Abstract
Abstract: Despite its economic importance, the microclimate in soybean canopies has not yet been studied in detail. Such a study can yield valuable information regarding the interaction of a crop with its environment. In this context, the aim of this study was to evaluate the solar radiation dynamic and yield responses for each canopy strata for two soybean cultivars with determined and undetermined growth habits. A field study was conducted during the 2013/2014 and 2014/2015 growing seasons in the city of Frederico Westphalen, Rio Grande do Sul, Brazil. The cultivar NA6411, with a determinate growth habit, presented a greater interception of radiation in the middle and lower canopies strata which results in higher soybean yield when compared to the cultivar TEC6029, and thus, can be recommended for cultivation in crop production systems. The contribution of the middle and upper canopy strata to the total yield formation was greater than that observed in the lower canopy strata due to the greater interception of solar radiation by these strata. To increase soybean yields, new studies regarding the microclimatic conditions of the soybean canopy should be developed to improve the maximum potential yield of the new soybean cultivars.
Key words
canopy architecture; Glycine max L.; radiation use efficiency; extinction coefficient
INTRODUCTION
Soybean (Glycine max (L.) Merril) is one of the most important oilseeds cultivated in the world due to its high content of protein and oil (Yang et al. 2016YANG XF, YU XQ, ZHOU Z, MA WJ and TANG GX. 2016. A high-efficiency Agrobacterium tumefaciens mediated transformation system using cotyledonary node as explants in soybean (Glycine max L.). Acta Physiol Plant 38: 1-10.) and its widespread use for human and animal consumption. In the crop year 2016/2017, 114.07 million tons of soybeans were produced in Brazil, and the estimate for 2017/2018 is 108.6 million tons (Conab 2017CONAB - COMPANHIA NACIONAL DE ABASTECIMENTO ACOMPANHAMENTO DA SAFRA BRASILEIRA DE GRÃOS. 2016/2017. Disponível em: <http://www.conab.gov.br/OlalaCMS/uploads/arquivos/16_12_22_12_08_27_boletim_graos_dezembro_2016.pdf>. Acessado em: 15 Jan 2018.
http://www.conab.gov.br/OlalaCMS/uploads...
). The use of new production technologies and the improvement of cultural practices, aiming at a more efficient use of natural resources led to a significant increase of the cultivation area in Brazil and, therefore, to higher oilseed quantities harvested (Schwerz et al. 2016SCHWERZ F, CARON BO, ELLI EF, SOUZA VQ, OLIVEIRA DM and ROCKENBACH AP. 2016. Soybean morphological and productive characteristics influenced by meteorological parameters and sowing dates. Científica 44: 121-130.).
Microclimatic studies of the soybean canopies were conducted by Egli et al. (1970)EGLI DB, PENDLETON JW and PETERS DB. 1970. Photosynthetic rate of three soybean communities as related to CO2 levels and solar radiation. Agron J 62: 411- 414., Hiebsch et al. (1976)HIEBSCH CK, KANEMASU ET and NICKELL CD. 1976. Effects of soybean leaflet type on net carbon dioxide exchange, water use and water use efficiency. Can J Plant Sci 56: 455-458., BaldocchiBALDOCCHI DD, VERMA SB and ROSENBERG NJ. 1983. Microclimate in the soybean canopy. Agric Meteorol 28: 321-337. et al. (1983, 1985BALDOCCHI DD, VERMA SB, ROSENBERG NJ, BLAD BL and SPECHT JE. 1985. Microclimate-plant architectural interactions: influence of leaf width on the mass and energy exchange of a soybean canopy. Agr For Meteorol 35: 1-20.). These authors concluded that some disagreement prevails on the question of how alteration in a plant architecture may affect momentum, mass and energy exchanges between a canopy and the atmosphere. The authors suggested that many of the uncertainties reported in the literature might have been resolved had detailed observations been made of microclimatic and physiological conditions. This fact confirms the importance of the detailed study of the solar radiation dynamics and the productive response in different canopy strata of soybean cultivars with different growth habits.
An optimized utilization of the photosynthetically active radiation (PAR) intercepted by the canopy of soybean plants may maximize their photosynthetic efficiency, and thus lead to an increased grain yield (Marchão et al. 2006MARCHÃO RL, BRASIL EM and XIMENES PA. 2006. Interceptação da radiação fotossinteticamente ativa e rendimento de grãos do milho adensado. Rev Bras Milho Sorgo 5: 170-181.). The plant growth is a function of accumulated biomass through photosynthesis. Biomass production in plants depends upon the quantity of photosynthetically active radiation absorbed by leaves (PARa) and the efficiency with which the leaves can convert the radiation into assimilates through photosynthesis. Thus, the intercepted photosynthetically active radiation (PARi) that is converted into biomass reveals the radiation use efficiency (RUE) by species (Monteith 1977MONTEITH JL. 1977. Climate and the efficiency of crop production en Britain. Philos T Roy Soc B 181: 277-294., Van Heerden et al. 2010VAN ROEKEL RJ and PURCELL LC. 2014. Soybean biomass and nitrogen accumulation rates and radiation use efficiency in a maximum yield environment. Crop Sci 54: 1189-1196.).
Canopy architecture is determinant for the plant’s photosynthetic capacity (Wells 1991WELLS R. 1991. Soybean growth response to plant density: relationships among canopy photosynthesis, leaf area, and light interception. Crop Sci 31: 755-761.). The soybean crop has an upper strata of dense foliage, which makes it difficult for light to penetrate into lower leaf strata (Rambo et al. 2004RAMBO L, COSTA AJ, PIRES FLJ and FERREIRA GF. 2004. Estimativa do potencial de rendimento por estrato do dossel da soja, em diferentes arranjos de plantas. Cienc Rural 34: 33-40.). The PAR transmission in a plant canopy, according to Beer’s law (Fagan et al. 2013FAGAN EB, MIQUELANTI NP, BONFIM TS, PEREIRA IS, CORRÊA LT and SOARES JN. 2013. Lei de Beer e sua relação com a ecofisiologia de plantas. Cerrado Agrociência 4: 78-79.), is a negative exponential function of the product of leaf area index by the extinction coefficient. Additionally, the irregularity of light distribution throughout the canopy may result in unequal contributions of the different plant sections to the overall yield. Greater light incidence into the lower leaf strata as well as diffuse radiation, can increase the production of reproductive organs in the lower canopy strata, and lead to an overall increase in yield.
Studies highlighting the importance of soybean canopy stratification, quantifying the contribution of each strata and its leaf area index, extinction coefficient, and crop yield were not found in the literature. In coffee (Marin et al. 2003MARIN M, CASTRO B, GAITAN A, PREISIG O, WINGFIELD BD and WINGFIELD MJ. 2003. Relationships of Ceratocystis frimbiata isolates from Colombian coffee-growing regions based on molecular data a pathogenicity. J Phytopathol 151: 395-405., Cunha and Volpe 2010CUNHA AR and VOLPE CA. 2010. Relações radiométricas no terço superior da copa do cafeeiro. Bragantia 69: 263-271.), apple (BulerBULER Z and MIKA A. 2009. The influence of canopy architecture on light interception and distribution in ‘sampion’ apple trees. J Fruit Omam Plant Res 18: 96-100. and Mika 2009) and orange trees (Pilau and Angelocci 2015PILAU FG and ANGELOCCI. LR. 2015. Área foliar e interceptação de radiação solar pela copa de uma laranjeira. Bragantia 74: 476-482.), for example, the contribution of different leaf strata to yield formation were analyzed with respect to their interception of solar radiation. Studies indicate differentiated contributions of leaf strata to crop yield. Thus, the identification of alternatives for the best use of the microclimatic conditions is essential to maximize the contribution of the different leaf strata, especially in relation to the radiation use efficiency, which can result in a significant increase in soybean yield.
Soybean growth can be determined by the microclimatic conditions and the management that is utilized during the growing season. These characteristics can be influenced by cultivar growth habit (determined growth or undetermined growth) (Zanon et al. 2015ZANON AJ, STRECK NA, RICHTER GL, BECKER CC, ROCHA TSMD, CERA JC, WINCK JEM, CARDOZO AP, TAGLIAPIETRA EL and WEBER PS. 2015. Branches contribution and leaf area index evolution in modern cultivars of soybean. Bragantia 74: 279-290.). Plants with a determined growth habit finish the formation of leaves upon reaching the reproductive period, which results in differences in leaf architecture, leaf area index and radiation use efficiency in the different leaf strata, when compared to plants with an indeterminate growth habit, which even after reaching the reproductive period, maintain the formation of vegetative organs. Additionally, the leaf morphology of plants with determined growth remains constant throughout the plant extension. This fact differentiates determined plants of indeterminate because these present sheets of smaller sizes in the upper strata of the plant. The flowering of determined plants begins in the middle strata of the plant and progresses to the upper and lower strata. Consequently, the development of pods and seeds follows this pattern (SediyamaSEDIYAMA T, PEREIRA GP, SEDIYAMA CS and GOMES JLL. 1996. Cultura da Soja: I Parte. Viçosa: UFV, 96 p. et al. 1996, 1999SEDIYAMA T, TEIXEIRA RC and REIS MC. 1999. Melhoramento da Soja. In: Borém A. Melhoramento de espécies cultivadas. Viçosa: UFV, cap. 14, p. 487-531.).
Although photosynthesis is the basis for biomass production by plants, only few studies consider RUE as an alternative to increase soybean yield. The solar radiation intercepted by the different strata of the plant canopy is one of the most important components for the growth analysis. However, to estimate the amounts of intercepted solar radiation, it is necessary to determine the leaf area index (LAI) and the light extinction coefficient (k).
Information that reveals the dynamics of microclimatic conditions in the canopy strata and its relations with the solar radiation and soybean yield responses are relevant in order to improve some management practices for the soybean producers. The aim of this study was to evaluate the solar radiation dynamics and yield responses in different leaf strata of two soybean cultivars with determined and undetermined growth habit.
MATERIALS AND METHODS
STUDY AREA AND EXPERIMENTAL DESIGN
A field study was conducted during the 2013/2014 and 2014/2015 growing seasons in the city of Frederico Westphalen-Rio Grande do Sul, Brazil, at the coordinates 27°23′48′′S, 53°25′45′′W and an altitude of 490 m (Fig. 1). According to the Köppen climate classification, the climate is Cfa, i.e., humid subtropical with mean annual temperatures of 19.1 °C and varying maximum and minimum temperatures of 38 and 0 °C, respectively (AlvaresALVARES CA, STAPE JL, SENTELHAS PC, MORAES G, LEONARDO J and SPARROVEK G. 2013. Köppen’s climate classification map for Brazil. Meteorol Z 22: 711-728. et al. 2013).

Geographical location of the experiment. The State of Rio Grande do Sul is highlighted in black on the bottom map, while the city of Frederico Westphalen is highlighted in black on the main map.
According to the soil mapping unit of Passo Fundo, the soil is classified as Ferralsols typical. Fertilization was carried out in response to a soil analysis following the recommendations for soybean crops (CCSF 2004COMISSÃO DE QUÍMICA E FERTILIDADE DO SOLO. 2004. Manual de adubação e calagem para os estados do Rio Grande do Sul e Santa Catarina. Porto Alegre: Sociedade Brasileira de Ciência do Solo – Núcleo Regional Sul, p. 400.). For this, a total of 80 kg ha-2 of triple superphosphate and 50 kg ha-2 of potassium chloride were applied at the time of sowing. The control of weeds and diseases was also carried out through phytosanitary treatments following the recommendations of products for the crop (Embrapa 2006EMBRAPA. 2006. Tecnologias de Produção de Soja – Paraná 2007. Londrina. EMBRAPA, 220 p.).
The experimental design was a randomized complete block, where was tested two soybean cultivars and two growing seasons, with four repetitions. In both crop years, the cultivars Nideira A 6411 RG (Sementes Nidera, São Paulo, Brazil) that is characterized by a determined growth habit with an early cycle and average height, and CCGL TEC 6029 RR (Cooperativa Central Gaúcha Ltda, Rio Grande do Sul, Brazil) that exhibits an undetermined growth habit with early cycle and high height, were used in this study. The cultivars evaluated in this study were coded as follows: NA6411 (Nideira A 6411 RG) and TEC6029 (CCGL TEC 6029 RR).
Seed sowing was carried out on 11/14/2013 and 11/08/2014 with a sowing machine. Thinning was performed seven days after emergence (DAE) resulting in a final planting density of 250,000 plants per hectare. The experimental unit consisted of five rows of 3.0 m in length, with spacing between the seeding lines of 0.45 m. Only the three central rows, withdrawing 0.3 m of the field edge were sampled for analyses. The experimental procedures were the same in both crop years.
During the entire growth period of the crop, in both years, data of air temperature (°C), rainfall (mm), and solar radiation (MJ m-2) were obtained from the meteorological station of the Instituto Nacional de Meteorologia (INMET), situated about 100 m from the experimental site. The water balance was calculated on a monthly basis according to Thorntwaite and Mather (1995) using an Excel (Microsoft Office Professional Plus 2016, Microsoft Corporation, Redmond, WA, U.S.A.) calculation spreadsheet as proposed by Rolim et al. (1998)ROLIM GS, SENTELHAS PC and BARBIERI V. 1998. Planilhas no ambiente EXCEL para os cálculos de balanços hídricos: normal, sequencial, de cultura e de produtividade real e potencial. Rev Bras Agrometeorol 6: 133-137., considering the water storage capacity available on the ground 100 mm (Elli et al. 2016ELLI EF, CARON BOC, PAULA GM, ELOY E, SCHWERZ F and SCHIMDT D. 2016. Ecophysiology of Peltophorum dubium Spr. In: agroforestry systems arrangements. Comunicata Scientiae 7: 464-472.).
CANOPY STRATIFICATION
The determination of leaf area index, specific leaf area, yield components (number of pods per plant, thousand grain weight, total yield), interception of solar radiation and the extinction coefficient were performed for each soybean canopy strata (Fig. 2). Data assessment was carried out every two weeks from 30 to 105 days after emergence (30, 45, 60, 75, 90 and 105 DAE). Four plants per plot were destructively sampled at each sampling date.

Schematic of the separation of the plant canopy of soybean plants into upper, middle and lower leaf strata. Classifications are delineated by the two parallel lines.
Except for the first sampling date (30 DAE), the sampled plants were divided into three canopy sections of similar size in order to individually analyze the contribution of each canopy strata, according to the location of their vegetative and reproductive structures. The criterion used to sectioning the plants canopy was that to constitute the lower strata the plant structures should be located from 0 % to 33 % of the height of the plants, the middle stratum of 33.34 % to 66.66 % and the upper stratum of 66.67 % to 100 %, respectively.
At the first sampling date (30 DAE), the dry matter of the entire plant was calculated by adding the dry weights of the individual plant organs: branches, stems and leaves. At all other sampling dates, branches, stems, and leaves were calculated according to its position in the canopy layer: lower, middle and upper (Fig. 2).
BIOMASS AND LEAF AREA ANALYSIS
The total dry matter (TDM) of the plants was determined from the sum of the components: leaves, stem, branches, and pods. Each component was gathered and placed in pre-identified individual paper bag. Samples were dried in a forced circulation oven at 60 °C until a constant mass was obtained. The samples were later weighed on a precision balance in order to obtain the dry mass of each component, which together resulted in the TDM.
Leaf area in each different canopy strata was measured with a leaf area meter LI-COR Model 3000 (LI-COR, Lincoln, Nebraska, U.S.A), with the value expressed in cm2. The leaf area index (LAI) was determined by the ratio between plant leaf area and soil area (SA) that it occupied (0.04 m2) with the following equation:
Where LA is the leaf area, and SA is the soil area occupied by the plant.
SOLAR RADIATION ANALYSIS
The incident solar radiation (W m-2) in each evaluation period was measured using a portable pyranometer (LICOR PY32164, Lincoln, Nebraska, U.S.A) coupled to a Datalogger (LICOR 1400, Lincoln, Nebraska, U.S.A). The values for intercepted global radiation (IGR) were measured serially at each individual leaf strata (lower, middle and upper) with the portable pyranometer which recorded measurements in the period from 10 h to 12 h. The values of intercepted solar radiation were obtained according to the following equation:
Where: Rn = incident radiation under each leaf strata of the canopy; Rt = incident radiation above the canopy.
The extinction coefficient (k) was calculated for each leaf strata using the following equation:
Where k = extinction coefficient, Rn = solar radiation measured under each canopy stratum (MJ m-2); Rt = radiation above the plant canopy (MJ m-2); LAI = leaf area index (m-2 m-2).
The production of dry matter was based on the model proposed by Monteith (1977), where dry matter production was calculated from intercepted photosynthetically active radiation (PARi) multiplied by the radiation use efficiency. The radiation use efficiency (RUE) was calculated by the relation between the average production of accumulated TDM and the PARi involved in the production of biomass according to the following expression:
Where TDM = total dry matter produced (g m-2); PARi = intercepted photosynthetically active radiation (MJ m-2) and RUE = radiation use efficiency in dry matter produced (g MJ-1). The value for the radiation use efficiency given by the angular coefficient represents the amount of accumulated biomass for each unit of intercepted radiation.
Values for intercepted photosynthetically active radiation were determined based on the model proposed by Varlet-GrancherVARLET-GRANCHER C, GOSSE G, CHARTIER M, SINOQUET H, BONHOMME R and ALLIRAND JM. 1989. Mise au point: rayonnement solaire absorbé ou intercepté par un couvert végétal. Agronomie 9: 419-439. et al. (1989):
Where: PARi = intercepted photosynthetically active radiation (MJ m-2); PARin = incident photosynthetically active radiation (MJ m-2); LAI = leaf area index; k = extinction coefficient, 0.67 and 0.60 were used for the extinction coefficient in the crop year 2013/2014 and 2014/2015, respectively, the values used for k were averages obtained in this study and, calculated using the equation described above.
The values of photosynthetically active radiation were estimated as 45 % of global solar radiation. This fraction represents the average values found by AssisASSIS FN and MENDEZ MEG. 1989. Relação entre radiação fotossinteticamente ativa e radiação global. Pesq Agropec Bras 24: 797-800. and Mendez (1989) for the area of Rio Grande do Sul. The estimate of accumulated photosynthetically active radiation was performed, based on methods by Monteith (1977) and Varlet-Grancher et al. (1989).
YIELD TRAITS AND DATA ANALYSIS
The yield components were analyzed by assessing ten randomly plants in each previously demarcated plot, at harvest time. The criterion used for the collection of these 10 plants was that these should represent in a reliable way the characteristics of growth and development of the total plants of each plot. The number of pods per plant (NPP) was counted. Additionally, the 1000-grain weight (GW) was assessed through threshing the pods, counting grains and weighed on a precision balance. To assess the yield, the total grains obtained in each canopy strata for each evaluation plot were weighed on a precision balance and the moisture of the grain converted for 13 %. The yield value obtained in g plot-1 was converted into kg ha-1.
Data were analyzed using the software Statistical Analysis System (SAS, Edition 8.0 2003SAS- STATISTICAL ANALYSIS SYSTEM. 2003. Getting started with the SAS Learning Edition. Cary, 200 p., SAS Institute Inc., Cary N.C., U.S.A.). Data were initially examined for homogeneity of variance, and then, subjected to analysis of variance in order to determine treatment effects and possible interactions among cultivar, crop year and canopy strata. The normality distribution of all data was checked using the Shapiro–Wilk test. The descriptive analysis were performed including the respective error bars, which represent the standard deviation of the average values. For the radiation use efficiency and soybean yield traits, a Tukey test (p<0.05) was performed to verify the difference between treatments.
RESULTS
METEOROLOGICAL CONDITIONS
Air temperature (maximum, minimum and average), incident solar radiation and water balance during the experimental period are shown in Figure 3. In the crop year 2013/2014, the temperature ranged between 12.4 °C and 36.8 °C with an average temperature of 22.8 °C. The flux of global solar radiation was 25.48 MJ m-2 day-1 on average, with a variation of 5.12 to 39.21 MJ m-2 day-1. Also, was observed a small water deficit in February (2013/2014). For the crop year 2014/2015, the air temperature ranged from 11.9 to 34.5 °C with an average temperature of 22.6 °C. The average flux of global solar radiation was 24.30 MJ m-2 day-1 with a variation of 5.55 to 38.12 MJ m-2 day-1. It was possible to identify periods of water deficit during the plant growth.

Monthly averages of minimum, maximum and average air temperatures, accumulated incident solar radiation, and water balance during the experimental period in Frederico Westphalen, Brazil, ‘A’ – crop year 2013/2014 and ‘B’ – crop year 2014/2015.
RADIATION USE EFFICIENCY AND ASSIMILATE PARTITIONING
The analysis of variance revealed a significant interaction “crop year x cultivar” for RUE. Dry matter formation was positively, linearly correlated with PARi during the crop cycle, with high correlation coefficients (Fig. 4). The highest RUE was observed in the crop year 2014/2015 for the cultivar TEC6029, which had 1.5070 g MJ-1, not differing from cultivar NA6411 which obtained 1.4676 g MJ-1. In the crop year 2013/2014, lower values of RUE were observed (1.1518 and 1.3839 g MJ-1 for the cultivars TEC6029 and NA6411 respectively).

Relationship between accumulated intercepted photosynthetically active radiation (PARiac) and dry matter produced (TDM) in a field experiment in Frederico Westphalen, Brazil, with the soybean cultivars ‘a’ – TEC6029 and ‘b’ – NA6411 (crop year 2013/2014); ‘c’ – TEC6029 and ‘d’ – NA6411 (crop year 2014/2015). Means followed by the same letter, lowercase comparing the crop years and uppercase comparing cultivar not differ by Tukey test at 0.05 probability of error.
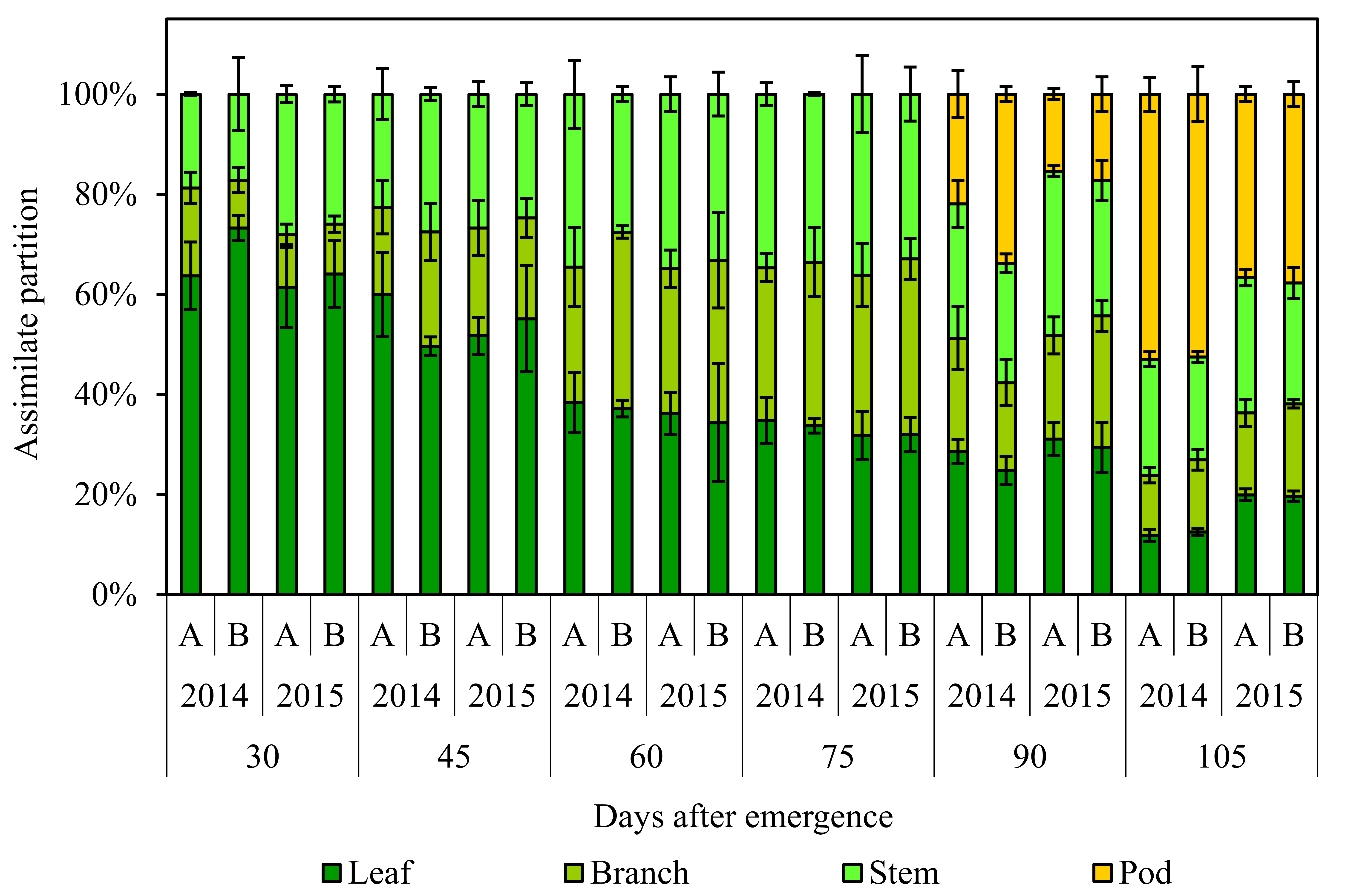
The pattern of dry matter accumulation in the leaves, stem, branches, and pods of the soybean plants was similar during all evaluations periods (Fig. 5). In the initial stages of plant growth, up to 45 DAE, the leaves, stems, and branches were responsible for 60, 25 and 15 % on average of total dry matter accumulation. From 90 DAE the dry matter of the pods was computed, and for the 105 DAE, the pods were responsible for 42 % of total dry matter accumulated by the soybean crop.

Assimilate partitioning to different structural components of soybean plants (leaf, branch, stem, and pod) for the cultivars ‘A’ - NA6411 and ‘B’ - TEC6029 in two crop years (2013/2014 and 2014/2015) throughout all evaluation periods in a field experiment in Frederico Westphalen, Brazil. Bars represent the standard deviation of mean values ± SE.
LEAF AREA INDEX AND INTERCEPTED GLOBAL RADIATION
In both cultivars studied, LAI averaged highest at 90 DAE (Fig. 6). In the crop year 2014/2015, the greatest LAI values were observed for the cultivar TEC6029 (4.85) and for NA6411 (4.34), while for 2013/2014 the lowest values were 4.40 and 4.15 for the cultivars TEC6029 and NA6411, respectively. Additionally, each plant canopy strata showed similar contributions considering the years and cultivars studied. In this way, the lower, middle and upper strata were responsible for 19.6, 48.1 and 32.3 % of the total LAI, respectively.
A positive linear increment of the specific leaf area up to 75 DAE was observed. After this period, a decrease in the values was found due to the physiological maturity (Fig. 6). The greatest average values for this characteristic was observed for the cultivar TEC6029 throughout the evaluation periods in both crop years.

Leaf area index (LAI), specific leaf area (SLA) and intercepted global radiation (IGR) in different canopy strata of the soybean cultivars ‘A’ – NA6411 and ‘B’ - TEC6029 in two crop years (2013/2014 and 2014/2015) throughout all evaluation periods in a field experiment in Frederico Westphalen, Brazil. Error bars represent the standard deviation of mean values ± SE.
The cultivars responded similarly in terms of the interception of incident solar radiation in the different crop years (Fig. 6); highest values were found at 90 DAE where the plants intercepted on average 89.90 % of the incident radiation, coinciding with the greater LAI values. Each plant canopy strata intercepts an amount of solar radiation dependent upon leaf area index. Additionally, the contribution of each plant canopy strata to total radiation interception at 90 DAE was 8.87, 15.95 and 65.07 % for the lower, middle and upper strata, respectively. A reduction of radiation interception was observed at 105 DAE, possibly due to the beginning of leaf senescence.
LIGHT EXTINCTION COEFFICIENT
The k estimate in the different plant canopy strata can be seen in Table I. The k values increased up to 75 DAE and decreased afterwards until senescence. Regarding the difference between cultivar and plant strata, the average values for the crop year 2013/2014 were 0.43, 0.66, 0.76, 0.86, 0.62 and 0.52, and for the crop year 2014/2015 average values of 0.29, 0.57, 0.65, 0.77, 0.61 and 0.45 were obtained during the evaluation periods (30, 45, 60, 75, 90 and 105 DAE), respectively. Considering all evaluation periods, we found an average k value of 0.67 in the crop year 2013/2014 and 0.59 in the crop year 2014/2015.
The greatest estimates of k were obtained in the crop year 2013/2014 for the cultivar NA6411, in a majority of the evaluation periods. The lower canopy strata showed higher k values, demonstrating the dynamics of solar radiation interception within the plant canopy being dependent upon the LAI of each individual strata.
SOYBEAN YIELD TRAITS IN EACH CANOPY STRATA
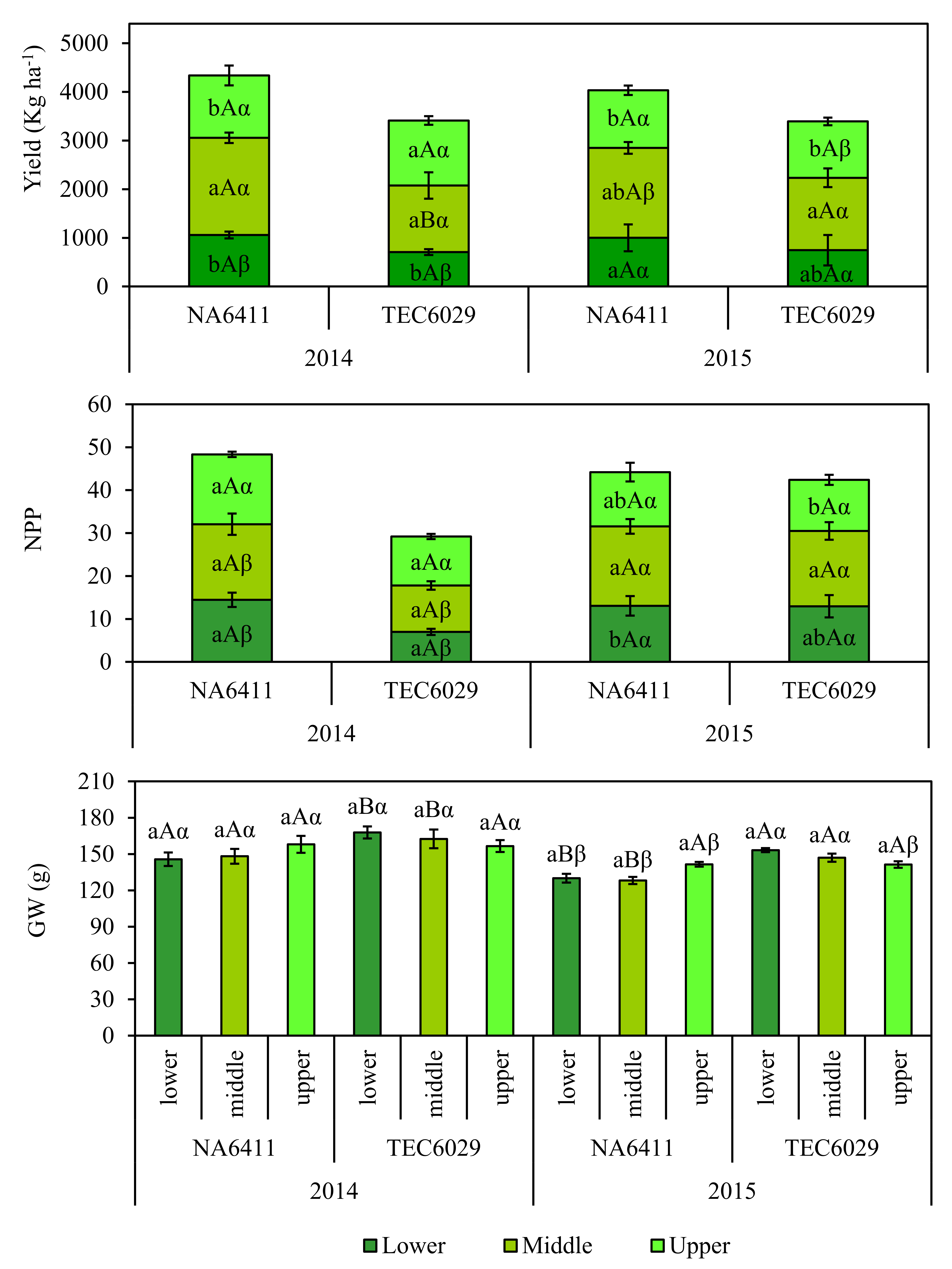
Here we reported results of a detailed investigation of the microclimate within different types of soybean canopies; one with determined growth habit and other with undetermined. Yield of cultivar NA6411 (4336.9 and 4032.2 kg ha-1 in the crop years 2013/2014 and 2014/2015, respectively) was significantly higher than cultivar TEC6029 (3411.6 and 3392.1 kg ha-1 in 2013/2014 and 2014/2015, respectively) (Fig. 7). Among the three plant canopy strata, the middle contributed most to the total yield, followed by the upper, and lower strata.

Yield, number of pods per plant (NPP) and 1000-grain weight (GW) in different canopy strata of the soybean cultivars NA6411 and TEC6029 in two crop years (2013/2014 and 2014/2015) in a field experiment in Frederico Westphalen, Brazil. Bars represent the standard deviation of mean values ± SE. * Means followed by the same letter, lowercase comparing the crop years, uppercase soybean cultivars and greek letters comparing the canopy strata did not differ significantly (Tukey test at 0.05 probability of error).
The number of pods per plant corresponded with the yield pattern, where the highest values were found in the cultivar NA6411 in both crop years (Fig. 7). The same trend was observed for the different canopy strata, as the middle and upper strata presented a larger contribution to the total number of pods per plant. Regarding 1000-grain weight, the greatest values were found in the crop year 2013/2014. The 1000-grain weight of the cultivar TEC6029 was greater than NA6411, in both crop years. Variations in 1000-grain weight were not observed for the different plant canopy strata and crop years.
DISCUSSION
RADIATION USE EFFICIENCY AND ASSIMILATE PARTITIONING
Each cultivar had a differentiated ability to convert radiation into photoassimilates. The lower radiation use efficiency obtained in the crop year 2013/2014 can be related to the water deficit which occurred in February 2014; together with an increase of the maximum air temperature (Fig. 3), this led to a senescence in the leaves of the middle and lower canopy strata. This premature senescence did not affect the grain yield.
The greater radiation use efficiency obtained by the cultivar TEC6029 in the crop year 2013/2014 may be related to the higher LAI associated with the greater use of available diffuse radiation in the lower plant canopy strata. Thus, greater leaf formation resulted in a better use of diffuse radiation, which is multidirectional, and more efficient in penetrating the plant canopy (BuriolBURIOL GA, STRECK NA, PETRY C and SCHNEIDER FM. 1995. Transmissividade a radiação solar do polietileno de baixa densidade utilizado em estufas. Cienc Rural 25: 1-4. et al. 1995).
According to CasaroliCASAROLI D, FAGAN EB, SIMON J, MEDEIROS SP, MANFRON PA, DOURADO NETO D, VAN LIER QJ, MULLER L and MARTIN TN. 2007. Solar radiation and physiologics aspects in soybean - a review. Revista da FZVA 14: 102-120. et al. (2007), the use of cultivars with more efficient leaf architecture may provide more efficient photosynthesis; however, these plants are often more demanding in water and nutrients due to the greater interception of solar radiation by the leaves and so, have a greater energetic demand. This response may affect radiation use efficiency of soybean plants.
The values of radiation use efficiency obtained in this study were similar to that has been observed by other authors; Souza et al. (2009)SOUZA PJOP, RIBEIRO A, ROCHA EJP, FARIAS JRB, LOUREIRO RS, BISPO CC and SAMPAIO L. 2009. Solar radiation use efficiency by soybean under field conditions in the Amazon region. Pesq Agropec Bras 44: 1211-1218. in a study conducted in two crop years (2007/2008 with the occurrence of drought and 2008/2009 without drought), found RUE values of 1.14 and 1.75 g MJ-1, respectively. VanVAN HEERDEN PDR, DONALDSON RA, WATT DA and SINGELS A. 2010. Biomass accumulation in sugarcane: unraveling the factors underpinning reduced growth phenomena. J Exp Bot 61: 2877-2887. Roekel and Purcell (2014), in a study conducted in Missouri, USA, found RUE values for soybean in different crop years (2012/2013 and 2013/2014) of 1.01 and 1.77 g MJ-1, respectively. Also, other results were reported in the literature: 1.23 g MJ-1 (SchöffelSCHÖFFEL ER and VOLPE CA. 2001. Eficiência de conversão da radiação fotossinteticamente ativa interceptada pela soja para produção de fitomassa. Rev Bras Agrometeorol 9: 241-249. and Volpe 2001); 1.31 g MJ-1 (AdeboyeADEBOYE OB, SCHULTZ B, ADELAKU O and PRASAD K. 2016. Impact of water stress on radiation interception and radiation use efficiency of soybeans (Glycine max L. Merr.) in Nigeria. Braz J Sci and Technol 3: 1-15. et al. 2016); 1.44 g MJ-1 (Singer et al. 2011SINGER JW, MEEK DW, SAUER TJ, PRUEGER JH and HATFIELD JL. 2011. Variability of light interception and radiation use efficiency in maize and soybean. Field Crops Res 121: 147-152.); and 1.78 g MJ-1 (Pengelly et al. 1999PENGELLY BC, BLAMEY FPC and MUCHOW RC. 1999. Radiation interception and the accumulation of biomass and nitrogen by soybean and three tropical annual forage legumes. Field Crops Res 63: 99-112.) for the soybean crop. The variation in the results found in the literature was probably due to the differences of the meteorological conditions to which plants were subjected. This was likely because the radiation use efficiency reflects the capacity of the plant to transform the intercepted radiation into dry matter.
Regarding the partitioning of assimilates produced, the biomass accumulated in the leaves decreased gradually throughout the crop cycle, especially during the maturation stage due to the senescence of the lower leaves and the reallocation of the assimilates for grain filling (Fig. 4).
The assimilate translocation to sink organs is governed by factors such as the proximity of the source to the sink. In this way, the leaves located in the lower stratum direct their assimilates to the roots, while the leaves of the upper strata send assimilates to the growing apices and the leaves of the middle strata for both directions (Taiz et al. 2017). Therefore, the position of the leaves within each plant canopy strata is important for the supply of assimilates to vegetative/reproductive structures. Studies of AliyevALIYEV JA. 2010. Photosynthesis, photorespiration and productivity of wheat and soybean genotypes. Biol Sci 65: 7-48. (2010) and Aliyev and Mirzoyev (2010)ALIYEV JA and MIRZOYEY RS. 2010. Photosynthesis and Productivity of Soybean (Glycine max L.). Biol Sci 65: 60-70. indicate that the soybean yield is related to the shape and arrangement of leaves in the canopy strata, photosynthetic activity and the period of grain filling.
The relationship between total dry matter and grain yield depends on the cultivar used and its interaction with the micrometeorological conditions. However, increased solar radiation use efficiency will not necessarily result in higher grain yields. This means that a cultivar can present variations in the efficiency of the use of solar radiation in different years, due to the interannual variability of solar radiation availability. This fact will not necessarily result in an increase or decrease in crop yield (Koester et al. 2014KOESTER RP, SKONECZA JA, CARY TR, DIERS BW and AINSWORTH EA. 2014. Historical gains in soybean (Glycine max Merr.) seed yield are driven by linear increases in light interception, energy conversion, and partitioning efficiencies. J Exp Bot 65: 3311-3321., Petter et al. 2016PETTER FA, DA SILVA JA, ZUFFO AM, ANDRADE FR, PACHECO LP and DE ALMEIDA FA. 2016. Elevada densidade de semeadura aumenta a produtividade da soja? Respostas da radiação fotossinteticamente ativa. Bragantia. 75: 173-183.).
MICROCLIMATIC RELATIONSHIP BETWEEN LEAF AREA INDEX AND INTERCEPTED SOLAR RADIATION
The soybean plants obtained a high leaf area production due to a rapid emission of leaves, promoting significant increases in the interception of solar radiation. However, with the beginning of the self-shading of the leaves, there is an increase in the light extinction coefficient. With the increase of LAI, up to a critical value, the solar radiation interception and the photosynthetic rate are also increased (Casaroli et al. 2007).
According to Souza et al. (2009), for the soybean crop, the critical LAI value to intercept 95 % of the solar radiation is around 4.0. In this context, the average values obtained in this study are above the critical value of LAI, averaging 4.4 at 90 DAE in the two evaluation years, and promoting the interception of 89.9 % of incident solar radiation.
The highest LAI and specific leaf area were obtained for the upper and middle strata, which promoted the formation of a dense leaf strata and making it difficult the solar radiation to penetrate into the lowest strata. In this way, due the reduction in the solar radiation availability in the lower canopy strata an unequal contribution of the plant sections to the total yield was observed (Fig. 7).
The greatest interception of solar radiation by the soybean cultivars occurred in the upper canopy strata, resulting in a greater photosynthetic rate (Taiz et al. 2017). It might be inferred that the lower strata of the leaves contributed less than they theoretically could for the photosynthetic process, while other strata operated more close to their limit. In this context, the use of alternatives in order to increase the photosynthetic rates of the middle and lower plant strata is important because it could lead to an increase in the overall grain yield of the soybean crop.
LIGHT EXTINCTION COEFFICIENT AND SOLAR RADIATION DYNAMICS
Soybean growth was correlated to an increase in the k values (Table I), coinciding with an increase in LAI. Increased LAI leads to a greater quantity of intercepted radiation, up to a certain value when the plants reached critical LAI and begin self-shading, resulting in an increase in the light extinction coefficient. According to BehlingBEHLING A, SANQUETTA DR, DALLA CORTE AP, NETTO SP, RODRIGUES AL, CARON BO and SIMON AA. 2016. Tracking leaf area index and coefficient of light extinction over the harvesting cycle of black wattle. J of For Res 27: 1211-1217. et al. (2016), this attenuation of light by the plant canopy is determined by leaf density (which can be expressed by LAI), and by the geometric and optical properties characteristics of the leaves. Thornley (1976)THORNLEY JHM. 1976. Mathematical models in plant physiology. Academic Press, London, p. 318. reported that the value of k might vary with leaf traits, sun’s incident angle, spacing, and latitude.
Light extinction coefficients (± standard error) in different canopy strata of the two soybean cultivars (NA6411 and TEC6029) in two crop years (2013/2014 and 2014/2015) at different evaluation periods in a field experiment in Frederico Westphalen, Brazil.
The k values observed in our study are similar to those obtained by BajgainBAJGAIN R, KAWASAKI Y, AKAMATSU Y, TANAKA Y, KAWAMURA H, KATSURA K and SHIRAIWA T. 2015. Biomass production and yield of soybean grown under converted paddy fields with excess water during the early growth stage. Field Crops Res 180: 221-227. et al. (2015) who reported k values of 0.85 and 0.65 for soybean cultivation under an excess of water and optimal humidity conditions, respectively. Procópio et al. (2003)PROCÓPIO SO, SANTOS JB, SILVA AA and COSTA LC. 2003. Desenvolvimento folhar das culturas da soja e do feijão e de planta daninha. Cienc Rural 33: 207-211., found k values of 0.52 and 0.93 for soybeans before and after the flowering stage, respectively. Additionally, Pengelly et al. (1999) found that the light extinction coefficient might vary from 0.5 to 0.6 for soybean crops, considering an average of the crop cycle. In a study conducted in Nigeria by Adeboye et al. (2016), the authors obtained average k values of 0.52 and 0.42 for two crop years 2013/2014 and 2014/2015, respectively.
YIELD TRAITS AND ITS RELATIONSHIP WITH PHOTOSYNTHETIC ACTIVITY
The highest grain yield was obtained by the cultivar with a determinate growth habit and was mainly influenced by the greater number of pods per plant in the middle plant strata, since for this growth type, the flowering begins in the middle layer and then goes to the upper and lower. This response was most evident for the crop year 2013/2014. Moreover, cultivars with determined growth habit had a lower dense layer of leaves in the upper strata, which allows a greater interception of radiation in the middle and lower canopies strata when compared to the cultivar with an indeterminate growth habit.
For the cultivar with indeterminate growth habit, it is observed a dense layer of leaves in the upper strata due to the continuous emission of leaves even after flowering, this fact can reduce the fixation of the flowers in the middle and lower strata. Therefore, considering the interception dynamics of the solar radiation and the greater contribution of the middle and lower strata to the cultivar with determined growth habit presented higher grain yield.
Studies that relate productivity and type of plant growth indicate a tendency of determined cultivars, under irrigated conditions, to produce more than the indeterminate cultivars (Kilgore-Norquest and Sneller 2000KILGORE-NORQUEST L and SNELLER CH. 2000. Effect of stem termination on soybean traits in southern U.S. production systems. Crop Sci 40: 83-90.). According to Perini et al. (2012)PERINI LJ, JÚNIOR NDSF, DESTRO D and PRETE CEC. 2012. Componentes da produção em cultivares de soja com crescimento determinado e indeterminado. Semin Ciênc Agrár 33: 2531-2544., the total number of pods was the yield traits that was most important for the indirect selection of more productive and determined growth cultivars.
In overall, the highest contribution of the medium and upper strata of the plants to the total yield may be related to the higher leaf area index and interception of the solar radiation obtained in these sections. These results of total yield and number of pod per plant may be related to a greater photosynthetic activity of these sections, resulting in an increase of the production and direction of assimilates for the reproductive structures which was seen in the medium and upper strata.
The results observed are consistent with those of Rambo et al. (2004), evaluating the potential yield of each canopy strata of the soybean crop. These authors found that the upper and middle plant strata contributed strongly to the soybean yield potential. The greater number of pods in each section explained this. Passos et al. (2011)PASSOS AMA, REZENDE PM, ALVARENGA AA, BALIZA DP, CARVALHO ER and ALCÂNTARA HP. 2011. Yield per plant and other characteristics of soybean plants treated with kinetin and potassium nitrate. Ciênc Agrotec 35: 965-972. reported that the middle and upper strata of the soybean canopy has a greater contribution to grain yield, due to the greater presence of pods in these sections. The authors also highlighted that the middle strata represented about 57.6 % of the total number of pods produced in the plants.
The stratified canopy study of soybean plants enables the identification of contrasting characteristics within the plant population, which together with the best use of the micrometeorological conditions, mainly of diffuse radiation, is essential for increasing the radiation use efficiency and consequently the grain yield.
Information generated in this study are relevant and provides knowledge for soybean producers and agricultural managers, especially regarding the microclimatic conditions and its relations with the canopy architecture, growth habit, as well as the contribution of each canopy strata to the total yield. Moreover, the results highlighted in this study can help to understand the dynamics within the plant’s canopy and adapt possible management strategies such as the use of microclimatic information for the monitoring of plant diseases in order to maximize grain yield in the lower plant strata.
CONCLUSIONS
The cultivar NA6411, with a determinate growth habit, presented a greater interception of solar radiation in the middle and lower canopies strata which results in higher soybean yield when compared to the cultivar TEC6029, and thus, can be recommended for cultivation in crop production systems.
The contribution of the middle and upper canopy strata to the total yield formation was greater than that observed in the lower canopy strata due to the greater interception of solar radiation by these strata. To increase soybean yields, new studies regarding the microclimatic conditions of the soybean canopy should be developed to improve the maximum potential yield of the new soybean cultivars.
ACKNOWLEGMENTS
The authors thank to Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the productivity scholarship of the co-author Braulio Otomar Caron and for their financial support for the author Felipe Schwerz.
REFERENCES
- ADEBOYE OB, SCHULTZ B, ADELAKU O and PRASAD K. 2016. Impact of water stress on radiation interception and radiation use efficiency of soybeans (Glycine max L. Merr.) in Nigeria. Braz J Sci and Technol 3: 1-15.
- ALIYEV JA. 2010. Photosynthesis, photorespiration and productivity of wheat and soybean genotypes. Biol Sci 65: 7-48.
- ALIYEV JA and MIRZOYEY RS. 2010. Photosynthesis and Productivity of Soybean (Glycine max L.). Biol Sci 65: 60-70.
- ALVARES CA, STAPE JL, SENTELHAS PC, MORAES G, LEONARDO J and SPARROVEK G. 2013. Köppen’s climate classification map for Brazil. Meteorol Z 22: 711-728.
- ASSIS FN and MENDEZ MEG. 1989. Relação entre radiação fotossinteticamente ativa e radiação global. Pesq Agropec Bras 24: 797-800.
- BAJGAIN R, KAWASAKI Y, AKAMATSU Y, TANAKA Y, KAWAMURA H, KATSURA K and SHIRAIWA T. 2015. Biomass production and yield of soybean grown under converted paddy fields with excess water during the early growth stage. Field Crops Res 180: 221-227.
- BALDOCCHI DD, VERMA SB and ROSENBERG NJ. 1983. Microclimate in the soybean canopy. Agric Meteorol 28: 321-337.
- BALDOCCHI DD, VERMA SB, ROSENBERG NJ, BLAD BL and SPECHT JE. 1985. Microclimate-plant architectural interactions: influence of leaf width on the mass and energy exchange of a soybean canopy. Agr For Meteorol 35: 1-20.
- BEHLING A, SANQUETTA DR, DALLA CORTE AP, NETTO SP, RODRIGUES AL, CARON BO and SIMON AA. 2016. Tracking leaf area index and coefficient of light extinction over the harvesting cycle of black wattle. J of For Res 27: 1211-1217.
- BULER Z and MIKA A. 2009. The influence of canopy architecture on light interception and distribution in ‘sampion’ apple trees. J Fruit Omam Plant Res 18: 96-100.
- BURIOL GA, STRECK NA, PETRY C and SCHNEIDER FM. 1995. Transmissividade a radiação solar do polietileno de baixa densidade utilizado em estufas. Cienc Rural 25: 1-4.
- CASAROLI D, FAGAN EB, SIMON J, MEDEIROS SP, MANFRON PA, DOURADO NETO D, VAN LIER QJ, MULLER L and MARTIN TN. 2007. Solar radiation and physiologics aspects in soybean - a review. Revista da FZVA 14: 102-120.
- COMISSÃO DE QUÍMICA E FERTILIDADE DO SOLO. 2004. Manual de adubação e calagem para os estados do Rio Grande do Sul e Santa Catarina. Porto Alegre: Sociedade Brasileira de Ciência do Solo – Núcleo Regional Sul, p. 400.
- CONAB - COMPANHIA NACIONAL DE ABASTECIMENTO ACOMPANHAMENTO DA SAFRA BRASILEIRA DE GRÃOS. 2016/2017. Disponível em: <http://www.conab.gov.br/OlalaCMS/uploads/arquivos/16_12_22_12_08_27_boletim_graos_dezembro_2016.pdf>. Acessado em: 15 Jan 2018.
» http://www.conab.gov.br/OlalaCMS/uploads/arquivos/16_12_22_12_08_27_boletim_graos_dezembro_2016.pdf>. - CUNHA AR and VOLPE CA. 2010. Relações radiométricas no terço superior da copa do cafeeiro. Bragantia 69: 263-271.
- EGLI DB, PENDLETON JW and PETERS DB. 1970. Photosynthetic rate of three soybean communities as related to CO2 levels and solar radiation. Agron J 62: 411- 414.
- ELLI EF, CARON BOC, PAULA GM, ELOY E, SCHWERZ F and SCHIMDT D. 2016. Ecophysiology of Peltophorum dubium Spr. In: agroforestry systems arrangements. Comunicata Scientiae 7: 464-472.
- EMBRAPA. 2006. Tecnologias de Produção de Soja – Paraná 2007. Londrina. EMBRAPA, 220 p.
- FAGAN EB, MIQUELANTI NP, BONFIM TS, PEREIRA IS, CORRÊA LT and SOARES JN. 2013. Lei de Beer e sua relação com a ecofisiologia de plantas. Cerrado Agrociência 4: 78-79.
- HIEBSCH CK, KANEMASU ET and NICKELL CD. 1976. Effects of soybean leaflet type on net carbon dioxide exchange, water use and water use efficiency. Can J Plant Sci 56: 455-458.
- KILGORE-NORQUEST L and SNELLER CH. 2000. Effect of stem termination on soybean traits in southern U.S. production systems. Crop Sci 40: 83-90.
- KOESTER RP, SKONECZA JA, CARY TR, DIERS BW and AINSWORTH EA. 2014. Historical gains in soybean (Glycine max Merr.) seed yield are driven by linear increases in light interception, energy conversion, and partitioning efficiencies. J Exp Bot 65: 3311-3321.
- MARCHÃO RL, BRASIL EM and XIMENES PA. 2006. Interceptação da radiação fotossinteticamente ativa e rendimento de grãos do milho adensado. Rev Bras Milho Sorgo 5: 170-181.
- MARIN M, CASTRO B, GAITAN A, PREISIG O, WINGFIELD BD and WINGFIELD MJ. 2003. Relationships of Ceratocystis frimbiata isolates from Colombian coffee-growing regions based on molecular data a pathogenicity. J Phytopathol 151: 395-405.
- MONTEITH JL. 1977. Climate and the efficiency of crop production en Britain. Philos T Roy Soc B 181: 277-294.
- PASSOS AMA, REZENDE PM, ALVARENGA AA, BALIZA DP, CARVALHO ER and ALCÂNTARA HP. 2011. Yield per plant and other characteristics of soybean plants treated with kinetin and potassium nitrate. Ciênc Agrotec 35: 965-972.
- PENGELLY BC, BLAMEY FPC and MUCHOW RC. 1999. Radiation interception and the accumulation of biomass and nitrogen by soybean and three tropical annual forage legumes. Field Crops Res 63: 99-112.
- PERINI LJ, JÚNIOR NDSF, DESTRO D and PRETE CEC. 2012. Componentes da produção em cultivares de soja com crescimento determinado e indeterminado. Semin Ciênc Agrár 33: 2531-2544.
- PETTER FA, DA SILVA JA, ZUFFO AM, ANDRADE FR, PACHECO LP and DE ALMEIDA FA. 2016. Elevada densidade de semeadura aumenta a produtividade da soja? Respostas da radiação fotossinteticamente ativa. Bragantia. 75: 173-183.
- PILAU FG and ANGELOCCI. LR. 2015. Área foliar e interceptação de radiação solar pela copa de uma laranjeira. Bragantia 74: 476-482.
- PROCÓPIO SO, SANTOS JB, SILVA AA and COSTA LC. 2003. Desenvolvimento folhar das culturas da soja e do feijão e de planta daninha. Cienc Rural 33: 207-211.
- RAMBO L, COSTA AJ, PIRES FLJ and FERREIRA GF. 2004. Estimativa do potencial de rendimento por estrato do dossel da soja, em diferentes arranjos de plantas. Cienc Rural 34: 33-40.
- ROLIM GS, SENTELHAS PC and BARBIERI V. 1998. Planilhas no ambiente EXCEL para os cálculos de balanços hídricos: normal, sequencial, de cultura e de produtividade real e potencial. Rev Bras Agrometeorol 6: 133-137.
- SAS- STATISTICAL ANALYSIS SYSTEM. 2003. Getting started with the SAS Learning Edition. Cary, 200 p.
- SCHÖFFEL ER and VOLPE CA. 2001. Eficiência de conversão da radiação fotossinteticamente ativa interceptada pela soja para produção de fitomassa. Rev Bras Agrometeorol 9: 241-249.
- SCHWERZ F, CARON BO, ELLI EF, SOUZA VQ, OLIVEIRA DM and ROCKENBACH AP. 2016. Soybean morphological and productive characteristics influenced by meteorological parameters and sowing dates. Científica 44: 121-130.
- SEDIYAMA T, PEREIRA GP, SEDIYAMA CS and GOMES JLL. 1996. Cultura da Soja: I Parte. Viçosa: UFV, 96 p.
- SEDIYAMA T, TEIXEIRA RC and REIS MC. 1999. Melhoramento da Soja. In: Borém A. Melhoramento de espécies cultivadas. Viçosa: UFV, cap. 14, p. 487-531.
- SINGER JW, MEEK DW, SAUER TJ, PRUEGER JH and HATFIELD JL. 2011. Variability of light interception and radiation use efficiency in maize and soybean. Field Crops Res 121: 147-152.
- SOUZA PJOP, RIBEIRO A, ROCHA EJP, FARIAS JRB, LOUREIRO RS, BISPO CC and SAMPAIO L. 2009. Solar radiation use efficiency by soybean under field conditions in the Amazon region. Pesq Agropec Bras 44: 1211-1218.
- TAIZ L, ZEIGER E, MØLLER IM and MURPHY A. 2017. Fisiologia Vegetal, 6a ed., Porto Alegre: Artmed, p. 888.
- THORNLEY JHM. 1976. Mathematical models in plant physiology. Academic Press, London, p. 318.
- THORNTHWAITE CW and MATHER JR. 1955. The water balance. Publication in climatology. Centerton, New Jersey, p. 104.
- VAN HEERDEN PDR, DONALDSON RA, WATT DA and SINGELS A. 2010. Biomass accumulation in sugarcane: unraveling the factors underpinning reduced growth phenomena. J Exp Bot 61: 2877-2887.
- VAN ROEKEL RJ and PURCELL LC. 2014. Soybean biomass and nitrogen accumulation rates and radiation use efficiency in a maximum yield environment. Crop Sci 54: 1189-1196.
- VARLET-GRANCHER C, GOSSE G, CHARTIER M, SINOQUET H, BONHOMME R and ALLIRAND JM. 1989. Mise au point: rayonnement solaire absorbé ou intercepté par un couvert végétal. Agronomie 9: 419-439.
- WELLS R. 1991. Soybean growth response to plant density: relationships among canopy photosynthesis, leaf area, and light interception. Crop Sci 31: 755-761.
- YANG XF, YU XQ, ZHOU Z, MA WJ and TANG GX. 2016. A high-efficiency Agrobacterium tumefaciens mediated transformation system using cotyledonary node as explants in soybean (Glycine max L.). Acta Physiol Plant 38: 1-10.
- ZANON AJ, STRECK NA, RICHTER GL, BECKER CC, ROCHA TSMD, CERA JC, WINCK JEM, CARDOZO AP, TAGLIAPIETRA EL and WEBER PS. 2015. Branches contribution and leaf area index evolution in modern cultivars of soybean. Bragantia 74: 279-290.
Publication Dates
-
Publication in this collection
09 Sept 2019 -
Date of issue
2019
History
-
Received
22 Jan 2018 -
Accepted
26 Oct 2018