
Abstract
Besides their ecological importance, epiphytic species of Orchidaceae play economic and social roles through their commercialization and some are at great risk of extinction. The objectives of this study were to characterize the leaf and root anatomy of fourteen epiphytic Orchidaceae species, which occur in indigenous territory in the Parque Estadual da Serra do Tabuleiro (P.E.S.T.), Santa Catarina, Brazil and to identify adaptive anatomical characteristics related to the epiphytic habit. The species are commercialized by the Guarani and were collected during interviews and guided tours with Guarani in the indigenous territory. The results reveal the species have many morphoanatomical structures that are useful during water shortages resulting from the epiphytic habit. Notable characteristics are related to reserving water (i.e., pseudobulbs and a hypodermis with water-storage cells) and resistance to desiccation in the leaf (i.e., conspicuous cuticle, suprastomatic chamber and extraxylary and pericyclic fibers) and root (i.e., tilosomes and/or exodermal thickening and cortex cells with phi thickenings or sclereids). Descriptions and the identification of adaptive characteristics of epiphytic plant species are useful for conservation and cultivation studies, especially for plants commercially used by the Guarani Indians.
Key words:
hypodermis; leaf; pseudobulb; root; trichome; velamen
Resumo
Além de sua importância ecológica, espécies epífitas de Orchidaceae desempenham papéis econômicos e sociais através de sua comercialização e algumas estão em grande risco de extinção. O objetivo deste estudo foi caracterizar a anatomia foliar e radicular de quatorze espécies epífitas de Orchidaceae, que ocorrem em território indígena no Parque Estadual da Serra do Tabuleiro (PEST), Santa Catarina, Brasil e identificar características anatômicas adaptativas relacionadas ao hábito epifitico. As espécies são comercializadas pelos Guarani e foram coletadas durante entrevistas e visitas guiadas com os Guarani no território indígena. Os resultados revelam que as espécies possuem muitas estruturas morfoanatômicas úteis durante a escassez de água resultante do hábito epifítico. Características notáveis estão relacionadas à reserva de água (pseudobulbos e hipoderme com células de armazenamento de água) e resistência à dessecação na folha (cutícula conspícua, câmara suprasomática e fibras extraxilares e pericíclicas) e raiz (i.e., tilossomas e / ou espessamento exodérmico e células do córtex com espessamentos phi ou esclereides). Descrições e identificação de características adaptativas de espécies de plantas epífitas são úteis para estudos de conservação e cultivo, especialmente para plantas utilizadas comercialmente pelos índios Guarani.
Palavras-chave:
hipoderme; folha; pseudobulbo; raiz; tricoma; velame
Introduction
The family Orchidaceae is the second largest angiosperm family, in number of species, and includes 736 genera in five subfamilies (Chase et al. 2015Chase MW, Cameron KM, Freudenstein JV, Pridgeon AM, Salazar G, Van den Berg C & Schuiteman A (2015) An updated classification of Orchidaceae. Botanical Journal of the Linnean Society 177: 151-174. DOI: <https://doi.org/10.1111/boj.12234>.
https://doi.org/10.1111/boj.12234...
). The most derived and richest subfamily is Epidendroideae, which includes approximately 650 genera and 18,000 species (Dressler 2005Dressler R (2005) How many Orchid species? Selbyana 26: 155-158.; Siqueira et al. 2014Siqueira CE, Zanin A & Neto LM (2014) Orchidaceae in Santa Catarina: update, geographic distribution and conservation. Check List 10: 1452-1478.). Orchidaceae are herbaceous, sometimes mycoheterotrophic, and can be epiphytic (73% of the species in the family), hemiepiphytic, terrestrial or rupicolous (Koch et al. 2014Koch AK, Santos JUM & Ilkiu-Borges AL (2014) Sinopse das Orchidaceae holoepífitas e hemiepífitas da Floresta Nacional de Caxiuanã, PA, Brasil. Hoehnea 41: 129-148.). In addition, among epiphytic plants, Orchidaceae dominate in number and diversity of species (Kersten 2010Kersten RDA (2010) Epífitas vasculares - histórico, participação taxonômica e aspectos relevantes, com ênfase na Mata Atlântica. Hoehnea 37: 9-38.) in all ecosystems worldwide, especially in neotropical regions (Dressler 2005; Chase et al. 2015). These species play an important ecological role because they can quickly catch and reintegrate energy and matter into an ecosystem (Nadkarni 1984Nadkarni NM (1984) Epiphyte biomass and nutrient capital of a Neotropical Elfin Forest. Biotropica 16: 249. DOI: <https://doi.org/10.2307/2387932>
https://doi.org/10.2307/2387932...
) that creates, through the accumulation of organic material, a rich source of nutrients available to above-ground fauna (Nadkarni 1984; Balachandar et al. 2019Balachandar M, Koshila Ravi R, Ranjithamani A & Muthukumar T (2019) Comparative vegetative anatomy and mycorrhizal morphology of three South Indian Luisia species (Orchidaceae) with the note on their epiphytic adaptations. Flora 251: 39-6.).
Epiphytic vascular plants are mainly found in tropical and subtropical forests on tree trunks and branches (Benzing 2008Benzing DH (2008) Vascular epiphytes. General biology and related biota. Vol. 1. Ed. Cambridge University Press, New York. 295p.). Since their roots are not in contact with the soil, water is the main limiting factor for their survival, growth and distribution (Zotz & Hietz 2001Zotz G & Hietz P (2001) The physiological ecology of vascular epiphytes: current knowledge, open questions. Journal of Experimental Botany 52: 2067-2078.). For this reason, these plants have developed different morphoanatomical adaptations in their leaves, stems and roots to deal with water deficit (Benzing 2008). In epiphytic Orchidaceae species, some adaptations have been reported, such as velamen that absorbs water, provides mechanical protection and reduces perspiration, presence of water-storage parenchyma in the pseudobulbs or in the leaves, and occurrence of Crassulacean Acid Metabolism (CAM) (Pridgeon et al. 1983Pridgeon AM, Stern WL & Benzing DH (1983) Tilosomes in roots of Orchidaceae: morphology and systematic occurrence. American Journal of Botany 70: 1365-1377. DOI: <https://doi.org/10.1002/j.1537-2197.1983.tb07926.x>.
https://doi.org/10.1002/j.1537-2197.1983...
; Benzing 1986; Bonates 1993Bonates LCM (1993) Estudos ecofisiológicos de Orchidaceae da Amazônia II - anatomia ecológica foliar de espécies com metabolismo CAM de uma campina da Amazôniza central. Acta Amazonica 23: 315-348.; Oliveira & Sajo 2001Oliveira VC & Sajo MG (2001) Morfo-anatomia caulinar de nove espécies de Orchidaceae. Acta Botanica Brasilica 15: 176-188.; Piazza et al. 2015Piazza LD, Smidt EC & Bona C (2015) Anatomia comparada dos órgãos vegetativos de espécies de Bulbophyllum seção Didactyle (Lindl.) Cogn. e Bulbophyllum seção. Xiphizusa Rchb.f. (Orchidaceae). Hoehnea 42: 171-183.). In addition, many anatomical characteristics vary among species, representing different strategies of epiphytic survival that can be used in taxonomic studies. Oliveira-Pires et al. (2003)Oliveira-Pires MDF, Semir J, Melo De Pinna GFDA & Felix LP (2003) Taxonomic separation of the genera Prosthechea and Encyclia (Laeliinae: Orchidaceae) using leaf and root anatomical features. Botanical Journal of the Linnean Society 143: 293-303. DOI: <https://doi.org/10.1046/j.1095-8339.2003.00195.x>.
https://doi.org/10.1046/j.1095-8339.2003...
analyzed species of Prosthechea Hook. and Encyclia Knowles & Westc. noted differences and identified unusual characteristics that are not found in other epiphytic species (e.g., flavonoid crystals in the roots, fiber strands in the subepidermal layer and thickness of the leaf cuticle), which they used to distinguish the studied genera.
Many species of orchids have been widely used by traditional communities as ornamental plants such species of the genus Arachnis (Foge et al. 2019Foge BA, Essomo SE, Bechem TE, Tabot PT, Arrey BD, Afanga Y & Assoua EM (2019) Market trends and ethnobotany of orchids of Mount Cameroon. Journal Ethnobiology Ethnomedicine 15: 2-11.), food such Vanilla planifolia (Bhatnagar et al. 2017Bhatnagar M, Sarkar N, Gandharv N, Apang O, Singh S & Ghosal S (2017) Evaluation of antimycobacterial, leishmanicidal and antibacterial activity of three medicinal orchids of Arunachal. Complementary and Alternative Medicine 17: 1-10. DOI: <https://doi.org/10.1186/s12906-017-1884-z>
https://doi.org/10.1186/s12906-017-1884-...
) and, especially, as phytotherapeutic medicines such Rhynchostylis retusa (Hinsley et al. 2018Hinsley A, Hugo JB, Michael FF, Stephan WG, Lauren M, Gardiner RSG, Pankaj K, Susanne M, Destario M, David LR, Sarina V, Shan W & Jacob P (2018) A review of the trade in orchids and its implications for conservation. Botanical Journal of the Linnean Society 8: 435-455.) by indigenous peoples of India. Some species such as Cyrtorchis arcuata, Diaphananthe millarii, Diaphananthe xanthopollinia used by traditional communities in South Africa also have bioactive compounds that can be studied for the production of medicines (Ng & Hew 2000Ng CKY & Hew CS (2000) Orchid pseudobulbs - “false” bulbs with a genuine importance in orchid growth and survival! Scientia Horticulturae 83: 165-172.; Chinsamy et al. 2011Chinsamy M, Finnie JF & Van Staden J (2011) A etnobotânica das orquídeas medicinais sul-africanas. Jornal Sul-Africano de Botânica 77: 2-9. DOI: <https://doi.org/10.1016/j.sajb.2010.09.015>
https://doi.org/10.1016/j.sajb.2010.09.0...
; Bhatnagar et al. 2017).
In Brazil, orchids are present on the indigenous culture, such as in Guarani traditional histories, and also as a source of income through the commercialization of orchid species by Guarani indigenous people in the southern part of the country (Stehmann et al. 2009Stehmann JR, Forzza RC, Salino A, Sobral M, Costa DP & Kamino LHY (Org.) (2009) Plantas da Floresta Atlântica. Instituto de Pesquisa Jardim Botânico do Rio de Janeiro, Rio de Janeiro. 500p.; Siqueira et al. 2014Siqueira CE, Zanin A & Neto LM (2014) Orchidaceae in Santa Catarina: update, geographic distribution and conservation. Check List 10: 1452-1478.; Blanco 2017Blanco GD (2017) Estudo etnobotânico e morfoanatômico de espécies de Orchidaceae utilizadas por grupos Guarani. Dissertação de Mestrado. Universidade Federal de Santa Catarina, Florianopolis. 163p.). In the urban centers of south Brazil, Guarani groups acquire part of their income commercializing handicrafts and living plants, mostly orchids. However, because some commercially important orchid species are threatened with extinction, the search for adequate management techniques is intensifying (Swarts & Dixon 2009Swarts ND & Dixon KW (2009) Terrestrial orchid conservation in the age of extinction. Annals of Botany 2: 543-556. DOI: <https://doi.org/10.1093/aob/mcp025>.
https://doi.org/10.1093/aob/mcp025...
). Successfully managing nurseries is severely limited by the lack of general comprehension regarding orchids’ anatomical adaptation mechanisms to the environment, thus making it harder to optimize orchid’s survival at nurseries (Duarte & Gandolfi 2017Duarte MM & Gandolfi S (2017) Forest ecology and management diversifying growth forms in tropical forest restoration: enrichment with vascular epiphytes. Forest Ecology and Management 401: 89-98. DOI: <https://doi.org/10.1016/j.foreco.2017.06.063>.
https://doi.org/10.1016/j.foreco.2017.06...
). In Monte Camarões, the sale of orchids is also an important source of income for indigenous groups, and for this reason these groups have sought strategies for the conservation of species (Foge et al. 2019Foge BA, Essomo SE, Bechem TE, Tabot PT, Arrey BD, Afanga Y & Assoua EM (2019) Market trends and ethnobotany of orchids of Mount Cameroon. Journal Ethnobiology Ethnomedicine 15: 2-11.).
Serra do Tabuleiro State Park is a fully protected Brazilian conservation unit. The mainland area of the park encompasses part of the Florianópolis, Palhoça, Santo Amaro da Imperatriz, Águas Mornas, São Bonifácio, São Martinho, Imaruí and Paulo Lopes municipalities (about 1% of the state), has different plant formations of the Atlantic Forest biome and harbors great biological diversity (Blanco 2017Blanco GD (2017) Estudo etnobotânico e morfoanatômico de espécies de Orchidaceae utilizadas por grupos Guarani. Dissertação de Mestrado. Universidade Federal de Santa Catarina, Florianopolis. 163p.). The flora of Santa Catarina State is considered the best known in Brazil, mainly because of the publications “Flora Ilustrada Catarinense,” which is still ongoing, and “Inventário Florıstico Florestal de Santa Catarina” (Hassemer et al. 2015Hassemer G, Giovanni R & Trevisan R (2015) The use of potential distribution models in the study of the distribution and conservation status of plants: the case of Plantago L. (Plantaginaceae) in Brazil. The Journal of the Torrey Botanical Society 143: 1-38.). In this region there are threatened or vulnerable species of Orchidaceae, such as from the genus Cattleya (Blanco 2017). In Santa Catarina State occur 121 genera and 560 accepted orchids species and 12 of them endemic (Siqueira et al. 2014Siqueira CE, Zanin A & Neto LM (2014) Orchidaceae in Santa Catarina: update, geographic distribution and conservation. Check List 10: 1452-1478.). Among these species, according to the criteria of the International Union for Conservation of Nature, 24 species fall within the vulnerable category, seven are endangered and four are critically endangered (Siqueira et al. 2014). Anatomical studies of Orchidaceae species in the state are very scarce, Warmling (2013)Warmling J (2013) Anatomia da folha e ramicaule de espécies da subtribo Pleurothallidinae (Orchidaceae). Dissertação de Mestrado. Universidade Federal de Santa Catarina, Florianópolis. 109p. and Avi & Rodrigues (2019)Avi RC & Rodrigues AC (2019) Comparative anatomy of the leaf and ramicaule in Pleurothallidinae (Orchidaceae) species. The Journal of the Torrey Botanical Society 146: 198-212. who worked with leaf and ramicaule of Pleurothallidinae species to taxonomy.
Thus, the objectives of the present study were to describe and compare the leaf and root anatomy of epiphytic Orchidaceae species that are commercially used by the Guarani in areas of Serra do Tabuleiro State Park, in order to better understand the adaptive capacity of these species in relation to the environment where they grow.
Material and Methods
Study site
The orchids were collected in December 2015 and January 2016, in the Mbyá-Guarani Morro dos Cavalos Indigenous Territory (or Itaty, in the Guarani language), in the municipality of Palhoça, Santa Catarina state, Brazil (27°48.060’S, 48°39.204’W) (Fig. 1a-b). The indigenous territory is located in remnants of Atlantic Forest, together with Serra do Tabuleiro State Park, where ombrophilous forest is the predominant vegetation (Bertho 2005Bertho AMM (2005) Os índios Guarani da Serra do Tabuleiro e a conservação da natureza (Uma perspetiva etnoambiental). Tese de Doutorado. Universidade Federal de Santa Catarina, Florianópolis. 224p.). According to the Köppen classification, coastal Santa Catarina State is subtropical with a hot summer (Cfa). The region has an average temperature of the coldest month below 18 ºC, average temperature of the hottest month above 22 ºC, infrequent frosts, a tendency for rainfall to be concentrated during the summer, and no defined dry season (Pandolfo et al. 2002Pandolfo C, Braga HJ, Silva JRVP, Massignam AM, Pereira ES, Thomé VMR & Valci FV (2002) Atlas climatológico digital do estado de Santa Catarina. Epagri, Florianópolis. 13p.).

a-b. Location of the study area - a. state of Santa Catarina; b. detail of the municipality of Palhoça, where the Mbyá-Guarani Morro dos Cavalos Indigenous Territory is located, in the Parque Estadual da Serra do Tabuleiro (P.E.S.T.).
Data collection
All species included in this study are used as important economic resources for Guarani, with no other reported uses besides for commercialization. It is possible that other uses of orchids for the Guarani have been kept undisclosed, to protect their own knowledge. The orchids species were selected based on interviews and guided tours with Guarani who collect them to commercialize at the Florianopolis downtown. For this, the following was needed: authorization from the community, in accordance with a FICF (Free and Informed Consent Form), authorization from the ethics committee at the Federal University of Santa Catarina, authorization from FUNAI (Fundação Nacional do Índio), and registration in SISBIO (Sistema de Autorização e Informação em Biodiversidade). During the guided tours, the indigenous collectors recognized plants in the natural environment and, when possible, collected at least three individuals of each species. The specimens collected were without flowers, so they were brought to the Botany Department at UFSC and maintained in the orchidarium until the appearance of flowers. When specimens were with flowers they were sent to specialists for identification (Tab. 1), and vouchers were deposited in the herbaria FLOR and Instituto Federal de Educação, Ciência e Tecnologia do Amazonas (EAFM).
Epiphytic species of Orchidaceae Epidendroideae subfamily occurring in indigenous territory in the Parque Estadual da Serra do Tabuleiro (P.E.S.T.), Santa Catarina, Brazil.
Anatomical analyses
For the anatomical analyses, regions of adult adventitious aerial roots 2 to 3 cm from the rhizome) and leaves from the third to the fifth node (middle third of the midrib) were fixed in FAA 70, (Johansen 1940Johansen DA (1940) Plant microtechnique. McGraw-Hill, New York. 523p.) in a vacuum for 24h and stocked in 70 % ethyl alcohol. After that the samples were pre-infiltrated in a solution of polyethylene glycol 1,500 U.S.P. (PEG) and 70 % ethyl alcohol at the proportion of 1:1 (v:v), in a dryer at 56 °C for 12 hours, and then infiltrated with pure PEG solution for 20 hours and embedded in PEG (Piazza et al. 2015Piazza LD, Smidt EC & Bona C (2015) Anatomia comparada dos órgãos vegetativos de espécies de Bulbophyllum seção Didactyle (Lindl.) Cogn. e Bulbophyllum seção. Xiphizusa Rchb.f. (Orchidaceae). Hoehnea 42: 171-183.). The blocks were sectioned (45 µm thick) using a rotary microtome (Leica RM 2235) with a steel blade. The sections were stained with 1% astra blue and 1% safranin and sealed with glycerinated gelatin (Kraus & Arduin 1997Kraus JE & Arduin M (1997) Manual básico de métodos em morfologia vegetal. EDUR, Rio de Janeiro. 198p.).
Results
Leaf
In all, 14 species were analyzed, seven belong to the tribe Cymbidieae and seven belong to tribe Epidendreae (Tab. 1). A pseudobulb was present in all species of Cymbidieae, except in Huntleya meleagris, and was absent in the species of Epidendreae, except Encyclia patens (Tab. 2). In all the species, the leaves are hypostomatic with stomata at the same level as the epidermal cells (Fig. 2a-c). The species Bifrenaria inodora, Gongora bufonia, Heterotaxis brasiliensis, Miltonia sp., Xylobium variegatum, Cattleya spp. and E. patens have a conspicuous stomatal crest that forms a suprastomatic chamber (Fig. 2c). Trichomes in depression were observed on both epidermis surfaces in B. inodora, G. bufonia, H. brasiliensis and E. patens (Fig. 2d-e; Tab. 2). The epidermis is unistratified, with square to rectangular (Fig. 2f) to round cells (Fig. 2a,d) that tend to be papillate, as in E. patens (Fig. 2c). The cuticle varies in thickness, is conspicuous in most species of both tribes (Tab. 2), such as Cattleya tigrina (Fig. 2f) and H. brasiliensis (Fig. 2g-h), and inconspicuous in G. bufonia, H. meleagris and X. variegatum of tribe Cymbidieae (Tab. 2) and E. patens (Fig. 2b-d; Tab. 2) of tribe Epidendreae.
Morpho-anatomical characteristics of epiphytic species of Orchidaceae Epidendroideae subfamily occurring in indigenous territory in the Parque Estadual da Serra do Tabuleiro (P.E.S.T.), Santa Catarina, Brazil. 1 = present; 0 = absent.

a-i. Transverse sections of the leaf of Orchidaceae epiphytic species - a-b. Huntleya meleagris - a. general structure; b. detail of the stomata at the same level as the other epidermal cells; c-e. Encyclia patens - c. detail of abaxial surface showing evident stomatal crest (arrowhead); d. mesophyll homogeneous with fibers extraxylary strands distributed randomly and vascular bundles near the adaxial surface; e. detail showing trichome; f. Cattleya tigrina - detail of the adaxial surface showing conspicuous cuticle and hypodermis with inverted “n” thickened wall cells (arrow); g-h. Heterotaxis brasiliensis - g. midrib with conspicuous water-storage parenchyma and a vascular bundle; h. detail showing epidermis of abaxial surface with conspicuous cuticle (empty arrow); i. Octomeria grandiflora - showing water-storage parenchyma occupying more than 80% of the leaf volume. Ab = abaxial epidermis; Ad = adaxial epidermis; C = cuticle (only on 2f); C = chlorenchyma; Ef = extraxylary fibers; Hp = hypodermis; Ph = phloem; Sc = sclerified cells; St = stomata; T = trichome; Vb = vascular bundle; X = xylem; Ws = water-storage cells. Scale: a-d, f-g = 100 µm; e, h = 50 µm.
The mesophyll is homogeneous in all studied species, with lobed or isodiametric chlorenchyma cells and few intercellular spaces (Figs. 2a,c,d,g; 3a-f). An adaxial hypodermis, mainly in the midrib, with water-storage parenchyma, is observed in most species (Fig. 2a,d,g,i; Tab. 2), except in B. inodora, G. bufonia, and X. variegatum of tribe Cymbidieae and Epidendrum vesicatum of Epidendreae (Fig. 3d-e; Tab. 2). The water-storage parenchyma is multiseriate and comprises elongated cells (Fig. 2a,d,g,i) and without any type of thickenings. In Octomeria grandiflora it is very conspicuous and accounts for more than 80% of the leaf volume (Fig. 2i). In C. tigrina, the hypodermis has cells with thick, ‘n’ shaped thickening walls on both surfaces of the epidermis (Fig. 2f). Extraxylary fiber strands in the mesophyll are common in the Cymbidieae species studied here, except in H. meleagris (Tab. 2). However, in Epidendreae, only E. patens and E. vesicatum have extraxylary fiber strands (Tab. 2). The fiber strands can occur in three forms: near the abaxial surface, in a single series (Fig. 3b), as observed in B. inodora, G. bufonia, H. brasiliensis, Miltonia flavescens, Miltonia spectabilis and E. vesicatum; near the epidermis of both surfaces, in two series, as in X. variegatum (Fig. 3c-d); or randomly distributed throughout the mesophyll, as in E. patens (Fig. 2d).

a-f. Transverse sections of the leaf of Orchidaceae epiphytic species - a. Octomeria grandiflora - chlorenchyma with two series of vascular bundles near the abaxial surface are observed; b. Gongora bufonia - homogeneous mesophyll and fibers extraxylary strands are observed near the abaxial surface; c-d. Xylobium variegatum - c. mesophyll with fibers extraxylary strands in two series, close to both surfaces; d. prominent midrib with three vascular bundles immersed in parenchyma cells that are surrounded by a sclerenchyma sheath; e. Gongora bufonia - prominent midrib with a central vascular bundle; f. Epidendrum vesicatum - midrib not prominent with a vascular bundle. Ab = abaxial epidermis; P = parenchyma; Ad = adaxial epidermis; C = chlorenchyma; Ef = extraxylary fiber; Ph = phloem; V = vascular bundle; Sc = sclerenchyma; T = trichome; X = xylem; Ws = water-storag clles. Scale: a-d = 100 µm.
The vascular system comprises collateral vascular bundles of different sizes, occurring in a series in the mesophyll (Figs. 2a,d,g; 3b), except in O. grandiflora that has two series of vascular bundles (Figs. 2i; 3a). Sclerified cells, possibly pericyclic fibers, completely surround the bundles in B. inodora, G. bufonia (Fig. 3b), X. variegatum (Fig. 3c), Cattleya intermedia, C. tigrina, E. patens and O. grandiflora (Fig. 3a), or partially surround the bundles in H. brasiliensis (Fig. 2g), H. meleagris (Fig. 2a), M. flavescens, M. spectabilis, Epidendrum ramosum, Epidendrum rigidum and E. vesicatum. The midrib of some species of Cymbidieae is prominent (Tab. 2) and may contain one collateral vascular bundle, as in B. inodora, G. bufonia (Fig. 3e) and H. meleagris, or three vascular bundles, as in X. variegatum (Fig. 3d), which are immersed in parenchyma cells that are surrounded by a sclerenchyma sheath. In the remaining Cymbidieae species and all of the Epidendreae species analyzed, the midrib is not prominent and has one vascular collateral bundle that can be partially surrounded by a sheath of sclerified cells (}Figs. 2a,g; 3f; Tab. 2).
Root
All the species studied have velamen that usually consists of elongated cells with reticulated thickenings without clear differentiation between epi and endovelamen (Fig. 4a-b,d-e) except in O. grandiflora (Fig. 4c) that have the endovelamen with a layer and the epivelamen with two and three layers of cells. In H. brasiliensis and M. flavecens (of tribe Cymbidieae) and C. intermedia, C. tigrina, E. vesicatum and O. grandiflora (of tribe Epidendreae), the velamen has up to four layers of cells (Fig. 4c,e; Tab. 2); in the remaining species there are more than four layers (Fig. 4a-b,d; Tab. 2). Root hairs were observed only in X. variegatum (Fig. 4a). Tilosomes occur in all species [Fig. 4a-b,d (arrow)] except in M. flavescens and O. grandiflora (Fig. 4c,e).
In the cortex, in all the species the exodermis is uniseriate with thin walls, except for O-shaped thickening in O. grandiflora (Fig. 4c; Tab. 2). Passage cells in the exodermis are common and can be observed in Miltonia spp., X. variegatum, Epidendrum spp. and O. grandiflora (Fig. 4a,c,e). The cortical parenchyma of the species studied have various layers of rounded cells, it is possible to observed cells with phi thickenings in B. inodora, G. bufonia and X. variegatum (Cymbidieae) (Fig. 4a-b,d; Tab. 2) and E. patens (Epidendreae), and isolated sclereids can be seen in H. meleagris, M. flavescens (Fig. 4e) and X. variegatum (Fig. 4a). Mycorrhizae were observed in O. grandiflora (Fig. 4c; Tab. 2) and H. brasiliensis, M. flavescens (Fig. 4e). The endodermis cells have thin or slightly thickened U-shaped or O-shaped walls with passage cells (Figs. 4a-f). The vascular cylinder of all studied species is polyarchy, with uniseriate pericycle of thick cells. Sclerified cells occur around the vascular elements and the pith is parenchymal except in. O. grandiflora (Fig. 4c), M. flavescens (Fig. 4e), M. spectabilis e G. bufonia.
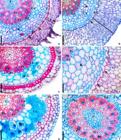
a-f. Transverse sections of the root of Orchidaceae epiphyte species - a. Xylobium variegatum - root hairs, velamen more than 4 layers, passage cells in the exodermis and cortex with sclerenchyma cells; b. Gongora bufonia - velamen with more than four layers and cortical cells with phi thickening; c. Octomeria grandiflora - velamen with three layers, thickened exodermis, cortical parenchyma with mycorrhizae; d. Bifrenaria inodora - tilosome and cortical cells with phi thickening are observed; e. Miltonia flavescens - cortical parenchyma with mycorrhizae and endodermis with passage cells; f. Bifrenaria inodora - polyartic vascular system and parenchymal pith. Pc = passage cells; Cx = cortex; Ed = endodermis; Ex = exodermis; My = mycorrhizae; Mx = metaxylema; P = pith; Pe = pericycle; Ph = phloem; Px = protoxylem; Rh = root hairs; Sc = sclereids; V = velamen; X = xylem. Phi thickening (arrow head); tilosomes (arrow). Scale: a-f = 100 µm.
Discussion
All of the species analyzed have hypostomatic leaves with stomates at the same level as the epidermal cells, characteristics common in Orchidaceae species (Yukawa & Stern 2002Yukawa T & Stern WL (2002) Comparative vegetative anatomy and systematics of Cymbidium (Cymbidieae: Orchidaceae). Botanical Journal of the Linnean Society 138: 383-419.; Zanenga-Godoy & Costa 2003; Silva et al. 2006Silva IV, Meira RMSA, Azevedo AA & Euclydes RMA (2006) Estratégias anatômicas foliares de treze espécies de Orchidaceae ocorrentes em um campo de altitude no Parque Estadual da Serra do Brigadeiro. Acta Botanica Brasilica 20: 741-750.). Some species have a suprastomatic chamber formed by conspicuous stomatal crests, which has been cited for epiphytic Orchidaceae species (Stern 2014) and is considered an adaptive characteristic that saves water (Oliveira & Sajo 1999Oliveira VC & Sajo MG (1999) Anatomia foliar de espécies epífitas de Orchidaceae. Revista Brasileira de Botânica 22: 365-374.).
Foliar trichomes of Orchidaceae species can be glandular or non-glandular, unicellular or multicellular and superficial or sunken; the most common type is sunken with up to five cells, with or without a glandular apex (Stern 2014Stern WL (2014) Anatomy of the monocotyledons Vol. X. Orchidaceae, Oxford University Press, New York. 288p.). In the present study, we observed trichomes in depression in four species; however, it was not possible to determine if the trichomes were glandular or non-glandular. According to Stern (2014), during development the trichome becomes sunken due to anticlinal divisions in the protoderm and eventual rupturing of the apical cell wall, resulting in a depression on the surface that is covered by an opaque brown residue. In addition, in Orchidaceae these trichomes do not function as significant absorption agents and can secrete mucilage that might help the lamina unfold (Pridgeon et al. 1983Pridgeon AM, Stern WL & Benzing DH (1983) Tilosomes in roots of Orchidaceae: morphology and systematic occurrence. American Journal of Botany 70: 1365-1377. DOI: <https://doi.org/10.1002/j.1537-2197.1983.tb07926.x>.
https://doi.org/10.1002/j.1537-2197.1983...
).
Variation in cuticle thickness in Orchidaceae species has been observed by other authors, such as Oliveira & Sajo (1999)Oliveira VC & Sajo MG (1999) Anatomia foliar de espécies epífitas de Orchidaceae. Revista Brasileira de Botânica 22: 365-374., Helbsing et al. (2000)Helbsing S, Riederer M & Zotz G. (2000) Cuticles of vascular epiphytes: efficient barriers for water loss after stomatal closure? Annals of Botany 86: 765-769., Riederer & Schreiber (2001)Riederer M & Schreiber L (2001) Protecting against water loss: analysis of the barrier properties of plant cuticles. Journal of Experimental Botany 52: 2023-2032., Oliveira & Sajo (2001) and Zanenga-Godoy & Costa (2003) that associate cuticle thickness to the environment and exposure to sun, where a thicker cuticle is related to more solar radiation. Other authors have used cuticle thickness as a characteristic to distinguish species (Rosso 1966Rosso SW (1966) The vegetative anatomy of the Cypripedioideae (Orchidaceae). Botanical Journal of the Linnean Society 59: 309-314. DOI: <https://doi.org/10.1016/j.landusepol.2016.08.032>.
https://doi.org/10.1016/j.landusepol.201...
; Oliveira Pires et al. 2003Oliveira-Pires MDF, Semir J, Melo De Pinna GFDA & Felix LP (2003) Taxonomic separation of the genera Prosthechea and Encyclia (Laeliinae: Orchidaceae) using leaf and root anatomical features. Botanical Journal of the Linnean Society 143: 293-303. DOI: <https://doi.org/10.1046/j.1095-8339.2003.00195.x>.
https://doi.org/10.1046/j.1095-8339.2003...
). In this study we observed that most species have a conspicuous cuticle, which is a character related to resistance to desiccation, as classified by Oliveria & Sajo (1999).
A pseudobulb is common in epiphytic species of Orchidaceae (Stern 2014Stern WL (2014) Anatomy of the monocotyledons Vol. X. Orchidaceae, Oxford University Press, New York. 288p.), is an important organ that stores water, carbohydrates and minerals (Ng & Hew 2000Ng CKY & Hew CS (2000) Orchid pseudobulbs - “false” bulbs with a genuine importance in orchid growth and survival! Scientia Horticulturae 83: 165-172.), and contributes to the survival capacity of these plants (Esau 1977Esau K (1977) Anatomy of seed plants. John Wiley & Sons, New York. 550p.; Pridgeon et al. 1983Pridgeon AM, Stern WL & Benzing DH (1983) Tilosomes in roots of Orchidaceae: morphology and systematic occurrence. American Journal of Botany 70: 1365-1377. DOI: <https://doi.org/10.1002/j.1537-2197.1983.tb07926.x>.
https://doi.org/10.1002/j.1537-2197.1983...
; Benzing 1986Benzing DH (1986) The vegetative basis of vascular epiphytism. Selbyana 9: 23-43.; Loeschen et al. 1993Loeschen VS, Martin CE, Smith M & Eder SL (1993) Leaf anatomy and and CO2 recycling during Crassulacean Acid Metabolism in twelve epiphytic species of Tillandsia (Bromeliaceae). International Journal of Plant Sciences 154: 100-106. DOI: <https://doi.org/10.1086/297095>.
https://doi.org/10.1086/297095...
; Silva et al. 2006Silva IV, Meira RMSA, Azevedo AA & Euclydes RMA (2006) Estratégias anatômicas foliares de treze espécies de Orchidaceae ocorrentes em um campo de altitude no Parque Estadual da Serra do Brigadeiro. Acta Botanica Brasilica 20: 741-750.). Similarly, a hypodermis with water-storage parenchyma is common in these species (Pridgeon et al. 1983; Oliveira & Sajo 1999Oliveira VC & Sajo MG (1999) Anatomia foliar de espécies epífitas de Orchidaceae. Revista Brasileira de Botânica 22: 365-374.; Dettke & Milaneze-Gutierre 2008Dettke GA & Milaneze-Gutierre MA (2008) Anatomia caulinar de espécies epífitas de Cactaceae, subfamília Cactoideae. Hoehnea 35: 583-595.; Costa 2014Costa TS (2014) Caracterização anato-fisiológica de Cattleya guttata Lindl. e Laelia tenebrosa Rolf. (Orchidaceae): efeito da radiação. Dissertação de Mestrado. Universidade Federal do Espírito Santo, Vítoria. 62p.; Ayensu & Williams 1972Ayensu ES & Williams NH (1972) Leaf anatomy of Palumbina and Odontoglossum subgenus Osmoglossum. American Orchid Society bulletin 41: 687-696.). In the present study, most of the species only have a hypodermis in the region of the midrib, except for O. grandiflora whose hypodermis is very conspicuous and occurs throughout the leaf blade, abaxially, and occupies more than 80% of the leaf volume. In C. tigrina, a hypodermis with thickened cell walls was observed on both leaf surfaces. The same was observed in other species of Cattleya by Costa (2014) and Carneiro et al. (2017)Carneiro GT, Gonçalves LM & Flores AS (2017) Anatomia foliar de duas espécies de Cattleya (Orchidaceae) endêmicas dos Escudos das Guianas. Biota Amazônia 7: 26-29., which might mean this is a taxonomic characteristic of the genus. According to Stern (2014), in species of Orchidaceae, the composition of the hypodermis is parenchyma and/or sclerenchyma that occur as either continuous or interrupted layers. The parenchymatous layers store water and the sclerenchyma fibers provide mechanical support.
Extraxylary fiber strands observed in the mesophyll in some of the species analyzed are common in Orchidaceae epiphytes and normally are associated with silica bodies (stegmata) (Oliveira & Sajo 1999Oliveira VC & Sajo MG (1999) Anatomia foliar de espécies epífitas de Orchidaceae. Revista Brasileira de Botânica 22: 365-374.); however, in the present study stegmata were not observed. These cells create mechanical resistance during dehydration that is comparable to coriaceous leaves (Oliveira & Sajo 1999; Dettke & Milaneze-Gutierre 2008Dettke GA & Milaneze-Gutierre MA (2008) Anatomia caulinar de espécies epífitas de Cactaceae, subfamília Cactoideae. Hoehnea 35: 583-595.; Silva et al. 2010Silva IV, Meira RMSA & Azevedo AA (2010) Anatomia de raízes de espécies de Orchidaceae do Parque Estadual da Serra do Brigadeiro, Minas Gerais. Hoehnea 37: 147-161.). All the species analyzed have sclerified cells that completely or partially surround the bundles. These cells are cited by Oliveira & Sajo (1999) as pericyclic fibers and, like extraxylary fibers, are common in epiphytic Orchidaceae species.
All of the species studied have multiseriate velamen. Velamen is an important tissue in Orchidaceae species that is related to epiphytism (Benzing 2008Benzing DH (2008) Vascular epiphytes. General biology and related biota. Vol. 1. Ed. Cambridge University Press, New York. 295p.) and absorbs water and minerals, reduces transpiration rates, incidence of light and loss of water in the cortex, and increases mechanical protection (Pridgeon et al. 1983Pridgeon AM, Stern WL & Benzing DH (1983) Tilosomes in roots of Orchidaceae: morphology and systematic occurrence. American Journal of Botany 70: 1365-1377. DOI: <https://doi.org/10.1002/j.1537-2197.1983.tb07926.x>.
https://doi.org/10.1002/j.1537-2197.1983...
; Benzing 1986; Joca et al. 2017Joca TAC, Oliveira DC, Zotz G, Winkler U & Moreira ASFP (2017) The velamen of epiphytic orchids: variation in structure and correlations with nutrient absorption. Flora 230: 66-74. DOI: <https://doi.org/10.1016/j.flora.2017.03.009>.
https://doi.org/10.1016/j.flora.2017.03....
). The number of velamen layers can have diagnostic value in Orchidaceae; although, Orchidaceae species can exhibit high phenotypic plasticity for this character (Sanford & Adanlawo 1973Sanford WW & Adanlawo I (1973) Velamen and exodermis characters of West African epiphytic orchids in relation to taxonomic grouping and habitat tolerance. Botanical Journal of the Linnean Society 66: 307-321. DOI: <https://doi.org/10.1111/j.1095-8339.1973.tb02178.x>.
https://doi.org/10.1111/j.1095-8339.1973...
; Pita & Menezes 2002Pita PB & Menezes NL (2002) Anatomia da raiz de espécies de Dyckia Schult. f. e Encholirium Mart. ex Schult. & Schult. f. (Bromeliaceae, Pitcairnioideae) da Serra do Cipó (Minas Gerais, Brasil), com especial referência ao velame. Revista Brasileira de Botânica 25: 25-34. DOI: <https://doi.org/10.1590/S0100-84042002000100005>.
https://doi.org/10.1590/S0100-8404200200...
; Silva & Milaneze-Gutierra 2004Silva CI & Milaneze-Gutierra MA (2004) Caracterização morfo-anatômica dos órgãos vegetativos de Cattleya walkeriana Gardner (Orchidaceae). Acta Scientiarum Biological Sciences 26: 91-100. DOI: <https://doi.org/10.4025/actascibiolsci.v26i1.1664>.
https://doi.org/10.4025/actascibiolsci.v...
; Mayer et al. 2008Mayer JLS, Ribas LLF, Bona C & Quoirin M (2008) Anatomia comparada das folhas e raízes de Cymbidium Hort. (Orchidaceae) cultivadas ex vitro e in vitro. Acta Botanica Brasilica 22: 323-332. DOI: <https://doi.org/10.1590/S0102-33062008000200003>.
https://doi.org/10.1590/S0102-3306200800...
; Joca et al. 2017). Here, the only species with clear differentiation between epi- and endovelame was O. grandiflora, the other species the distinction between epi- and endovelamen are not as clear. Agreement with Porembski & Barthlott (1988)Porembski S & Barthlott W (1988) Velamen radicum micromorphology and classification of Orchidaceae. Nordic Journal of Botany 8: 117-137. O. grandiflora has the Pleurothallis type and the other species the Cymbidium type velamen is the type that we think more similar.
Most of the species here analyzed have tilosomes. These are thickenings from the walls of cells of the innermost velamen cell layer adjacent to thin-walled passage cells of the exodermis in roots of many epiphytic orchids (Pridgeon et al. 1983Pridgeon AM, Stern WL & Benzing DH (1983) Tilosomes in roots of Orchidaceae: morphology and systematic occurrence. American Journal of Botany 70: 1365-1377. DOI: <https://doi.org/10.1002/j.1537-2197.1983.tb07926.x>.
https://doi.org/10.1002/j.1537-2197.1983...
; Silva et al. 2010Silva IV, Meira RMSA & Azevedo AA (2010) Anatomia de raízes de espécies de Orchidaceae do Parque Estadual da Serra do Brigadeiro, Minas Gerais. Hoehnea 37: 147-161.; Stern 2014Stern WL (2014) Anatomy of the monocotyledons Vol. X. Orchidaceae, Oxford University Press, New York. 288p.; Kedrovski & Sajo 2019Kedrovski HR & Sajo MG (2019) What are tilosomes? An update and new perspectives. Acta Botanica Brasilica 33: 106-115.). It is believed that tilosomes are associated with the epiphytic habit of Orchidaceae species and involved in the absorption of water, increasing the contact surface of the inner part of the velamen with the atmosphere, as well as preventing water loss via the passage cells in the exodermis, since their composition is mostly lignin, a highly hydrophobic phenolic compound (Pridgeon et al. 1983; Benzing 1986Benzing DH (1986) The vegetative basis of vascular epiphytism. Selbyana 9: 23-43., 2008; Silva et al. 2010). Kedrovski & Sajo (2019) suggest that tilosomes in Anathallis sclerophylla (Lindl.) have two functional phases: in young tissues they increase the symplast connection and thus improve outside-inside transport and in mature tissues they direct solutes to passage cells. According to Pridgeon et al. (1983) seven broad morphological types are recognized which can characterize specific orchid subtribes. Species Cymbidium type velamen show a webbed or meshed tilosomes structure (Pridgeon et al. 1983).
The main functions of the root exodermis are to control the passage of nutrients and prevent the loss of water in the cortex (Benzing 2008Benzing DH (2008) Vascular epiphytes. General biology and related biota. Vol. 1. Ed. Cambridge University Press, New York. 295p.; Silva & Milaneze-Gutierre 2004Silva CI & Milaneze-Gutierra MA (2004) Caracterização morfo-anatômica dos órgãos vegetativos de Cattleya walkeriana Gardner (Orchidaceae). Acta Scientiarum Biological Sciences 26: 91-100. DOI: <https://doi.org/10.4025/actascibiolsci.v26i1.1664>.
https://doi.org/10.4025/actascibiolsci.v...
). In all of the species studied, except O. grandiflora, the exodermis has thin walls with visible passage cells. In Orchidaceae, wall thickness in the exodermis varies (Benzing 1986; Silva et al. 2010). The species with thin exodermis walls had tilosomes, corroborating what was observed by Benzing (2008).
Some of the analyzed species have root cortical cells with phi thickenings and/or sclereids. The phi thickenings are lignified and form wide bands that are sometimes ramified and generally form a continuous sheath in the center of the cortical region of the roots (Oliveira Pires et al. 2003Oliveira-Pires MDF, Semir J, Melo De Pinna GFDA & Felix LP (2003) Taxonomic separation of the genera Prosthechea and Encyclia (Laeliinae: Orchidaceae) using leaf and root anatomical features. Botanical Journal of the Linnean Society 143: 293-303. DOI: <https://doi.org/10.1046/j.1095-8339.2003.00195.x>.
https://doi.org/10.1046/j.1095-8339.2003...
). Phi thickenings have already been cited for other Orchidaceae species (Oliveira Pires et al. 2003; Moreira & Isaias 2008Moreira ASFP & Isaias RM (2008) Comparative Anatomy of the absorption roots of terrestrial and epiphytic Orchids. Brazilian Archives of Biology and Technology 51: 11-17.; Idris & Collings 2015Idris NA & Collings DA (2015) The life of phi: the development of phi thickenings in roots of the orchids of the genus Miltoniopsis. Planta 241: 489-506. DOI: <https://doi.org/10.1007/s00425-014-2194-z>.
https://doi.org/10.1007/s00425-014-2194-...
), as well as other families, such as Geraniaceae (Haas et al. 1976Haas DL, Carothers ZB, Robbins RR, Journal A, Jul N & Robbins R (1976) Observations on the Phi-Thickenings and Casparian Strips in Pelargonium Roots. American Journal of Botany 63: 863-867. DOI: <https://doi.org/10.2307/2442047>.
https://doi.org/10.2307/2442047...
), Rosaceae (Weerdenburg & Peterson 1983Weerdenburg CA & Peterson CA (1983) Structural changes in phi thickenings during primary and secondary growth in roots Apple (Pyrus malus) Rosaceae. Canadian Journal of Botany 61: 2570-2576. DOI: <https://doi.org/10.1139/b83-282>.
https://doi.org/10.1139/b83-282...
) and Brassicaceae (Fernandez-Garcia et al. 2009Fernandez-Garcia N, Lopez-Perez L, Hernandez M & Olmos E (2009) Role of phi cells and the endodermis under salt stress in Brassica oleracea. New Phytologist 18: 347-360.), and gymnosperm species (Gerrath et al. 2005Gerrath JM, Matthes U, Purich M & Larson DW (2005) Root environmental effects on phi thickening production and root morphology in three gymnosperms. Canadian Journal of Botany 83: 379-385. DOI: <https://doi.org/10.1139/b05-011>.
https://doi.org/10.1139/b05-011...
). According to Idris & Collings (2015), the function of phi thickenings in orchid species is still poorly understood and could be associated with water stress, keeping plant tissue from collapsing when under pressure from a lack of water, pathogen attacks and providing mechanical protection; although, some works have shown that they are not very effective against water loss (Zanenga-Godoy & Costa 2003; Piazza et al. 2015Piazza LD, Smidt EC & Bona C (2015) Anatomia comparada dos órgãos vegetativos de espécies de Bulbophyllum seção Didactyle (Lindl.) Cogn. e Bulbophyllum seção. Xiphizusa Rchb.f. (Orchidaceae). Hoehnea 42: 171-183.; Joca et al. 2017Joca TAC, Oliveira DC, Zotz G, Winkler U & Moreira ASFP (2017) The velamen of epiphytic orchids: variation in structure and correlations with nutrient absorption. Flora 230: 66-74. DOI: <https://doi.org/10.1016/j.flora.2017.03.009>.
https://doi.org/10.1016/j.flora.2017.03....
).
The endodermis is a highly specialized, single layer of cells that comprises the innermost layer of the cortex that surrounds the stele (Beck 2005Beck CB (2005) An introduction to plant structure and development. Vol. 1. Cambridge University Press, Cambridge. 459p.) and is an important apoplastic barrier that prevents water and ion loss and protects the vascular cylinder against pathogens through Casparian band and wall thickening (Muthukumar & Kowsalya 2017Muthukumar T & Kowsalya A (2017) Comparative anatomy of aerial and substrate roots of Acampe praemorsa (Rox.) Blatt. & McCann. Flora 226: 17-28.). Studies have shown that environmental factors such as drought tend to promote cell wall augmentation of the endodermis and reduce or eliminate passage cells (Peterson & Enstone 1996Peterson CA & Enstone DE (1996) Function of passage cells in the endodermis and exodermis of roots. Physiologia Plantarum 97: 592-598.). Here the species analyzed were mostly thin or slightly thickened wall endodermis with evident passage cells, showing low resistance to desiccation. Moreover, all species have thick walled and sclerenchymatous cells around vascular elements and pith parenchyma except four of them, which have slightly sclerified pith cells. The thickening of cells is a common feature of plant roots growing in arid regions to cope up with desiccation and to avoid cell collapse, in aerial roots, they appear to provide mechanical strength to the root (Muthukumar & Kowsalya 2017).
A common characteristic in epiphytic Orchidaceae is the presence of fungal hyphae, which are involved in forming mycorrhizae (Cevallos et al. 2017Cevallos S, Sánchez-rodríguez A, Decock C, Declerck S & Suárez JP (2017) Are there keystone mycorrhizal fungi associated to tropical epiphytic orchids? Mycorrhiza 27: 225-232. DOI: <https://doi.org/10.1007/s00572-016-0746-8>.
https://doi.org/10.1007/s00572-016-0746-...
; Joca et al. 2017Joca TAC, Oliveira DC, Zotz G, Winkler U & Moreira ASFP (2017) The velamen of epiphytic orchids: variation in structure and correlations with nutrient absorption. Flora 230: 66-74. DOI: <https://doi.org/10.1016/j.flora.2017.03.009>.
https://doi.org/10.1016/j.flora.2017.03....
). Mycorrhizae in epiphytic Orchidaceae species have been described in other works (Cevallos et al. 2017) and are the result of a mutualistic symbiotic relationship where the fungus helps absorb organic and inorganic nutrients (Boldrini et al. 2010Boldrini RF, Santos WO, Cruz ZM & Ramos AC (2010) Bases da associação micorrízica orquidóide Bases of the orchidoid mycorrhizal association. Natureza on Line 8: 140-145.; Cevallos et al. 2017). Recent studies have shown the importance of the presence of fungi to the survival of epiphytic species, mainly in relation to seed development and germination of orchids (Gonçalves et al. 2014; Marillac et al. 2014Marillac C, Nunes C & Araújo LG (2014) Isolation and characterization of mycorrhizal fungi in Cyrtopodium vernum Rchb.F. & Warm (Orchidaceae). Revista Da Ciência Agraria 57: 244-249.).
As discussed, the Orchidaceae species analyzed in the present study exhibit different morphoanatomical adaptations that are used to save water due to their epiphytic habit. These characteristics can occur in different parts of the root, stem and leaf. Among the adaptive characters highlighted in this work, we can distinguish those related to storing water and those related to resistance to desiccation (Oliveira & Sajo 1999Oliveira VC & Sajo MG (1999) Anatomia foliar de espécies epífitas de Orchidaceae. Revista Brasileira de Botânica 22: 365-374.). In the first case, we cite the presence of pseudobulbs and a hypodermis with water-storage cells. In the second case, we note the conspicuous cuticle, suprastomatic chamber and extraxylary and pericyclic fibers in the leaves, and tilosomes and/or exodermal thickenings and cortex cells with phi thickenings or sclereids in the root.
All of the species studied have pseudobulbs or leaves with a hypodermis with water-storage cells, except E. vesicatum that lacks characters related to water storage. For the characteristics related to resistance to desiccation, most of the species analyzed have four to six attributes, accept H. meleagris and E. ramosun that have only three and E. rigidum that has only two; these species do not have pseudobulbs, only a hypodermis with water-storage cells in the midrib of the leaf. Our results suggest that E. vesicatum, H. meleagris, E. ramosun and E. rigidum are less tolerant to hydric stress compared to the other species analyzed. By learning more about the adaptive characteristics of the species studied here, we can learn more about how to conserve them and more efficient cultivation strategies.
Acknowledgements
The authors thank the specialists Carlos Eduardo de Siqueira, Luís Funez and Nestor Fabio Alzate, for identifying the species; the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), for the scholarship awarded to the first author; the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), for the financial support and productivity research grant awarded to the third author; and FAPESC (covenant 11320/2012-4).
References
- Avi RC & Rodrigues AC (2019) Comparative anatomy of the leaf and ramicaule in Pleurothallidinae (Orchidaceae) species. The Journal of the Torrey Botanical Society 146: 198-212.
- Ayensu ES & Williams NH (1972) Leaf anatomy of Palumbina and Odontoglossum subgenus Osmoglossum American Orchid Society bulletin 41: 687-696.
- Balachandar M, Koshila Ravi R, Ranjithamani A & Muthukumar T (2019) Comparative vegetative anatomy and mycorrhizal morphology of three South Indian Luisia species (Orchidaceae) with the note on their epiphytic adaptations. Flora 251: 39-6.
- Beck CB (2005) An introduction to plant structure and development. Vol. 1. Cambridge University Press, Cambridge. 459p.
- Benzing DH (1986) The vegetative basis of vascular epiphytism. Selbyana 9: 23-43.
- Benzing DH (2008) Vascular epiphytes. General biology and related biota. Vol. 1. Ed. Cambridge University Press, New York. 295p.
- Bertho AMM (2005) Os índios Guarani da Serra do Tabuleiro e a conservação da natureza (Uma perspetiva etnoambiental). Tese de Doutorado. Universidade Federal de Santa Catarina, Florianópolis. 224p.
- Bhatnagar M, Sarkar N, Gandharv N, Apang O, Singh S & Ghosal S (2017) Evaluation of antimycobacterial, leishmanicidal and antibacterial activity of three medicinal orchids of Arunachal. Complementary and Alternative Medicine 17: 1-10. DOI: <https://doi.org/10.1186/s12906-017-1884-z>
» https://doi.org/10.1186/s12906-017-1884-z - Blanco GD (2017) Estudo etnobotânico e morfoanatômico de espécies de Orchidaceae utilizadas por grupos Guarani. Dissertação de Mestrado. Universidade Federal de Santa Catarina, Florianopolis. 163p.
- Boldrini RF, Santos WO, Cruz ZM & Ramos AC (2010) Bases da associação micorrízica orquidóide Bases of the orchidoid mycorrhizal association. Natureza on Line 8: 140-145.
- Bonates LCM (1993) Estudos ecofisiológicos de Orchidaceae da Amazônia II - anatomia ecológica foliar de espécies com metabolismo CAM de uma campina da Amazôniza central. Acta Amazonica 23: 315-348.
- Carneiro GT, Gonçalves LM & Flores AS (2017) Anatomia foliar de duas espécies de Cattleya (Orchidaceae) endêmicas dos Escudos das Guianas. Biota Amazônia 7: 26-29.
- Cevallos S, Sánchez-rodríguez A, Decock C, Declerck S & Suárez JP (2017) Are there keystone mycorrhizal fungi associated to tropical epiphytic orchids? Mycorrhiza 27: 225-232. DOI: <https://doi.org/10.1007/s00572-016-0746-8>.
» https://doi.org/10.1007/s00572-016-0746-8 - Chase MW, Cameron KM, Freudenstein JV, Pridgeon AM, Salazar G, Van den Berg C & Schuiteman A (2015) An updated classification of Orchidaceae. Botanical Journal of the Linnean Society 177: 151-174. DOI: <https://doi.org/10.1111/boj.12234>.
» https://doi.org/10.1111/boj.12234 - Chinsamy M, Finnie JF & Van Staden J (2011) A etnobotânica das orquídeas medicinais sul-africanas. Jornal Sul-Africano de Botânica 77: 2-9. DOI: <https://doi.org/10.1016/j.sajb.2010.09.015>
» https://doi.org/10.1016/j.sajb.2010.09.015 - Costa TS (2014) Caracterização anato-fisiológica de Cattleya guttata Lindl. e Laelia tenebrosa Rolf. (Orchidaceae): efeito da radiação. Dissertação de Mestrado. Universidade Federal do Espírito Santo, Vítoria. 62p.
- Dettke GA & Milaneze-Gutierre MA (2008) Anatomia caulinar de espécies epífitas de Cactaceae, subfamília Cactoideae. Hoehnea 35: 583-595.
- Dressler R (2005) How many Orchid species? Selbyana 26: 155-158.
- Duarte MM & Gandolfi S (2017) Forest ecology and management diversifying growth forms in tropical forest restoration: enrichment with vascular epiphytes. Forest Ecology and Management 401: 89-98. DOI: <https://doi.org/10.1016/j.foreco.2017.06.063>.
» https://doi.org/10.1016/j.foreco.2017.06.063 - Esau K (1977) Anatomy of seed plants. John Wiley & Sons, New York. 550p.
- Fernandez-Garcia N, Lopez-Perez L, Hernandez M & Olmos E (2009) Role of phi cells and the endodermis under salt stress in Brassica oleracea New Phytologist 18: 347-360.
- Foge BA, Essomo SE, Bechem TE, Tabot PT, Arrey BD, Afanga Y & Assoua EM (2019) Market trends and ethnobotany of orchids of Mount Cameroon. Journal Ethnobiology Ethnomedicine 15: 2-11.
- Gerrath JM, Matthes U, Purich M & Larson DW (2005) Root environmental effects on phi thickening production and root morphology in three gymnosperms. Canadian Journal of Botany 83: 379-385. DOI: <https://doi.org/10.1139/b05-011>.
» https://doi.org/10.1139/b05-011 - Haas DL, Carothers ZB, Robbins RR, Journal A, Jul N & Robbins R (1976) Observations on the Phi-Thickenings and Casparian Strips in Pelargonium Roots. American Journal of Botany 63: 863-867. DOI: <https://doi.org/10.2307/2442047>.
» https://doi.org/10.2307/2442047 - Hassemer G, Giovanni R & Trevisan R (2015) The use of potential distribution models in the study of the distribution and conservation status of plants: the case of Plantago L. (Plantaginaceae) in Brazil. The Journal of the Torrey Botanical Society 143: 1-38.
- Helbsing S, Riederer M & Zotz G. (2000) Cuticles of vascular epiphytes: efficient barriers for water loss after stomatal closure? Annals of Botany 86: 765-769.
- Hinsley A, Hugo JB, Michael FF, Stephan WG, Lauren M, Gardiner RSG, Pankaj K, Susanne M, Destario M, David LR, Sarina V, Shan W & Jacob P (2018) A review of the trade in orchids and its implications for conservation. Botanical Journal of the Linnean Society 8: 435-455.
- Idris NA & Collings DA (2015) The life of phi: the development of phi thickenings in roots of the orchids of the genus Miltoniopsis Planta 241: 489-506. DOI: <https://doi.org/10.1007/s00425-014-2194-z>.
» https://doi.org/10.1007/s00425-014-2194-z - Joca TAC, Oliveira DC, Zotz G, Winkler U & Moreira ASFP (2017) The velamen of epiphytic orchids: variation in structure and correlations with nutrient absorption. Flora 230: 66-74. DOI: <https://doi.org/10.1016/j.flora.2017.03.009>.
» https://doi.org/10.1016/j.flora.2017.03.009 - Johansen DA (1940) Plant microtechnique. McGraw-Hill, New York. 523p.
- Kedrovski HR & Sajo MG (2019) What are tilosomes? An update and new perspectives. Acta Botanica Brasilica 33: 106-115.
- Kersten RDA (2010) Epífitas vasculares - histórico, participação taxonômica e aspectos relevantes, com ênfase na Mata Atlântica. Hoehnea 37: 9-38.
- Koch AK, Santos JUM & Ilkiu-Borges AL (2014) Sinopse das Orchidaceae holoepífitas e hemiepífitas da Floresta Nacional de Caxiuanã, PA, Brasil. Hoehnea 41: 129-148.
- Kraus JE & Arduin M (1997) Manual básico de métodos em morfologia vegetal. EDUR, Rio de Janeiro. 198p.
- Loeschen VS, Martin CE, Smith M & Eder SL (1993) Leaf anatomy and and CO2 recycling during Crassulacean Acid Metabolism in twelve epiphytic species of Tillandsia (Bromeliaceae). International Journal of Plant Sciences 154: 100-106. DOI: <https://doi.org/10.1086/297095>.
» https://doi.org/10.1086/297095 - Marillac C, Nunes C & Araújo LG (2014) Isolation and characterization of mycorrhizal fungi in Cyrtopodium vernum Rchb.F. & Warm (Orchidaceae). Revista Da Ciência Agraria 57: 244-249.
- Mayer JLS, Ribas LLF, Bona C & Quoirin M (2008) Anatomia comparada das folhas e raízes de Cymbidium Hort. (Orchidaceae) cultivadas ex vitro e in vitro Acta Botanica Brasilica 22: 323-332. DOI: <https://doi.org/10.1590/S0102-33062008000200003>.
» https://doi.org/10.1590/S0102-33062008000200003 - Moreira ASFP & Isaias RM (2008) Comparative Anatomy of the absorption roots of terrestrial and epiphytic Orchids. Brazilian Archives of Biology and Technology 51: 11-17.
- Muthukumar T & Kowsalya A (2017) Comparative anatomy of aerial and substrate roots of Acampe praemorsa (Rox.) Blatt. & McCann. Flora 226: 17-28.
- Nadkarni NM (1984) Epiphyte biomass and nutrient capital of a Neotropical Elfin Forest. Biotropica 16: 249. DOI: <https://doi.org/10.2307/2387932>
» https://doi.org/10.2307/2387932 - Ng CKY & Hew CS (2000) Orchid pseudobulbs - “false” bulbs with a genuine importance in orchid growth and survival! Scientia Horticulturae 83: 165-172.
- Oliveira-Pires MDF, Semir J, Melo De Pinna GFDA & Felix LP (2003) Taxonomic separation of the genera Prosthechea and Encyclia (Laeliinae: Orchidaceae) using leaf and root anatomical features. Botanical Journal of the Linnean Society 143: 293-303. DOI: <https://doi.org/10.1046/j.1095-8339.2003.00195.x>.
» https://doi.org/10.1046/j.1095-8339.2003.00195.x - Oliveira VC & Sajo MG (1999) Anatomia foliar de espécies epífitas de Orchidaceae. Revista Brasileira de Botânica 22: 365-374.
- Oliveira VC & Sajo MG (2001) Morfo-anatomia caulinar de nove espécies de Orchidaceae. Acta Botanica Brasilica 15: 176-188.
- Pandolfo C, Braga HJ, Silva JRVP, Massignam AM, Pereira ES, Thomé VMR & Valci FV (2002) Atlas climatológico digital do estado de Santa Catarina. Epagri, Florianópolis. 13p.
- Peterson CA & Enstone DE (1996) Function of passage cells in the endodermis and exodermis of roots. Physiologia Plantarum 97: 592-598.
- Piazza LD, Smidt EC & Bona C (2015) Anatomia comparada dos órgãos vegetativos de espécies de Bulbophyllum seção Didactyle (Lindl.) Cogn. e Bulbophyllum seção. Xiphizusa Rchb.f. (Orchidaceae). Hoehnea 42: 171-183.
- Pita PB & Menezes NL (2002) Anatomia da raiz de espécies de Dyckia Schult. f. e Encholirium Mart. ex Schult. & Schult. f. (Bromeliaceae, Pitcairnioideae) da Serra do Cipó (Minas Gerais, Brasil), com especial referência ao velame. Revista Brasileira de Botânica 25: 25-34. DOI: <https://doi.org/10.1590/S0100-84042002000100005>.
» https://doi.org/10.1590/S0100-84042002000100005 - Pridgeon AM, Stern WL & Benzing DH (1983) Tilosomes in roots of Orchidaceae: morphology and systematic occurrence. American Journal of Botany 70: 1365-1377. DOI: <https://doi.org/10.1002/j.1537-2197.1983.tb07926.x>.
» https://doi.org/10.1002/j.1537-2197.1983.tb07926.x - Porembski S & Barthlott W (1988) Velamen radicum micromorphology and classification of Orchidaceae. Nordic Journal of Botany 8: 117-137.
- Riederer M & Schreiber L (2001) Protecting against water loss: analysis of the barrier properties of plant cuticles. Journal of Experimental Botany 52: 2023-2032.
- Rosso SW (1966) The vegetative anatomy of the Cypripedioideae (Orchidaceae). Botanical Journal of the Linnean Society 59: 309-314. DOI: <https://doi.org/10.1016/j.landusepol.2016.08.032>.
» https://doi.org/10.1016/j.landusepol.2016.08.032 - Sanford WW & Adanlawo I (1973) Velamen and exodermis characters of West African epiphytic orchids in relation to taxonomic grouping and habitat tolerance. Botanical Journal of the Linnean Society 66: 307-321. DOI: <https://doi.org/10.1111/j.1095-8339.1973.tb02178.x>.
» https://doi.org/10.1111/j.1095-8339.1973.tb02178.x - Silva CI & Milaneze-Gutierra MA (2004) Caracterização morfo-anatômica dos órgãos vegetativos de Cattleya walkeriana Gardner (Orchidaceae). Acta Scientiarum Biological Sciences 26: 91-100. DOI: <https://doi.org/10.4025/actascibiolsci.v26i1.1664>.
» https://doi.org/10.4025/actascibiolsci.v26i1.1664 - Silva IV, Meira RMSA & Azevedo AA (2010) Anatomia de raízes de espécies de Orchidaceae do Parque Estadual da Serra do Brigadeiro, Minas Gerais. Hoehnea 37: 147-161.
- Silva IV, Meira RMSA, Azevedo AA & Euclydes RMA (2006) Estratégias anatômicas foliares de treze espécies de Orchidaceae ocorrentes em um campo de altitude no Parque Estadual da Serra do Brigadeiro. Acta Botanica Brasilica 20: 741-750.
- Siqueira CE, Zanin A & Neto LM (2014) Orchidaceae in Santa Catarina: update, geographic distribution and conservation. Check List 10: 1452-1478.
- Stehmann JR, Forzza RC, Salino A, Sobral M, Costa DP & Kamino LHY (Org.) (2009) Plantas da Floresta Atlântica. Instituto de Pesquisa Jardim Botânico do Rio de Janeiro, Rio de Janeiro. 500p.
- Stern WL (2014) Anatomy of the monocotyledons Vol. X. Orchidaceae, Oxford University Press, New York. 288p.
- Swarts ND & Dixon KW (2009) Terrestrial orchid conservation in the age of extinction. Annals of Botany 2: 543-556. DOI: <https://doi.org/10.1093/aob/mcp025>.
» https://doi.org/10.1093/aob/mcp025 - Warmling J (2013) Anatomia da folha e ramicaule de espécies da subtribo Pleurothallidinae (Orchidaceae). Dissertação de Mestrado. Universidade Federal de Santa Catarina, Florianópolis. 109p.
- Weerdenburg CA & Peterson CA (1983) Structural changes in phi thickenings during primary and secondary growth in roots Apple (Pyrus malus) Rosaceae. Canadian Journal of Botany 61: 2570-2576. DOI: <https://doi.org/10.1139/b83-282>.
» https://doi.org/10.1139/b83-282 - Yukawa T & Stern WL (2002) Comparative vegetative anatomy and systematics of Cymbidium (Cymbidieae: Orchidaceae). Botanical Journal of the Linnean Society 138: 383-419.
- Zanenga-Godoy R & Costa CG (2003) Anatomia foliar de quatro espécies do gênero Cattleya Lindl. (Orchidaceae) do planalto central brasileiro. Acta Amazonica 17: 101-118.
- Zotz G & Hietz P (2001) The physiological ecology of vascular epiphytes: current knowledge, open questions. Journal of Experimental Botany 52: 2067-2078.
Edited by
Publication Dates
-
Publication in this collection
30 Apr 2021 -
Date of issue
2021
History
-
Received
12 Nov 2019 -
Accepted
29 Mar 2020




