Abstract
Capsicum is represented by peppers and sweet peppers and comprises a group with remarkable genetic variability. Different cultivated Capsicum peppers of Brazil were evaluated by using CMA3 and DAPI specific fluorochromes. There was high polymorphism of highly GC-rich CMA heterochromatic bands among the analyzed species, ranging from six (BAGC 114; C. annuum) to 26 blocks (BAGC 81; C. baccatum). Heterochromatin percentage ranged from 3.14% (BAGC 114; C. annuum) to 8.72% (BAGC 81; C. baccatum), corroborating the variation in the number of heterochromatic bands, particularly those distributed in the terminal and subterminal regions of the chromosomes. The information reported in this paper supports the cytogenetic characterization of the domesticated peppers accessions belonging to the Capsicum Germplasm Active Bank of the Federal University of Piauí (BAGC-UFPI). Moreover, the present data helped to better understand the karyotype features of peppers and provide additional information that could contribute to the improvement and maintenance of Capsicum genetic breeding programs.
Keywords:
CMA/DAPI staining; genetic diversity; genetic resources
INTRODUCTION
Capsicum L. genus is native to Tropical and Subtropical America, comprising ~42 species represented by peppers and bell peppers, spices widely used in the worldwide cuisine (Carrizo García et al. 2016Carrizo GarcíaCBarfussMHJSehrEMBarbozaGESamuelRMosconeEAEhrendorferF2016 Phylogenetic relationships, diversification and expansion of chili peppers (Capsicum, Solanaceae). Annals of Botany 118:35-51, Barboza et al. 2019BarbozaGECarrizo GarcíaCGonzálezSLScaldaferroMReyesX2019 Four new species of Capsicum (Solanaceae) from the tropical Andes and an update on the phylogeny of the genus. PLoS ONE 14:1-26, Barboza et al. 2020aBarboza GE, Bianchetti LDB and Stehmann JR (2020a) Capsicum carassense (Solanaceae), a new species from the Brazilian Atlantic Forest. PhytoKeys 140: 125-38., Barboza et al. 2020bBarboza GE, Carrizo García C, Scaldaferro M and Bohs L (2020b) An amazing new Capsicum (Solanaceae) species from the Andean-Amazonian Piedmont. PhytoKeys 163: 13-29). The five cultivated and domesticated species of the genus are: Capsicum annuum L., C. chinense Jacq., C. frutescens L., C. baccatum L. and C. pubescens R. & P. (Ribeiro et al. 2020RibeiroCReifschneiderFCarvalhoSBianchettiBianchettiBusoG2020 Embrapa’s Capsicum breeding program-looking back… into the future. Crop Breeding, Genetics and Genomics 2:e200001).
Brazil is considered to be a secondary diversity center of peppers with domesticated, semi- domesticated and wild Capsicum species (Barboza et al. 2020a, Ribeiro et al. 2020RibeiroCReifschneiderFCarvalhoSBianchettiBianchettiBusoG2020 Embrapa’s Capsicum breeding program-looking back… into the future. Crop Breeding, Genetics and Genomics 2:e200001). Studies of genetic diversity characterization in cultivated plant species are essential for breeding programs. In this context, different methodologies have been applied to explore the genetic divergence among germplasm accessions, e.g., phenotype, biochemical, molecular, and cytogenetic characterization (Costa et al. 2019CostaGNSilvaBMPLopesACACarvalhoLCBGomesRLF2019 Selection of pepper accessions with ornamental potential. Revista Caatinga 32:566-574, Assis et al. 2020AssisRBabaVYCintraLAGonçalvesLSARodriguesRVanzelaALL2020 Genome relationships and LTR-retrotransposon diversity in three cultivated Capsicum L (Solanaceae) species .. BMC Genomics 21:1-14, Nankar et al. 2020NankarNATodorovaVTringovskaIPasevGRadeva-IvanovaVIvanovaVKostovaKostova2020 A step towards Balkan Capsicum annuum L. core collection: Phenotypic and biochemical characterization of 180 accessions for agronomic, fruit quality, and virus resistance traits. PLoS ONE 15:1-28).
Cytogenetic studies using different approaches have been providing important information about the intra- and interspecific Capsicum diversity and have contributed to the systematics, genetics, evolution, and genetic breeding of the genus (Moscone et al. 1993MosconeEALambrouMHunzikerATEhrendorferF1993 Giemsa C-banded karyotypes in Capsicum (Solanaceae). Plant Systematics and Evolution 186:213-229, Moscone et al. 1996MosconeEALambrouMEhrendorferF1996 Fluorescent chromosome banding in the cultivated species of Capsicum (Solanaceae)*. Plant Systematics and Evolution 202:37-63, 2007MosconeEAScaldaferroMAGrabieleMCecchiniNMGarcíaYSJarretRDaviñaJRDucasseDABarbozaGEEhrendorferF2007 The evolution of chili peppers (Capsicum - Solanaceae): A cytogenetic perspective. Acta Horticulturae 745:137-170, Scaldaferro et al. 2013ScaldaferroMAGrabieleMMosconeEA2013 Heterochromatin type, amount and distribution in wild species of chili peppers (Capsicum, Solanaceae). Genetic Resources and Crop Evolution 60:693-709, Scaldaferro et al. 2016ScaldaferroMACruzVRCecchiniCecchiniMosconeEA2016 FISH and AgNor mapping of the 45S and 5S rRNA genes in wild and cultivated species of Capsicum (Solanaceae). Genome 59:95-113, Martins et al. 2018MartinsLVPeronAPLopesACAGomesRFLCarvalhoRFeitozaLL2018 Heterochromatin distribution and histone modification patterns of H4K5 acetylation and phosphorylation in Capsicum L. Crop Breeding and Applied Biotechnology 18:161-168, Zhou et al. 2019ZhouHCWaminalNEKimHH2019 In silico mining and FISH mapping of a chromosome specific satellite DNA in Capsicum annuum L. Genes & Genomics 41:1001-1006). Moreover, meiotic analysis has provided essential information regarding reproduction, fertility, recombination, meiotic irregularities and gamete viability. These karyological data, together with molecular and morphoagronomical data, help breeders plan and execute intra- and interspecific crosses of Capsicum species (Pozzobon et al. 2006PozzobonMTSchifino-WittmannMTBianchettiLDB2006 Chromosome numbers in wild and semidomesticated Brazilian Capsicum L. (Solanaceae) species: do x = 12 and x = 13 represent two evolutionary lines? Botanical Journal of the Linnean Society 151:259-269, Pozzobon et al. 2015PozzobonMTBianchettiLBSantosSCarvalhoSICReifschneiderReifschneiderRibeiroCSC2015 Comportamento meiótico em acessos de Capsicum chinense Jacq. do Banco de Germoplasma da Embrapa, Brasil. Revista Brasileira de Biociências 13:96-100).
Capsicum species have basic chromosome numbers n = 12 and n = 13. Species with 2n = 2x = 24 have symmetric karyotypes, and the chromosome number n = 12 is considered to be the ancestral state of Capsicum (Carrizo García et al. 2016Carrizo GarcíaCBarfussMHJSehrEMBarbozaGESamuelRMosconeEAEhrendorferF2016 Phylogenetic relationships, diversification and expansion of chili peppers (Capsicum, Solanaceae). Annals of Botany 118:35-51). This number is present in the domesticated species C. annuum, C. chinense, C. frutescens, C. baccatum and C. pubescens. On the other hand, the 2n = 2x = 26 group is represented by wild species in South America and exhibits asymmetric karyotype formula, as found in C. campylopodium and C. mirabile (Pozzobon et al. 2006PozzobonMTSchifino-WittmannMTBianchettiLDB2006 Chromosome numbers in wild and semidomesticated Brazilian Capsicum L. (Solanaceae) species: do x = 12 and x = 13 represent two evolutionary lines? Botanical Journal of the Linnean Society 151:259-269, Moscone et al. 2007MosconeEAScaldaferroMAGrabieleMCecchiniNMGarcíaYSJarretRDaviñaJRDucasseDABarbozaGEEhrendorferF2007 The evolution of chili peppers (Capsicum - Solanaceae): A cytogenetic perspective. Acta Horticulturae 745:137-170, Barboza et al. 2020b).
The fluorochrome banding with CMA (chromomycin A3) and DAPI (4’-6-diamidino-2- phenylindole) is a classical technique widely used in cytogenetics studies to evaluate the general composition and distribution of the constitutive heterochromatin (CH) (Schweizer 1976SchweizerD1976 Reverse fluorescent chromosome banding with chromomycin and DAPI. Chromosoma 58:307-324). CMA staining reveals the GC (Guanine-Cytosine)-rich regions, while the DAPI staining reveals the AT (Adenine-Thymine)-rich regions in the chromosomes (Guerra 2000GuerraM2000 Patterns of heterochromatin distribution in plant chromosomes. Genetics and Molecular Biology 23:1029-1041).
Previous studies on Capsicum identified four types of CH by using the CMA/DAPI banding pattern: highly GC-rich and AT-reduced (CMA++/DAPI-); (2) highly AT-rich and GC-reduced (CMA-/DAPI++); (3) moderately GC-rich and AT-neutral (CMA+/DAPI0); and (4) moderately GC-rich and moderately AT-rich (CMA+/DAPI+) (Moscone et al. 2007MosconeEAScaldaferroMAGrabieleMCecchiniNMGarcíaYSJarretRDaviñaJRDucasseDABarbozaGEEhrendorferF2007 The evolution of chili peppers (Capsicum - Solanaceae): A cytogenetic perspective. Acta Horticulturae 745:137-170, Scaldaferro et al. 2013ScaldaferroMAGrabieleMMosconeEA2013 Heterochromatin type, amount and distribution in wild species of chili peppers (Capsicum, Solanaceae). Genetic Resources and Crop Evolution 60:693-709, Romero-da Cruz and Forni-Martins 2015Romero-da Cruz MV and Forni-Martins2015 Capsicum chinense DC. IAPT/IOPB chromosome data 20. Taxon 64:E33, Romero-da Cruz et al. 2017Romero-da Cruz MV, Urdampilleta JD, Forni-Martins ER and Moscone EA2017 Cytogenetic markers for the characterization of Capsicum annuum L. cultivars. Plant Biosystems 151:84-91, Barboza et al. 2019BarbozaGECarrizo GarcíaCGonzálezSLScaldaferroMReyesX2019 Four new species of Capsicum (Solanaceae) from the tropical Andes and an update on the phylogeny of the genus. PLoS ONE 14:1-26). Most of the CH bands are located in terminal and subterminal regions of the chromosomes, except in C. flexuosum and C. campylopodium, which show intercalary CH bands. In some cases, the centromeric heterochromatin is visualized as weak CMA+/DAPI0 markers in some cultivated taxa, as observed in C. chinense and C. frutescens (Moscone et al. 2007MosconeEAScaldaferroMAGrabieleMCecchiniNMGarcíaYSJarretRDaviñaJRDucasseDABarbozaGEEhrendorferF2007 The evolution of chili peppers (Capsicum - Solanaceae): A cytogenetic perspective. Acta Horticulturae 745:137-170).
Grabiele et al. (2018AssisRBabaVYCintraLAGonçalvesLSARodriguesRVanzelaALL2020 Genome relationships and LTR-retrotransposon diversity in three cultivated Capsicum L (Solanaceae) species .. BMC Genomics 21:1-14) reported a new type of satellite DNA in Capsicum, composed of inactive rDNA 18S-25S. The complete unity of rDNA 35S is amplified, disperse and organized in tandem in the genome of the species with n = 12, and it is the main component of highly GC-rich heterochromatin, except for C. recurvatum and C. rhomboideum (both with n = 13).
Because of their socioeconomic importance and aiming to preserve the genetic diversity of peppers in the country, the Federal University of Piauí has created the Capsicum Germplasm Active Bank (BAGC-UFPI). Currently, the BAGC-UFPI has more than 250 pepper accessions belonging to different Brazilian regions (Northeast, North, Southeast, Midwest and South). Moreover, UFPI performs initiative genetics pre-breeding studies in Capsicum (Sousa et al. 2015, Martins et al. 2018MartinsLVPeronAPLopesACAGomesRFLCarvalhoRFeitozaLL2018 Heterochromatin distribution and histone modification patterns of H4K5 acetylation and phosphorylation in Capsicum L. Crop Breeding and Applied Biotechnology 18:161-168, Costa et al. 2019CostaGNSilvaBMPLopesACACarvalhoLCBGomesRLF2019 Selection of pepper accessions with ornamental potential. Revista Caatinga 32:566-574).
In this context, the aim of this study was to increase knowledge of the karyotype constitution of the socioeconomic important Capsicum species. This paper described in detail the number, distribution pattern, and percentage of CH bands in the karyotype of 16 pepper accessions of different Brazilian regions (Northeast, Midwest, Southeast and South) belonging to BAGC-UFPI by CMA/DAPI banding technique. The present data contribute to better understanding the dynamics of the CH distribution pattern in Capsicum species that can probably be related to the genetic diversity observed in this economically important genus.
MATERIAL AND METHODS
Plant materials
Seeds of 16 pepper accessions were obtained from BAGC-UFPI (Table 1), located in Teresina, Piauí, Brazil. The accessions were selected on the basis of previous intra- and interspecific morphological diversity characterization study (unpublished data).
Chromosome preparation
Root tips obtained from the germinated seeds were pretreated with p-dichlorobenzene (0.015 g mL-1) for 2 hours at room temperature, fixed in solution (ethanol: acetic acid v/v) for at least 24 hours, and stored at -20 °C until use.
CMA/DAPI fluorochrome staining
The protocol described by Schweizer and Ambros (1994SchweizerDAmbrosPF1994 Chromosome banding: stain combinations for specific regions. In Gosden JR (ed) Chromosome analysis protocols. Human Press, Totowa, p. 97-112) was followed with minor modifications. For each accession, root tips were digested with an enzymatic solution containing 2% cellulase (Onozuka R-10) and 20% pectinase (Sigma-Aldrich). The slides were stained with 10 μL of CMA (0.5 mg mL-1) for 1 hour, counterstained with 10 μL of DAPI (2 mg mL-1) for 30 min, mounted in glycerol/McIlvaine (1:1) and stored for three days before analysis.
Image analyses and morphometry
The five metaphases of each accession were photographed using a DF7000GT digital camera coupled to a Leica DM4B microscope. The images were optimized for brightness and contrast using Adobe Photoshop CS3. Chromosome sizes were measured using the Drawid v0.26 software (Kirov et al. 2017KirovIKhustalevaLLaereKVSolovievAMeeusSRomanovDFesenkoI2017 DRAWID: user-friendly java software for chromosome measurements and idiogram drawing. Comparative Cytogenetics 11:747-757). Idiograms were constructed using Corel DRAW (2017), and chromosome morphologies and parameters (Table 1) were classified according to Guerra (2002GuerraM2002 Como observar cromossomos: um guia de técnicas em citogenética vegetal, animal e humana. FUNPEC, Ribeirão Preto, 131p). Heterochromatin percentage (%) of GC-rich heterochromatin of each accession and the total size of the heterochromatic blocks were compared to the total size of the chromosome set, as described by Fonsêca et al. (2010FonsêcaAFerreiraTFerreiraFerreiraJJSantosTRBMosiolekMBellucciEKamiJGeptsPGeffroyVSchweizerDSantosKGBPedrosa-HarandA2010 Cytogenetic map of common bean (Phaseolus vulgaris L.). Chromosome Research 18:487-502).
RESULTS AND DISCUSSION
Capsicum accessions showed 2n = 24 chromosomes with metacentric (M) and submetacentric (SM) morphologies (Table 1, Figures 1 and 2). There was a Solanum-like prophase condensation pattern, with early-condensed proximal regions and late-condensed terminal chromatin (Figure 2i) (Feitoza et al. 2017FeitozaLCostaLGuerraM2017 Condensation patterns of prophase/prometaphase chromosome are correlated with H4K5 histone acetylation and genomic DNA contents in plants. PLoS ONE 12:1-14). Its condensation pattern is related to the chromatin organization at interphase, being also related to the nuclei type. The semi-reticulated nuclei were identified in all analyzed accessions (Figure 2h and 2k), with simple and well-distributed chromocenters, as previously identified by Scaldaferro et al. (2016ScaldaferroMACruzVRCecchiniCecchiniMosconeEA2016 FISH and AgNor mapping of the 45S and 5S rRNA genes in wild and cultivated species of Capsicum (Solanaceae). Genome 59:95-113) and Martins et al. (2018MartinsLVPeronAPLopesACAGomesRFLCarvalhoRFeitozaLL2018 Heterochromatin distribution and histone modification patterns of H4K5 acetylation and phosphorylation in Capsicum L. Crop Breeding and Applied Biotechnology 18:161-168).
Accession, common name, scientific name, number of diploid chromosomes (2n), range of chromosomal size (RCS), Ratio between long and short arms (R) of each chromosome pair, karyotype formula (KF), total chromosomal length (TCL), chromosomal medium length (CML) and number of CMA/DAPI bands (CMA3/DAPI). ++ represents more strongly stained CMA bands, while + represents slightly stained CMA bands. 0 represents AT neutral and - AT reduced bands. Heterochromatin percentage (%)

Double staining with CMA/DAPI fluorochromes in Capsicum 81 and 114 accessions. a1 and b1 represent the chromosomes counterstained with DAPI (in blue). a2 and b2 represent the chromosomes stained with CMA. a3 and b3 represent the merge of DAPI and CMA images. C. baccatum (a1-a3) shows the higher number of CMA heterochromatic bands (26), while C. annuum (b1-b3) had the smallest number of CMA bands (six). Arrows indicate large CMA++ blocks. All inserts indicate small CMA+ blocks in a region that is difficult to detect. Bar = 10 μm.

Double staining with CMA/DAPI fluorochromes showing high polymorphism of the CMA heterochromatic bands in different analyzed Capsicum accessions. The number of each accession is indicated on the left side of the metaphases. Arrows indicate large CMA++ blocks. All inserts indicate small CMA+ blocks in a region that is difficult to detect. Bar = 10 μm.
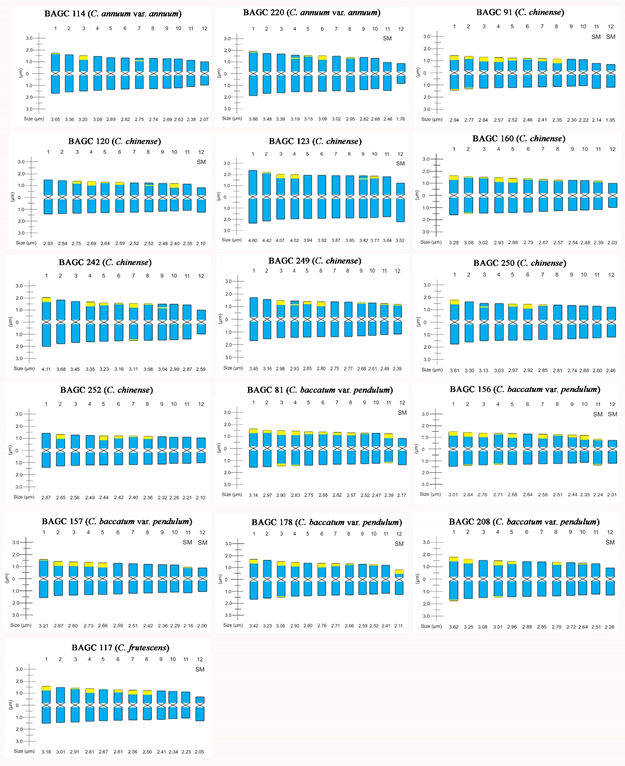
Chromosomal Size (RCS) ranged from 1.76 µm in C. annuum (BAGC 220) to 4.80 µm in C. chinense (BAGC 123), while Total Chromosomal Length (TCL) ranged from 57.61 µm in C. frutescens (BAGC 117) to 78.46 µm in C. chinense (BAGC 123) (Table 1). Variation regarding RCS and TCL in different Capsicum accessions is common and has been previously reported. Sousa et al. (2011SousaSAMMartinsKLPereiraTN2011 Polimorfismo cromossômico em Capsicum chinense Jacq. Ciência Rural 41:1777-1783) found RCS variation from 2.59 µm to 4.12 µm in C. chinense from four accessions collected in different regions of Brazil, while TCL ranged from 82.40 to 84.18 µm. Sousa et al. (2015) identified RCS variation from 3.29 µm in C. chinense to 7.48 µm in C. baccatum. In C. frutescens, the authors found TCL divergence from 105.96 to 144.44 µm. Similarly, Martins et al. (2018MartinsLVPeronAPLopesACAGomesRFLCarvalhoRFeitozaLL2018 Heterochromatin distribution and histone modification patterns of H4K5 acetylation and phosphorylation in Capsicum L. Crop Breeding and Applied Biotechnology 18:161-168) found RCS from 1.96 µm to 5.94 µm, while TCL ranged from 66.55 µm to 117.56 µm in different domesticated Capsicum accessions belonging to BACC-UFPI. These differences could be related to unequal degrees of chromosome condensation during cell division (Moscone 1990MosconeEA1990 Chromosome studies on Capsicum (Solanaceae) I. Karyotype analysis in C. chacoense. Brittonia 42:147-154, Martins et al. 2018), differences in pretreatment (Pozzobon et al. 2006PozzobonMTSchifino-WittmannMTBianchettiLDB2006 Chromosome numbers in wild and semidomesticated Brazilian Capsicum L. (Solanaceae) species: do x = 12 and x = 13 represent two evolutionary lines? Botanical Journal of the Linnean Society 151:259-269), and/or different classes of repetitive DNA sequences, such as Copia and Gypsy LTR-retrotransposons that can result in genome size variation of the species (Assis et al. 2020AssisRBabaVYCintraLAGonçalvesLSARodriguesRVanzelaALL2020 Genome relationships and LTR-retrotransposon diversity in three cultivated Capsicum L (Solanaceae) species .. BMC Genomics 21:1-14).
Three karyotype formulas were identified in the analyzed accessions: seven accessions (BAGC 81, 1117, 120, 123, 178, 208, and 220) exhibited 11M + 1SM, and six (BAGC 114, 160, 242, 249, 250, and 252) exhibited 12M (Table 1). Only three accessions (BAGC 91, 156, and 157) showed 10M + 2SM, which may suggest the presence of three different cytotypes of Capsicum accessions at BAGC-UFPI (Sousa et al. 2015Sousa, WRN, Lopes ACA, Carvalho R, Gomes RLF and Peron AP2015 Karyotypic characterization of Capsicum sp. accessions. Acta Scientiarum 37:147-153, Martins et al. 2018MartinsLVPeronAPLopesACAGomesRFLCarvalhoRFeitozaLL2018 Heterochromatin distribution and histone modification patterns of H4K5 acetylation and phosphorylation in Capsicum L. Crop Breeding and Applied Biotechnology 18:161-168). Further studies using more detailed cytomolecular techniques, e.g., 5S and 35S ribosomal DNA and/or transposable elements such as FISH probes, are needed to investigate the karyotype constitution of these accessions in detail.
The double-staining technique with CMA and DAPI fluorochromes allowed the identification of two heterochromatin banding patterns: CMA++/DAPI- and CMA+/DAPI0 (Figures 1 and 2). No DAPI bands were found in the present study, as opposed to previous works with wild Capsicum species (Moscone et al. 1996MosconeEA1990 Chromosome studies on Capsicum (Solanaceae) I. Karyotype analysis in C. chacoense. Brittonia 42:147-154, Moscone et al. 2007, Scaldaferro et al. 2013ScaldaferroMAGrabieleMMosconeEA2013 Heterochromatin type, amount and distribution in wild species of chili peppers (Capsicum, Solanaceae). Genetic Resources and Crop Evolution 60:693-709). DAPI bands were only found in domesticated (C. pubescens) and wild species (C. campylopodium, C. pereirae and C. praetermissum) (Moscone et al. 2007). For a better understanding of the distribution pattern of the CMA bands in the haploid set, in addition to determining the morphology and size of the chromosomes, all the accessions of the karyotype were schematically represented as idiograms (Figure 3).

Idiograms representing the size, morphology and distribution of the CMA bands (yellow bands) in the karyotypes of different Capsicum pepper accessions belonging to the BAGC-UFPI.
All the accessions showed variable number, size, distribution, and type of the heterochromatin blocks. At least two terminal CMA++/DAPI- bands were identified in all accessions, probably corresponding to the nucleolar organizer regions (NORs). The presence of this CMA++/DAPI- pair is commonly described in Capsicum species and it seems to be universal within the genus Capsicum (Moscone et al. 1996MosconeEALambrouMEhrendorferF1996 Fluorescent chromosome banding in the cultivated species of Capsicum (Solanaceae)*. Plant Systematics and Evolution 202:37-63, Martins et al. 2018MartinsLVPeronAPLopesACAGomesRFLCarvalhoRFeitozaLL2018 Heterochromatin distribution and histone modification patterns of H4K5 acetylation and phosphorylation in Capsicum L. Crop Breeding and Applied Biotechnology 18:161-168, Assis et al. 2020AssisRBabaVYCintraLAGonçalvesLSARodriguesRVanzelaALL2020 Genome relationships and LTR-retrotransposon diversity in three cultivated Capsicum L (Solanaceae) species .. BMC Genomics 21:1-14).
The accession BAGC 114 (C. annuum var. annuum) had the smallest number of CH bands, with six CMA blocks. Similar results were found by Moscone et al. (1996MosconeEA1990 Chromosome studies on Capsicum (Solanaceae) I. Karyotype analysis in C. chacoense. Brittonia 42:147-154) and Martins et al. (2018MartinsLVPeronAPLopesACAGomesRFLCarvalhoRFeitozaLL2018 Heterochromatin distribution and histone modification patterns of H4K5 acetylation and phosphorylation in Capsicum L. Crop Breeding and Applied Biotechnology 18:161-168). The authors highlighted that its species has a smaller number of and simpler heterochromatin banding patterns. On the other hand, the accession BAGC 117, commonly known as “malagueta preta” (C. frutescens), showed 12 CMA bands (Figure 2b): four CMA++/DAPI- pairs and two CMA+/DAPI0 pairs. Differently, Moscone et al. (1996) identified 18 GC-rich bands and Martins et al. (2018) found four and six GC-rich bands for its species.
According to Carrizo García et al. (2016Carrizo GarcíaCBarfussMHJSehrEMBarbozaGESamuelRMosconeEAEhrendorferF2016 Phylogenetic relationships, diversification and expansion of chili peppers (Capsicum, Solanaceae). Annals of Botany 118:35-51), C. annuum, C. chinense, and C. frutescens domesticated species and C. galapagoense wild species belong to the Annuum clade, previously known as “white-flowered” group. The species belonging to its clade are closely related regarding karyotype features, including chromosomes with small size, low DNA content, and low GC-rich heterochromatin constitution, mainly distributed at terminal regions of the chromosomes (Moscone et al. 1993MosconeEALambrouMHunzikerATEhrendorferF1993 Giemsa C-banded karyotypes in Capsicum (Solanaceae). Plant Systematics and Evolution 186:213-229, Moscone et al. 1996MosconeEALambrouMEhrendorferF1996 Fluorescent chromosome banding in the cultivated species of Capsicum (Solanaceae)*. Plant Systematics and Evolution 202:37-63, Moscone et al. 200MosconeEAScaldaferroMAGrabieleMCecchiniNMGarcíaYSJarretRDaviñaJRDucasseDABarbozaGEEhrendorferF2007 The evolution of chili peppers (Capsicum - Solanaceae): A cytogenetic perspective. Acta Horticulturae 745:137-1707).
C. chinense accessions showed a highly variable number of heterochromatic blocks. For example, the accessions BAGC 123, 250 and 252 (Figure 2c, 2d, 2n and 2o) showed a small number of bands: four CMA++/DAPI- and eight CMA+/DAPI0 in BAGC 120, and four CMA++/DAPI- and six CMA+/DAPI0 in BAGC 123 and 250, respectively, and six CMA++/DAPI- and four CMA+/DAPI0 in BAGC 252. Similar results were found by Romero-da-Cruz and Forni-Martins (2015). The authors found small terminal GC-rich bands in the small and long arms of four chromosome pairs of C. chinense.
The accessions belonging to C. annuum (BAGC 114 and 220) and C. chinense (BAGC 120, 123, 242, 249 and 250) showed an additional small pair of intercalary bands moderately rich in GC (CMA+/DAPI0). BAGC 91 and 160 (Figure 2a and 2g) are distinguished from the others because of the presence of 20 GC bands, i.e., six CMA++ and 14 CMA+ bands. In general, the CH is not homogeneous, varying quantitatively and qualitatively within and between species (Guerra 2000GuerraM2000 Patterns of heterochromatin distribution in plant chromosomes. Genetics and Molecular Biology 23:1029-1041, Roa and Guerra 2015RoaFGuerraM2015 Non-random distribution of 5S rDNA sites and its association with 45S rDNA in plant chromosomes. Cytogenetic and Genome Research 146:243249, Mate-Sucre et al. 2020Mate-Sucre Y, Costa L, Gagnon E, Lewis GP, Leitch IJ and Souza G2020 Revisiting the cytomolecular evolution of the Caesalpinia group (Leguminosae): a broad sampling reveals new correlations between cytogenetic and environmental variables. Plant Systematics and Evolution 306:1-13). Moscone et al. (2007MosconeEAScaldaferroMAGrabieleMCecchiniNMGarcíaYSJarretRDaviñaJRDucasseDABarbozaGEEhrendorferF2007 The evolution of chili peppers (Capsicum - Solanaceae): A cytogenetic perspective. Acta Horticulturae 745:137-170) found polymorphism regarding number and size of CMA bands in C. annuum cultivars that showed highly GC-rich heterochromatin, with distal and interstitial moderately GC-rich bands distribution among the cultivars.
C. baccatum var. pendulum accessions stand out for the number and variation of the heterochromatic bands. The accessions BAGC 157, 178, 208, 156 and 81 have 12, 18, 16, 24 and 26 GC-rich blocks, respectively (Figures 1 and 2). Variations of the number and brightness intensity of the blocks were previously reported for C. baccatum cytotypes. Moscone et al. (1996MosconeEALambrouMEhrendorferF1996 Fluorescent chromosome banding in the cultivated species of Capsicum (Solanaceae)*. Plant Systematics and Evolution 202:37-63) found variation from 24 to 28 GC blocks, and Aguilera et al. (2017AguileraPMDebatHJGrabieleM2017 An integrated physical map of the cultivated hot chili pepper, Capsicum baccatum var. pendulum. International Journal of Agriculture & Biology 19:455-469) identified 32 terminal CMA+ sites in C. baccatum var. pendulum chromosomes by the CMA/DA/DAPI banding technique. Similarly, Martins et al. (2018MartinsLVPeronAPLopesACAGomesRFLCarvalhoRFeitozaLL2018 Heterochromatin distribution and histone modification patterns of H4K5 acetylation and phosphorylation in Capsicum L. Crop Breeding and Applied Biotechnology 18:161-168) reported 10 to 18 CMA bands in C. baccatum var. pendulum accessions belonging to the same BAGC-UFPI germplasm bank. Differently from other domesticated pepper species, its species are differentiated by a larger karyotype length (and larger DNA genome, 3.2 Gb), greater presence of GC-rich heterochromatin and a more complex heterochromatic band pattern (Moscone et al. 2007, Grabiele et al. 2014GrabieleMDebatHJAguileraPMDebatHJForni-MartinsERMartíDA2014 Cytogenetic characterization of the germplasm of wild chili peppers: Capsicum baccatum var. praetermissum DC. IAPT/IOPB chromosome data 18. Taxon: 63:E6, Grabiele et al. 2018, Kim et al. 2017KimSParkJYeomSKimY-MSeoEKi-TaeKMyung-ShinKLeeJMCheongKShinHSKimSBHanHLeeJParkMLeeHALeeHYLeeYOhSLeeJHChoiEChoiELeeSEJeonJKimHChoiGSongHLeeJLeeSCKwonJKLeeHYKooNHongYKimRWKangWHHuhJHKangBCYangTJLeeYHBennetzenJLChoiD2017 New reference genome sequences of hot pepper reveal the massive evolution of plant disease resistance genes by retroduplication. Genome Biology 18:1-11, Assis et al. 2020AssisRBabaVYCintraLAGonçalvesLSARodriguesRVanzelaALL2020 Genome relationships and LTR-retrotransposon diversity in three cultivated Capsicum L (Solanaceae) species .. BMC Genomics 21:1-14).
Regarding the percentage of GC-rich heterochromatin (Table 1), there was variation from 3.14 - 4.29% in C. annuum var. annuum (BAGC 114 and 220, respectively) to 5.65 - 8.72% in C. baccatum var. pendulum (BAGC 157 and 81, respectively). In C. chinense, the percentage ranged from 4.36% (BAGC 120) to 8.53% (BAGC 91), while in C. frutescens (BAGC 117), it was found that CH composes 8.57% of the total genome.
There is an association between the number and percentage of the CH blocks, despite minor divergences. These differences are probably related to the evolutionary dynamics of the DNA sequences of CH constitution that play an important role in the karyotype evolution of Capsicum (Scaldaferro et al. 2013ScaldaferroMAGrabieleMMosconeEA2013 Heterochromatin type, amount and distribution in wild species of chili peppers (Capsicum, Solanaceae). Genetic Resources and Crop Evolution 60:693-709). The accessions with smaller and higher % of CH were the same with smaller and higher CMA banding blocks, namely BAGC 81 (C. annuum var. annuum) and BAGC 81 C. baccatum var. pendulum, respectively.
Our results corroborate the variation in CH content (1.72% to 38.91%) previously found within and among species, with an average value of 10.90% (Moscone et al. 1996MosconeEALambrouMEhrendorferF1996 Fluorescent chromosome banding in the cultivated species of Capsicum (Solanaceae)*. Plant Systematics and Evolution 202:37-63, Moscone et al. 2003MosconeEABaranyiMEbertIGreilhuberJEhrendorferFHunzikerAT2003 Analysis of nuclear DNA content in Capsicum (Solanaceae) by flow cytometry and feulgen densitometry. Annals of Botany 92:21-29, Moscone et al. 2007, Scaldaferro et al. 2013ScaldaferroMAGrabieleMMosconeEA2013 Heterochromatin type, amount and distribution in wild species of chili peppers (Capsicum, Solanaceae). Genetic Resources and Crop Evolution 60:693-709). In most of the analyzed taxa, there is a positive association between karyotype size and CH amount, indicating that its heterochromatin contributes to the differences found in the chromosome size and probably in the genome size of Capsicum (Moscone et al. 1996MosconeEALambrouMEhrendorferF1996 Fluorescent chromosome banding in the cultivated species of Capsicum (Solanaceae)*. Plant Systematics and Evolution 202:37-63, Moscone et al. 2007, Scaldaferro et al. 2013, Scaldaferro et al. 2016, Martins et al. 2018MartinsLVPeronAPLopesACAGomesRFLCarvalhoRFeitozaLL2018 Heterochromatin distribution and histone modification patterns of H4K5 acetylation and phosphorylation in Capsicum L. Crop Breeding and Applied Biotechnology 18:161-168). A great variation in genome size is common in plants, probably owing to the large fraction of repetitive DNA found in their genomes. In Capsicum, a genus with species of large genome size, this variation could be explained by the large accumulation of transposable elements (Park et al. 2012ParkMJoSHKnownJ-KParkJAhnJHKimSLeeY-HYangT-JHurC-GKangB-CKimB-DChoiD2012 Comparative analysis of pepper and tomato reveals euchromatin expansion of pepper genome caused by differential accumulation of Ty3/Gypsy-like elements. BMC Genomics 12:1-13).
Previous genetic diversity studies using different approaches (morphological, agronomic, cytogenetic, and molecular) have provided additional data about the general genomic/phenotypic features of the species. Cytogenetics is an important tool for understanding pepper karyotypes, as well as serving as a basis for conservation activities and applied research in genetic breeding (Grabiele et al. 2018GrabieleMDebatHJScaldaferroMAAguileraPMMosconeEASeijoJGDucasseDA2018 Highly GC-rich heterochromatin in chili peppers (Capsicum-Solanaceae): A cytogenetic and molecular characterization. Scientia Horticulturae 238:391-399, Costa et al. 2019CostaGNSilvaBMPLopesACACarvalhoLCBGomesRLF2019 Selection of pepper accessions with ornamental potential. Revista Caatinga 32:566-574, Assis et al. 2020AssisRBabaVYCintraLAGonçalvesLSARodriguesRVanzelaALL2020 Genome relationships and LTR-retrotransposon diversity in three cultivated Capsicum L (Solanaceae) species .. BMC Genomics 21:1-14). The CMA/DAPI technique identified the presence and high variation of GC-rich CH in all the analyzed pepper accessions. Polymorphisms of the heterochromatic blocks could be confirmed within and among Capsicum domesticated species.
A general pattern was found for CMA marks, particularly located at terminal regions of the chromosomes, with a small number of marks occurring in the intercalary regions of C. annuum and C. chinense chromosomes. The additional information generated in this study will contribute to a better characterization and understating of karyotype polymorphisms of the Brazilian pepper domesticated accessions belonging to BAGC-UFPI. Moreover, these data will provide additional information that can help the genetic breeding programs of Capsicum species. Capsicum genetic variation is the main support for the genetic breeding program of the genus. Additionally, cytogenetics characterization and molecular and morphological studies are essential for segregated population management and the development of strategies for conservation of pepper germplasms.
ACKNOWLEDGMENTS
We are thankful to the Brazilian agencies CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) and FAPEPI (Fundação de Amparo à Pesquisa do Estado do Piauí) for the scholarships. We would also like to thank MCTI/CNPQ/Universal, project number 457201/2014-2 for providing financial support.
REFERENCES
- AssisRBabaVYCintraLAGonçalvesLSARodriguesRVanzelaALL2020 Genome relationships and LTR-retrotransposon diversity in three cultivated Capsicum L (Solanaceae) species .. BMC Genomics 21:1-14
- AguileraPMDebatHJGrabieleM2017 An integrated physical map of the cultivated hot chili pepper, Capsicum baccatum var. pendulum. International Journal of Agriculture & Biology 19:455-469
- BarbozaGECarrizo GarcíaCGonzálezSLScaldaferroMReyesX2019 Four new species of Capsicum (Solanaceae) from the tropical Andes and an update on the phylogeny of the genus. PLoS ONE 14:1-26
- Barboza GE, Bianchetti LDB and Stehmann JR (2020a) Capsicum carassense (Solanaceae), a new species from the Brazilian Atlantic Forest. PhytoKeys 140: 125-38.
- Barboza GE, Carrizo García C, Scaldaferro M and Bohs L (2020b) An amazing new Capsicum (Solanaceae) species from the Andean-Amazonian Piedmont. PhytoKeys 163: 13-29
- Carrizo GarcíaCBarfussMHJSehrEMBarbozaGESamuelRMosconeEAEhrendorferF2016 Phylogenetic relationships, diversification and expansion of chili peppers (Capsicum, Solanaceae). Annals of Botany 118:35-51
- CostaGNSilvaBMPLopesACACarvalhoLCBGomesRLF2019 Selection of pepper accessions with ornamental potential. Revista Caatinga 32:566-574
- FeitozaLCostaLGuerraM2017 Condensation patterns of prophase/prometaphase chromosome are correlated with H4K5 histone acetylation and genomic DNA contents in plants. PLoS ONE 12:1-14
- FonsêcaAFerreiraTFerreiraFerreiraJJSantosTRBMosiolekMBellucciEKamiJGeptsPGeffroyVSchweizerDSantosKGBPedrosa-HarandA2010 Cytogenetic map of common bean (Phaseolus vulgaris L.). Chromosome Research 18:487-502
- GrabieleMDebatHJAguileraPMDebatHJForni-MartinsERMartíDA2014 Cytogenetic characterization of the germplasm of wild chili peppers: Capsicum baccatum var. praetermissum DC. IAPT/IOPB chromosome data 18. Taxon: 63:E6
- GrabieleMDebatHJScaldaferroMAAguileraPMMosconeEASeijoJGDucasseDA2018 Highly GC-rich heterochromatin in chili peppers (Capsicum-Solanaceae): A cytogenetic and molecular characterization. Scientia Horticulturae 238:391-399
- GuerraM2000 Patterns of heterochromatin distribution in plant chromosomes. Genetics and Molecular Biology 23:1029-1041
- GuerraM2002 Como observar cromossomos: um guia de técnicas em citogenética vegetal, animal e humana. FUNPEC, Ribeirão Preto, 131p
- KimSParkJYeomSKimY-MSeoEKi-TaeKMyung-ShinKLeeJMCheongKShinHSKimSBHanHLeeJParkMLeeHALeeHYLeeYOhSLeeJHChoiEChoiELeeSEJeonJKimHChoiGSongHLeeJLeeSCKwonJKLeeHYKooNHongYKimRWKangWHHuhJHKangBCYangTJLeeYHBennetzenJLChoiD2017 New reference genome sequences of hot pepper reveal the massive evolution of plant disease resistance genes by retroduplication. Genome Biology 18:1-11
- KirovIKhustalevaLLaereKVSolovievAMeeusSRomanovDFesenkoI2017 DRAWID: user-friendly java software for chromosome measurements and idiogram drawing. Comparative Cytogenetics 11:747-757
- MartinsLVPeronAPLopesACAGomesRFLCarvalhoRFeitozaLL2018 Heterochromatin distribution and histone modification patterns of H4K5 acetylation and phosphorylation in Capsicum L. Crop Breeding and Applied Biotechnology 18:161-168
- Mate-Sucre Y, Costa L, Gagnon E, Lewis GP, Leitch IJ and Souza G2020 Revisiting the cytomolecular evolution of the Caesalpinia group (Leguminosae): a broad sampling reveals new correlations between cytogenetic and environmental variables. Plant Systematics and Evolution 306:1-13
- MosconeEA1990 Chromosome studies on Capsicum (Solanaceae) I. Karyotype analysis in C. chacoense. Brittonia 42:147-154
- MosconeEALambrouMHunzikerATEhrendorferF1993 Giemsa C-banded karyotypes in Capsicum (Solanaceae). Plant Systematics and Evolution 186:213-229
- MosconeEALambrouMEhrendorferF1996 Fluorescent chromosome banding in the cultivated species of Capsicum (Solanaceae)*. Plant Systematics and Evolution 202:37-63
- MosconeEABaranyiMEbertIGreilhuberJEhrendorferFHunzikerAT2003 Analysis of nuclear DNA content in Capsicum (Solanaceae) by flow cytometry and feulgen densitometry. Annals of Botany 92:21-29
- MosconeEAScaldaferroMAGrabieleMCecchiniNMGarcíaYSJarretRDaviñaJRDucasseDABarbozaGEEhrendorferF2007 The evolution of chili peppers (Capsicum - Solanaceae): A cytogenetic perspective. Acta Horticulturae 745:137-170
- NankarNATodorovaVTringovskaIPasevGRadeva-IvanovaVIvanovaVKostovaKostova2020 A step towards Balkan Capsicum annuum L. core collection: Phenotypic and biochemical characterization of 180 accessions for agronomic, fruit quality, and virus resistance traits. PLoS ONE 15:1-28
- ParkMJoSHKnownJ-KParkJAhnJHKimSLeeY-HYangT-JHurC-GKangB-CKimB-DChoiD2012 Comparative analysis of pepper and tomato reveals euchromatin expansion of pepper genome caused by differential accumulation of Ty3/Gypsy-like elements. BMC Genomics 12:1-13
- PozzobonMTSchifino-WittmannMTBianchettiLDB2006 Chromosome numbers in wild and semidomesticated Brazilian Capsicum L. (Solanaceae) species: do x = 12 and x = 13 represent two evolutionary lines? Botanical Journal of the Linnean Society 151:259-269
- PozzobonMTBianchettiLBSantosSCarvalhoSICReifschneiderReifschneiderRibeiroCSC2015 Comportamento meiótico em acessos de Capsicum chinense Jacq. do Banco de Germoplasma da Embrapa, Brasil. Revista Brasileira de Biociências 13:96-100
- RibeiroCReifschneiderFCarvalhoSBianchettiBianchettiBusoG2020 Embrapa’s Capsicum breeding program-looking back… into the future. Crop Breeding, Genetics and Genomics 2:e200001
- RoaFGuerraM2015 Non-random distribution of 5S rDNA sites and its association with 45S rDNA in plant chromosomes. Cytogenetic and Genome Research 146:243249
- Romero-da Cruz MV and Forni-Martins2015 Capsicum chinense DC. IAPT/IOPB chromosome data 20. Taxon 64:E33
- Romero-da Cruz MV, Urdampilleta JD, Forni-Martins ER and Moscone EA2017 Cytogenetic markers for the characterization of Capsicum annuum L. cultivars. Plant Biosystems 151:84-91
- ScaldaferroMAGrabieleMMosconeEA2013 Heterochromatin type, amount and distribution in wild species of chili peppers (Capsicum, Solanaceae). Genetic Resources and Crop Evolution 60:693-709
- ScaldaferroMACruzVRCecchiniCecchiniMosconeEA2016 FISH and AgNor mapping of the 45S and 5S rRNA genes in wild and cultivated species of Capsicum (Solanaceae). Genome 59:95-113
- SchweizerD1976 Reverse fluorescent chromosome banding with chromomycin and DAPI. Chromosoma 58:307-324
- SchweizerDAmbrosPF1994 Chromosome banding: stain combinations for specific regions. In Gosden JR (ed) Chromosome analysis protocols. Human Press, Totowa, p. 97-112
- SousaSAMMartinsKLPereiraTN2011 Polimorfismo cromossômico em Capsicum chinense Jacq. Ciência Rural 41:1777-1783
- Sousa, WRN, Lopes ACA, Carvalho R, Gomes RLF and Peron AP2015 Karyotypic characterization of Capsicum sp. accessions. Acta Scientiarum 37:147-153
- ZhouHCWaminalNEKimHH2019 In silico mining and FISH mapping of a chromosome specific satellite DNA in Capsicum annuum L. Genes & Genomics 41:1001-1006
Publication Dates
-
Publication in this collection
14 Feb 2022 -
Date of issue
2022
History
-
Received
07 July 2021 -
Accepted
11 Nov 2021 -
Published
20 Feb 2022