Abstract:
Several studies have successfully used polyethylene glycol (PEG) and/or abscisic acid (ABA) to reinduce desiccation tolerance in germinated seeds of orthodox species, improving our understanding of the mechanisms that underlie this trait. In this context, treatments with PEG and ABA were tested to assess their efficiency in reducing the sensitivity to desiccation and increasing the storage period of Magnolia ovata seeds. The drying of seeds to 10% water did not significantly affect germination, nor was germination impaired in seeds that were dried and treated with ABA. However, seeds treated with PEG had lower viability. After storage, ABA-treated seeds with as low as 5% water content (WC) had higher germination rates, the treatment with 100 µM ABA providing the greatest storage capacity. No changes were observed in the activity of catalase, peroxidase, or superoxide dismutase or in the abundance of total or heat-resistance proteins.
Index terms:
enzymatic activity; forest seeds; intermediate seeds; proteins
Resumo:
Vários estudos têm utilizado com sucesso o polietilenoglicol (PEG) e/ou ácido abscísico (ABA) para induzir a tolerância à dessecação em sementes germinadas de espécies ortodoxas, melhorando nosso entendimento dos mecanismos que fundamentam essa característica. Nesse contexto, tratamentos com PEG e ABA foram testados para avaliar sua eficiência em reduzir a sensibilidade à dessecação e aumentar o período de armazenamento das sementes de Magnolia ovata. A secagem das sementes a 10% de teor de água não afetou significativamente a germinação, independentemente de terem sido previamente tratadas ou não com ABA. No entanto, sementes tratadas com PEG apresentaram menor viabilidade. Após o armazenamento, as sementes tratadas com ABA e dessecadas a 5% de teor de água, apresentaram maiores taxas de germinação, sendo que o tratamento com ABA 100 µM proporcionou a maior capacidade de armazenamento. Não foram observadas alterações na atividade da catalase, peroxidase ou superóxido dismutase ou na abundância de proteínas totais ou resistentes ao calor.
Termos para indexação:
atividade enzimática; sementes florestais; sementes intermediárias; proteínas
INTRODUCTION
Desiccation tolerance is the ability to withstand water loss and survive after rehydration without permanent damage (Dekkers et al., 2015DEKKERS, B.J.; COSTA, M.C.; MAIA, J.; BENTSINK, L.; LIGTERINK, W.; HILHORST, H.W. Acquisition and loss of desiccation tolerance in seeds: from experimental model to biological relevance. Planta, v.241, n.3, p.563-577, 2015. http://dx.doi.org/10.1007/s00425-014-2240-x.
http://dx.doi.org/10.1007/s00425-014-224...
). Seeds with these characteristics are called orthodox. The preservation of these seeds, whether for short or long periods, is directly influenced by their desiccation tolerance (Gonçalves et al., 2017GONÇALVES, M.I.F.; FARIA, J.M.R.; JOSÉ, A.C.; TONETTI, O.A.O.; MARQUES, E.R. Desiccation tolerance and antioxidant enzymatic activity in Citrus reschni seeds exposed to various drying rates. Seed Science and Technology, v.45, n.2, p.411-427, 2017. https://doi.org/10.15258/sst.2017.45.2.08.
https://doi.org/10.15258/sst.2017.45.2.0...
). The difficulty in preserving the physiological quality of unorthodox seeds has prevented the preservation of certain species. Recalcitrant seeds are dispersed with high water content and high metabolic activity, and their viability is lost as the water content decreases, so they cannot be stored at low temperatures with low water content (Pereira et al., 2020PEREIRA, L.C.V.; MAYRINCK, R. C.; ZAMBON, C. R.; JOSE, A.C.; FARIA, J.M.R. Storage of short-lived seeds of Inga vera subsp. affinis in osmotic medium. Seed Science Research, v.30, p.1-5, 2020. http://dx.doi.org/10.1017/S0960258520000185
http://dx.doi.org/10.1017/S0960258520000...
).
Seed viability during drying and storage is directly related to their ability to eliminate reactive oxygen species (ROS) produced during these processes. If they are not eliminated, ROS can damage macromolecules or act as signals that induce programmed cell death. Desiccation tolerant seeds, during the drying, increases cytoplasmic viscosity, forming the glassy state, decreasing all chemical reactions, thus helping to maintain viability during drying and storage. However, desiccation sensitive seeds cannot maintain the integrity of their membranes and organelles under low water content (Oliver et al., 2020OLIVER, M.J.; FARRANT, J.M.; HILHORST, H.W.M.; MUNDREE, S.; WILLIAMS, B.; BEWLEY, J.D. Desiccation tolerance: avoiding cellular damage during drying and rehydration. Annual Review of Plant Biology, v.71, n.1, p.435-460, 2020. https://www.annualreviews.org/doi/full/10.1146/annurev-arplant-071219-105542.
https://www.annualreviews.org/doi/full/1...
).
The reinduction of desiccation tolerance in germinated orthodox seeds has been performed in an attempt to understand the mechanisms that make the drying and storage of seeds sensitive to desiccation unviable. For this purpose, seed conditioning with polyethylene glycol (PEG) and abscisic acid (ABA) has provided good results (Buitink et al., 2003BUITINK, J.; THOMAS, M.; GISSOT, L.; LEPRINCE, O. Starvation, osmotic stress and desiccation tolerance lead to expression of different genes of the regulatory b and g subunits of the SnRK1 complex in germinating seeds of Medicago truncatula. Plant Cell and Environment, v.27, n.1, p.55-67, 2003. https://doi.org/10.1046/j.0016-8025.2003.01125.x
https://doi.org/10.1046/j.0016-8025.2003...
; Vieira et al., 2010VIEIRA, C.V.; SILVA, E.A.A.; ALVARENGA, A.A.; CASTRO, E.M.; TOOROP, P.E. Stress-associated factors increase after desiccation of germinated seeds of Tabebuia impetiginosa Mart. Plant Growth Regulation, v.62, n.3, p.257-263, 2010. https://doi.org/10.1007/s10725-010-9496-3.
https://doi.org/10.1007/s10725-010-9496-...
; Maia et al., 2011MAIA, J.; DEKKERS, B.J.; PROVART, N.J.; LIGTERINK, W.; HILHORST, H.W. The re-establishment of desiccation tolerance in germinated Arabidopsis thaliana seeds and its associated transcriptome. Plos One, v.6, n. 12, 2011. http://dx.doi.org/10.1371/journal.pone.0029123.
http://dx.doi.org/10.1371/journal.pone.0...
; Adu-Gyamfi and Wetten, 2012ADU-GYAMFI, R.; WETTEN, A. Cryopreservation of cocoa (Theobroma cacao L.) somatic embryos by vitrification. Cryo-Letters, v.33, n.6, p.494-505, 2012. https://pubmed.ncbi.nlm.nih.gov/23250409/
https://pubmed.ncbi.nlm.nih.gov/23250409...
; Dekkers et al., 2015DEKKERS, B.J.; COSTA, M.C.; MAIA, J.; BENTSINK, L.; LIGTERINK, W.; HILHORST, H.W. Acquisition and loss of desiccation tolerance in seeds: from experimental model to biological relevance. Planta, v.241, n.3, p.563-577, 2015. http://dx.doi.org/10.1007/s00425-014-2240-x.
http://dx.doi.org/10.1007/s00425-014-224...
). The osmotic potential of PEG, exposure time and concentration of ABA may influence the response to reinduction of desiccation tolerance (Maia et al., 2011MAIA, J.; DEKKERS, B.J.; PROVART, N.J.; LIGTERINK, W.; HILHORST, H.W. The re-establishment of desiccation tolerance in germinated Arabidopsis thaliana seeds and its associated transcriptome. Plos One, v.6, n. 12, 2011. http://dx.doi.org/10.1371/journal.pone.0029123.
http://dx.doi.org/10.1371/journal.pone.0...
), which varies between species. Some proteins affected by these treatments are associated to antioxidant activities and responses to stress, inducing the synthesis of protective molecules, such as sugars and late embryogenic abundant (LEA) proteins and heat shock proteins (HSPs) (Buitink et al., 2003BUITINK, J.; THOMAS, M.; GISSOT, L.; LEPRINCE, O. Starvation, osmotic stress and desiccation tolerance lead to expression of different genes of the regulatory b and g subunits of the SnRK1 complex in germinating seeds of Medicago truncatula. Plant Cell and Environment, v.27, n.1, p.55-67, 2003. https://doi.org/10.1046/j.0016-8025.2003.01125.x
https://doi.org/10.1046/j.0016-8025.2003...
; Maia et al., 2011MAIA, J.; DEKKERS, B.J.; PROVART, N.J.; LIGTERINK, W.; HILHORST, H.W. The re-establishment of desiccation tolerance in germinated Arabidopsis thaliana seeds and its associated transcriptome. Plos One, v.6, n. 12, 2011. http://dx.doi.org/10.1371/journal.pone.0029123.
http://dx.doi.org/10.1371/journal.pone.0...
). It is believed that the use of PEG, combined or not with ABA, can reduce the sensitivity to desiccation of seeds sensitive to desiccation and storage.
Magnolia ovata, belonging to the family Magnoliaceae, has great ecological importance specially in restoration of riparian forests due to its adaptation to swampy soils. Its seeds are classified as intermediate in terms of their tolerance to desiccation and storage (José et al., 2011JOSÉ, A.C.; SILVA, E.A.A.; DAVIDE, A.C.; MELO, A.J.S.; TOOROP, P.E. Effects of drying rate and storage time on Magnolia ovata Spreng. seed viability. Seed Science and Technology , v.39, n.2, p.425-434, 2011. http://dx.doi.org/10.15258/sst.2011.39.2.14.
http://dx.doi.org/10.15258/sst.2011.39.2...
). This study aimed to evaluate the effect of seed conditioning with PEG and ABA on the reduction of desiccation sensitivity and to prolong the longevity of stored M. ovata seeds.
MATERIALS AND METHODS
Fruit collection and seed processing
The collection was performed in Lavras, Minas Gerais, Brazil, from seven trees. The fruits were taken to the Forest Nursery of the Universidade Federal de Lavras. After the fruits naturally opened, the seeds were processed using a sieve in running water to remove the red aril that covers the seed. The seeds were taken to the Forest Seeds Laboratory in the Department of Forest Sciences and kept in a single layer on a bench to dry off the surface water with a paper towel. The seeds were stored in a cold chamber (5 °C) in a plastic bag for 5 days.
Determination of water content
The water content was determined by the oven-drying method at 105 °C for 24 hours (Brasil, 2013BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Instruções para análise de sementes de espécies florestais. Secretaria de Defesa Agropecuária. Brasília: MAPA/ACS, 2013. 97p. https://www.gov.br/agricultura/pt-br/assuntos/laboratorios/arquivos-publicacoes-laboratorio/florestal_documento_pdf-ilovepdf-compressed.pdf
https://www.gov.br/agricultura/pt-br/ass...
), with four replications of 10 seeds each. The calculation was performed on a wet basis, and the results are expressed as percentages.
Osmotic conditioning and ABA treatment
Osmotic conditioning and ABA treatment of the seeds were performed in PEG 6000 and ABA solutions under the following conditions: 5 µM ABA; 50 µM ABA; 100 µM ABA; -2.1 MPa PEG; 5 µM ABA + -2.1 MPa PEG; 50 µM ABA + -2.1 MPa PEG; or 100 µM ABA + -2.1 MPa PEG.
For conditioning, the seeds were kept in plastic trays containing PEG and ABA solutions (200 mL of solution for each kilo of seeds). The amount of solution used did not fully cover the seed layer, to allow oxygenation. The trays were covered with aluminum foil, following the recommendation of Costa et al. (2015COSTA, M.C.; RIGHETTI, K.; NIJVEEN, H.; YAZDANPANAH, F.; LIGTERINK, W.; BUITINK, J.; HILHORST, H.W. A gene co-expression network predicts functional genes controlling the re-establishment of desiccation tolerance in germinated Arabidopsis thaliana seeds. Planta, n.242, v.2, p.435-449, 2015. http://dx.doi.org/10.1007/s00425-015-2283-7.
http://dx.doi.org/10.1007/s00425-015-228...
) and placed in a germination chamber at 10 °C for 72 hours. The solutions were replaced daily to keep the water potential constant during the conditioning period. After conditioning, the seeds were washed in running water and kept for 1 hour at 20 °C to remove surface water. Next, the water content was determined, and the seeds were dried.
Seed drying
After conditioning, the seeds were dried following the recommendation of Magistrali et al. (2013MAGISTRALI, P.R.; JOSÉ, A.C.; FARIA, J.M.R.; GASPARIN, E. Physiological behavior of Genipa americana L. seeds regarding the capacity for desiccation and storage tolerance. Journal of Seed Science , v.35, n.4, p.495-500, 2013. http://dx.doi.org/10.1590/S2317-15372013000400011.
http://dx.doi.org/10.1590/S2317-15372013...
). The seeds were placed in single layer inside hygrostat boxes (box with a lid and internal air circulation generated by a small fan, allowing the homogenization of the relative humidity) containing one of saline solutions (Table 1). Seeds were kept in each condition until reach the target water content. When the water content was not obtained, seeds were transferred to a hygrostat with lower relative humidity, and so on.
Saline solutions used for drying M. ovata seeds to obtain different equilibrium relative humidities.
During drying, seeds were weighed daily until the water content reached 15%, 10%, or 5%. Upon reaching the target weight, the water content was measured by the oven-drying method.
Seed storage
After reaching 10% and 5% water content (WC), seeds were stored at -21 °C in a hermetically sealed container. After 45 days, germination tests were performed to determine seed viability.
Evaluation of seed quality
Before starting the germination tests, seeds were pre humidified for 24 hours at 25 °C in a single layer in a Gerbox® containing 50 mL of water at the bottom. Soon after, seeds were superficially disinfected with 2% sodium hypochlorite for 3 minutes, and the germination tests were performed on Petri dishes containing two sheets of filter paper moistened with water, which were kept at 25 °C with a photoperiod of 12 hours. Five replications of 20 seeds each were used.
The evaluations were performed at regular intervals of 3 days using root protrusion (≥ 2 mm) as the germination test. The test was completed at 42 days, when the formation of normal seedlings was observed. Germination speed index (GSI) was evaluated daily by calculating the number of germinated seeds until stabilization using the formula proposed by Maguire (1962MAGUIRE, J.D. Speed of germination aid in selection and evaluation for seedling emergence and vigor. Crop Science, v.2, n.2, p.176-77, 1962. http://dx.doi.org/10.2135/cropsci1962.0011183X000200020033x.
http://dx.doi.org/10.2135/cropsci1962.00...
).
Electrophoretic pattern of total and heat-resistance proteins
Electrophoretic pattern of total and heat-resistance proteins were evaluated at the initial (20%); 15%, 10%, and 5% water contents, with and without PEG or PEG+ABA (100 µM ABA; -2.1 MPa PEG, -2.1 MPa PEG + 100 µM ABA), before and after storage for 45 days with 10% and 5% WC. Protein extraction was performed according to José et al. (2011JOSÉ, A.C.; SILVA, E.A.A.; DAVIDE, A.C.; MELO, A.J.S.; TOOROP, P.E. Effects of drying rate and storage time on Magnolia ovata Spreng. seed viability. Seed Science and Technology , v.39, n.2, p.425-434, 2011. http://dx.doi.org/10.15258/sst.2011.39.2.14.
http://dx.doi.org/10.15258/sst.2011.39.2...
).
Electrophoresis was performed in a discontinuous acrylamide gel (6% stacking gel and 12.5% acrylamide separating gel). A total of 40 µg of protein from each sample was loaded. The run was performed in a vertical apparatus with sodium dodecyl sulfate buffer at 200 V for approximately 6 hours. Gel fixation, staining and digitalization were performed according to José et al. (2011JOSÉ, A.C.; SILVA, E.A.A.; DAVIDE, A.C.; MELO, A.J.S.; TOOROP, P.E. Effects of drying rate and storage time on Magnolia ovata Spreng. seed viability. Seed Science and Technology , v.39, n.2, p.425-434, 2011. http://dx.doi.org/10.15258/sst.2011.39.2.14.
http://dx.doi.org/10.15258/sst.2011.39.2...
).
Enzyme activity
Extraction of catalase (CAT) and superoxide dismutase (SOD) was performed using 0.2 M Tris-HCl buffer pH 8.0 + 0.1% mercaptoethanol, at a ratio of 200 µL per 50 mg of macerated seeds. For the extraction of peroxidase (PO), phosphate extraction buffer (0.034 M dibasic sodium phosphate; 0.2 M sucrose; 2.56 g PVP; 0.003 M DTT; 5.7 mM ascorbic acid; 2.5 mM sodium borate; 1 g PEG 6000; 0.2% β-mercaptoethanol) was used. The tubes were vortexed, kept in the refrigerator for 12 hours and centrifuged for 30 minutes at 14,000 rpm at 4 °C.
For all enzymes, 40 µL of each sample was loaded into discontinuous polyacrylamide gel channels, using 7.5% polyacrylamide in the separating gel and 4.5% polyacrylamide in the stacking gel. The buffer for the gel/electrode system used was tris-glycine pH 8.9, and the run was performed at 120 V for 5 hours. After electrophoresis, it was used a specific method for developing the gels described by Alfenas et al. (2006ALFENAS, A.C. Eletroforese e marcadores bioquímicos em plantas e microorganismos. Viçosa: Editora UFV, 2006. 627p.), using the appropriate substrate for each enzyme. After development, the gels were scanned and analyzed by visual comparison using ImageJ® software to quantify the intensity of the observed bands.
Statistical analysis
The experiment was set up in a completely randomized design. In the first part of the experiment, which consisted of the analysis of the effects of conditioning and drying on seed viability, a 3×8 factorial design (water contents x conditioning treatments) was used, while the storage experiment was analyzed in a 2×8 factorial design (water contents x conditioning treatments). Data were analyzed using R for Windows software generalized linear model analysis, and the means were compared by the least significant difference (LSD) test at 5% probability.
RESULTS AND DISCUSSION
After processing, the seeds had 20% WC. In general, drying to 10% WC did not affect germination, except when the seeds had been conditioned in PEG, in which case a reduction in viability was observed.
Seed drying to 5% WC reduced the viability of untreated seeds (control) and of seeds treated with ABA. Viability was similar between these treatments, as was the GSI. However, viability was drastically reduced when the seeds were conditioned in PEG or PEG + ABA and dried to 5% WC (Figures 1A and 1B). In all treatments where osmotic conditioning with PEG was performed, the seeds had reduced viability, reduced formation of normal seedlings and GSI. In seeds with 10% WC, the use of 5 µM, 50 µM, or 100 µM ABA led to a higher GSI than in the control (Figures 1E and 1F). The use of PEG, combined or not with ABA, negatively influenced the formation of normal seedlings, the lowest values being observed when seeds were dried to 10% or 5% WC (Figures 1C and 1D). In seeds dried to 5% WC, the best result was obtained in those treated with 5 µM ABA. All treatments that included PEG at 10% and 5% WC showed no formation of normal seedlings.

Germination percentage, normal seedling percentage, and GSI of M. ovata seeds subjected to PEG conditioning and treatment with ABA. Panels A and B: Germination percentage; C and D: normal seedling percentage; E and F: germination speed index. In panels B, D, and F, the same letters, lowercase in a column and uppercase in a row, indicate the absence of significant differences by the LSD test at the 5% probability level between the treatments and between the water contents, respectively.
PEG has been successfully used to reestablish desiccation tolerance in seeds with orthodox behavior after germination, when they lose desiccation tolerance (Maia et al., 2011MAIA, J.; DEKKERS, B.J.; PROVART, N.J.; LIGTERINK, W.; HILHORST, H.W. The re-establishment of desiccation tolerance in germinated Arabidopsis thaliana seeds and its associated transcriptome. Plos One, v.6, n. 12, 2011. http://dx.doi.org/10.1371/journal.pone.0029123.
http://dx.doi.org/10.1371/journal.pone.0...
; Costa et al., 2015COSTA, M.C.; RIGHETTI, K.; NIJVEEN, H.; YAZDANPANAH, F.; LIGTERINK, W.; BUITINK, J.; HILHORST, H.W. A gene co-expression network predicts functional genes controlling the re-establishment of desiccation tolerance in germinated Arabidopsis thaliana seeds. Planta, n.242, v.2, p.435-449, 2015. http://dx.doi.org/10.1007/s00425-015-2283-7.
http://dx.doi.org/10.1007/s00425-015-228...
). However, for M. ovata seeds, the use of PEG, combined or not with ABA, impaired seed germination. PEG is considered an inert, nontoxic product because it is not absorbed by seeds during conditioning (Santos et al., 2011SANTOS, A.R.F.; SILVA-MANN, R.; FERREIRA, R.A. Restrição hídrica em sementes de jenipapo (Genipa americana L.). Revista Árvore, v.35, n.2, p. 213-220, 2011. http://dx.doi.org/10.1590/S0100-67622011000200006.
http://dx.doi.org/10.1590/S0100-67622011...
). The reason why using PEG combined with ABA reduced seed survival after drying is unknown. In the literature, it is common to observe a positive effect of this combination, however, in some cases, may cause damage that affects the viability of seeds, as observed by Ribeiro et al. (2016RIBEIRO, D.E.; ALVARENGA, A.A.; MARTINS, J.R.; RODRIGUES, A.C.; MAIA, V.O. Germinação e reindução da tolerância à dessecação em sementes de Senna multijuga (Rich.) Irwin et Barn. Ciência Florestal, v.26, n.4, p. 1133-1140, 2016. https://www.redalyc.org/pdf/534/53449110009.pdf
https://www.redalyc.org/pdf/534/53449110...
) in Senna multijuga seeds.
The best results for stored seeds were obtained when they were treated with 50 µM or 100 µM ABA and dried to 5% WC. Under these conditions, germination was higher than in dry seeds with up to 10% WC (Figures 2A and 2B). It should be noted, however, that normal seedlings were not produced in none of the treatments. ABA acts in biosynthesis and genes involved in the biosynthesis of many molecules such as non-reducing sugars and LEA proteins, contributing to the re-induction of DT (Silva et al., 2017SILVA, A.C.; DAVIDE, L.C.; BRAZ, G.T.; MAIA, J.; CASTRO, E.M.; SILVA, E.A.A. Re-induction of desiccation tolerance in germinated cowpea seeds. South African Journal of Botany, v.113, p.34-39, 2017. https://doi.org/10.1016/j.sajb.2017.07.011
https://doi.org/10.1016/j.sajb.2017.07.0...
). Neem seeds (Azadirachta indica), a species with similar behavior to M. ovata regarding the tolerance to desiccation and storage, when dried to 5% WC also improved germination after storage, compared to those stored with higher water contents (Sacandé et al., 2000SACANDÉ, M.; BUITINK, J.; HOEKSTRA, F.A. A study of water relations in neem (Azadirachta indica) seed that is characterized by complex storage behavior. Journal of Experimental Botany, v.51, n.344, p.635-643, 2000. https://doi.org/10.1093/jexbot/51.344.635
https://doi.org/10.1093/jexbot/51.344.63...
). According to the authors, this could be explained by the fact that below 9% WC there is formation of cytoplasmic glasses and reduction in the amount of freezable water.

Germination percentage and germination speed index (GSI) of M. ovata seeds subjected to different treatments and stored for 45 days at -21 °C. Panel A: Germination percentage; C: germination speed index. In panels B and D, the same letters, lowercase in a column and uppercase in a row, indicate the absence of significant differences by the LSD test at the 5% probability level between the treatments and between the water contents, respectively.
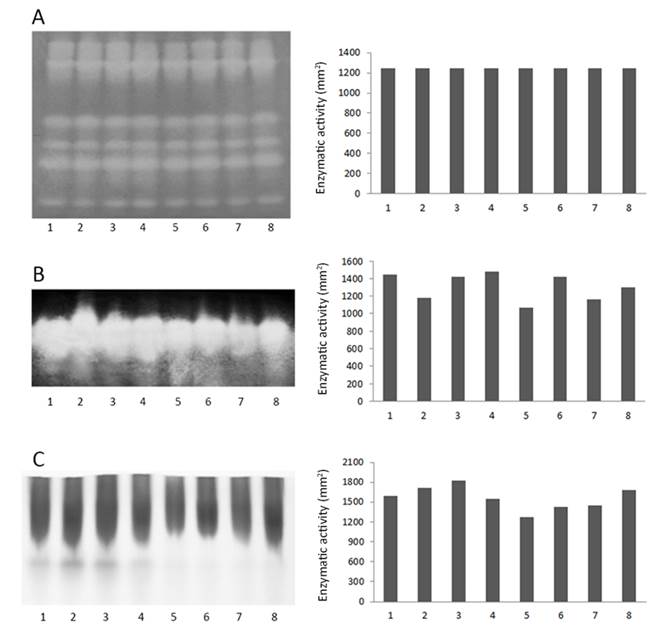
No changes in SOD activity were observed during drying and storage (Figures 3A and 4A). The activity of the CAT enzyme (Figure 3B) decreased during seed drying. However, CAT activity in seeds dried to 15% WC increased when they were treated with PEG and ABA, compared to the control. In general, PO activity was reduced with seed drying (Figure 3C), but a consistent trend was not observed.

Enzymatic activity in M. ovata seeds after PEG conditioning and ABA treatment. Left side: Electrophoresis gels after staining to visualize the enzymes. Right side: Numerical representation of band intensity. Panel A: Superoxide dismutase; B: catalase; C: peroxidase. Treatments: 1: 20% water; 2: 15% water; 3: Seeds treated with 100 µM ABA and dried to 15% water; 4: seeds treated with -2.1 MPa PEG and dried to 15% water; 5: seeds treated with -2.1 MPa PEG + 100 µM ABA and dried to 15% water; 6: 10% water; 7: seeds treated with 100 µM ABA and dried to 10% water; 8: seeds treated with -2.1 MPa PEG and dried to 10% water; 9: seeds treated with -2.1 MPa PEG + 100 µM ABA and dried to 10% water; 10: 5% water; 11: seeds treated with 100 µM ABA and dried to 5% water; 12: seeds treated with -2.1 MPa PEG and dried to 5% water; 13: seeds treated with -2.1 MPa PEG + 100 µM ABA and dried to 5% water.

Enzymatic activity in M. ovata seeds after PEG conditioning, ABA treatment, and drying, followed by storage for 45 days at -21 °C. Left side: Electrophoresis gels after staining for visualization of the enzymes. Right side: Numerical representation of band intensity. Panel A: Superoxide dismutase; B: catalase; C: peroxidase. Treatments: 1: 10% water; 2: seeds conditioned with 100 µM ABA and dried to 10% water; 3: seeds conditioned with -2.1 MPa PEG and dried to 10% water; 4: seeds conditioned in -2.1 MPa PEG + 100 µM ABA and dried to 10% water; 5: seeds dried to 5% water; 6: seeds treated with 100 µM ABA and dried to 5% water; 7: seeds conditioned in -2.1 MPa PEG, dried to 5% water; 8: seeds conditioned in -2.1 MPa PEG + 100 µM ABA, dried to 5% water.
With drying, M. ovata seeds progressively lost viability. In seeds sensitive to desiccation, the dysregulation of metabolism is directly related to the generation of ROS. As water is lost, the metabolism becomes unbalanced, causing the accumulation of ROS (Varghese et al., 2011VARGHESE, B.; BERJAK, P.; VARGHESE, D.; PAMMENTER, N.W. Differential drying rates of recalcitrant Trichilia dregeana embryonic axes: a study of survival and oxidative stress metabolism. Physiologia Plantarum, v.142, n.4, p.326-338, 2011. http://dx.doi.org/10.1111/j.1399-3054.2011.01469.x.
http://dx.doi.org/10.1111/j.1399-3054.20...
), and because the antioxidant system is inefficient at removing ROS, they cause irreversible damage to lipids, proteins, and fatty acids (Kurek et al., 2019KUREK, K.; PLITTA-MICHALAK, B.; RATAJCZAK, E. Reactive oxygen species as potential drivers of the seed aging process. Plants, v.8, n.174, 2019. https://doi.org/10.3390/plants8060174
https://doi.org/10.3390/plants8060174...
).
Different parts of the seed have varying levels of desiccation tolerance (Maia et al., 2011MAIA, J.; DEKKERS, B.J.; PROVART, N.J.; LIGTERINK, W.; HILHORST, H.W. The re-establishment of desiccation tolerance in germinated Arabidopsis thaliana seeds and its associated transcriptome. Plos One, v.6, n. 12, 2011. http://dx.doi.org/10.1371/journal.pone.0029123.
http://dx.doi.org/10.1371/journal.pone.0...
). The lack of formation of normal seedlings may be related to this factor because during the tests, root protrusion was observed in some treatments after drying at low water contents, though without development of the shoots. The mechanisms of desiccation tolerance include the action of heat-resistance proteins, among which are LEA proteins (Manfre et al., 2009MANFRE, A.J.; LAHATTE, G.A.; CLIMER, C.R.; MARCOTTE JR., W.R. Seed dehydration and the establishment of desiccation tolerance during seed maturation is altered in the Arabidopsis thaliana mutant atem6-1. Plant and Cell Physiology, v.50, n.2, p.243-253, 2009. http://dx.doi.org/10.1093/pcp/pcn185.
http://dx.doi.org/10.1093/pcp/pcn185...
).
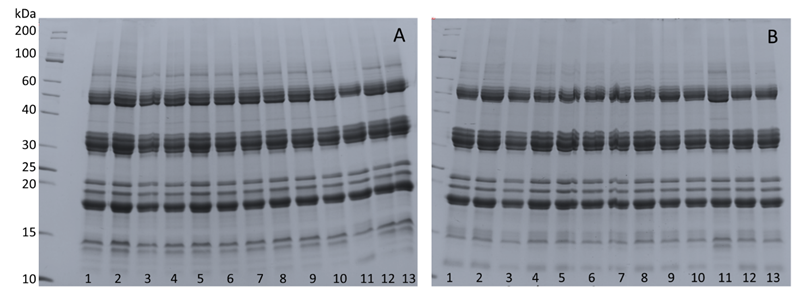
No changes were observed in the electrophoretic pattern of total or heat-resistance proteins. Most likely, the treatments used were not efficient in activating protection through these proteins, or the analytical method was not sensitive enough to detect these changes, as also observed by Gonçalves et al. (2015GONÇALVES, L.H.N.; SANTOS, H.O.; VON PINHO, E.V.R.; ANDRADE, T.; PINHO, I.V.V.; PEREIRA, R.W. Physiological quality and expression of genes in seeds of Handroanthus serratifolius subjected to drying. Journal of Seed Science, v.37, n.2, p. 102-110, 2015. http://dx.doi.org/10.1590/2317-1545v37n2144303.
http://dx.doi.org/10.1590/2317-1545v37n2...
) in studies with Handroanthus serratifolius seeds.
There was a trend toward decreased PO enzyme activity during drying and storage (Figure 4C), but without a consistent pattern. The same was observed in Citrus reshni seeds, in which the activity of CAT and SOD enzymes increased at the beginning of drying, but as the seed water content dropped, a drastic reduction in the activity of these enzymes was observed (Gonçalves et al., 2017GONÇALVES, M.I.F.; FARIA, J.M.R.; JOSÉ, A.C.; TONETTI, O.A.O.; MARQUES, E.R. Desiccation tolerance and antioxidant enzymatic activity in Citrus reschni seeds exposed to various drying rates. Seed Science and Technology, v.45, n.2, p.411-427, 2017. https://doi.org/10.15258/sst.2017.45.2.08.
https://doi.org/10.15258/sst.2017.45.2.0...
). Studying the storage of Oryza sativa seeds, Gao et al. (2016GAO, J.; FU, H.; ZHOU, X.; CHEN, Z.; LUO, Y.; CUI, B.; CHEN, G.; LIU, J. Comparative proteomic analysis of seed embryo proteins associated with seed storability in rice (Oryza sativa L.) during natural aging. Plant Physiology and Biochemistry, v.103, n.3, p.31-44, 2016. https://doi.org/10.1016/j.plaphy.2016.02.026.
https://doi.org/10.1016/j.plaphy.2016.02...
) did not find a correlation between the activities of the antioxidant enzymes SOD, CAT, and PO. In Ginkgo biloba seeds (Tommasi et al., 2006TOMMASI, F.; PACIOLLA, C.; PINTO, M.C.; GARA, L. Effects of storage temperature on viability, germination and antioxidant metabolism in Ginkgo biloba L. seeds. Plant Physiology and Biochemistry , n.44, p.359-368, 2006. https://doi.org/10.1016/j.plaphy.2006.06.014.
https://doi.org/10.1016/j.plaphy.2006.06...
), the responses of PO and CAT enzymes were similar to those of M. ovata seeds: CAT activity was less affected than PO, with little change during storage. It is believed that the inability of seeds to increase the activities of these enzymes during desiccation and storage may be one of the causes of the damage that occurs during storage (Tommasi et al., 2006). In the present study, CAT seemed to be less affected by storage (Figure 4B). For both enzymes, seeds with 5% WC had slightly lower activity than seeds with 10% WC. This is related to the finding that seeds with 5% WC showed greater viability (Figure 2).
Data on ROS production and antioxidant activity in recalcitrant seed tissues in response to desiccation are inconsistent between species. The same can occur with intermediate seeds, as we did not observe a consistent pattern of activity of the studied enzymes. This may be due to the great variability between the species of these two groups, such as the water content at the time of dispersal, the rate of water loss, and the response to drying (Berjak and Pammenter, 2013BERJAK, P.; PAMMENTER, N. Implications of the lack of desiccation tolerance in recalcitrance seeds. Frontiers in Plant Science, v.4, p.1-9, 2013. http://dx.doi.org/10.3389/fpls.2013.00478.
http://dx.doi.org/10.3389/fpls.2013.0047...
).
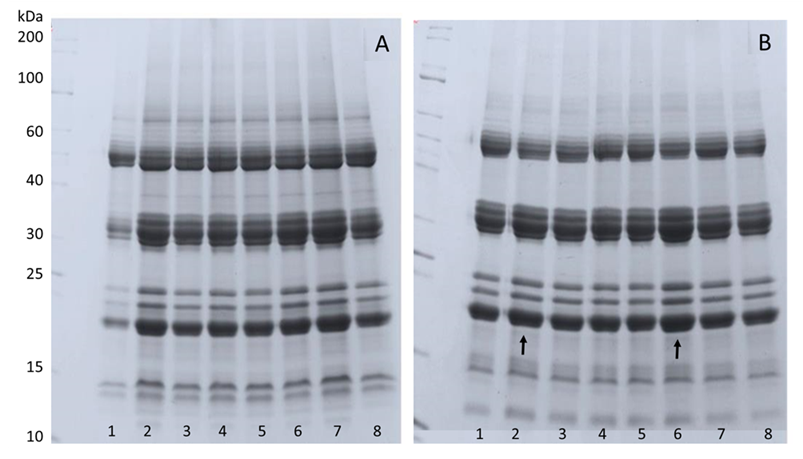
The electrophoretic profile of total proteins (Figure 5A) and heat-resistance proteins (Figure 5B) showed no differences between treatments. The same electrophoretic pattern was seen for total (Figure 6A) and heat-resistant proteins (Figure 6B) in seeds that were stored for 45 days at -21 °C. Stored seeds, treated with 100 µM ABA at 10% and 5% WC showed the highest band intensities of the heat-resistance proteins (Figure 6B). This higher protein abundance observed in these treatments was apparently associated positively with desiccation tolerance (Figure 2A).

Electrophoretic pattern of total and heat-resistance proteins of M. ovata after different treatments and with different water contents. Panel A: Total proteins; B: heat-resistance proteins. Treatments: 1: 20% water content (WC); 2: 15% WC; 3: 100 µM ABA and dried to 15% WC; 4: -2.1 MPa PEG and dried to 15% WC; 5: -2.1 MPa PEG + 100 µM ABA and dried to 15% WC; 6: 10% WC; 7: 100 μM ABA and dried to 10% WC; 8: -2.1 MPa PEG and dried to 10% WC; 9: -2.1 MPa PEG + 100 µM ABA and dried to 10% WC; 10: 5% WC; 11: 100 µM ABA and dried to 5% WC; 12: -2.1 MPa PEG and dried to 5% WC; 13: -2.1 MPa PEG + 100 µM ABA and dried to 5% WC.

Electrophoretic patterns of total and heat-resistant proteins of M. ovata after different treatments at different water contents stored for 45 days at -21 °C. Panel A: Total proteins; B: heat-resistance proteins. Treatments: 1: 10% water content (WC); 2: 100 µM ABA and dried to 10% WC; 3: -2.1 MPa PEG and dried to 10% WC; 4: -2.1 MPa PEG + 100 µM ABA and dried to 10% WC; 5: 5% WC; 6: 100 µM ABA and dried to 5% WC; 7: -2.1 MPa PEG and dried to 5% WC; 8: -2.1 MPa PEG + 100 µM ABA and dried to 5% WC. Arrows indicate the treatments with the highest band intensity.
Even without the formation of normal seedlings after storage, the treatment of seeds with 50 and 100 µM ABA led to a higher germination percentage than the other treatments. The application of ABA may have activated other mechanisms that allow greater tolerance of M. ovata seeds, such as the presence of dehydrins, which are associated with changes in endogenous ABA levels (Smolikova et al., 2020SMOLIKOVA, G.; LEONOVA, T.; VASHURINA, N.; FROLOV, A.; MEDVEDEV, S. Desiccation tolerance as the basis of long-term seed viability. International Journal of Molecular Sciences, v.22, n.1, 2020. https://doi.org/10.3390/ijms22010101
https://doi.org/10.3390/ijms22010101...
). In studies with Arabidopsis thaliana seeds, high ABA concentrations stimulated the formation or release of protective sugars (Finkelstein, 2013FINKELSTEIN, R. Abscisic acid synthesis and response. Arabidopsis Book, v.11, e0166, 2013. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3833200/
https://www.ncbi.nlm.nih.gov/pmc/article...
). These sugars assist in the structural and functional preservation of proteins and membranes during the drying and storage processes (Hoekstra et al., 2001HOEKSTRA, F.A.; GOLOVINA, E.A.; BUITINK J. Mechanisms of plant desiccation tolerance. Trends in Plant Science, v.6, n.9, p.431-438, 2001. https://doi.org/10.1016/S1360-1385(01)02052-0.
https://doi.org/10.1016/S1360-1385(01)02...
), contributing to the maintenance of seed viability.
CONCLUSIONS
Drying seeds to 10% WC does not affect the viability of M. ovata seeds.
The use of PEG combined or not with ABA, negatively affected seed viability.
The treatment of seeds dried to 5% WC with ABA from 5 to 100 µM can increase their storage capacity at low temperatures (-21 °C).
The activity of SOD, CAT, and PO and electrophoretic profile of total proteins were not affected by drying or storage of M. ovata seeds, however, the electrophoretic profile of heat-resistance proteins showed changes in band intensities and should be investigated in further studies.
ACKNOWLEDGMENTS
A.C.J. and J.M.R.F. thanks the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the research productivity fellowships (Processes 310976/2018-9 and 311556/2018-3, respectively). The other authors thanks the following agencies for the scholarships: CNPq; Fundação de Amparo à Pesquisa do Estado de Minas Gerais - FAPEMIG; and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - CAPES.
REFERENCES
- ADU-GYAMFI, R.; WETTEN, A. Cryopreservation of cocoa (Theobroma cacao L.) somatic embryos by vitrification. Cryo-Letters, v.33, n.6, p.494-505, 2012. https://pubmed.ncbi.nlm.nih.gov/23250409/
» https://pubmed.ncbi.nlm.nih.gov/23250409/ - ALFENAS, A.C. Eletroforese e marcadores bioquímicos em plantas e microorganismos Viçosa: Editora UFV, 2006. 627p.
- BERJAK, P.; PAMMENTER, N. Implications of the lack of desiccation tolerance in recalcitrance seeds. Frontiers in Plant Science, v.4, p.1-9, 2013. http://dx.doi.org/10.3389/fpls.2013.00478
» http://dx.doi.org/10.3389/fpls.2013.00478 - BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Instruções para análise de sementes de espécies florestais Secretaria de Defesa Agropecuária. Brasília: MAPA/ACS, 2013. 97p. https://www.gov.br/agricultura/pt-br/assuntos/laboratorios/arquivos-publicacoes-laboratorio/florestal_documento_pdf-ilovepdf-compressed.pdf
» https://www.gov.br/agricultura/pt-br/assuntos/laboratorios/arquivos-publicacoes-laboratorio/florestal_documento_pdf-ilovepdf-compressed.pdf - BUITINK, J.; THOMAS, M.; GISSOT, L.; LEPRINCE, O. Starvation, osmotic stress and desiccation tolerance lead to expression of different genes of the regulatory b and g subunits of the SnRK1 complex in germinating seeds of Medicago truncatula Plant Cell and Environment, v.27, n.1, p.55-67, 2003. https://doi.org/10.1046/j.0016-8025.2003.01125.x
» https://doi.org/10.1046/j.0016-8025.2003.01125.x - COSTA, M.C.; RIGHETTI, K.; NIJVEEN, H.; YAZDANPANAH, F.; LIGTERINK, W.; BUITINK, J.; HILHORST, H.W. A gene co-expression network predicts functional genes controlling the re-establishment of desiccation tolerance in germinated Arabidopsis thaliana seeds. Planta, n.242, v.2, p.435-449, 2015. http://dx.doi.org/10.1007/s00425-015-2283-7
» http://dx.doi.org/10.1007/s00425-015-2283-7 - DEKKERS, B.J.; COSTA, M.C.; MAIA, J.; BENTSINK, L.; LIGTERINK, W.; HILHORST, H.W. Acquisition and loss of desiccation tolerance in seeds: from experimental model to biological relevance. Planta, v.241, n.3, p.563-577, 2015. http://dx.doi.org/10.1007/s00425-014-2240-x
» http://dx.doi.org/10.1007/s00425-014-2240-x - FINKELSTEIN, R. Abscisic acid synthesis and response. Arabidopsis Book, v.11, e0166, 2013. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3833200/
» https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3833200/ - GAO, J.; FU, H.; ZHOU, X.; CHEN, Z.; LUO, Y.; CUI, B.; CHEN, G.; LIU, J. Comparative proteomic analysis of seed embryo proteins associated with seed storability in rice (Oryza sativa L.) during natural aging. Plant Physiology and Biochemistry, v.103, n.3, p.31-44, 2016. https://doi.org/10.1016/j.plaphy.2016.02.026
» https://doi.org/10.1016/j.plaphy.2016.02.026 - GONÇALVES, L.H.N.; SANTOS, H.O.; VON PINHO, E.V.R.; ANDRADE, T.; PINHO, I.V.V.; PEREIRA, R.W. Physiological quality and expression of genes in seeds of Handroanthus serratifolius subjected to drying. Journal of Seed Science, v.37, n.2, p. 102-110, 2015. http://dx.doi.org/10.1590/2317-1545v37n2144303
» http://dx.doi.org/10.1590/2317-1545v37n2144303 - GONÇALVES, M.I.F.; FARIA, J.M.R.; JOSÉ, A.C.; TONETTI, O.A.O.; MARQUES, E.R. Desiccation tolerance and antioxidant enzymatic activity in Citrus reschni seeds exposed to various drying rates. Seed Science and Technology, v.45, n.2, p.411-427, 2017. https://doi.org/10.15258/sst.2017.45.2.08
» https://doi.org/10.15258/sst.2017.45.2.08 - HOEKSTRA, F.A.; GOLOVINA, E.A.; BUITINK J. Mechanisms of plant desiccation tolerance. Trends in Plant Science, v.6, n.9, p.431-438, 2001. https://doi.org/10.1016/S1360-1385(01)02052-0
» https://doi.org/10.1016/S1360-1385(01)02052-0 - JOSÉ, A.C.; SILVA, E.A.A.; DAVIDE, A.C.; MELO, A.J.S.; TOOROP, P.E. Effects of drying rate and storage time on Magnolia ovata Spreng. seed viability. Seed Science and Technology , v.39, n.2, p.425-434, 2011. http://dx.doi.org/10.15258/sst.2011.39.2.14
» http://dx.doi.org/10.15258/sst.2011.39.2.14 - KUREK, K.; PLITTA-MICHALAK, B.; RATAJCZAK, E. Reactive oxygen species as potential drivers of the seed aging process. Plants, v.8, n.174, 2019. https://doi.org/10.3390/plants8060174
» https://doi.org/10.3390/plants8060174 - MAGISTRALI, P.R.; JOSÉ, A.C.; FARIA, J.M.R.; GASPARIN, E. Physiological behavior of Genipa americana L. seeds regarding the capacity for desiccation and storage tolerance. Journal of Seed Science , v.35, n.4, p.495-500, 2013. http://dx.doi.org/10.1590/S2317-15372013000400011
» http://dx.doi.org/10.1590/S2317-15372013000400011 - MAIA, J.; DEKKERS, B.J.; PROVART, N.J.; LIGTERINK, W.; HILHORST, H.W. The re-establishment of desiccation tolerance in germinated Arabidopsis thaliana seeds and its associated transcriptome. Plos One, v.6, n. 12, 2011. http://dx.doi.org/10.1371/journal.pone.0029123
» http://dx.doi.org/10.1371/journal.pone.0029123 - MAGUIRE, J.D. Speed of germination aid in selection and evaluation for seedling emergence and vigor. Crop Science, v.2, n.2, p.176-77, 1962. http://dx.doi.org/10.2135/cropsci1962.0011183X000200020033x
» http://dx.doi.org/10.2135/cropsci1962.0011183X000200020033x - MANFRE, A.J.; LAHATTE, G.A.; CLIMER, C.R.; MARCOTTE JR., W.R. Seed dehydration and the establishment of desiccation tolerance during seed maturation is altered in the Arabidopsis thaliana mutant atem6-1. Plant and Cell Physiology, v.50, n.2, p.243-253, 2009. http://dx.doi.org/10.1093/pcp/pcn185
» http://dx.doi.org/10.1093/pcp/pcn185 - OLIVER, M.J.; FARRANT, J.M.; HILHORST, H.W.M.; MUNDREE, S.; WILLIAMS, B.; BEWLEY, J.D. Desiccation tolerance: avoiding cellular damage during drying and rehydration. Annual Review of Plant Biology, v.71, n.1, p.435-460, 2020. https://www.annualreviews.org/doi/full/10.1146/annurev-arplant-071219-105542
» https://www.annualreviews.org/doi/full/10.1146/annurev-arplant-071219-105542 - PEREIRA, L.C.V.; MAYRINCK, R. C.; ZAMBON, C. R.; JOSE, A.C.; FARIA, J.M.R. Storage of short-lived seeds of Inga vera subsp. affinis in osmotic medium. Seed Science Research, v.30, p.1-5, 2020. http://dx.doi.org/10.1017/S0960258520000185
» http://dx.doi.org/10.1017/S0960258520000185 - RIBEIRO, D.E.; ALVARENGA, A.A.; MARTINS, J.R.; RODRIGUES, A.C.; MAIA, V.O. Germinação e reindução da tolerância à dessecação em sementes de Senna multijuga (Rich.) Irwin et Barn. Ciência Florestal, v.26, n.4, p. 1133-1140, 2016. https://www.redalyc.org/pdf/534/53449110009.pdf
» https://www.redalyc.org/pdf/534/53449110009.pdf - SACANDÉ, M.; BUITINK, J.; HOEKSTRA, F.A. A study of water relations in neem (Azadirachta indica) seed that is characterized by complex storage behavior. Journal of Experimental Botany, v.51, n.344, p.635-643, 2000. https://doi.org/10.1093/jexbot/51.344.635
» https://doi.org/10.1093/jexbot/51.344.635 - SANTOS, A.R.F.; SILVA-MANN, R.; FERREIRA, R.A. Restrição hídrica em sementes de jenipapo (Genipa americana L.). Revista Árvore, v.35, n.2, p. 213-220, 2011. http://dx.doi.org/10.1590/S0100-67622011000200006
» http://dx.doi.org/10.1590/S0100-67622011000200006 - SILVA, A.C.; DAVIDE, L.C.; BRAZ, G.T.; MAIA, J.; CASTRO, E.M.; SILVA, E.A.A. Re-induction of desiccation tolerance in germinated cowpea seeds. South African Journal of Botany, v.113, p.34-39, 2017. https://doi.org/10.1016/j.sajb.2017.07.011
» https://doi.org/10.1016/j.sajb.2017.07.011 - SMOLIKOVA, G.; LEONOVA, T.; VASHURINA, N.; FROLOV, A.; MEDVEDEV, S. Desiccation tolerance as the basis of long-term seed viability. International Journal of Molecular Sciences, v.22, n.1, 2020. https://doi.org/10.3390/ijms22010101
» https://doi.org/10.3390/ijms22010101 - TOMMASI, F.; PACIOLLA, C.; PINTO, M.C.; GARA, L. Effects of storage temperature on viability, germination and antioxidant metabolism in Ginkgo biloba L. seeds. Plant Physiology and Biochemistry , n.44, p.359-368, 2006. https://doi.org/10.1016/j.plaphy.2006.06.014
» https://doi.org/10.1016/j.plaphy.2006.06.014 - VARGHESE, B.; BERJAK, P.; VARGHESE, D.; PAMMENTER, N.W. Differential drying rates of recalcitrant Trichilia dregeana embryonic axes: a study of survival and oxidative stress metabolism. Physiologia Plantarum, v.142, n.4, p.326-338, 2011. http://dx.doi.org/10.1111/j.1399-3054.2011.01469.x
» http://dx.doi.org/10.1111/j.1399-3054.2011.01469.x - VIEIRA, C.V.; SILVA, E.A.A.; ALVARENGA, A.A.; CASTRO, E.M.; TOOROP, P.E. Stress-associated factors increase after desiccation of germinated seeds of Tabebuia impetiginosa Mart. Plant Growth Regulation, v.62, n.3, p.257-263, 2010. https://doi.org/10.1007/s10725-010-9496-3
» https://doi.org/10.1007/s10725-010-9496-3
Publication Dates
-
Publication in this collection
23 Feb 2022 -
Date of issue
2022
History
-
Received
06 Sept 2021 -
Accepted
21 Dec 2021