Abstract
The aging process of reservoirs has been extensively investigated; however, little is known about how fish populations are adjusted after many years of impoundment. Thus, this study aimed to compare the diet, length-weight relationship, sizes classes, variation in size, and age of Astyanax lacustris Lütken, 1875 collected from lotic and lentic habitats of an aging reservoir. The study group consisted of 730 captured fishes. We found that specimens collected from lotic habitats had a wider range of size classes (1.0 to 12.0 cm), were linked to a high frequency of juveniles (48.7%), and had greater feeding activity (higher stomach fullness). In contrast, fishes collected from lentic environments exhibited high rates of capture (78.4%), increased frequency of adults (87.3%), and higher values of mean standard length. The length-weight relationship indicated that lentic fishes were heavier than fish collected from lotic areas. Moreover, we observed 37 food items in A. lacustris diet, mainly plant material, algae, Cladocera, Hymenoptera, Coleoptera, and Ephemeroptera. Differences among the diet of fishes between sites were evidenced with Permanova (p < 0.05). Astyanax lacustris can be considered a persistent species in Chavantes Reservoir after aging, encountering conditions to complete its life span and adjusting to food resources.
Key words
Characidae; native species; reservoir aging; upper Paraná River basin
INTRODUCTION
Reservoirs built to generate electricity are conspicuous components of the global landscape (Grill et al. 2015GRILL G, LEHNER B, LUMSDON AE, MACDONALD GK, ZARFL C & LIERMANN CR. 2015. An index-based framework for assessing patterns and trends in river fragmentation and flow regulation by global dams at multiple scales. Environ Res Lett 10: 015001.). There are over 59,000 large dams in the world (ICOLD 2018ICOLD - INTERNATIONAL COMMISSION ON LARGE DAMS. 2018. World register on dams – General synthesis. Available at: http://www.icold-cigb.org/GB/World_register/general_synthesis.asp.
http://www.icold-cigb.org/GB/World_regis...
) built for multiple purposes, such as irrigation, flow containment, and water supply (Tundisi & Matsumura-Tundisi 2003TUNDISI JG & MATSUMURA-TUNDISI T. 2003. Integration of research and management in optimizing multiple uses of reservoirs: the experience in South America and Brazilian case studies. Aquat Biodiversity 500: 231-242.), but the main use is to produce hydropower (Agostinho et al. 2016AGOSTINHO AA, GOMES LC, SANTOS NCL, ORTEGA JCG & PELICICE FM. 2016. Fish assemblages in Neotropical reservoirs: Colonization patterns, impacts and management. Fish Res 173: 26-36.).
The construction of dams and associated reservoirs causes irreversible impacts, affecting the structure and composition of the fish fauna (Poff et al. 2007POFF NL, OLDEN JD, MERRITT DM & PEPIN DM. 2007. Homogenization of regional river dynamics by dams and global biodiversity implications. Proc Natl Acad Sci 104: 5732-5737., Pelicice et al. 2015PELICICE FM, POMPEU PS & AGOSTINHO AA. 2015. Large reservoirs as ecological barriers to downstream movements of Neotropical migratory fish. Fish Fish 16: 697-715.). The alterations in the dammed environments tend to reach trophic stabilization over time, i.e., as its functional age progresses (Agostinho et al. 2007AGOSTINHO AA, GOMES LC & PELICICE FM. 2007. Ecologia e manejo de recursos pesqueiros em reservatórios do Brasil, 1st ed., Maringá: Eduem, 501 p., Miranda & Krogman 2015MIRANDA LE & KROGMAN RM. 2015. Functional age as an indicator of reservoir senescence. Fish 40: 170-176., Lima et al. 2018LIMA FP, NOBILE AB, FREITAS-SOUZA D, CARVALHO ED & VIDOTTO-MAGNONI AP. 2018. Can dams affect the trophic structure of ichthyofauna? A long-term effects in the Neotropical region. Iheringia Série Zool 108: e2018030.).
Dams impact biodiversity and ecosystem services. Aquatic richness and diversity are impacted by the transformation from lotic to lentic environment, fragmentation, and success of invasive species in reservoirs (Turgeon et al. 2019TURGEON K, TURPIN C & GREGORY-EAVES I. 2019. Dams have varying impacts on fish communities across latitudes: A quantitative synthesis. Ecol Letters. 22: 1501-1516. doi: https://doi.org/10.1111/ele.13283.). In Brazil, several studies have assessed such impacts on aquatic biodiversity (Vasconcelos et al. 2014VASCONCELOS LP, ALVES DC & GOMES LC. 2014. Fish reproductive guilds downstream of dams. J Fish Biol 85: 1489-1506.). However, few specifically determined the influence of reservoirs on the population biology of fish species (Delariva et al. 2013DELARIVA RL, HAHN NS & KASHIWAQUI EAL. 2013. Diet and trophic structure of the fish fauna in a subtropical ecosystem: impoundment effects. Neotrop Ichthyol 11: 891-904.), especially over several years after dam closure.
Fish species are differentially influenced by each phase of reservoir formation. After the filling phase, reservoirs undergo a heterotrophic period (upsurge period), with increase in fish abundance (Agostinho et al. 2016AGOSTINHO AA, GOMES LC, SANTOS NCL, ORTEGA JCG & PELICICE FM. 2016. Fish assemblages in Neotropical reservoirs: Colonization patterns, impacts and management. Fish Res 173: 26-36.). During the post-heterotrophic period, environmental filters, referred as abiotic factors that prevent the establishment or persistence of species in a given location (Kraft et al. 2015KRAFT NJ, ADLER PB, GODOY O, JAMES EC, FULLER S & LEVINE JM. 2015. Community assembly, coexistence and the environmental filtering metaphor. Funct Ecol 29: 592-599.) gradually remove pre-existing fluvial species (Gomes & Miranda 2001GOMES LC & MIRANDA LE. 2001. Riverine characteristics dictate composition of fish assemblages and limit fisheries in reservoirs of the Upper Paraná River Basin. Regulated Rivers: Research & Management: An International Journal Devoted to River Research and Management 17: 67-76., Agostinho et al. 2016AGOSTINHO AA, GOMES LC, SANTOS NCL, ORTEGA JCG & PELICICE FM. 2016. Fish assemblages in Neotropical reservoirs: Colonization patterns, impacts and management. Fish Res 173: 26-36.). The absence of pre-adapted species colonizing the pelagic zone of reservoirs leads to a concentrated biological diversity and abundance in the littoral zone (Mol et al. 2007MOL JH, MÉRONA BD, OUBOTER PE & SAHDEW S. 2007. The fish fauna of Brokopondo Reservoir, Suriname, during 40 years of impoundment. Neotrop Ichthyol 5: 351-368., Agostinho et al. 2016AGOSTINHO AA, GOMES LC, SANTOS NCL, ORTEGA JCG & PELICICE FM. 2016. Fish assemblages in Neotropical reservoirs: Colonization patterns, impacts and management. Fish Res 173: 26-36.), with the presence of several small and medium-sized non-migratory species and their juveniles. Their presence is due to the amounts of nutrient and food supply, and a higher spatial heterogeneity, which provides shelter to individuals (Casatti et al. 2003CASATTI L, MENDES HF & FERREIRA KM. 2003. Aquatic macrophytes as feeding site for small fishes in the Rosana Reservoir, Paranapanema River, Southeastern Brazil. Braz J Biol 63: 213-222., Pelicice & Agostinho 2006PELICICE FM & AGOSTINHO AA. 2006. Feeding ecology of fishes associated with Egeria spp. patches in a tropical reservoir, Brazil. Ecol Fresh Fish 15: 10-19.). The proliferation of small characins has been documented immediately after the formation of several reservoirs (Agostinho et al. 1999AGOSTINHO AA, MIRANDA LE, BINI LM, GOMES LC, THOMAZ SM & SUZUKI HI. 1999. Patterns of colonization in neotropical reservoirs, and prognoses on aging. In: Tundisi JG & Straskraba M (Orgs), Theoretical reservoir ecology and its applications: Instituto Internacional de Ecologia, São Carlos, p. 227-265., 2007). However, this abundance decreases sharply along time, as the age of the reservoir increases (Agostinho et al. 2007AGOSTINHO AA, GOMES LC & PELICICE FM. 2007. Ecologia e manejo de recursos pesqueiros em reservatórios do Brasil, 1st ed., Maringá: Eduem, 501 p.).
The presence of large tributaries, dam design, and operational procedures are some of the factors that influence the persistence of some species (Agostinho et al. 2007AGOSTINHO AA, GOMES LC & PELICICE FM. 2007. Ecologia e manejo de recursos pesqueiros em reservatórios do Brasil, 1st ed., Maringá: Eduem, 501 p., 2016). Though many studies present fish ecology data (trophic and community analysis) during the initial years of reservoir formation (Loureiro-Crippa & Hahn 2006LOUREIRO-CRIPPA VE & HAHN NS. 2006. Use of food resources by the fish fauna of a small reservoir (rio Jordão, Brazil) before and shortly after its filling. Neotrop Ichthyol 4: 357-362., Cantanhêde et al. 2008CANTANHêDE G, HAHN NS, FUGI R & GUBIANI ÉA. 2008. Alterations on piscivorous diet following change in abundance of prey after impoundment in a Neotropical river. Neotrop Ichthyol 6: 631-636.), few have measured the population-scale (Abelha & Goulart 2008ABELHA MCF & GOULART E. 2008. Population structure, condition factor and reproductive period of Astyanax paranae (Eigenmann, 1914) (Osteichthyes: Characidae) in a small and old Brazilian reservoir. Braz Arch Biol Technol 51: 503-512.) and the long-term ecological changes in the community (Mol et al. 2007MOL JH, MÉRONA BD, OUBOTER PE & SAHDEW S. 2007. The fish fauna of Brokopondo Reservoir, Suriname, during 40 years of impoundment. Neotrop Ichthyol 5: 351-368.). In this manner, it is advised to monitor fish populations to detect possible changes in community on the post-stabilization period of Brazilian reservoirs (Dei Tos et al. 2002DEI TOS C, BARBIERI G, AGOSTINHO AA, GOMES LC & SUZUKI HI. 2002. Ecology of Pimelodus maculatus (Siluriformes) in the Corumbá reservoir, Brazil. Cybium 26: 275-282.).
In Brazil, at least 195 out of 655 reservoirs have more than 40 years of operation (ANEEL 2019ANEEL - AGÊNCIA NACIONAL DE ENERGIA ELÉTRICA. 2019. Acompanhamento de estudos e projetos hidroenergéticos. Available at: http://www2.aneel.gov.br/aplicacoes/capacidadebrasil/UsinaListaSelecao.asp.
http://www2.aneel.gov.br/aplicacoes/capa...
). The ichthyofauna may have undergone alterations over time, and the species that persisted over the years may present different survival strategies (Agostinho et al. 2007AGOSTINHO AA, GOMES LC & PELICICE FM. 2007. Ecologia e manejo de recursos pesqueiros em reservatórios do Brasil, 1st ed., Maringá: Eduem, 501 p.). Since the lentic areas can become less favorable for survival with the aging of the reservoir – and some species are still abundant in old reservoirs – we hypothesize that fish species may have better conditions to complete their life span in the tributaries of the reservoir, searching for riverine conditions that rivers once had before damming.
Astyanax lacustris (early Astyanax altiparanae) (Lucena & Soares 2016LUCENA CD & SOARES HG. 2016. Review of species of the Astyanax bimaculatus “caudal peduncle spot” subgroup sensu Garutti & Langeani (Characiformes, Characidae) from the rio La Plata and rio São Francisco drainages and coastal systems of southern Brazil and Uruguay. Zootaxa 4072: 101-125.) is highly abundant in reservoirs. This small-sized species is widely distributed in the Upper Paraná River basin (Langeani et al. 2007LANGEANI F, CASTRO RMC, OYAKAWA OT, SHIBATTA OA, PAVANELLI CS & CASATTI L. 2007. Diversidade da ictiofauna do Alto Rio Paraná: composição atual e perspectivas futuras. Biota Neotrop 7: 181-197.) and performs short-distance migration (Graça & Pavanelli 2007GRAçA WJ & PAVANELLI CS. 2007. Peixes da planície de inundação do alto rio Paraná e áreas adjacentes, 1st ed., Maringá: Eduem, 241 p.). Moreover, A. lacustris is abundant in rivers and reservoirs (Orsi et al. 2004ORSI ML, CARVALHO ED & FORESTI F. 2004. Biologia populacional de Astyanax altiparanae Garutti & Britski (Teleostei, Characidae) do médio rio Paranapanema, Paraná, Brasil. Rev Bras Zool 21: 207-218.) and can easily adapt to recently impounded rivers (Dias et al. 2005DIAS RM, BAILLY D, ANTôNIO RR, SUZUKI HI & AGOSTINHO AA. 2005. Colonization of the Corumbá Reservoir (Corumbá River, Paraná River basin, Goiás state, Brazil) by “lambari” Astyanax altiparanae (Tetragonopterinae; Characidae). Braz Arch Biol Technol 48: 467-476., Hahn & Fugi 2008HAHN NS & FUGI R. 2008. Environmental changes, habitat modifications and feeding ecology of freshwater fish. In: Cyrino JEP, Bureau DP & Kapoor BG (Eds), Feeding and Digestive Functions in Fishes: Science Publishers, p. 35-65.). The species, known by its reproductive plasticity, reproduces several times in a year in both the lotic and lentic habitats (Orsi 2010ORSI ML. 2010. Estratégias reprodutivas de peixes da região média-baixa do Rio Paranapanema, reservatório de Capivara. São Paulo: Blucher, 113 p.). Astyanax lacustris is a diurnal predator, with good visual acuity (Orsi et al. 2004ORSI ML, CARVALHO ED & FORESTI F. 2004. Biologia populacional de Astyanax altiparanae Garutti & Britski (Teleostei, Characidae) do médio rio Paranapanema, Paraná, Brasil. Rev Bras Zool 21: 207-218.) and colonizes the mid-water and mid-channel habitats (Casatti 2002CASATTI L. 2002. Alimentação dos peixes em um riacho do Parque Estadual Morro do Diabo, bacia do Alto Paraná, sudeste do Brasil. Biota Neotrop 2: 1-14.). The species is a generalist and opportunistic consumer of drift items (Bennemann et al. 2000BENNEMANN ST, SHIBATTA OA & GARAVELLO JC. 2000. Peixes do rio Tibagi: uma abordagem ecológica, 1st ed., Londrina: Eduel, 62 p., Lowe-McConnell 1999LOWE-MCCONNELL RH. 1999. Estudos ecológicos de comunidades de peixes tropicais, São Paulo: Edusp, 534 p., Maroneze et al. 2011MARONEZE DM, TUPINAMBáS TH, ALVEZ CBM, VIEIRA F, POMPEU PS & CALLISTO M. 2011. Fish as ecological tools to complement biodiversity inventories of benthic macroinvertebrates. Hydrobiol 673: 29-40.), including macroinvertebrates (Lima et al. 2018LIMA FP, NOBILE AB, FREITAS-SOUZA D, CARVALHO ED & VIDOTTO-MAGNONI AP. 2018. Can dams affect the trophic structure of ichthyofauna? A long-term effects in the Neotropical region. Iheringia Série Zool 108: e2018030.). Since A. lacustris can be used to investigate the aging processes of reservoirs, we aimed to characterize the population biology of A. lacustris in the lentic environments of the Chavantes Reservoir and its tributaries from the Middle Paranapanema River, Paraná River basin, Brazil.
MATERIALS AND METHODS
Study area
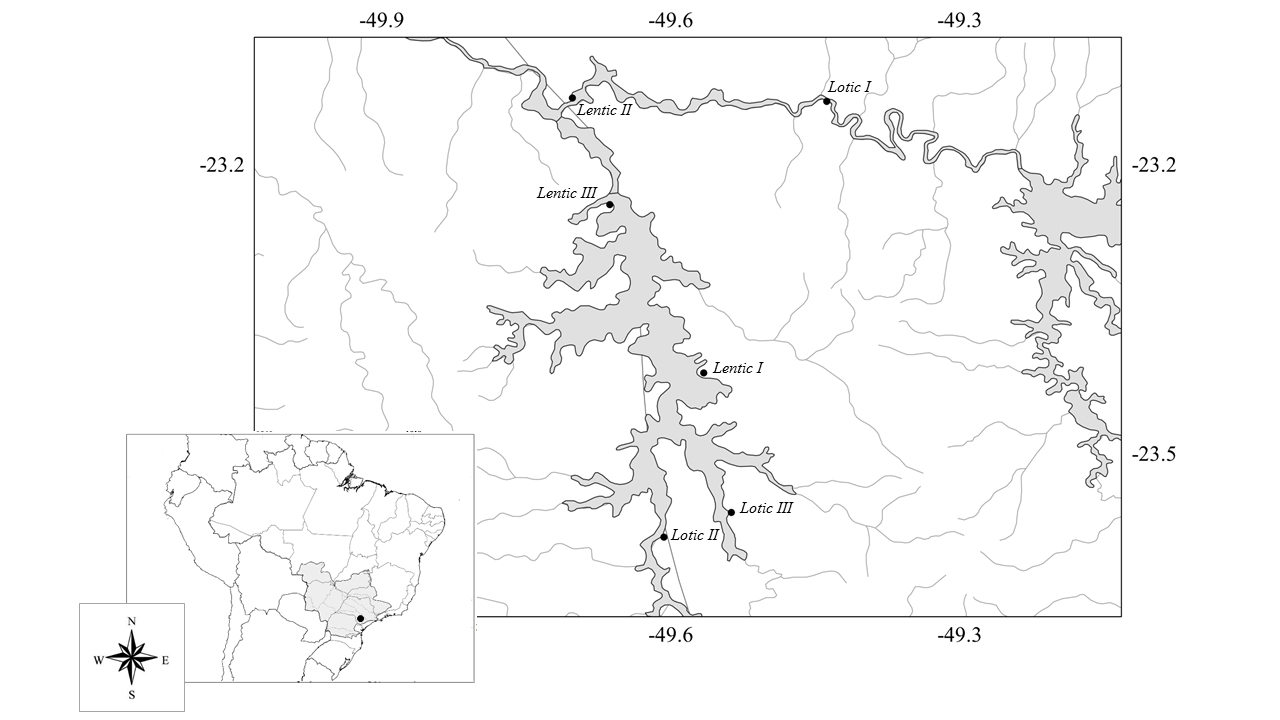
The Paranapanema River is a tributary on the East side of the Paraná River (Fig. 1) that has a drainage area of 106,500 Km2 and elevation of 474 m (ANA 2016ANA - AGENCIA NACIONAL DE ÁGUAS. 2016. Plano Integrado de Recursos Hídricos da Unidade de Gestão de Recursos Hídricos Paranapanema. Agência Nacional Das Águas – Brasília, 354 p.). The Chavantes Reservoir, active for 46 years of operation, is currently classified as old (according to Miranda & Krogman 2015MIRANDA LE & KROGMAN RM. 2015. Functional age as an indicator of reservoir senescence. Fish 40: 170-176.). The Chavantes Reservoir (-49.635111; -23.380378) is located in the middle of the Paranapanema River and is 80 m deep near the dam (Nogueira et al. 2006NOGUEIRA MG, JORCIN A, VIANNA NC & BRITTO YC. 2006. Reservatórios em cascata e os efeitos na limnologia e organização das comunidades bióticas (fitoplâncton, zooplâncton e zoobentos), um estudo de caso no rio Paranapanema (SP/PR). In: Nogueira MG, Henry R & Jorcin A (Eds), Ecologia de reservatórios: Impactos potenciais, ação de manejo e sistemas em cascatas: Rima, São Carlos, p. 435-459.). The water hydraulic retention time is approximately 418 days, which results in a defined thermal stratification below 20 m (Nogueira et al. 2006NOGUEIRA MG, JORCIN A, VIANNA NC & BRITTO YC. 2006. Reservatórios em cascata e os efeitos na limnologia e organização das comunidades bióticas (fitoplâncton, zooplâncton e zoobentos), um estudo de caso no rio Paranapanema (SP/PR). In: Nogueira MG, Henry R & Jorcin A (Eds), Ecologia de reservatórios: Impactos potenciais, ação de manejo e sistemas em cascatas: Rima, São Carlos, p. 435-459.). The reservoir has distinct limnological characteristics and its trophic state is oligo-mesotrophic (Perbiche-Neves & Nogueira 2010PERBICHE-NEVES G & NOGUEIRA MG. 2010. Multi-dimensional, effects on Cladoceran (Crustacea, Anomopoda) assemblages in two cascade reservoirs in Southeast Brazil. Lakes Reservoirs: Res Manage 15: 139-152.) (Fig. 1). Six sampling sites were selected with distinct limnological and habitat characteristics (see Perbiche-Neves & Nogueira 2010PERBICHE-NEVES G & NOGUEIRA MG. 2010. Multi-dimensional, effects on Cladoceran (Crustacea, Anomopoda) assemblages in two cascade reservoirs in Southeast Brazil. Lakes Reservoirs: Res Manage 15: 139-152.). Three were lotic and three were lentic environments (Table I).

Characterization of the lotic and lentic sampling sites studied, influenced by the Chavantes Reservoir, Paranapanema River.
Sampling
Astyanax lacustris were sampled quarterly from October 2005 to July 2006 (IBAMA/ICMBio license: 15549-1). At each sampling stations, fish were captured with gillnets (five sets with five nets each, mesh sizes ranging from 3 to 7 cm between opposite knots), comprising 500 m2 of the net in every sample. Gillnets were set late in the afternoon and removed the following morning (14-hour exposure). Complementarily, juveniles were caught in marginal areas with seining nets (0.6 m2, 0.5 cm mesh) and sieve (0.4 m2, 0.5 cm mesh), over one hour by five people.
Fish specimens were anesthetized and euthanized by immersion in a water solution with eugenol and confirmation of death was made by checking breathing movement. After that, fish were fixed in 10% formalin for at least 48h, transferred to 70% ethanol and identified in the laboratory (Graça & Pavanelli 2007GRAçA WJ & PAVANELLI CS. 2007. Peixes da planície de inundação do alto rio Paraná e áreas adjacentes, 1st ed., Maringá: Eduem, 241 p., Ota et al. 2018OTA RR, DEPRá GC, GRAçA WJ & PAVANELLI CS. 2018. Peixes da planície de inundação do alto rio Paraná e áreas adjacentes: revised, annotated and updated. Neotrop Ichthyol 16: e170094.).
Biometric data, such as standard length (cm) and total weight (g) of the specimens were measured. Subsequently, the fish specimens were dissected to collect their stomachs and were macroscopically analyzed to identify their stage of maturation and sex (Vazzoler 1996VAZZOLER AEM. 1996. Biologia da reprodução de peixes teleósteos: teoria e prática. Maringá: Eduem, 169 p.). The stomachs collected were fixed in 10% formaldehyde, and were transferred to 70% alcohol after thirty days. The stomach fullness was visually classified on a scale of 0 (empty), 1 (< 25% content), 2 (> 25 < 50%), 3 (> 50 < 75%) and 4 (> 75%) (Walsh & Rankine 1979WALSH M & RANKINE P. 1979. Observations on the diet of mackerel in the North Sea and the west of Britain. In: International Council for the Exploration of the Sea, p. 1-45.). Voucher specimens were deposited in the fish collection at the Laboratory of Fish Biology and Genetics (LBP), at the Institute of Biosciences, UNESP, Botucatu, São Paulo, Brazil (LBP 9169).
The stomach content (higher than 50%) was examined using stereoscopic and optical microscopes. The food items were identified to the lowest possible taxonomic level and their weight was quantified with an analytical scale (precision 0.0001 g) (Hyslop 1980HYSLOP EJ. 1980. Stomach analysis: a review of methods and their application. J Fish Biol 17: 411-429.). To calculate the relative weight of algae and detritus we first weighted the total stomach content and calculated items as a percentage from the total.
Data analysis
To test for populational differences among sites, we investigated the length-weight relationship, histogram of size classes, frequencies of juveniles and adults, frequency of feeding activity and diet composition. The descriptive statistics obtained for standard length (SL) were median and quartiles (25 and 75%). The median SL values were statistically tested with the non-parametric test Kruskal-Wallis (followed by Dunn multiple comparison test) among sampled stations, as the data presented a non-normal distribution.
The length-weight relationship was determined using the linear regression: log TW = log a + b log TL, where TW is the total weight (in grams), TL is the total length (in cm), “a” is the intercept and “b” is the slope of the linear regression (Nobile et al. 2015NOBILE AB, BRAMBILLA EM, DE LIMA FP, FREITAS-SOUZA D, BAYONA-PEREZ IL & CARVALHO ED. 2015. Length-weight relationship of 37 fish species from the Taquari River (Paranapanema Basin, Brazil). J App Ichthyol 31: 580-582.).
The histogram of length class was plotted for the different (lotic and lentic habitats). The length classes were determined using the Sturges method (Sturges 1926STURGES HA. 1926. The choice of a class interval. J Am Stat Assoc 21: 65-66.). The analysis of the adult and young proportion for the different sampling stations was established following standard length and stage of gonadal development (Vazzoler 1996VAZZOLER AEM. 1996. Biologia da reprodução de peixes teleósteos: teoria e prática. Maringá: Eduem, 169 p.). The “chi-square” non-parametric test (χ2) was applied in absolute frequency and represented in relative frequency of young and adults.
A covariance analysis (ANCOVA) was applied to test whether the regression lines (weight-length relationship) differed between the lentic and lotic habitat. Two null hypotheses were tested: (i) the slopes of the regression lines (b) are parallel to each other; (ii) the Y intercepts of regression lines (a) are all equal (McDonald 2014MCDONALD JH. 2014. Handbook of Biological Statistics (3rd ed.). Sparky House Publishing, Baltimore, Maryland. Available at: http://www.biostathandbook.com/ancova.html.
http://www.biostathandbook.com/ancova.ht...
).
The differences in dietary compositions between lentic and lotic habitats were tested using permutational multivariate analysis of variance (PERMANOVA, Anderson et al. 2008ANDERSON MJ, GORLEY RN & CLARKE KR. 2008. PERMANOVA+ for PRIMER. Guide to Software and Statistical Methods. PRIMER-E, Ltd., Plymouth.). The analysis was applied to a matrix of food items (individuals in rows and food items in columns). The Mahalanobis distance was selected as a measure of dissimilarity, because it corrects some of the limitations of the Euclidean distance (Linden 2009LINDEN R. 2009. Técnicas de agrupamento. Revista de Sistemas de Informação da FSMA 4: 18-36.). We used 999 permutations to test the significance of the F statistic derived from PERMANOVA. Statistical analyses were conducted in the R Programming Environment and the Vegan package (The R Project for Statistical computing, http://www.r-project.org; Oksanen et al. 2017OKSANEN J ET AL. 2017. Vegan: Community Ecology Package. R package Version 2.4-3. https://CRAN.R-project.org/package=vegan.
https://CRAN.R-project.org/package=vegan...
).
The frequency of stomach fullness was calculated, and the Multi-Response Permutation Procedure (MRPP) (McCune & Grace 2002MCCUNE B & GRACE JB. 2002. Analysis of ecological communities. Oregon, Mjm Software Design, 300 p.) was used to test the significance of differences between the lotic and lentic habitats. MRPP provides a test statistic (T) that describes the separation between the groups (the more negative the T value, the stronger the separation), a measure of “effect size” (A) which is a measure of the degree of group homogeneity, compared to random expectation (where Amax = 1 when all items are identical within groups), and a p-value, which is useful for evaluating the likelihood that an observed difference is due to chance (McCune & Grace 2002MCCUNE B & GRACE JB. 2002. Analysis of ecological communities. Oregon, Mjm Software Design, 300 p.). As the input distance matrix, we selected the Bray-Curtis distance, with “n/sum(n)” as the weighting factor.
The trophic niche breadth was calculated with corrected Levin’s index (B) (Hurlbert 1978HURLBERT SH. 1978. The measurement of niche overlap and some relatives. Ecol 59: 67-77.). The B value ranges from 0 (species feed on few food resources) to n (species feed on all resources in similar proportions). The statistical analyses were performed in Statistica 7.0 (Statsoft 2004STATSOFT INC. 2004. Statistica (data analysis software system), version 7.0. Tulsa. Available at: http://www.statsoft.com.
http://www.statsoft.com...
) and PC-Ord v. 5.0 (McCune & Mefford 2011MCCUNE B & MEFFORD MJ. 2011. PC-ORD: Multivariate Analysis of Ecological Data. Version 6. Mjm Software, Gleneden Beach, Oregon.). For all analyses the level of significance was set to 5%.
RESULTS
We collected 730 specimens of A. lacustris. A higher number of individuals were caught in all the lentic sampling sites (Lotic I=53; Lotic II=47; Lotic III=61; Lentic I=339; Lentic II=112; Lentic III=118).
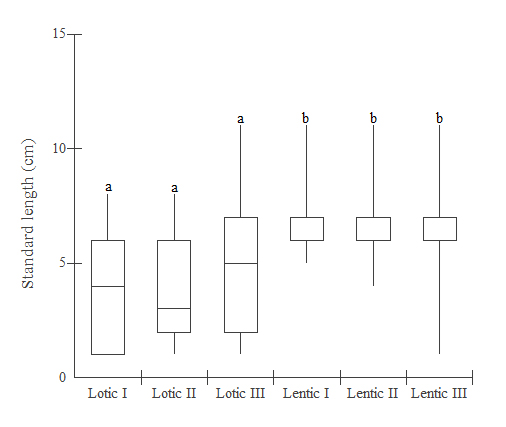
The lotic sample sites showed a normal standard-length distribution (Kolmogorov-Smirnov, p > 0.05), while the lentic sample sites showed non-normal distribution (Kolmogorov-Smirnov, p < 0.001). Kruskal-Wallis test showed statistically significant differences for this variable (H = 117.51, GL = 5, p < 0.0001). The highest median value was obtained for Lentic I (7.0 cm) (Dunn’s test p < 0.05) and the lowest median value in Lotic II (3.6 cm) (Dunn’s test p < 0.05). Dunn’s multiple comparisons test showed statistically significant differences between lotic and lentic sites (Dunn’s test, p < 0.001) but not within the same habitats (p > 0.05) (Fig. 2).

Median and quartiles of standard length (cm) of Astyanax lacustris from the lentic and lotic sites of Chavantes Reservoir, Paranapanema River. Different letters indicate the statistical difference (Kruskal-Wallis, p < 0.001).
The sampling sites of lotic habitat showed broader frequency distribution of length classes than the lentic habitat. Lotic II presented two higher frequencies in the size class (1.1–2.0 cm and 6.1–7.0 cm), Lotic I showed peaks in size classes 2.1–3 cm and 6.1–7.0 cm and Lotic III had higher frequency of individuals from 1.1–2.0 and 8.1–9.0 cm (Fig. 3a). The lentic habitat had a high frequency of individuals in only a few size classes, with 86.5% of individuals between the size classes of 5.1– 6.0 cm and 7.1 – 8.0 cm (Fig. 3b).

Histograms (size classes of standard length) of Astyanax lacustris individuals in the lotic (a) and lentic habitats (b) of Chavantes Reservoir, Middle Paranapanema River.
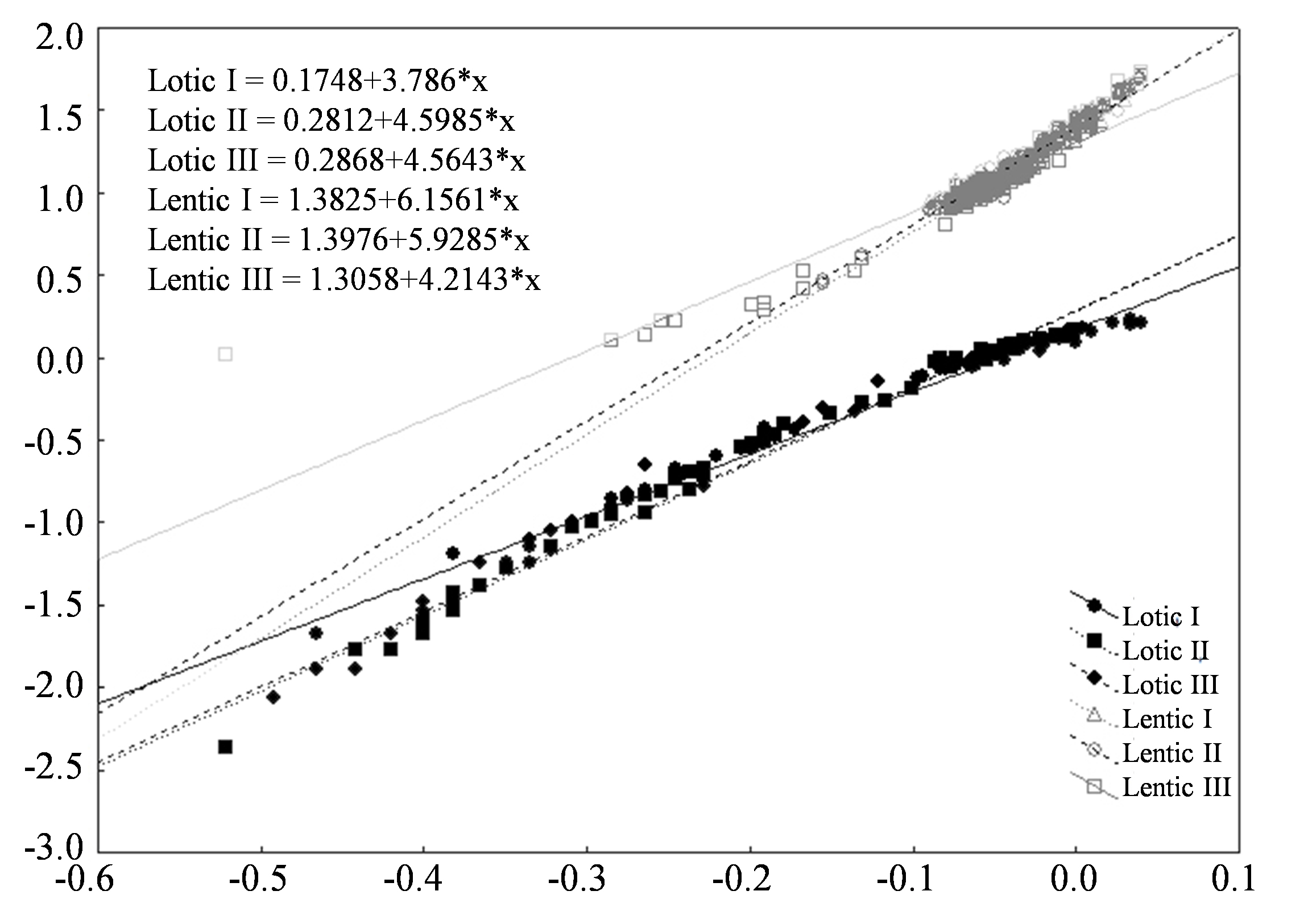
The analysis of the length-weight relationship showed that the assumption of parallelism was not achieved. The null hypothesis of all slope “b” being equal was then rejected (interaction environment * Ls significant, F=135,23, p <0.001). The fish of the lentic areas were heavier (indicated by “a”) then fish of lotic areas of the same size (Fig. 4).

Slope and equations for data of length and weight of fishes in lotic and lentic habitats of Chavantes Reservoir, Middle Paranapanema River (see Table I for more details).
Regarding the frequency of adults and young specimens, there were higher proportions of juveniles in lotic environments (Figure 5). The highest proportion of young individuals was obtained in Lotic III (59.3%) and the highest frequency of adults was obtained in Lentic I (88.8%). All lentic environments showed statistically significant differences between adult and young proportions when using the chi-square test (χ2 Lentic I=96.46; Lentic II=10.33; Lentic III=37.23).

Adult and young proportion of Astyanax lacustris in lotic and lentic habitats of Chavantes Reservoir, Middle Paranapanema River. * = statistically significant differences based on the chi-square test.
The diet of A. lacustris was composed of 37 food items (from plant material aquatic and terrestrial insects, crustaceans, mollusks, detritus, and sediment). Based on the percentage volume of the diet, plant material was the most consumed item in Lotic I, II, and III, complemented by Hymenoptera (at all Lotic sites) and Isoptera (Lotic I), Cladocera (Lotic II) and Algae (Lotic III). In the lentic sites, A. lacustris feed on Coleoptera (adults) and plant material (Lentic I), Hymenoptera (adults) and Decapoda (Lentic II) and Ephemeroptera (nymphs) and plant material (Lentic III) (Table II). Of the 21 comparisons performed between the six sites, nine showed p-values < 0.05 for Permanova: Lentic II vs all lotic sites, lotic I vs lotic II and III, lentic I vs lotic I and III. The diet of fishes of the lentic sites presented no statistical differences (Table III). The niche breadth (B) varied from 2.6 (Lotic II) to 5.6 (Lentic I).
Variation in standard length range and number of individuals (N) selected for diet analysis; the number of items found in the diet, trophic niche breadth (B), and diet composition (% weight) of the food resources consumed by Astyanax lacustris in lotic and lentic sites influenced by the Chavantes Reservoir, Paranapanema River.
Permanova results comparing Astyanax lacustris diet from lotic and lentic sites of Chavantes Reservoir, Middle Paranapanema River. p-values <0.05 are shown in bold.
Lentic I, II and III had a higher frequency of stomachs fulness with repletion levels of 0 and 1, while Lotic I, II, and III had a higher frequency with repletion levels of 2, 3, and 4 (partially filled and full). The three lotic sites differed significantly from the three lentic sites (MRPP, T = -2.71, A = 0.21, p = 0.02).
DISCUSSION
The wider amplitude of length and distribution of adult and juvenile fishes at lotic sites indicate that fish are well adapted to these habitats. This pattern was also observed in A. altiparanae from another stretch of the Paranapanema River (Orsi et al. 2004ORSI ML, CARVALHO ED & FORESTI F. 2004. Biologia populacional de Astyanax altiparanae Garutti & Britski (Teleostei, Characidae) do médio rio Paranapanema, Paraná, Brasil. Rev Bras Zool 21: 207-218.), which was associated with the quality of food resources in environments with low influence of damming. The school of juveniles that generally form groups of 30 to 50 individuals (Casatti 2002CASATTI L. 2002. Alimentação dos peixes em um riacho do Parque Estadual Morro do Diabo, bacia do Alto Paraná, sudeste do Brasil. Biota Neotrop 2: 1-14., Suzuki & Orsi 2008SUZUKI FM & ORSI ML. 2008. Formação de cardumes por Astyanax altiparanae (Teleostei, Characidae) no rio Congonhas, Paraná, Brasil. Rev Bras Zool 25: 566-569.) may encounter conditions to survive and develop at the lotic habitats. Fast-water habitats have unique ecological advantages including lower predation pressure and increased foraging efficiency (Werner et al. 1983WERNER EE, GILLIAM JF, HALL DJ & MITTELBACH GG. 1983. An experimental test of the effects of predation risk on habitat use in fish. Ecol 64: 1540-1548., Lujan & Conway 2015LUJAN NK & CONWAY KW. 2015. Life in the fast lane: a review of rheophily in freshwater fishes. In: Riesch R, Tobler M & Plath M (Eds), Extremophile Fishes, Springer, Cham, p. 107-136.) due to input of allochthonous material (Orsi et al. 2004ORSI ML, CARVALHO ED & FORESTI F. 2004. Biologia populacional de Astyanax altiparanae Garutti & Britski (Teleostei, Characidae) do médio rio Paranapanema, Paraná, Brasil. Rev Bras Zool 21: 207-218.). Marginal lagoons, present in Lotic I and II, can also enhance the shelter, survival, and recruitment of young fishes (Agostinho & Zalewski 1995AGOSTINHO AA & ZALEWSKI M. 1995. The dependence of fish community structure and dynamics on floodplain and riparian ecotone in Paraná River, Brazil. Hydrobiol 303: 141-148.).
Habitat selection by fish is related to foraging profitability and predation risk (McIvor & Odum 1988MCIVOR CC & ODUM WE. 1988. Food, predation risk and micro-habitat selection in a marsh fish assemblage. Ecol 69: 1341-1353.), which can be exerted either by native or non-native species. In the Chavantes Reservoir, two of the most noteworthy predator fish species are non-native and widespread in lentic environments (Cichla kelberi and Plagioscion squamosissimus) (Vidotto-Magnoni 2009VIDOTTO-MAGNONI AP. 2009. Ecologia trófica das assembléias de peixes do reservatório de Chavantes (Médio rio Paranapanema, SP/PR). [PhD Thesis]. Botucatu, SP: UNESP.) and in the Tibagi River, A. lacustris is main prey of P. squamosissimus (Bennemann et al. 2000BENNEMANN ST, SHIBATTA OA & GARAVELLO JC. 2000. Peixes do rio Tibagi: uma abordagem ecológica, 1st ed., Londrina: Eduel, 62 p.).
Lentic habitats presented higher abundance of individuals and fish heavier than the lotic ones. Similar results were observed by Abelha & Goulart (2008)ABELHA MCF & GOULART E. 2008. Population structure, condition factor and reproductive period of Astyanax paranae (Eigenmann, 1914) (Osteichthyes: Characidae) in a small and old Brazilian reservoir. Braz Arch Biol Technol 51: 503-512. with A. paranae from Alagados Reservoir, in state of Paraná and by Orsi et al. (2004)ORSI ML, CARVALHO ED & FORESTI F. 2004. Biologia populacional de Astyanax altiparanae Garutti & Britski (Teleostei, Characidae) do médio rio Paranapanema, Paraná, Brasil. Rev Bras Zool 21: 207-218. for A. lacustris in lentic areas of Tibagi River near to Capivara Reservoir. The marginal areas of lentic habitats are often occupied by species pre-adapted to lacustrine condition (Agostinho et al. 2016AGOSTINHO AA, GOMES LC, SANTOS NCL, ORTEGA JCG & PELICICE FM. 2016. Fish assemblages in Neotropical reservoirs: Colonization patterns, impacts and management. Fish Res 173: 26-36.). However, a small feeding activity was observed in specimens collected from the lentic sites, indicating that damming could be affecting the offer of preferential food resources. Pereira et al. (2016)PEREIRA LS, AGOSTINHO AA & DELARIVA RL. 2016. Effects of river damming in Neotropical piscivorous and omnivorous fish: feeding, body condition and abundances. Neotrop Ichthyol 14: 1-12. attributed the reduction in feeding activity of A. lacustris to the piscivory displayed by the species after the damming of Salto Caxias, Iguaçu River. Carnivorous fish usually have higher rate of empty stomachs, which can be associated to the faster digestion (Gerking 1994GERKING SD. 1994. Feeding ecology of fish, San Diego: Academic Press, 416 p.), as observed in lentic areas.
The feeding behavior of species from genus Astyanax corresponds to what Gerking (1994)GERKING SD. 1994. Feeding ecology of fish, San Diego: Academic Press, 416 p. describes as switching from one food source to a more advantageous source, which occurs in opportunistic species over a given period of time, both from the bottom (Abelha et al. 2006ABELHA MCF, GOULART E, KASHIWAQUI EAL & SILVA MRD. 2006. Astyanax paranae Eigenmann, 1914 (Characiformes: Characidae) in the Alagados Reservoir, Paraná, Brazil: diet composition and variation. Neotrop Ichthyol 4: 349-356.) and the surface of the water (Cassemiro et al. 2002CASSEMIRO FAZ, HAHN NS & FUGI R. 2002. Avaliação da dieta de Astyanax altiparanae Garutti & Britski, 2000 (Osteichthyes, Tetragonopterinae) antes e após a formação do reservatório de Salto Caxias, Estado do Paraná, Brasil. Acta Sci Biol Sci 24: 419-425.). This opportunistic behavior was observed by Lobón-Cerviá & Bennemann (2000)LOBóN-CERVIá J & BENNEMANN ST. 2000. Temporal trophic shifts and feeding in two sympatric, neotropical, omnivorous fishes: Astyanax bimaculatus and Pimelodus maculatus in Rio Tibagi (Paraná, Southern Brazil). Archiv für Hydrobiol 149: 285-306. in Astyanax altiparanae in Tibagi River and by Castro & Carvalho (2014)CASTRO RJ & CARVALHO ED. 2014. Trophic ecology of the fish in the Rio dos Veados (Upper Paranapanema River, SP) during the dry and rainy seasons. Biodiversidade 13: 1-23. in Jurimirim Reservoir (Paranapanema River). The differences in the diet of A. lacustris among the lotic and lentic sites and the trophic niche breadth indicates that these species exhibit an opportunistic behavior. Plant material, the primary food resource in Lotic sites, is a preferential food item of A. lacustris in the large rivers (Bennemann et al. 2005BENNEMANN ST, GEALH ST, ORSI ML & SOUSA LM. 2005. Ocorrência e estrutura trófica de quatro espécies de Astyanax (Characidade) em diferentes rios da bacia do rio Tibagi, Paraná, Brasil. Iheringia Sér Zool 95: 247-254.) and reservoirs (Lima et al. 2018LIMA FP, NOBILE AB, FREITAS-SOUZA D, CARVALHO ED & VIDOTTO-MAGNONI AP. 2018. Can dams affect the trophic structure of ichthyofauna? A long-term effects in the Neotropical region. Iheringia Série Zool 108: e2018030.), which is also a common feeding resource for other species of the genus, such as A. taeniatus (Manna et al. 2012MANNA LR, REZENDE CF & MAZZONI R. 2012. Plasticity in the diet of Astyanax taeniatus in a coastal stream from south-east Brazil. Braz J Biol 72: 919-928.). An herbivorous feeding habit is an adaptation to the use of vegetation on river margins, reservoirs, and cultivated pastures and was observed for this species in Iguaçu River before the closure of Salto Caxias Dam (Pereira et al. 2016PEREIRA LS, AGOSTINHO AA & DELARIVA RL. 2016. Effects of river damming in Neotropical piscivorous and omnivorous fish: feeding, body condition and abundances. Neotrop Ichthyol 14: 1-12.). Terrestrial insects (Hymenoptera and Coleoptera) were the main food resources of A. lacustris in Lentic I and II and are well documented for this species in rivers and reservoirs (Casatti 2002CASATTI L. 2002. Alimentação dos peixes em um riacho do Parque Estadual Morro do Diabo, bacia do Alto Paraná, sudeste do Brasil. Biota Neotrop 2: 1-14., Bennemann et al. 2005BENNEMANN ST, GEALH ST, ORSI ML & SOUSA LM. 2005. Ocorrência e estrutura trófica de quatro espécies de Astyanax (Characidade) em diferentes rios da bacia do rio Tibagi, Paraná, Brasil. Iheringia Sér Zool 95: 247-254., Bonato et al. 2012BONATO KO, DELARIVA RL & SILVA JC. 2012. Diet and trophic guilds of fish assemblage in two streams with different anthropic impacts in the Northwest of Paraná, Brazil. Rev Bras Zool 29: 27-38., Delariva et al. 2013DELARIVA RL, HAHN NS & KASHIWAQUI EAL. 2013. Diet and trophic structure of the fish fauna in a subtropical ecosystem: impoundment effects. Neotrop Ichthyol 11: 891-904.). The differences observed between lotic sites (Paranapanema, Itararé and Verde rivers) can be inferred by the differences in conditions (size, width, depth) that may interfere in the composition and availability of different food items.
Species that persist many years after reservoir formation may have the behavioral personality of boldness (a trait where individual are prone to take risks and explore the environment faster when exposed to novelty). Boldness has implications for survival, reproduction, and many other behavioral and life-history traits (Budaev 1997BUDAEV SV. 1997. ‘Personality’ in the guppy (Poecilia reticulata): A correlated study of exploratory behavior and social tendency. J Comp Psychol 111: 399-411., Binoy 2015BINOY V. 2015. Comparative analysis of boldness in four species of freshwater teleosts. Indian J Fish 62: 128-130., Ariyomo & Watt 2015ARIYOMO TO & WATT PJ. 2015. Effect of hunger level and time of day on boldness and aggression in the zebrafish Danio rerio. J Fish Biol 86: 1852-1859.), especially for those like A. lacustris that form schools (Suzuki & Orsi 2008SUZUKI FM & ORSI ML. 2008. Formação de cardumes por Astyanax altiparanae (Teleostei, Characidae) no rio Congonhas, Paraná, Brasil. Rev Bras Zool 25: 566-569.). A variation in the boldness spectrum may influence the risk of predation, diet, and consequently growth and fitness of the species (Wilson et al. 1994WILSON DS, CLARK AB, COLEMAN K & DEARSTYNE T. 1994. Shyness and boldness in humans and other animals. Trends Ecol Evol 9: 442-446.). Understanding the boldness spectrum of species should be subject of future research.
Our hypothesis was partially corroborated, since A. lacustris encountered superior conditions for recruitment in lotic habitats, adjacent to reservoir. Moreover, this species showed an ability to adapt to the impoundments using different food resources at the lentic sites and colonizing littoral areas.
REFERENCES
- ABELHA MCF & GOULART E. 2008. Population structure, condition factor and reproductive period of Astyanax paranae (Eigenmann, 1914) (Osteichthyes: Characidae) in a small and old Brazilian reservoir. Braz Arch Biol Technol 51: 503-512.
- ABELHA MCF, GOULART E, KASHIWAQUI EAL & SILVA MRD. 2006. Astyanax paranae Eigenmann, 1914 (Characiformes: Characidae) in the Alagados Reservoir, Paraná, Brazil: diet composition and variation. Neotrop Ichthyol 4: 349-356.
- AGOSTINHO AA, GOMES LC & PELICICE FM. 2007. Ecologia e manejo de recursos pesqueiros em reservatórios do Brasil, 1st ed., Maringá: Eduem, 501 p.
- AGOSTINHO AA, GOMES LC, SANTOS NCL, ORTEGA JCG & PELICICE FM. 2016. Fish assemblages in Neotropical reservoirs: Colonization patterns, impacts and management. Fish Res 173: 26-36.
- AGOSTINHO AA, MIRANDA LE, BINI LM, GOMES LC, THOMAZ SM & SUZUKI HI. 1999. Patterns of colonization in neotropical reservoirs, and prognoses on aging. In: Tundisi JG & Straskraba M (Orgs), Theoretical reservoir ecology and its applications: Instituto Internacional de Ecologia, São Carlos, p. 227-265.
- AGOSTINHO AA & ZALEWSKI M. 1995. The dependence of fish community structure and dynamics on floodplain and riparian ecotone in Paraná River, Brazil. Hydrobiol 303: 141-148.
- ANA - AGENCIA NACIONAL DE ÁGUAS. 2016. Plano Integrado de Recursos Hídricos da Unidade de Gestão de Recursos Hídricos Paranapanema. Agência Nacional Das Águas – Brasília, 354 p.
- ANDERSON MJ, GORLEY RN & CLARKE KR. 2008. PERMANOVA+ for PRIMER. Guide to Software and Statistical Methods. PRIMER-E, Ltd., Plymouth.
- ANEEL - AGÊNCIA NACIONAL DE ENERGIA ELÉTRICA. 2019. Acompanhamento de estudos e projetos hidroenergéticos. Available at: http://www2.aneel.gov.br/aplicacoes/capacidadebrasil/UsinaListaSelecao.asp
» http://www2.aneel.gov.br/aplicacoes/capacidadebrasil/UsinaListaSelecao.asp - ARIYOMO TO & WATT PJ. 2015. Effect of hunger level and time of day on boldness and aggression in the zebrafish Danio rerio. J Fish Biol 86: 1852-1859.
- BENNEMANN ST, GEALH ST, ORSI ML & SOUSA LM. 2005. Ocorrência e estrutura trófica de quatro espécies de Astyanax (Characidade) em diferentes rios da bacia do rio Tibagi, Paraná, Brasil. Iheringia Sér Zool 95: 247-254.
- BENNEMANN ST, SHIBATTA OA & GARAVELLO JC. 2000. Peixes do rio Tibagi: uma abordagem ecológica, 1st ed., Londrina: Eduel, 62 p.
- BINOY V. 2015. Comparative analysis of boldness in four species of freshwater teleosts. Indian J Fish 62: 128-130.
- BONATO KO, DELARIVA RL & SILVA JC. 2012. Diet and trophic guilds of fish assemblage in two streams with different anthropic impacts in the Northwest of Paraná, Brazil. Rev Bras Zool 29: 27-38.
- BUDAEV SV. 1997. ‘Personality’ in the guppy (Poecilia reticulata): A correlated study of exploratory behavior and social tendency. J Comp Psychol 111: 399-411.
- CANTANHêDE G, HAHN NS, FUGI R & GUBIANI ÉA. 2008. Alterations on piscivorous diet following change in abundance of prey after impoundment in a Neotropical river. Neotrop Ichthyol 6: 631-636.
- CASATTI L. 2002. Alimentação dos peixes em um riacho do Parque Estadual Morro do Diabo, bacia do Alto Paraná, sudeste do Brasil. Biota Neotrop 2: 1-14.
- CASATTI L, MENDES HF & FERREIRA KM. 2003. Aquatic macrophytes as feeding site for small fishes in the Rosana Reservoir, Paranapanema River, Southeastern Brazil. Braz J Biol 63: 213-222.
- CASSEMIRO FAZ, HAHN NS & FUGI R. 2002. Avaliação da dieta de Astyanax altiparanae Garutti & Britski, 2000 (Osteichthyes, Tetragonopterinae) antes e após a formação do reservatório de Salto Caxias, Estado do Paraná, Brasil. Acta Sci Biol Sci 24: 419-425.
- CASTRO RJ & CARVALHO ED. 2014. Trophic ecology of the fish in the Rio dos Veados (Upper Paranapanema River, SP) during the dry and rainy seasons. Biodiversidade 13: 1-23.
- DEI TOS C, BARBIERI G, AGOSTINHO AA, GOMES LC & SUZUKI HI. 2002. Ecology of Pimelodus maculatus (Siluriformes) in the Corumbá reservoir, Brazil. Cybium 26: 275-282.
- DELARIVA RL, HAHN NS & KASHIWAQUI EAL. 2013. Diet and trophic structure of the fish fauna in a subtropical ecosystem: impoundment effects. Neotrop Ichthyol 11: 891-904.
- DIAS RM, BAILLY D, ANTôNIO RR, SUZUKI HI & AGOSTINHO AA. 2005. Colonization of the Corumbá Reservoir (Corumbá River, Paraná River basin, Goiás state, Brazil) by “lambari” Astyanax altiparanae (Tetragonopterinae; Characidae). Braz Arch Biol Technol 48: 467-476.
- GERKING SD. 1994. Feeding ecology of fish, San Diego: Academic Press, 416 p.
- GRAçA WJ & PAVANELLI CS. 2007. Peixes da planície de inundação do alto rio Paraná e áreas adjacentes, 1st ed., Maringá: Eduem, 241 p.
- GOMES LC & MIRANDA LE. 2001. Riverine characteristics dictate composition of fish assemblages and limit fisheries in reservoirs of the Upper Paraná River Basin. Regulated Rivers: Research & Management: An International Journal Devoted to River Research and Management 17: 67-76.
- GRILL G, LEHNER B, LUMSDON AE, MACDONALD GK, ZARFL C & LIERMANN CR. 2015. An index-based framework for assessing patterns and trends in river fragmentation and flow regulation by global dams at multiple scales. Environ Res Lett 10: 015001.
- HAHN NS & FUGI R. 2008. Environmental changes, habitat modifications and feeding ecology of freshwater fish. In: Cyrino JEP, Bureau DP & Kapoor BG (Eds), Feeding and Digestive Functions in Fishes: Science Publishers, p. 35-65.
- HURLBERT SH. 1978. The measurement of niche overlap and some relatives. Ecol 59: 67-77.
- HYSLOP EJ. 1980. Stomach analysis: a review of methods and their application. J Fish Biol 17: 411-429.
- ICOLD - INTERNATIONAL COMMISSION ON LARGE DAMS. 2018. World register on dams – General synthesis. Available at: http://www.icold-cigb.org/GB/World_register/general_synthesis.asp
» http://www.icold-cigb.org/GB/World_register/general_synthesis.asp - KRAFT NJ, ADLER PB, GODOY O, JAMES EC, FULLER S & LEVINE JM. 2015. Community assembly, coexistence and the environmental filtering metaphor. Funct Ecol 29: 592-599.
- LANGEANI F, CASTRO RMC, OYAKAWA OT, SHIBATTA OA, PAVANELLI CS & CASATTI L. 2007. Diversidade da ictiofauna do Alto Rio Paraná: composição atual e perspectivas futuras. Biota Neotrop 7: 181-197.
- LIMA FP, NOBILE AB, FREITAS-SOUZA D, CARVALHO ED & VIDOTTO-MAGNONI AP. 2018. Can dams affect the trophic structure of ichthyofauna? A long-term effects in the Neotropical region. Iheringia Série Zool 108: e2018030.
- LINDEN R. 2009. Técnicas de agrupamento. Revista de Sistemas de Informação da FSMA 4: 18-36.
- LOBóN-CERVIá J & BENNEMANN ST. 2000. Temporal trophic shifts and feeding in two sympatric, neotropical, omnivorous fishes: Astyanax bimaculatus and Pimelodus maculatus in Rio Tibagi (Paraná, Southern Brazil). Archiv für Hydrobiol 149: 285-306.
- LOUREIRO-CRIPPA VE & HAHN NS. 2006. Use of food resources by the fish fauna of a small reservoir (rio Jordão, Brazil) before and shortly after its filling. Neotrop Ichthyol 4: 357-362.
- LOWE-MCCONNELL RH. 1999. Estudos ecológicos de comunidades de peixes tropicais, São Paulo: Edusp, 534 p.
- LUCENA CD & SOARES HG. 2016. Review of species of the Astyanax bimaculatus “caudal peduncle spot” subgroup sensu Garutti & Langeani (Characiformes, Characidae) from the rio La Plata and rio São Francisco drainages and coastal systems of southern Brazil and Uruguay. Zootaxa 4072: 101-125.
- LUJAN NK & CONWAY KW. 2015. Life in the fast lane: a review of rheophily in freshwater fishes. In: Riesch R, Tobler M & Plath M (Eds), Extremophile Fishes, Springer, Cham, p. 107-136.
- MANNA LR, REZENDE CF & MAZZONI R. 2012. Plasticity in the diet of Astyanax taeniatus in a coastal stream from south-east Brazil. Braz J Biol 72: 919-928.
- MARONEZE DM, TUPINAMBáS TH, ALVEZ CBM, VIEIRA F, POMPEU PS & CALLISTO M. 2011. Fish as ecological tools to complement biodiversity inventories of benthic macroinvertebrates. Hydrobiol 673: 29-40.
- MCCUNE B & GRACE JB. 2002. Analysis of ecological communities. Oregon, Mjm Software Design, 300 p.
- MCCUNE B & MEFFORD MJ. 2011. PC-ORD: Multivariate Analysis of Ecological Data. Version 6. Mjm Software, Gleneden Beach, Oregon.
- MCDONALD JH. 2014. Handbook of Biological Statistics (3rd ed.). Sparky House Publishing, Baltimore, Maryland. Available at: http://www.biostathandbook.com/ancova.html
» http://www.biostathandbook.com/ancova.html - MCIVOR CC & ODUM WE. 1988. Food, predation risk and micro-habitat selection in a marsh fish assemblage. Ecol 69: 1341-1353.
- MIRANDA LE & KROGMAN RM. 2015. Functional age as an indicator of reservoir senescence. Fish 40: 170-176.
- MOL JH, MÉRONA BD, OUBOTER PE & SAHDEW S. 2007. The fish fauna of Brokopondo Reservoir, Suriname, during 40 years of impoundment. Neotrop Ichthyol 5: 351-368.
- NOBILE AB, BRAMBILLA EM, DE LIMA FP, FREITAS-SOUZA D, BAYONA-PEREZ IL & CARVALHO ED. 2015. Length-weight relationship of 37 fish species from the Taquari River (Paranapanema Basin, Brazil). J App Ichthyol 31: 580-582.
- NOGUEIRA MG, JORCIN A, VIANNA NC & BRITTO YC. 2006. Reservatórios em cascata e os efeitos na limnologia e organização das comunidades bióticas (fitoplâncton, zooplâncton e zoobentos), um estudo de caso no rio Paranapanema (SP/PR). In: Nogueira MG, Henry R & Jorcin A (Eds), Ecologia de reservatórios: Impactos potenciais, ação de manejo e sistemas em cascatas: Rima, São Carlos, p. 435-459.
- OKSANEN J ET AL. 2017. Vegan: Community Ecology Package. R package Version 2.4-3. https://CRAN.R-project.org/package=vegan
» https://CRAN.R-project.org/package=vegan - ORSI ML. 2010. Estratégias reprodutivas de peixes da região média-baixa do Rio Paranapanema, reservatório de Capivara. São Paulo: Blucher, 113 p.
- ORSI ML, CARVALHO ED & FORESTI F. 2004. Biologia populacional de Astyanax altiparanae Garutti & Britski (Teleostei, Characidae) do médio rio Paranapanema, Paraná, Brasil. Rev Bras Zool 21: 207-218.
- OTA RR, DEPRá GC, GRAçA WJ & PAVANELLI CS. 2018. Peixes da planície de inundação do alto rio Paraná e áreas adjacentes: revised, annotated and updated. Neotrop Ichthyol 16: e170094.
- PELICICE FM & AGOSTINHO AA. 2006. Feeding ecology of fishes associated with Egeria spp. patches in a tropical reservoir, Brazil. Ecol Fresh Fish 15: 10-19.
- PELICICE FM, POMPEU PS & AGOSTINHO AA. 2015. Large reservoirs as ecological barriers to downstream movements of Neotropical migratory fish. Fish Fish 16: 697-715.
- PERBICHE-NEVES G & NOGUEIRA MG. 2010. Multi-dimensional, effects on Cladoceran (Crustacea, Anomopoda) assemblages in two cascade reservoirs in Southeast Brazil. Lakes Reservoirs: Res Manage 15: 139-152.
- PEREIRA LS, AGOSTINHO AA & DELARIVA RL. 2016. Effects of river damming in Neotropical piscivorous and omnivorous fish: feeding, body condition and abundances. Neotrop Ichthyol 14: 1-12.
- POFF NL, OLDEN JD, MERRITT DM & PEPIN DM. 2007. Homogenization of regional river dynamics by dams and global biodiversity implications. Proc Natl Acad Sci 104: 5732-5737.
- SUZUKI FM & ORSI ML. 2008. Formação de cardumes por Astyanax altiparanae (Teleostei, Characidae) no rio Congonhas, Paraná, Brasil. Rev Bras Zool 25: 566-569.
- STATSOFT INC. 2004. Statistica (data analysis software system), version 7.0. Tulsa. Available at: http://www.statsoft.com
» http://www.statsoft.com - STURGES HA. 1926. The choice of a class interval. J Am Stat Assoc 21: 65-66.
- TUNDISI JG & MATSUMURA-TUNDISI T. 2003. Integration of research and management in optimizing multiple uses of reservoirs: the experience in South America and Brazilian case studies. Aquat Biodiversity 500: 231-242.
- TURGEON K, TURPIN C & GREGORY-EAVES I. 2019. Dams have varying impacts on fish communities across latitudes: A quantitative synthesis. Ecol Letters. 22: 1501-1516. doi: https://doi.org/10.1111/ele.13283.
- VASCONCELOS LP, ALVES DC & GOMES LC. 2014. Fish reproductive guilds downstream of dams. J Fish Biol 85: 1489-1506.
- VAZZOLER AEM. 1996. Biologia da reprodução de peixes teleósteos: teoria e prática. Maringá: Eduem, 169 p.
- VIDOTTO-MAGNONI AP. 2009. Ecologia trófica das assembléias de peixes do reservatório de Chavantes (Médio rio Paranapanema, SP/PR). [PhD Thesis]. Botucatu, SP: UNESP.
- WALSH M & RANKINE P. 1979. Observations on the diet of mackerel in the North Sea and the west of Britain. In: International Council for the Exploration of the Sea, p. 1-45.
- WERNER EE, GILLIAM JF, HALL DJ & MITTELBACH GG. 1983. An experimental test of the effects of predation risk on habitat use in fish. Ecol 64: 1540-1548.
- WILSON DS, CLARK AB, COLEMAN K & DEARSTYNE T. 1994. Shyness and boldness in humans and other animals. Trends Ecol Evol 9: 442-446.
Publication Dates
-
Publication in this collection
07 July 2021 -
Date of issue
2021
History
-
Received
20 May 2019 -
Accepted
25 Feb 2020