Abstract
The objective of this study was to analyze the morphofunctional attributes of Mixed Ombrophyllous Forest species of different successional categories. Fruits were collected from forest remnants in Curitiba, Paraná. It was analyzed the biometrics of seeds and seedlings of all species and the following ratios were estimated: shoot/seedling length, shoot/seedling mass, cotyledons/seedling mass, eophyll/seedling mass and seedling/ seed mass. Statistical differences among sucessional groups were observed in many of the evaluated traits. The variability among species is mainly explained by seedling size, tissue investment for light interception and shoot resource allocation. Late secondary species have larger seedlings with characteristics that indicate slower initial development, dependent on seed reserves. The pioneers species have smaller seedlings but faster development and earlier dependence on photosynthesis. These morphological and functional differences can result in different strategies for the establishment of seedlings in the successional process. The cluster analysis results on formation of two major groups. The first group is comprised of the late secondary species. The second group includes two distinct subgroups that separate the pioneers, and most of the early secondary species. These results indicate that the number and quality of available data should be considered in the definition of successional categories adopted in ecological studies.
Key words:
araucaria forest; forest ecology; regeneration dynamics; seedling ecology
Resumo
O objetivo deste estudo foi analisar os atributos morfofuncionais das espécies da Floresta Ombrófila Mista, pertencentes a diferentes categorias sucessoriais. Frutos foram coletados de remanescentes florestais em Curitiba, Paraná. Foi realizada a biometria das sementes e plântulas e estimados os seguintes índices: comprimento da parte aérea/plântula; massa da parte aérea/plântula; massa dos cotilédones/plântula; massa dos eofilos/plântula e massa da plântula/semente. Diferenças estatísticas entre os grupos foram observadas em muitas das características avaliadas. A variabilidade entre as espécies é explicada pelo tamanho das plântulas, investimento em tecidos para interceptação da luz e alocação de recursos na parte aérea. As secundárias tardias têm plântulas maiores com características que indicam um desenvolvimento inicial mais lento, dependente das reservas da semente. As pioneiras têm plântulas menores, desenvolvimento mais rápido e são dependentes da fotossíntese. Essas diferenças morfofuncionais resultam em diferentes estratégias de estabelecimento das plântulas no processo sucessional. A análise de agrupamento resultou na formação de dois grupos principais: o primeiro composto pelas espécies secundárias tardias e o segundo formado por dois subgrupos distintos, que separam as pioneiras e a maioria das secundárias iniciais. Estes resultados indicam que o número e a qualidade dos dados disponíveis devem ser considerados na definição de categorias sucessoriais adotadas em estudos ecológicos.
Palavras-chave:
dinâmica de regeneração; ecologia de plântulas; ecologia florestal; floresta com araucária
Introduction
Environmental heterogeneity combined with variable responses of plants to abiotic (humidity, light, temperature, water and nutrient availability) and biotic (competition, herbivory and facilitation) factors implies the differentiation of ecological niches. Different species occupy distinct niches with spatial and temporal dimensions, overlapping or not, according to their adaptive response patterns (Giller 1984Giller PS (1984) Community structure and the niche. Chapman and Hall, Londan, New York. 176p.; Poorter 2007Poorter L (2007) Are species adapted to their regeneration niche, adult niche, or both? The American Naturalist 169: 433-442.; Kitajima & Poorter 2008Kitajima K & Poorter L (2008) Functional basis for resource niche partitioning by tropical trees. In: Carson WP & Schnitzer SA (eds.) Tropical forest community ecology. Blackwell Science, Oxford. Pp. 160-181.).
In ecological succession, changes in the composition and structure of plant communities over time influence the distribution of niches. Species occupy different positions in this process, according to their responses to environmental features and their ability to disperse and become established (Bazzaz 1979Bazzaz FA (1979) The Physiological ecology of plant succession. Annual Review of Ecology and Systematics 10: 351-371.; Egerton 2009Egerton FN (2009) Homage to Frederic E. Clements, historian of plant succession studies. Bulletin of the Ecological Society of America 90: 43-79.). Therefore, the species can be classified into successional groups, according to the plant communities stages in which they are established (Piña-Rodrigues et al. 1990Pina-Rodrigues FCM, Costa LGS & Reis A (1990) Estratégias de estabelecimento de espécies arbóreas e o manejo de florestas tropicais. In: Anais do 6º Congresso Florestal Brasileiro. Campos do Jordão. Pp. 676-684.; Rees et al. 2010Rees M, Condit R, Crawley M, Pacala S & Tilman D (2010) Long-term studies of vegetation dynamics. Science 293: 650-655.).
There are some general trends of correlations between seed dispersal/seedling establishment characteristics and the successional stages. In the early stages of succession, species with dry fruits and numerous light, small, anemochorous seeds prevail. The seedlings are dependent on photosynthesis and tend to have relatively rapid development. Such species colonize gaps or open areas and are regarded as “pioneers” (Piña- Rodrigues et al. 1990Pina-Rodrigues FCM, Costa LGS & Reis A (1990) Estratégias de estabelecimento de espécies arbóreas e o manejo de florestas tropicais. In: Anais do 6º Congresso Florestal Brasileiro. Campos do Jordão. Pp. 676-684.; Gogosz et al. 2015Gogosz AM, Boeger MRT, Cosmo NL & Nogueira AC (2015) Morfologia de diásporos e plântulas de espécies arbóreas da floresta com araucária, no sul do Brasil. Floresta 45: 819- 832.).
In later successional stages, species with fleshy fruits, zoochorous dispersal, with few seeds are predominant. The seeds are larger and possess a reserve tissue. The seedlings, in turn, have slower development and are dependent on reserves present in the seeds and/or cotyledons. These species grow in shaded environments, and are considered “late secondary species” (Primack 1990Primack RB (1990) Seed physiology, seed germination and seedling ecology - commentary. In: Bawa KS & Hadley M (eds.) Reproductive ecology of tropical forest plants. Unesco, Parthenon, Paris. Pp. 233-236.; Piña-Rodrigues et al. 1990Pina-Rodrigues FCM, Costa LGS & Reis A (1990) Estratégias de estabelecimento de espécies arbóreas e o manejo de florestas tropicais. In: Anais do 6º Congresso Florestal Brasileiro. Campos do Jordão. Pp. 676-684.; Garwood 1996Garwood MD (1996) Functional morphology of tropical tree seedlings. In: Swaine MD (eds.) Ecology of tropical forest tree seedlings. Unesco, Parthenon, Paris. Pp. 59-129.; Ibarra-Manrìquez et al. 2001Ibarra-Manríquez G, Ramos MM & Oyama K (2001) Seedling functional types in a lowland rain forest in Mexico. American Journal of Botany 88: 1801- 1812.; Rees et al. 2010Rees M, Condit R, Crawley M, Pacala S & Tilman D (2010) Long-term studies of vegetation dynamics. Science 293: 650-655.; Gogosz et al. 2015Gogosz AM, Boeger MRT, Cosmo NL & Nogueira AC (2015) Morfologia de diásporos e plântulas de espécies arbóreas da floresta com araucária, no sul do Brasil. Floresta 45: 819- 832.). There are also species whose seed dispersal and seedling growth are intermediate, classified as “early secondary species” (Piña-Rodrigues et al. 1990Pina-Rodrigues FCM, Costa LGS & Reis A (1990) Estratégias de estabelecimento de espécies arbóreas e o manejo de florestas tropicais. In: Anais do 6º Congresso Florestal Brasileiro. Campos do Jordão. Pp. 676-684.; Gogosz et al. 2015Gogosz AM, Boeger MRT, Cosmo NL & Nogueira AC (2015) Morfologia de diásporos e plântulas de espécies arbóreas da floresta com araucária, no sul do Brasil. Floresta 45: 819- 832.).
Therefore, the study of functional traits of plants during their early development contributes to the understanding of the complex process of plant communities succession. Functional traits are plant attributes that enhance establishment, growth, survival, reproduction, and, ultimately, plant fitness (Poorter 2007Poorter L (2007) Are species adapted to their regeneration niche, adult niche, or both? The American Naturalist 169: 433-442.). Several studies have related morphological characteristics of seeds and seedlings to the ecological factors of the environment in which they develop (Burger 1972Burger HD (1972) Seedlings of some tropical trees and shrubs mainly of South East Asia. PUDOC, Wageningen. 399p.; Vogel 1980Vogel EF (1980) Seedlings of dicotyledons: structure, development, types: descriptions of 150 woody Malesian taxa. Centre for Publishing and Documentation, Wageningen. Pp. 26-48.; Kitajima 1996Kitajima K (1996) Cotyledon functional morphology and patterns of seed reserve utilization by neotropical tree seedlings. In: Swaine MD (ed.) Ecology of tropical forest tree seedling. Unesco, Parthenon, Paris. Pp. 193-210.; Wright et al. 2000Wright IJ, Clifford HT, Kidson R, Reed ML, Rice BL & Westoby MA (2000) Survey of seed and seedling characters in 1744 Australian dicotyledon species: cross-species trait correlations and correlated trait- shifts within evolutionary lineages. Biological Journal of the Linnean Society 69: 521-547.; Ibarra-Manríquez et al. 2001Ibarra-Manríquez G, Ramos MM & Oyama K (2001) Seedling functional types in a lowland rain forest in Mexico. American Journal of Botany 88: 1801- 1812.; Kitajima 2002Kitajima K (2002) Do shade-tolerant tropical tree seedlings depend longer on seed reserves? Functional growth analysis of three Bignoniaceae species. Functional Ecology 16: 433-444., 2003Kitajima K (2003) Impact of cotyledon and leaf removal on seedling survival in three tree species with contrasting cotyledon functions. Biotropica 35: 429-434.; Ressel et al. 2004Ressel K, Guilherme FAG, Schiavini I & Oliveira PE (2004) Ecologia morfofuncional de plântulas de espécies arbóreas da Estação Ecológica do Panga, Uberlândia, Minas Gerais. Revista Brasileira de Botânica 27: 311-323.) and the successional process (Foster & Janson 1985Foster S & Janson CH (1985) The relationship between seed size and establishment conditions in tropical woody plants. Ecology 66: 773-780.; Luttge 1997Lüttge U (1997) Physiological ecology of tropical plants. Springer, New York. Pp. 103-148.; Souza & Vàlio 2003Souza RP & Vàlio IFM (2003) Seedling growth of fifteen Brazilian tropical tree species differing in successional status. Revista Brasileira de Botânica 26: 35-47.; Ressel et al. 2004Ressel K, Guilherme FAG, Schiavini I & Oliveira PE (2004) Ecologia morfofuncional de plântulas de espécies arbóreas da Estação Ecológica do Panga, Uberlândia, Minas Gerais. Revista Brasileira de Botânica 27: 311-323.; Gogosz et al. 2015Gogosz AM, Boeger MRT, Cosmo NL & Nogueira AC (2015) Morfologia de diásporos e plântulas de espécies arbóreas da floresta com araucária, no sul do Brasil. Floresta 45: 819- 832.).
Among the vegetation types present in southern Brazil, Mixed Ombrophyllous Forest (Araucaria Forest) stands out due to its imposing vegetation and rich biodiversity. Nevertheless, during the process of territorial occupation and agricultural expansion, this phytogeographical unit was sorely devastated. Currently there are few, fragmented and degraded secondary forests remnants (Ipardes 2014Ipardes - Instituto Paranaense de Desenvolvimento Econômico e Social (2014) Base ambiental: Unidades de Conservação e remanescentes vegetais - Paraná. Disponível em <http://www.ipardes.gov.br/index.php>. Acesso em 13 outubro 2016.
http://www.ipardes.gov.br/index.php...
). To recover and conserving these forests is a major challenge. For this it is necessary understand their wide biological diversity and the ecological interactions. In this context, it is essential to study the early stages of plants, especially tree species, with potential for regeneration of forest areas.
The adaptive responses of plants to environmental factors in early development can result in morphological and functional differences in seedling establishment. Therefore, the study of these responses generates knowledge about the autecology of the species and provides information about the ecological successional process of plant communities, contributing to complement existing successional classifications. Thus, the objective of this study was to analyze the morphofunctional attributes of Mixed Ombrophyllous Forest species, belonging to different successional categories, to investigate: i. if the use of morphofunctional attributes of seeds and seedlings can be useful for a robust classification of species in successional categories; ii. what are the proper attributes to base as successional categories.
Material and Methods
Plant material
Plant samples were collected between 2009 and 2011 in remnants of Araucaria forest in the metropolitan region of Curitiba, state of Paraná, Brazil (Tab. 1). Araucaria forest is a typical phytogeographical unit in southern Brazil, and occurs in the humid subtropical climate region, with an annual average temperature of 18 ºC and winter averages below 15 °C (Brazão & Santos 1997Brazão JEM & Santos MM (1997) Vegetação. In: IBGE (ed.) Recursos naturais e meio ambiente: uma visão do Brasil. IBGE, Rio de Janeiro. Pp. 111-124.). The predominant soils are oxisols, argissols and cambisols (Embrapa 1999Embrapa (1999) Centro Nacional de Pesquisa de Solos. Sistema brasileiro de classificação de solos. Embrapa Produção de Informações, Brasília; Embrapa Solos, Rio de Janeiro. 412p.).
Sucessional classification
The species were previously divided in successional groups, based on Gogosz et al. (2015)Gogosz AM, Boeger MRT, Cosmo NL & Nogueira AC (2015) Morfologia de diásporos e plântulas de espécies arbóreas da floresta com araucária, no sul do Brasil. Floresta 45: 819- 832., Pina-Rodrigues et al. (1999), which classified the same set of species from the qualitative characteristics of fruits, seeds, autoecological features (dispersal mode, response to light, fruit and seed morphology, longevity etc.). Three sucessional categories were considered: pioneers, early secondary and late secondary species (Tab. 1).
Methods
Ripe fruits of selected species were collected using a variety of techniques. The diaspores/seeds were removed from fruits manually and dried in the laboratory under ambient temperature conditions. When necessary, the seeds were kept in cold storage (5 °C) in plastic bags. Fifty diaspores/seeds per species were selected for biometric analysis with size measurements (length and diameter in mm) with a digital caliper (ZAAS-1.0004). Dry mass (mg) was estimated with a precision scale; for species whose dry mass was less than the minimum recorded by the precision scale, eight samples of 100 seeds were used.
Five hundred diaspores/seeds of each species (from at least seven trees) were sown in plastic trays containing vermiculite. The seedlings were kept at ambient temperature, ranging from 22ºC to 26 ºC, and average humidity of 70% and watered regularly every two days. Light was provided by lamps with white light foxlux® bulbs (127 Volts, 25 Watts) with a 12:00 hour photoperiod. Seedlings were grown under an average luminosity of 347±55 µmol.s-1.m-2, measured daily. The seedling stage was defined as the phase between seed germination and the formation of the first leaf or eophyll, as proposed by Souza (2003)Souza RP & Vàlio IFM (2003) Seedling growth of fifteen Brazilian tropical tree species differing in successional status. Revista Brasileira de Botânica 26: 35-47.. Development stages of seedlings were defined as: Stage I - early germination (primary root emission); Stage II - cotyledon emission, and Stage III - full expansion of the first pair of eophylls. The elapsed time at each stage was recorded (in days), using five seedlings per species.
Morphometric analysis was based on 25 seedlings (Stage III) per species, and root (mm), shoot (mm) and seedling (mm) lengths were measured with a digital caliper. Dry mass was determined using a precision analytical balance. Root (mg), shoot (mg), cotyledon (mg), and eophyll (mg) dry mass and the total mass of the seedlings (mg) were measured. Shoot biometrics refers to the shoot itself plus the cotyledons and eophylls. The following indices were used to represent the allocation of reserves to the different parts of the seedling: shoot /seedling length ratio, shoot/ seedling mass ratio, cotyledon/seedling mass ratio, eophyll /seedling mass ratio and seedling/seed mass ratio (Poorter 1999Poorter L (1999) Growth responses of 15 rain-forest tree species to a light gradient: the relative importance of morphological and physiological traits. Functional Ecology 13: 396-410.; Wrigth & Westoby 2000Wright IJ & Westoby M (2000) Cross-species relationships between seedling relative growth rate, nitrogen productivity and root vs leaf function in 28 Australian woody species. Functional Ecology 14: 97-107.; Souza & Vàlio 2003Souza RP & Vàlio IFM (2003) Seedling growth of fifteen Brazilian tropical tree species differing in successional status. Revista Brasileira de Botânica 26: 35-47.; Markesteijn & Poorter 2009Markesteijn L & Poorter L (2009) Seedling root morphology and biomass allocation of 62 tropical tree species in relation to drought- and shade- tolerance. Journal of Ecology 97: 311-325.).
The areas of cotyledons and eophylls were estimated from scanned images using Sigma- Scan Pro software (version 5.0, SPSS Inc., Chicago, IL, USA). The following traits were calculated: cotyledon specific area (cotyledon area/ cotyledon mass, mm2.mg-1); eophyll specific area (eophyll area/eophyll mass, mm2.mg-1) and total photosynthetic area (mm2), represented by the sum of cotyledon area and eophyll area.
For anatomical analysis the median region of the cotyledons and eophylls from ten seedlings per species were selected. These samples were fixed in FAA 50 (Berlyn & Miksche 1976Berlyn GP & Miksche JP (1976) Botanical microtechnique and cytochemistry. The Iowa State Press, Ames. 326p.), preserved in 50% ethanol and embedded in the historesin Leica®. Cross sections were obtained using a rotary microtome Leica RM-4125, distended on slides, stained with 0.05% toluidine blue at pH 6.8 (O’Brien et al. 1965O’Brien TP, Feder N & Mccully ME (1965) Polychromatic staining of plat cell walls by toluidine blue. Protoplasma 59: 368-373.) and mounted with Permount® (Kraus & Arduin 1997Kraus JE & Arduin M (1997) Manual básico de métodos em morfologia vegetal. EDUR, Rio de Janeiro. 155p.). Total thickness and the thickness of the palisade and spongy parenchyma of cotyledons and eophylls (µm) were measured in transverse sections using a light microscope (Olympus, CX23) with an ocular micrometer.
Statistical analysis
The sucessional groups previously formed (pioneers, early secondary and late secondary) were compared to each other to verify if they differ in the morphological characteristics evaluated. The average and standard deviation of all quantitative variables were calculated. The Shapiro-Wilk test was used to verify the normality of the data; when the assumptions of normality were not met a logarithmic transformation was employed. Subsequently, in order to verify possible variation among averages of the sucessional groups, an analysis of variance (oneway ANOVA) was performed. When statistical differences were found, the averages were compared using Tukey test, with a significance level of 5% (p < 0.05). Pearson correlation analysis was applied in order to verify if there were correlations among variables. Principal component analysis (PCA) was used to identify which variables contribute most to the variation of the data. Eigenvalue criterion and the graph of the slope (screeplot) were employed to determine the number of axes. In order to assess the degree of similarity between the species studied, we performed a cluster analysis using a matrix distance based on Euclidean distance and the Ward’s minimum variance method. Cluster analyses with a cofenetic index greater than 0.5 were considered significant. Past software, version 1.34 (Hammer et al. 2001Hammer O, Harper DAT & Ryan PD (2001) PAST: Paleontological statistics software package for education and data analysis. Paleontologia Eletrônica 4(1). Version 2.17. Disponível em <http://nhm2.uio.no/norlex/past/download.html>. Acesso em 15 junho 2013.
http://nhm2.uio.no/norlex/past/download....
) was used for all statistical analyzes. Two other methods were tested in the software used: “Paired Group” and “Single Linkage”. The results of such methods are very similar to that of “Wards”. This was the method with higher cofenetic index. According to Valentin (2000)Valentin JL (2000) Ecologia numérica: uma introdução à análise multivariada de dados ecológicos. Interciência, Rio de Janeiro. Pp. 53-65., the choice of the grouping method depends on criteria based on the lowest degree of distortion and its ability to better evidence the structure of the data.
Results
The studied ecological groups differed in several variables related to seedling size (Tab. 2). The pioneer species had the lowest masses of root, shoot, seed and seedling; and lower cotyledon area and total photosynthetic areas than the late secondary species. The early secondary species did not differ from other groups in these traits (Tab. 2).
Mean values and respective standard deviations of seed and seedling morphological traits from different ecological groups, Araucaria forest, Brazil.
The pioneer species had lower seedling and root length and cotyledon masses in comparison to early and late successional species, with no significant differences between these latter two groups. The cotyledon/seedling mass ratio of the pioneer species was lower than that of the early secondary species and did not differ from the late secondary species. Only the averages of eophyll/ seedling mass ratio and seedling/seed mass ratio were higher in pioneer species than the other groups. No differences were found among groups (Tab. 2) for the remaining variables. In nine of the 16 characteristics analyzed the pioneer species are similar to the early secondary ones. However, in five characteristics the initial secondary resemble the late secondary (Tab. 2).
Considering development stages, the late secondary species took the longest time to complete the seedling stage. The time to expose the cotyledons (Stage II) was longer in early and late secondary species compared to the pioneer species. The total time of expansion of the first pair of eophylls (Stage III), was greater in late secondary species than in the other groups (Fig. 1; Tab. 2).
Seedling development stages from Araucaria Forest species, Brazil. Stage I - early germination (primary root emission); stage II - cotyledon emission, and stage III - full expansion of the eophyll first pair. (Csc: Clethra scabra; Mp:Moquiniastrum polymorphum; Lt: Lamanonia ternata; St: Schinus terebinthifolius; Ld: Luehea divaricata; Cs: Casearia sylvestris; Cf: Cedrela fissilis; Lp: Lafoensia pacari; Pc: Psidium cattleianum; Gk:Gymnanthes klotzschiana; Ha: Handroanthus albus; Ae: Allophylus edulis; Ip: Ilex paraguariensis; Mc: Mollinedia clavigera; Ps: Psychotria suterella; Cx: Campomanesia xanthocarpa, Cd: Casearia decandra e Cdi:Cinnamodendron dinisii).
The first three axes of the principal component analysis explained 73% of the variation in the data (Tab. 3). The first axis accounted for 36% of the variation, with size characteristics seedlings and seeds being most important. Among these, the most significant are, in order of importance, dry mass and the length of seedling, seed dry mass, eophyll total thickness and total photosynthetic area. The ratios of seedling/seed mass, eophyll/ seedling mass and cotyledon specific area were negatively correlated with the first axis (Tabs. 3, 4; Fig. 2). The second axis (21%) represents seedling investment in area for light interception, as evidenced by cotyledon specific area, spongy parenchyma thickness and eophyll total thickness. This axis is negatively correlated with cotyledon total thickness, shoot/seedling mass ratio; shoot/ seedling length ratio and cotyledon/seedling mass ratio (Tabs. 3, 4; Fig. 2). The third axis (16%), in turn, is represented by variables related to resource allocation in the shoot - both in length and mass - the total photosynthetic area and eophyll/seedling mass ratio. The cotyledon/seedling mass ratio is negatively correlated with this axis (Tabs. 3, 4).
Eigenvalues and percentage (%) of total variance of Principal Component Analysis for seedling morphological characteristics from Araucaria Forest species, Brazil.
Correlations among variables and principal components of Principal Component Analysis for seedling morphological characteristics from Araucaria Forest species, Brazil.
Principal component analysis of morphological characteristics of Araucaria Forest species, Brazil. SL: seedling length; SM: seedlings mass; SdM: seed mass; CTT: cotyledon total thickness; EMS: eophyll/seedling mass ratio; ETT: eophyll total thickness; EPT: eophyll spongy parenchyma thickness; DS: development stages; CSA: cotyledons specific area; ESA: eophyll specific area; PTA: total photosynthetic area; ShMS: shoot/seedling mass ratio; ShLS: shoot/seedling length ratio; CMS: cotyledon/seedling mass ratio; SMSd: seedling/seed mass ratio. Symbols: (+) - pioneer; (▲) - early secondary; (■) - late secondary.
The most correlated species on the first axis were the late secondary species (Mollinedia clavigera, Cinnamodendron dinisii, and Campomanesia xanthocarpa), while the pioneer species (Clethra scabra, Moquiniastrum polymorphum and Luehea divaricata) are negatively correlated with this axis (Figs. 2, 3). In the second axis, however, there was no clear distinction between ecological groups, since late secondary (Allophylus edulis, C. xanthocarpa), pioneer (Schinus terebinthifolius, L. divaricata), and early secondary species (Lafoensia pacari) (Figs. 2, 3) were grouped together. The third axis is only positively correlated with two late secondary species (C. xanthocarpa and Allophylus edulis).
Principal component analysis of Araucaria Forest species, Brazil. SL: seedling length; SM: seedlings mass; SdM: seed mass; CTT: cotyledon total thickness; EMS: eophyll/seedling mass ratio; ETT: eophyll total thickness; EPT: eophyll spongy parenchyma thickness; DS: development stages; CSA: cotyledons specific area; ESA: eophyll specific area; PTA: total photosynthetic area; ShMS: shoot/seedling mass ratio; ShLS: shoot/seedling length ratio; CMS: cotyledon/seedling mass ratio; SMSd: seedling/seed mass ratio. (Csc: Clethra scabra; Mp: Moquiniastrum polymorphum; Lt: Lamanonia ternata; St: Schinus terebinthifolius; Ld: Luehea divaricata; Cs: Casearia sylvestris; Cf: Cedrela fissilis; Lp: Lafoensia pacari; Pc: Psidium cattleianum; Gk: Gymnanthes klotzschiana; Ha: Handroanthus albus; Ae: Allophylus edulis; Ip: Ilex paraguariensis; Mc: Mollinedia clavigera; Ps: Psychotria suterella; Cx: Campomanesia xanthocarpa, Cd: Casearia decandra e Cdi: Cinnamodendron dinisii).
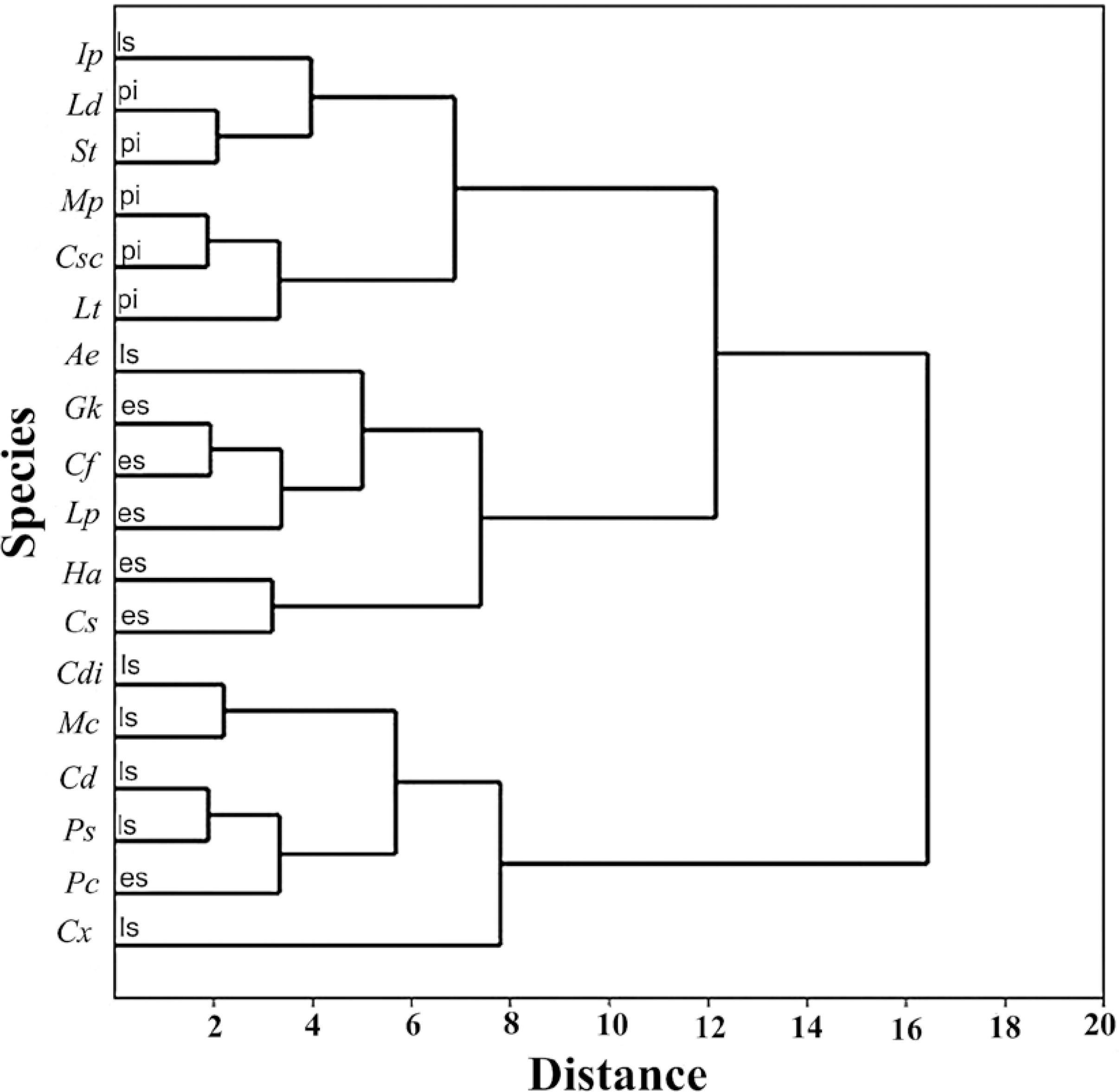
The cluster analysis using the morphofunctional variables (seedling length, seedling mass, seed mass, cotyledon total thickness, eophyll total thickness, spongy parenchyma thickness, development stages, cotyledon specific area, eophyll specific area, total photosynthetic area, shoot/seedling mass ratio, shoot/seedling length ratio, cotyledon/seedling mass ratio, eophyll/ seedling mass ratio and seedling/seed mass ratio) resulted in two main groups.
The first group is comprised of the late secondary species (M. clavigera, Psychotria suterella, C. xanthocarpa, Casearia decandra, and C. dinisii) and an early secondary species (Psidium cattleianum). The second group includes two distinct subgroups that separate the pioneers (C. scabra, M. polymorphum, Lamanonia ternata, S. terebinthifolius and L. divaricata), and most of the early secondary species (Cedrela fissilis, Casearia sylvestris, L. pacari, Gymnanthes klotzschiana, Handroanthus albus). The exceptions were Ilex paraguariensis, grouped with the pioneers, and A. edulis, included in the early secundary group (Fig. 4). Most analyzed traits (as log variables) were highly correlated (r > 0.5), as shown inTable 5.
Cluster analysis of Araucaria Forest species, Brazil. (Csc: Clethra scabra; Mp: Moquiniastrum polymorphum; Lt: Lamanonia ternata; St: Schinus terebinthifolius; Ld: Luehea divaricata; Cs: Casearia sylvestris; Cf: Cedrela fissilis; Lp: Lafoensia pacari; Pc: Psidium cattleianum; Gk:Gymnanthes klotzschiana; Ha: Handroanthus albus; Ae: Allophylus edulis; Ip: Ilex paraguariensis; Mc: Mollinediaclavigera; Ps: Psychotria suterella; Cx: Campomanesia xanthocarpa, Cd: Casearia decandra e Cdi: Cinnamodendron dinisii). Ecological group (pi: pioneer; es: early secondary; ls: late secondary).
Correlations between seedling morphological characteristics from Araucaria Forest species, Brazil. Coefficients > 0.5 were considered significant.
Discussion
Some general trends correlating seed and seedling characteristics to ecological groups found in this study are consistent with those observed by other authors (Foster & Janson 1985Foster S & Janson CH (1985) The relationship between seed size and establishment conditions in tropical woody plants. Ecology 66: 773-780.; Glesson & Tilman 1994Garwood MD (1996) Functional morphology of tropical tree seedlings. In: Swaine MD (eds.) Ecology of tropical forest tree seedlings. Unesco, Parthenon, Paris. Pp. 59-129.; Souza & Vàlio 2003Souza RP & Vàlio IFM (2003) Seedling growth of fifteen Brazilian tropical tree species differing in successional status. Revista Brasileira de Botânica 26: 35-47.; Moles & Westoby 2004Moles AT & Westoby M (2004) Seedling survival and seed size: a synthesis of the literature. Journal of Ecology 92: 372-383.; Coll et al. 2008Coll L, Potvin C, Messier C & Delagrange S (2008) Root architecture and allocation patterns of eight native tropical species with different successional status used in open-grown mixed plantations in Panama. Trees 22: 585-596.). In general, among the studied species, the pioneers have less biomass and smaller seeds and seedlings, compared to species that grow in shaded areas (Ng 1978Ng FSP (1978) Strategies of establishment in Malayan forest trees. In: Tomlinson PB & Zimmermann MH (eds.) Tropical trees as living systems. University Press, Cambridge. Pp. 129-162.; Foster & Janson 1985Foster S & Janson CH (1985) The relationship between seed size and establishment conditions in tropical woody plants. Ecology 66: 773-780.; Hladik & Miquel 1990Hladik A & Miquel S (1990) Seedling types and plant establishment in an African rains forest. In: Bawa KS, Hadley M (eds.) Reproductive ecology of tropical forest plants. Unesco, Parthenon, Paris. Pp. 261-282.; Wright & Westoby 1994Wright IJ & Wetoby M (1994) Cross-species relationships between seedling relative as correlates of seedling shade tolerance of 13 tropical trees. Oecologia 98: 419-428.; Garwood 1996Garwood MD (1996) Functional morphology of tropical tree seedlings. In: Swaine MD (eds.) Ecology of tropical forest tree seedlings. Unesco, Parthenon, Paris. Pp. 59-129.; Kitajima 1996Kitajima K (1996) Cotyledon functional morphology and patterns of seed reserve utilization by neotropical tree seedlings. In: Swaine MD (ed.) Ecology of tropical forest tree seedling. Unesco, Parthenon, Paris. Pp. 193-210.; Saverimuttu & Westoby 1996Saverimuttu T & Westoby M (1996) Seedling longevity under deep shade in relation to seed size. The Journal of Ecology, 84: 681-689.; Wright et al. 2000Wright IJ & Westoby M (2000) Cross-species relationships between seedling relative growth rate, nitrogen productivity and root vs leaf function in 28 Australian woody species. Functional Ecology 14: 97-107.).
The large amount of seed reserves in late secondary species can be allocated to different parts of the seedling during development, thereby producing larger seedlings, with greater competitive ability (Stebbins 1971Stebbins GL (1971) Adaptive radiation of reproductive characteristics in angiosperms, II: seeds and seedlings. Annual Review of Ecology and Systematics 2: 237-260.; Saverimuttu & Westoby 1996Saverimuttu T & Westoby M (1996) Seedling longevity under deep shade in relation to seed size. The Journal of Ecology, 84: 681-689.; Paz & Martinez-Ramos 2003Paz H & Martínez-Ramos M (2003) Seed mass and seedling performance within eight species of Psychotria (Rubiaceae). Ecology 84: 439-450.; Green & Juniper 2004Green PT & Juniper PA (2004) Seed-seedling allometry in tropical rain forest trees: seed mass-related patts of resource allocation and ‘reserve effect’. Journal of Ecology 92: 937-408.; Baraloto et al. 2005Baraloto C, Michel P & Goldberg DE (2005) Seed mass, seedling size and neotropical tree seedling establishment. Journal of Ecology 93: 1156-1166.), and survival (Baraloto et al. 2005Baraloto C, Michel P & Goldberg DE (2005) Seed mass, seedling size and neotropical tree seedling establishment. Journal of Ecology 93: 1156-1166.; Baraloto & Forget 2007Baraloto C & Forget PM (2007) Seed size, seedling morphology, and response to deep shade and damage in neotropical rain forest trees. American Journal of Botany 94: 901-911.) in shaded environments. This trend becomes clear when considering the positive correlation between seed mass and the biometric variables of seedlings, as has been previously observed by several other authors (Sworn & Westoby 1992; Green & Juniper 2004Green PT & Juniper PA (2004) Seed-seedling allometry in tropical rain forest trees: seed mass-related patts of resource allocation and ‘reserve effect’. Journal of Ecology 92: 937-408.; Baraloto et al. 2005Baraloto C, Michel P & Goldberg DE (2005) Seed mass, seedling size and neotropical tree seedling establishment. Journal of Ecology 93: 1156-1166.). In absolute values, the late secondary seedlings have greater biomass, total length and photosynthetic area. These results represent a greater investment in size in these species.
However, despite the smaller seedling of pioneer species, they develop more rapidly and are less dependent on seed mass, given the seedling/ seed mass ratio. This index represents how much a seedling grows in relation to seed size. Some studies report that species with smaller seeds tend to have higher growth rates (Sworn & Westoby 1992; Gleeson & Tilman 1994Gleeson SK & Tilman D (1994) Plant allocation, growth rate and sucessional status. Functional Ecology 8: 543-550.; Garwood 1996Embrapa (1999) Centro Nacional de Pesquisa de Solos. Sistema brasileiro de classificação de solos. Embrapa Produção de Informações, Brasília; Embrapa Solos, Rio de Janeiro. 412p.; Soriano et al. 2011Soriano D, Orozco-Segovia A, Marquez-Guzman J, Kitajima K, Gamboa-Debuen A & Huante P (2011) Seed reserve composition in 19 tree species of a tropical deciduous forest in Mexico and its relationship to seed germination and seedling growth. Annals of Botany 107: 939-951.) facilitating rapid colonization in early development stages, such as in the pioneer species (Souza & Vàlio 2003Souza RP & Vàlio IFM (2003) Seedling growth of fifteen Brazilian tropical tree species differing in successional status. Revista Brasileira de Botânica 26: 35-47.).
In addition, a negative correlation between seed mass and cotyledon specific area, observed in the pioneer species, indicates that in relative terms there is greater efficiency in the allocation to photosynthetic tissue in seedlings from smaller seeds. This corroborates the expected strategy of species in early stages of succession. Several studies have found that shade-tolerant species tend to have lower leaf specific area in addition to a lower growth rate (Baraloto et al. 2005Baraloto C, Michel P & Goldberg DE (2005) Seed mass, seedling size and neotropical tree seedling establishment. Journal of Ecology 93: 1156-1166.; Poorter et al. 2006Poorter L, Bongers L & Bongers F (2006) Architecture of 54 moist-forest tree species: traits, trade-offs, and functional groups. Ecology 87: 1289-1301.; Coll et al. 2008Coll L, Potvin C, Messier C & Delagrange S (2008) Root architecture and allocation patterns of eight native tropical species with different successional status used in open-grown mixed plantations in Panama. Trees 22: 585-596.; Markesteijn & Poorter 2009Markesteijn L & Poorter L (2009) Seedling root morphology and biomass allocation of 62 tropical tree species in relation to drought- and shade- tolerance. Journal of Ecology 97: 311-325.). In general, these species have larger amounts of reserve substances, which facilitates survival in the understory (Gogosz et al. 2015Gogosz AM, Boeger MRT, Cosmo NL & Nogueira AC (2015) Morfologia de diásporos e plântulas de espécies arbóreas da floresta com araucária, no sul do Brasil. Floresta 45: 819- 832.). The positive correlation between seed mass and cotyledon mass suggests that species with larger seeds produce cotyledons with larger mass, and probably with larger reserves, as also observed by Green & Juniper (2004)Green PT & Juniper PA (2004) Seed-seedling allometry in tropical rain forest trees: seed mass-related patts of resource allocation and ‘reserve effect’. Journal of Ecology 92: 937-408..
In general, plants that grow inside forests, such as late secondary species, have slower development and are more dependent on seed reserves. Species of open environments (pioneers) have faster development, are less dependent on reserves and have a greater contribution from photosynthesis early in growth (Baraloto et al. 2005Baraloto C, Michel P & Goldberg DE (2005) Seed mass, seedling size and neotropical tree seedling establishment. Journal of Ecology 93: 1156-1166.; Markesteijn & Poorter 2009Markesteijn L & Poorter L (2009) Seedling root morphology and biomass allocation of 62 tropical tree species in relation to drought- and shade- tolerance. Journal of Ecology 97: 311-325.; Gogosz et al. 2015Gogosz AM, Boeger MRT, Cosmo NL & Nogueira AC (2015) Morfologia de diásporos e plântulas de espécies arbóreas da floresta com araucária, no sul do Brasil. Floresta 45: 819- 832.), as seen in this study. This also becomes evident when observing the presence of reserve tissue in the late secondary species.
In this study, late secondary species allocated more biomass to cotyledons than the pioneer species. These reserves in the cotyledons permit the survival of seedlings until the photosynthetic apparatus is fully developed, enabling them to overcome any periods of carbon deficit (Kitajima 1994Kitajima K (1994) Relative importance of photosynthetic traits and allocation patterns correlates of seedling shade tolerance of 13 tropical tree species. Oecologia 98: 419-428.; Ibarra- Manriquez et al. 2001Ibarra-Manríquez G, Ramos MM & Oyama K (2001) Seedling functional types in a lowland rain forest in Mexico. American Journal of Botany 88: 1801- 1812.; Green & Juniper 2004Green PT & Juniper PA (2004) Seed-seedling allometry in tropical rain forest trees: seed mass-related patts of resource allocation and ‘reserve effect’. Journal of Ecology 92: 937-408.). This trend is generally valid for species with reserve cotyledons (Baraloto et al. 2005Baraloto C, Michel P & Goldberg DE (2005) Seed mass, seedling size and neotropical tree seedling establishment. Journal of Ecology 93: 1156-1166.). However, even species with foliaceous cotyledons differ in the amount of available reserves (Garwood 1996Garwood MD (1996) Functional morphology of tropical tree seedlings. In: Swaine MD (eds.) Ecology of tropical forest tree seedlings. Unesco, Parthenon, Paris. Pp. 59-129.), as observed in the studied species. Although all species studied had photosynthetic cotyledons, many of the late secondary also had reserves in this organ. Pioneer species have thin (membranous) cotyledons, as opposed to late secondary species, that tend to have thicker cotyledons (Gogosz et al. 2015Gogosz AM, Boeger MRT, Cosmo NL & Nogueira AC (2015) Morfologia de diásporos e plântulas de espécies arbóreas da floresta com araucária, no sul do Brasil. Floresta 45: 819- 832.). These differences were evident in the exposure time of the cotyledons, since in most of the late secondary species the cotyledons remained surrounded by the seed coats for a longer period of time, and probably acting as haustorial and/or reserve organ in this stage.
The development and establishment of seedlings are complex processes, where functions such as photosynthesis and reserve absorption are distributed among different organs dynamically (Garwood 1996Garwood MD (1996) Functional morphology of tropical tree seedlings. In: Swaine MD (eds.) Ecology of tropical forest tree seedlings. Unesco, Parthenon, Paris. Pp. 59-129.). Thus, a fully formed cotyledon can be photosynthetically active in an earlier stage, yet also provide a certain amount of reserve material. In the studied species, this is probably more relevant in seedlings with greater cotyledon biomass and those from larger seeds.
In addition to seedling morphological traits, anatomical features are also important for plant species growth (Garwood 1996Garwood MD (1996) Functional morphology of tropical tree seedlings. In: Swaine MD (eds.) Ecology of tropical forest tree seedlings. Unesco, Parthenon, Paris. Pp. 59-129.). According Gogosz (2013)Gogosz AM (2013) Morfologia funcional de plântulas como indicador fisionômico da dinâmica de regeneração de espécies arbóreas da Floresta Ombrófila Mista, Paraná.Tese de Doutorado. Universidade Federal do Paraná, Curitiba. 93p., variation in eophyll total thickness among ecological groups is a result of the greater thickness of spongy parenchyma. This tissue has lobed cells and large intercellular spaces to improve light absorption by chloroplasts, because the spongy parenchyma tends to have more efficient internal reflection (Vogelmann 1993Vogelmann TC (1993) Plant tissue optics. Annual Review of Plant Physiology and Plant Molecular Biology 44: 231-251.). Moreover, this structure allows more efficient diffusion of CO2 into the mesophyll, increasing photosynthetic performance (Lambers et al. 1998Lambers H, Chapin FS & Pons TL (1998) Plant physiological ecology. Springer, New York. Pp. 321-367.). Therefore, the greater eophyll thickness of the late secondary species is a strategy for intercepting light, since they develop in low light availability.
The morphological and functional differences observed among the groups may result in different strategies of seedlings establishment in the successional process, related to the different niches of species regeneration. Such differences were observed in many of the evaluated traits. In general, the variability among species is mainly explained by seedling size, tissue investment for light interception and the shoot resource allocation.
Late secondary species have larger seedlings with characteristics that indicate slower initial development, dependent on seed reserves. The pioneers species have smaller seedlings but faster development and earlier dependence on photosynthesis. Analyzing the results obtained for most of the characteristics of the early secondary species, it is verified that these are similar with the other groups.
The morphofunctional traits of seeds and seedlings analyzed in this study allowed the classification of species in different groups, partially corresponding to three successional groups previously defined by Gogosz et al. (2015)Gogosz AM, Boeger MRT, Cosmo NL & Nogueira AC (2015) Morfologia de diásporos e plântulas de espécies arbóreas da floresta com araucária, no sul do Brasil. Floresta 45: 819- 832.. The exceptions - Ilex paraguariensis, grouped with the pioneers, and A. edulis, included in the early secondary group - have some characteristics typical of pioneers, such as small seeds and rapid initial development, respectively. However, morphological and autoecological characteristics typical of late secondary species (fleshy fruits, zoochory, greater longevity and shadow tolerance) are predominant in these species (Gogosz et al. 2015Gogosz AM, Boeger MRT, Cosmo NL & Nogueira AC (2015) Morfologia de diásporos e plântulas de espécies arbóreas da floresta com araucária, no sul do Brasil. Floresta 45: 819- 832.). Therefore, the use of the new attributes analyzed by the study, subsidy to Gogosz et al. (2015)Gogosz AM, Boeger MRT, Cosmo NL & Nogueira AC (2015) Morfologia de diásporos e plântulas de espécies arbóreas da floresta com araucária, no sul do Brasil. Floresta 45: 819- 832..
The formation of a group that bring together the subgroups of pioneers and early secondary species indicates greater similarity between these two categories, when compared with the late secondary species.
Swaine & Whitmore (1988)Swaine MD & Whitmore TC (1988) On the definition of ecological species groups in tropical rain forests. Vegetatio 75: 81-86. proposed that tree species should be divided into pioneers and non-pioneers (equivalent to late secondary species) due the difficulty to identify subgroups when there is a lack of quantitative data for many species. Thus, several species cited in the literature as early secondary species would be considered by Swaine & Whitmore (1988)Swaine MD & Whitmore TC (1988) On the definition of ecological species groups in tropical rain forests. Vegetatio 75: 81-86. as long-lived pioneers.
These results indicate that the number and quality of available data should be considered in the definition of successional categories adopted in ecological studies. When there are few variables studied, it is preferable to classify the species into only two sucessional groups (eg.: pioneers vs non- pioneers). With sufficient information available, the classification into three groups, as adopted in this study, is efficient. Further analysis of other ecological (survival), physiological (growth rate, light compensation point) and anatomical features (thickness and number of layers the leaf tissues, stomatal density, reserve amount) may contribute to the identification and interpretation of morphofunctional patterns and to improve the classification systems.
Acknowledgements
The authors are grateful to CAPES (Coordination for Advances in Scientific Research) for providing the scholarship to the first author and CNPq (National Research Council) process n. 301971/2013-7 for providing the research assistantship to MRT Boeger.
References
- Baraloto C, Michel P & Goldberg DE (2005) Seed mass, seedling size and neotropical tree seedling establishment. Journal of Ecology 93: 1156-1166.
- Baraloto C & Forget PM (2007) Seed size, seedling morphology, and response to deep shade and damage in neotropical rain forest trees. American Journal of Botany 94: 901-911.
- Bazzaz FA (1979) The Physiological ecology of plant succession. Annual Review of Ecology and Systematics 10: 351-371.
- Berlyn GP & Miksche JP (1976) Botanical microtechnique and cytochemistry. The Iowa State Press, Ames. 326p.
- Brazão JEM & Santos MM (1997) Vegetação. In: IBGE (ed.) Recursos naturais e meio ambiente: uma visão do Brasil. IBGE, Rio de Janeiro. Pp. 111-124.
- Burger HD (1972) Seedlings of some tropical trees and shrubs mainly of South East Asia. PUDOC, Wageningen. 399p.
- Coll L, Potvin C, Messier C & Delagrange S (2008) Root architecture and allocation patterns of eight native tropical species with different successional status used in open-grown mixed plantations in Panama. Trees 22: 585-596.
- Egerton FN (2009) Homage to Frederic E. Clements, historian of plant succession studies. Bulletin of the Ecological Society of America 90: 43-79.
- Embrapa (1999) Centro Nacional de Pesquisa de Solos. Sistema brasileiro de classificação de solos. Embrapa Produção de Informações, Brasília; Embrapa Solos, Rio de Janeiro. 412p.
- Foster S & Janson CH (1985) The relationship between seed size and establishment conditions in tropical woody plants. Ecology 66: 773-780.
- Garwood MD (1996) Functional morphology of tropical tree seedlings. In: Swaine MD (eds.) Ecology of tropical forest tree seedlings. Unesco, Parthenon, Paris. Pp. 59-129.
- Giller PS (1984) Community structure and the niche. Chapman and Hall, Londan, New York. 176p.
- Gleeson SK & Tilman D (1994) Plant allocation, growth rate and sucessional status. Functional Ecology 8: 543-550.
- Gogosz AM (2013) Morfologia funcional de plântulas como indicador fisionômico da dinâmica de regeneração de espécies arbóreas da Floresta Ombrófila Mista, Paraná.Tese de Doutorado. Universidade Federal do Paraná, Curitiba. 93p.
- Gogosz AM, Boeger MRT, Cosmo NL & Nogueira AC (2015) Morfologia de diásporos e plântulas de espécies arbóreas da floresta com araucária, no sul do Brasil. Floresta 45: 819- 832.
- Green PT & Juniper PA (2004) Seed-seedling allometry in tropical rain forest trees: seed mass-related patts of resource allocation and ‘reserve effect’. Journal of Ecology 92: 937-408.
- Hammer O, Harper DAT & Ryan PD (2001) PAST: Paleontological statistics software package for education and data analysis. Paleontologia Eletrônica 4(1). Version 2.17. Disponível em <http://nhm2.uio.no/norlex/past/download.html>. Acesso em 15 junho 2013.
» http://nhm2.uio.no/norlex/past/download.html - Hladik A & Miquel S (1990) Seedling types and plant establishment in an African rains forest. In: Bawa KS, Hadley M (eds.) Reproductive ecology of tropical forest plants. Unesco, Parthenon, Paris. Pp. 261-282.
- Ibarra-Manríquez G, Ramos MM & Oyama K (2001) Seedling functional types in a lowland rain forest in Mexico. American Journal of Botany 88: 1801- 1812.
- Ipardes - Instituto Paranaense de Desenvolvimento Econômico e Social (2014) Base ambiental: Unidades de Conservação e remanescentes vegetais - Paraná. Disponível em <http://www.ipardes.gov.br/index.php>. Acesso em 13 outubro 2016.
» http://www.ipardes.gov.br/index.php - Kitajima K (1994) Relative importance of photosynthetic traits and allocation patterns correlates of seedling shade tolerance of 13 tropical tree species. Oecologia 98: 419-428.
- Kitajima K (1996) Cotyledon functional morphology and patterns of seed reserve utilization by neotropical tree seedlings. In: Swaine MD (ed.) Ecology of tropical forest tree seedling. Unesco, Parthenon, Paris. Pp. 193-210.
- Kitajima K (2002) Do shade-tolerant tropical tree seedlings depend longer on seed reserves? Functional growth analysis of three Bignoniaceae species. Functional Ecology 16: 433-444.
- Kitajima K (2003) Impact of cotyledon and leaf removal on seedling survival in three tree species with contrasting cotyledon functions. Biotropica 35: 429-434.
- Kitajima K & Poorter L (2008) Functional basis for resource niche partitioning by tropical trees. In: Carson WP & Schnitzer SA (eds.) Tropical forest community ecology. Blackwell Science, Oxford. Pp. 160-181.
- Kraus JE & Arduin M (1997) Manual básico de métodos em morfologia vegetal. EDUR, Rio de Janeiro. 155p.
- Lambers H, Chapin FS & Pons TL (1998) Plant physiological ecology. Springer, New York. Pp. 321-367.
- Lüttge U (1997) Physiological ecology of tropical plants. Springer, New York. Pp. 103-148.
- Markesteijn L & Poorter L (2009) Seedling root morphology and biomass allocation of 62 tropical tree species in relation to drought- and shade- tolerance. Journal of Ecology 97: 311-325.
- Moles AT & Westoby M (2004) Seedling survival and seed size: a synthesis of the literature. Journal of Ecology 92: 372-383.
- Ng FSP (1978) Strategies of establishment in Malayan forest trees. In: Tomlinson PB & Zimmermann MH (eds.) Tropical trees as living systems. University Press, Cambridge. Pp. 129-162.
- O’Brien TP, Feder N & Mccully ME (1965) Polychromatic staining of plat cell walls by toluidine blue. Protoplasma 59: 368-373.
- Paz H & Martínez-Ramos M (2003) Seed mass and seedling performance within eight species of Psychotria (Rubiaceae). Ecology 84: 439-450.
- Pina-Rodrigues FCM, Costa LGS & Reis A (1990) Estratégias de estabelecimento de espécies arbóreas e o manejo de florestas tropicais. In: Anais do 6º Congresso Florestal Brasileiro. Campos do Jordão. Pp. 676-684.
- Poorter L (1999) Growth responses of 15 rain-forest tree species to a light gradient: the relative importance of morphological and physiological traits. Functional Ecology 13: 396-410.
- Poorter L (2007) Are species adapted to their regeneration niche, adult niche, or both? The American Naturalist 169: 433-442.
- Poorter L, Bongers L & Bongers F (2006) Architecture of 54 moist-forest tree species: traits, trade-offs, and functional groups. Ecology 87: 1289-1301.
- Primack RB (1990) Seed physiology, seed germination and seedling ecology - commentary. In: Bawa KS & Hadley M (eds.) Reproductive ecology of tropical forest plants. Unesco, Parthenon, Paris. Pp. 233-236.
- Rees M, Condit R, Crawley M, Pacala S & Tilman D (2010) Long-term studies of vegetation dynamics. Science 293: 650-655.
- Ressel K, Guilherme FAG, Schiavini I & Oliveira PE (2004) Ecologia morfofuncional de plântulas de espécies arbóreas da Estação Ecológica do Panga, Uberlândia, Minas Gerais. Revista Brasileira de Botânica 27: 311-323.
- Saverimuttu T & Westoby M (1996) Seedling longevity under deep shade in relation to seed size. The Journal of Ecology, 84: 681-689.
- Soriano D, Orozco-Segovia A, Marquez-Guzman J, Kitajima K, Gamboa-Debuen A & Huante P (2011) Seed reserve composition in 19 tree species of a tropical deciduous forest in Mexico and its relationship to seed germination and seedling growth. Annals of Botany 107: 939-951.
- Souza LA (2003) Morfologia e anatomia vegetal: célula, tecidos, órgãos e plântula. UEPG, Ponta Grossa. 200p.
- Souza RP & Vàlio IFM (2003) Seedling growth of fifteen Brazilian tropical tree species differing in successional status. Revista Brasileira de Botânica 26: 35-47.
- Stebbins GL (1971) Adaptive radiation of reproductive characteristics in angiosperms, II: seeds and seedlings. Annual Review of Ecology and Systematics 2: 237-260.
- Swaine MD & Whitmore TC (1988) On the definition of ecological species groups in tropical rain forests. Vegetatio 75: 81-86.
- Valentin JL (2000) Ecologia numérica: uma introdução à análise multivariada de dados ecológicos. Interciência, Rio de Janeiro. Pp. 53-65.
- Vogel EF (1980) Seedlings of dicotyledons: structure, development, types: descriptions of 150 woody Malesian taxa. Centre for Publishing and Documentation, Wageningen. Pp. 26-48.
- Vogelmann TC (1993) Plant tissue optics. Annual Review of Plant Physiology and Plant Molecular Biology 44: 231-251.
- Wright IJ & Wetoby M (1994) Cross-species relationships between seedling relative as correlates of seedling shade tolerance of 13 tropical trees. Oecologia 98: 419-428.
- Wright IJ & Westoby M (2000) Cross-species relationships between seedling relative growth rate, nitrogen productivity and root vs leaf function in 28 Australian woody species. Functional Ecology 14: 97-107.
- Wright IJ, Clifford HT, Kidson R, Reed ML, Rice BL & Westoby MA (2000) Survey of seed and seedling characters in 1744 Australian dicotyledon species: cross-species trait correlations and correlated trait- shifts within evolutionary lineages. Biological Journal of the Linnean Society 69: 521-547.
Edited by
Publication Dates
-
Publication in this collection
18 Mar 2019 -
Date of issue
2019
History
-
Received
07 Feb 2017 -
Accepted
25 Feb 2018