical37: 182-194.
- Vencovsky R and Barriga P (1992) Genética biométrica no fitomelhoramento. Revista Brasileira de Genética, Ribeirão Preto, 496p.
Publication Dates
-
Publication in this collection
Dec 2015
History
-
Received
11 Nov 2014 -
Accepted
15 Jan 2015