
ABSTRACT
The genus Piptadenia Benth. has undergone several changes in its circumscriptions throughout its history, given the difficulty of its phylogenetic placement. Considering that palynological contributions have been significant for differentiating taxa within the Mimoseae tribe , the present study aims to characterize the representatives of the genus as well as describe the pollen grains of some taxa still unpublished in the literature. Three types of pollen grains were identified based on the number of grains per polyad considering the number of pollen grains and their ornamentation. However, other palynological characters are similar among species, supporting the monophyly of the genus. All species showed similarities, especially regarding size and shape, varying mainly in the number of pollen grains composing the polyads (8, 12, and 16). This variation has already been confirmed by morphological and molecular data. This study presents unprecedented palynological characterization of the species: Piptadenia affinis, P. cuzcoënsis, P. gonoacantha, P. imatacae, P. killipii, P. micracantha, and P. robusta.
Keywords:
diversity; exine; genus; morphology; palynotaxonomy; Piptadenia Benth; pollens; taxonomic
INTRODUCTION
In Bentham's study (1840), the genus Piptadenia Benth. was divided into three sections: Eupiptadenia Benth., Pityrocarpa (Benth.) Britton & Rose, and Niopa (Benth.) Britton. Furthermore, Piptadenia was also included in the tribe Piptadenieae due to the presence of glands on the anthers. Nonetheless, the boundaries of the tribe were expanded to include taxa lacking endosperm in the seed (Bentham, 1875). Piptadenia sensu Bentham (1840) was distinguished from other genera of the tribe by the presence of flattened legumes and thin valves. The genus lacks specialized fruit, which is characteristic of other Mimoseae Bronn genera, such as Entada Adans. and Anadenanthera Speg. Fruits were unknown for some of the species treated by Bentham (1840), and even a consistent fruit type was not present within the Piptadenia.
Later, Brenan (1955) reorganized Piptadenia and closely related genera based mainly on fruit shape, dehiscence type, and seed characteristics. Piptadenia was restricted to species with winged seeds and without endosperm, while the genus Pityrocarpa Benth. included species with non-winged seeds, presence of endosperm, and smooth testa. Brenan's typification (1965) was not accepted, and Pityrocarpa was again included within Piptadenia, while species with winged seeds were transferred to Parapiptadenia Brenan by Cowan & Brenan (1960). Subsequently, phylogenetic analyses by Jobson & Luckow (2007) demonstrated the polyphyly of Piptadenia (sensu Brenan, 1955).
Jobson & Luckow (2007) also confirmed the polyphyly of Piptadenia, previously reported by Luckow et al. (2003). These findings indicated the genus Pityrocarpa to be the sister group of Pseudopiptadenia Rauschert, despite weak support. Additionally, a polytomy among the genera Parapiptadenia, Microlobius C.Presl, and Stryphnodendron Mart. was evidenced. The species Piptadenia viridiflora (Kunth.) appears separated from Pityrocarpa, suggesting its repositioning, possibly as a new genus. In addition to phylogenetic analyses, the pollen morphology of Piptadenia viridiflora (polyads with 8 pollen grains, acalymmate, ovoid-shaped, and disposed in two opposite tetrahedral tetrads) also showed a distinction from the group.
Polyads with eight pollen grains are recorded in some species from the extinct informal group Piptadenia such as Pityrocarpa moniliformis (Benth.) Luckow & Jobson, Microlobius foetidus (Jacq.) M.Sousa & G.Andrade, and species of Mimosa L. However, the bitetrad arrangement observed in Piptadenia viridiflora is common in Mimosa species (Lima et al., 2008), unlike the arrangement in Pityrocarpa moniliformis and M. foetidus, which exhibit pollen grains arranged in six peripheral and two central positions.
The exine ornamentation of the pollen grains in Piptadenia viridiflora was found to be distinct from other species within the Piptadenia group. The isolated position of P. viridiflora, as revealed by molecular analyses and supported by morphological evidence, led Ribeiro et al. (2018) to propose the establishment of the new genus Lachesiodendron P.G. Ribeiro, L.P. Queiroz & Luckow. Consequently, the monophyly of Piptadenia was strongly supported following the exclusion of P. viridiflora.
Although Piptadenia can be identified by a combination of macromorphological characters, its phylogenetic positioning has represented a challenge, leading to the proposal of several circumscriptions throughout its history. Palynological studies have played a significant role in differentiating taxa within the Mimoseae, a group extensively studied for its pollen morphology (Erdtman, 1952; Van Campo & Guinet, 1961; Barros, 1963; Barth & Yoneshigue, 1966; Guinet, 1969; Sorsa, 1969; Barth & Silva, 1963; Jiménez, 1996; Silvestre-Capelato & Melhem, 1997; Caccavari, 1987; 2002; Du Bocage et al., 2008; Lima et al., 2008; Santos & Romão, 2008; Buril & Santos, 2010; Santos-Silva et al., 2013; Cruz et al., 2017; Medina-Acosta et al., 2018; Duarte et al., 2021; Soares et al., 2022).
Caccavari (1987; 2002) reported that Piptadenia s.l. can be considered as a eurypalynous group. Despite the significant diversity of pollen within the Piptadenia group, morphological and structural traits remain useful for distinguishing it from other Mimoseae groups, whose species exhibit pollen grains arranged in tetrads and/or polyads (Caccavari, 2002).
This study aims to advance palynological knowledge of representatives of the genus Piptadenia, including descriptions of the pollen morphology of species that remain unpublished in the specialized literature, and discuss palynological data that could be relevant in a taxonomic context.
MATERIALS AND METHODS
Pollen samples from 21 (twenty-one) species of the genus Piptadenia were analyzed. Pollen material was collected from exsiccates deposited in the following herbaria: BMCB, CEPEC, F, HUEFS, IAN, INPA, MBM, MO, NY, RB, SP, and US (acronyms following Thiers 2023 [continuously updated]). Whenever possible, three specimens were sampled for each species. The analyzed material is cited in Appendix 1.
Pollen grains were acetolyzed following the methodology proposed by Erdtman (1960), in which the pollen material was removed from the anthers of fertile floral buds and remained in the acetolysis solution for two minutes. Permanent slides were mounted in glycerin gelatin and sealed with paraffin. For each sample, four slides were mounted in transparent gelatin and one in gelatin stained with safranin, for better observation of the characters. The grains were measured and microphotographed using a Leica ICC50 W light microscope (LM). The mounted slides were included into the pollen library of the Laboratory of Plant Micromorphology of the State University of Feira de Santana (PUEFS).
For scanning electron microscopy analysis (SEM), acetolyzed pollen grains were washed and dehydrated in an ascending hydroethanolic series (50%, 70%, 90%, and 100%), remaining about 10 minutes in each bath. The absolute alcohol containing the pollen grains was dripped directly onto the specimen holder of the scanning electron microscope. After total drying, they were metalized by evaporation of gold in a high vacuum and electron micrographed in the JEOL 6390LV microscope from the Electronic Microscopy Platform of the Oswaldo Cruz Foundation - Gonçalo Moniz Research Center.
For palynological characterization, the following morphometric characters were measured: major and minor diameter, whenever possible, in 25 randomly selected polyads. The other parameters, sexine, nexine, and exine thickness of ten grains, were also randomly selected.
The morphometric characters were statistically analyzed by calculating the arithmetic mean (x̄), the sample standard deviation (s), the standard deviation of the mean (sx̄), the coefficient of variation (CV), and the 95% confidence interval (CI) for pollen parameter measurements with a sample size of 25, For the other measurements with a sample size of ten, only the arithmetic mean was calculated.
Pollen grains were described according to their main morphological attributes: size, shape, number of grains per polyad, types of apertures, types of ornamentation, and exine ornamentation. The palynological terminology followed Punt et al. (2007) and Halbritter et al. (2018).
The specimens analyzed are listed below:
Piptadenia adiantoides (Spreng.) J.F. Macbr.- Brazil, Minas Gerais: Serra do Cipó., Santana do Riacho, Pirani, J.R. 5029 (HUEFS); Brazil, Bahia: Feira de Santana, Moraes, M.V. 471ª (HUEFS); - , Oliveira, R.P. 599 (HUEFS).
Piptadenia buchtienii Barneby - Dorr, L.J; Barnett, L.C. & Lewis, M. s/n (NYBG); Bolivia, La Paz: Nor Yungas, Beck, St. G 8727 (NYBG).
Piptadenia floribunda Kleinhoonte - Brazil, Pará: Almeirim, Pires, M.J. & Silva, N. s/n (INPA); -, Mori, S. & Bolten, A. 12 (NYBG); -, Gran Ville de Crozier, J.J. F. 13823 (NYBG).
Piptadenia gonoacantha (Mart.) J.F.Macbr. - Brazil, São Paulo: São Paulo, Rossi, L. 145 (HUEFS); -, Campos, M.B.S. 57 (HUEFS); Brazil, Minas Gerais: Juiz de Fora, Pifano, D.S. 252 (CESJ).
Piptadenia imatacae Barneby - Ecuador, Sucumbios: Putumayo, Cerón, E. C. 9794 (MO).
Piptadenia irwinii G.P.Lewis - Brazil, Bahia: Palmeiras, Queiroz, L.P. de 12652 (HUEFS); Bahia, Brazil: Licínios de Almeida, Jardim, J.G. 3303 (HUEFS); Brazil, Bahia: Morro da Torre, Jequié, Queiroz, L. P. de 12887 (HUEFS).
Piptadenia killipii J.F.Macbr.- Peru, San Martín: San Martín, Belsmaw, C. M. 3159 (F); San Martín, Peru: Alto Rio Huallaga, Klug, G. 4333 (F); Bahia, Brazil: Juçari, Belém, R.P.; Pinheiro, R.S 2345 (IAN).
Piptadenia laxipinna G.M.Barroso - Brazil, Minas Gerais: São Pedro do Suaçuí, Davidse, T.P.G. Ramamoorthy & Vital, D.M. 11491 (MO); -, Timaná, M. & Smith, P. 1465 (MO).
Piptadenia micracantha Benth. - Brazil, Minas Gerais: Tombos, Oliveira, J.E. de 327 (HUEFS); Brazil, Bahia: Igrapiúna, Reserva Ecológica Michelin, Queiroz, L.P.de 327 (HUEFS).
Piptadenia paniculata Benth. - Brazil, Minas Gerais: Juiz de Fora, Krieger L. 1093 (HUEFS); Brazil, Paraná: Morretes, Hatschbach, G. 1650 (HUEFS); Brazil, Rio de Janeiro: Guapimirim, Estação Ecológica Estadual de Paraíso, de Lima, H. C. 4357 (HUEFS).
Piptadenia peruviana (J.F.Macbr.) Barneby - Peru, Loreto: Klug, G., 2927 (NYBG); -, Schunke Vigo, J. & Grahan, J. G. 514981 (MO).
Piptadenia pteroclada Benth - Brazil, Amazonas: São Paulo de Olivença, Froes, R. L. 20805 (IAN); Ecuador, Sucumbíos: Lago Agrio Bosque Húmedo Tropical, Neill, D. 7201 (MBM); Peru, Loreto: Retinga Central, Padre Isla, Prov. Maynas, Encarnacións, F. & Merjia, K. 25137 (MBM); Peru, Loreto: Maynas, Plowman, T.; Schultes, R.E. &. Tovar, O. 18 (INPA).
Piptadenia ramosissima (Mart. ex Colla) Benth. - Brazil, Bahia: Jussari, Nascimento, F.M.F 1564 (HUEFS); Brazil, Bahia: Rui Barbosa, Serra do Orobó, Queiroz, L.P.de 9915 (HUEFS).
Piptadenia retusa (Jacq.) P.G.Ribeiro, Seigler & Ebinger - Brazil, Sergipe: Monte Alegre de Sergipe, Machado, W. J 362 (HUEFS); Brazil, Sergipe: Canindé de São Francisco, Silva, A.C.C. 289 (HUEFS); - , Aristeiguieta, L. 7470 (MO).
Piptadenia santosii Barneby ex G.P.Lewis - Brazil, Minas Gerais: Salto da Divisa, Lombardi, J.A. 5044 (BMCB); Brazil, Bahia: Prado, Santos, T.S. 2729 (CEPEC).
Piptadenia trisperma (Vell.) Benth. - Brazil, Rio de Janeiro: Rio de Janeiro, Jacarepaguá, Hoehne, F.C. 111 (SP); Brazil, Rio de Janeiro: Maricá, Lewis, G.P. & de Lima, H.C. 1191 (HUEFS).
Piptadenia uaupensis (Benth.) Spruce - Brazil, Amazonas: Tonantins, Ducke 1592 (IAN); Knowles, E. 551 (INPA); Brazil, Pará: Santarém, Silva, M. & s/n (MG).
Piptadenia uliginosa Britton & Killip - Colombia, Bolívar: Volcanes: Killip, E.P. & Albert, C. S. 14140 (US); Colombia, Campeche Atlántico: Depto. del Atlantico, Dugand, A. 5328 (US); -, Lima, J.R. 571 (RB); - , Lima, J.R. 647 (RB); Lima, J.R. 460 (RB).
RESULTS
Pollen grains in Piptadenia are represented by polyads ranging in size from 15.7-43.6 µm (small to medium) (Table 1), with 8, 12, and 16 pollen grains (Fig. 1 K ). Polyads are mostly spheroidal, with pollen grains regularly arranged (Table 2). The type of aperture was not evidenced. Individual pollen grains are spheroidal and irregularly arranged.

Polyads of Piptadenia (Leguminosae) species analyzed under Light Microscopy (LM) and Scanning Electron Microscopy (SEM). P. adiantoides - A. Ovoidal polyad, B. Rugulate ornamentation. P. affinis - C.Polyads with 8 pollen grains. P. buchtieniiD. Detail of rugulate ornamentation. P. cuzcoënsis - E. Polyads with 12 pollen grains, F. Rugulate ornamentation. P. floribunda - G. Rugulate ornamentation. P. gonoacantha - H. Areolate ornamentation. P. imatacae - I. Polyads with 16 pollen grains, J. Detail of rugulate ornamentation. P. irwinii - K. Ovoidal polyad, L. Verrucate ornamentation. Scales: 10 µm (A, C, E,I, K), 5 µm (B, F, G, H, K), 1 µm (D, J).
Exine
Commonly, the exine exhibited a thickness ranging from 0.76 μm in P. peruviana to 1.37 μm in P. floribunda, with sexine equal to nexine in general. Sexine thicker than nexine is presented in pollen grains from P. affinis, P. cuzcoensis, P. floribunda, P. peruviana, P. santosii, and P. uaupensis, while nexine thicker than sexine was registered only in P. pteroclada. The analyzed species exhibited four patterns regarding the exine ornamentation of the polyads:
a) Rugulate in P. adiantoides, P. affinis, P. buchtienii, P. cuzcoënsis, P. floribunda, P. imatacae, P. laxipinna, P. micracantha, P. pteroclada, P. robusta, P. santosii and P. uliginosa (Fig. 1 B, D, F-G, J ; Fig. 2 C, E, J ; Fig. 3 D, F, J );
b) Areolate in P. gonoacantha, P. killipii, P. peruviana, P. ramosissima and (Fig. 1 H ; Fig. 2 B, H, L );
c) Verrucate in P. irwinii, P. retusa, P. trisperma, and P. uaupensis (Fig. 1 L ; Fig. 2 L ; Fig. 3 B, H );
d) Granulate in P. paniculata (Fig.2-G).
The presence of Ubisch bodies was registered in P. affinis, P. buchtienii, P. gonoacantha, P. paniculata, P. pteroclada, and P. uliginosa. In other hand, regarding the shape of the polyads, the ovoid shape was registered in P. adiantoides, P. affinis, P. irwinii, P. killipii, P. pteroclada, P. ramosissima, P. retusa, and P. santosii, (Fig. 1 A ; Fig. 3 A, E ) while the remaining species exhibited a spheroidal shape (Fig. 2 D ; Fig. 3 G, I, K ).

Pollen grains of species of Piptadenia (Leguminosae) analyzed under Light Microscopy (LM) and Scanning Electron Microscopy (SEM). P. killipii - A. Ovoidal polyad, B. Areolate ornamentation. P. laxipinna -C. Rugulate ornamentation. P. micracantha - D. Spheroidal polyad, E. Rugulate ornamentation. P. paniculata- F. Polyad with 16 pollen grains, G. Granulate ornamentation. P. peruviana - H. Areolate ornamentation. P.pteroclada - I. Ovoidal polyad, J. Rugulate ornamentation. P. ramosissima - K. Polyad with 8 pollen grains,L. Detail of areolate ornamentation. Scales: 10 µm (A, F, I, K), 5 µm (D, G, J, L), 1 µm (B, C, E, H).
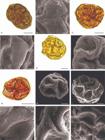
Pollen grains of species of Piptadenia (Leguminosae) analyzed under Light Microscopy (LM) and Scanning Electron Microscopy (SEM). P. retusa - A. Ovoidal polyad, B. Verrucate ornamentation. P. robusta - C. Polyad with 12 pollen grains, D. Rugulate ornamentation. P. santosii - E. Ovoidal polyad, F. Rugulate ornamentation. P. trisperma - G. Spheroidal polyad, H. Verrucate ornamentation. P. uaupensis - I. Spheroidal polyad, J. Rugulate ornamentation. P. uliginosa - K. Spheroidal polyad, L. Rugulate ornamentation. Scales: 10 µm (A, C, E, G), 5 µm (B, F, H, I), 1 µm (D, J, K, L).
Based on the number of pollen grains per polyad, three types of pollen grains are here recognized:
Type 1 - Polyads with16 pollen grains (Fig. 3 C, I ), predominantly spheroidal, ovoid only in P. killipii, the largest diameter measuring 29.86-38.1 µm. Rugulate exine ornamentation in P. laxipinna, P. imatacae, and P. uliginosa, areolate in P. killipii, P. peruviana, granulate in P. paniculata, and verrucate in P. uaupensis. Included taxa: P. imatacae, P. peruviana, P. uliginosa, P. killipii, P. laxipinna, P. paniculata, and P. uaupensis.
Type 2 - Polyads with 12 pollen grains (Fig. 2 F, K ); predominantly spheroidal, occasionally ovoid in Piptadenia adiantoides, Piptadenia pteroclada, and Piptadenia santosii. The largest diameter measuring 19.33-42.5 µm. Areolate exine ornamentation in P. gonoacantha, and rugulate exine in P. adiantoides, P. buchtienii, P. cuzcoënsis, P. floribunda, P. pteroclada, P. robusta, and P. santosii. Included taxa: P. adiantoides, P. buchtienii, P. cuzcoënsis, P. floribunda, P. gonoacantha, P. pteroclada, P. robusta, and P. santosii.
Type 3 - Polyads with eight pollen grains (Fig. 1 C, E ); predominantly ovoid, possibly spheroidal only in P. micracantha and P. trisperma. The largest diameter measuring 17.3-28.4 µm. Verrucate exine ornamentation in P. irwinii, P. retusa, P. trisperma, areolate in P. ramosissima, and rugulate in P. affinis and P. micracantha.
DISCUSSION
Pollen Morphology
The species of Piptadenia exhibit similarities in the size and shape of their polyads but show variation in the number of pollen grains per polyad and in exine ornamentation. This aligns with findings reported by Caccavari (2002), who highlighted significant palynological variation among Piptadenia species.
The size of the polyads ranged from small to medium, with medium-sized polyads being predominant. This observation is consistent with findings in the literature for various species of the genus (Barth & Yoneshigue, 1966; Sorsa, 1969; Buril & Santos, 2010; Cruz et al., 2017; Ybert et al., 2017).
In terms of shape, spheroidal polyads were predominant among most species, while ovoid polyads were observed in eight species. Caccavari (2002) described ovoid polyads in the majority of the species studied. Although that research also reported subspheroidal and occasionally biconvex polyads, these characteristics were not observed in the present study. Moreover, Buril and Santos (2010) documented polyads ranging from spherical to elliptical in P. stipulacea (Benth.) Ducke (synonymous with P. retusa (Jacq.) P.G. Ribeiro, Seigler & Ebinger). However, in this study, P. retusa exhibited an ovoid shape.
Ybert et al. (2017) analyzed P. paniculata, demonstrating that it exhibits a circular shape when viewed from its shortest diameter and an ellipsoidal shape when viewed from its longest diameter. Similarly, P. trisperma was described as ellipsoidal, with Soares et al. (2022) characterizing it as circular in frontal view and elliptical in lateral view. However, in the present study, both P. paniculata and P. trisperma displayed a spheroidal shape.
Most species exhibited polyads composed of 12 pollen grains (eight species), followed by 16 pollen grains (seven species) and 8 pollen grains (six species). This pattern was previously noted by Kenrick and Knox (1982), who described similar polymorphism in the genus Acacia Mill., associating it with the number of mitoses undergone by pollen cells. On the other hand, Caccavari (2002) described polyads containing 8, 12, or 16 pollen grains in the Piptadenia species analyzed, and these same quantities were observed in the species examined in this study.
Piptadenia retusa exhibited polyads composed of eight pollen grains. This finding contrasts with the data reported by Buril and Santos (2010), who described polyads of this species as consisting of 12 pollen grains arranged in six peripheral and six central positions. Additionally, P. paniculata and P. trisperma exhibited polyads with 16 and 8 pollen grains, respectively, findings consistent with those of Barth and Yoneshigue (1966), Cruz et al. (2017), and Ybert et al. (2017). However, these results differ from those of Soares et al. (2022), who reported polyads with 16 pollen grains for P. trisperma.
Polyads are a common feature in the Mimoseae tribe Initially, it was thought that polyads enhances pollen dispersal efficiency (Kenrick & Knox, 1982). However, Wyatt et al. (2000) argued that this hypothesis is unlikely, as it does not explain why polyads would evolve as a solution to low pollination efficiency in only a limited number of groups. Instead, the authors proposed that the structure of polyads may function to protect pollen grains from dehydration in arid environments (Wyatt et al., 2000).
Four patterns of exine ornamentation were observed in this study: rugulate (found in most species), areolate, verrucate, and granulate, with the latter characterized by granula of various sizes and the presence of microspines (observed only in P. paniculata). Caccavari (2002) described the pollen grains of the Piptadenia group, which included not only Piptadenia but also the genera Adenopodia C.Presl, Anadenanthera, Microlobius, Mimosa, Parapiptadenia, Piptadeniopsis Burkart, Pseudopiptadenia, and Stryphnodendron. In these genera, the exine is described as having granula, areolae, rugulae, or verrucae, with surface patterns ranging from psilate to irregular, undulated, or suprareticulated. However, Caccavari did not provide specific descriptions of exine ornamentation for each taxon and considered exine ornamentation to be a non-distinctive character for the group.
Buril & Santos (2010) observed an areolate exine with irregularly fused areolae in Piptadenia retusa under SEM. However, our results indicate that the exine ornamentation in this species is characterized as verrucate. Additionally, these authors classified P. retusa as part of a distinct and isolated pollen type, which also includes Pityrocarpa moniliformis and Parapiptadenia zehntneri (Harms) M.P.Lima & H.C.Lima. This pollen type is characterized by small-sized, acalymmate polyads, with pollen grains arranged in six peripheral and six central positions, varying in shape from spherical to elliptical, and an exine with irregularly fused areolae.
Under SEM observation, Cruz et al. (2017) described Piptadenia trisperma as having a rugulate exine. However, the voucher cited in their study does not correspond to the species mentioned. P. trisperma, as later characterized by Soares et al. (2022), was described as having pollen grains with psilate-foveolate ornamentation. However, the specimens cited in their study did not display the characteristic spike inflorescence of the genus, suggesting possible identification issues with the voucher used by the authors. In contrast, the present study found that P. trisperma exhibited a verrucate exine.
Taxonomic Implications of Pollen Morphology
Our results highlight that the pollen morphology of Piptadenia species is largely consistent, particularly in terms of size and shape. The main variation lies in the number of pollen grains composing the polyads (8, 12, or 16) and the patterns of exine ornamentation. These findings, combined with morphological and molecular data, provide robust support for the monophyly of the genus, as confirmed by Ribeiro et al. (2018).
Caccavari (2002) demonstrated that the pollen characteristics of taxa within the Piptadenia group support the systematic relationships among the genera included in this group. These characteristics include the use of polyads as dispersal units and the number of pollen grains per polyad (8, 12, 16, 20, and 32), with 8, 12, and 16 being the most common. Additionally, pollen grains are frequently ovoid in shape and exhibit an irregular arrangement of grains, a pattern typically observed in Piptadenia, Stryphnodendron, Parapiptadenia, and Pseudopiptadenia. Furthermore, Caccavari (2002) highlighted the exine structure, which displayed distinctive features such as white lines on the lamellae of the endexine and ectexine, covering the pore and forming a vestibule. However, the author emphasized that exine sculpture is not a distinguishing characteristic of the group.
The genus Piptadenia predominantly occurs in rainforests. However, P. adiantoides, P. gonoacantha, and P. paniculata are also distributed in semi-deciduous forests or gallery forests within the Caatinga or Cerrado biomes (Ribeiro, 2017). Notably, P. adiantoides and P. gonoacantha share rugulate exine ornamentation, while P. paniculata displays an ornamentation pattern with granula of varying sizes and the presence of microspines. Interestingly, verrucate ornamentation, which has been associated with pollination syndromes involving large pollinators such as bats (Banks et al., 2014), had not previously been observed in this group or cited in the reviewed literature. However, this study identified a verrucate ornamentation pattern in P. irwinii, P. retusa, P. trisperma, and P. uaupensis.
Santos-Silva et al. (2013) classified the pollen grains of species from Mimosa ser. Leiocarpe Benth. into two types based on polyad shape and the number of grains per polyad: tetrad and bitetrad types, emphasizing the diversity of pollen morphology within the group. However, their findings do not support the monophyly of Mimosa ser. Leiocarpe based solely on palynological characteristics, as observed in this study.
The most common ornamentation among the studied genera is rugulate, which, according to Punt (1986), is well adapted for pollination by insects. This aligns with the fact that the reproductive success of many Fabaceae species is closely tied to their relationship with bees as primary pollinators, which represent the predominant pollination syndrome in the family (Arroyo, 1981).
Caccavari (2002) highlighted the common pollen characteristics within the Piptadenia group and related taxa, emphasizing their relevance in supporting systematic relationships, particularly regarding exine structure. Similarly, Lewis & Elias (1981) demonstrated that palynological data provided strong evidence of taxonomic affinity among the group and its genera, reinforcing the systematic position of the Piptadenia group. However, subsequent molecular analyses by Jobson & Luckow (2007) and the reassignment of Piptadenia viridiflora to the genus Lachesiodendron by Ribeiro et al. (2018), which supported the monophyly of Piptadenia, revealed a misalignment between palynological tools and the newly proposed phylogenetic arrangement.
The palynological diversity of Neotropical species supports the hypothesis of Leguminosae diversification originating in Central America. This is evidenced by greater morphological diversity in pollen grains of taxa from tropical regions (Piptadenia, Stryphnodendron, and Mimosa) compared to the lower diversity observed in subtropical taxa such as Anadenanthera, Adenopodia, and Microlobius (Caccavari, 2002).
Polyads observed in the genus Mimosa are, in general, highly similar to those found in some species of Piptadenia, which represent a challenge in terms of differentiating between these two groups based solely on pollen morphology (Caccavari, 1987). Barneby (1991) suggested that this character state represents an ancestral trait, linking Mimosa to other genera within the Piptadenia group that share similar pollen grain organization.
According to Caccavari (1987, 2002), Piptadenia s.l. is eurypalynous and can be readily distinguished from other groups based on pollen morphology. However, subsequent phylogenetic studies have redefined the group's circumscription, leading to the segregation of species and the establishment of new genera (Jobson & Luckow, 2007; Ribeiro et al., 2018).
The pollen characteristics of the analyzed species of Piptadenia in this study were largely similar among the examined species, except for differences in the number of pollen grains per polyad (8, 12, or 16) and the ornamentation of the exine (areolate, granulate, rugulate, or verrucate), which varied among species. These two traits enabled the distinction of specific pollen types. Consequently, the palynological data, along with macromorphological and phylogenetic analysis, support the monophyly of the genus. However, palynological data alone are insufficient for delimiting the genus Piptadenia, as taxa within the informal Piptadenia group exhibit overlapping characteristics.
This study provides the first palynological characterization of several species: Piptadenia affinis, P. cuzcoënsis, P. gonoacantha, P. imatacae, P. killipii, P. micracantha, and P. robusta. These findings contribute valuable data to the taxonomic and phylogenetic studies of the group.
Acknowledgments
The authors thank the Fundação de Amparo à Pesquisa do Estado da Bahia (FAPESB) for the fellowship granted to the first author, to the curators of the herbaria BMCB, CEPEC, F, HUEFS, IAN, INPA, MBM, MO, NY, RB, SP, and US. The authors also thank the Laboratório de Micromorfologia Vegetal da Universidade Estadual de Feira de Santana (LAMIV-UEFS) for the use of their light microscopy laboratory facilities and the Instituto Oswaldo Cruz (FIOCRUZ) - Research Center Gonçalo Moniz, for access to the SEM., to the anonymous reviewers for their time and their good comments that certainly have contributed to the improvement of this manuscript. This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brazil (CAPES) - Finance Code 001.
References
- Arroyo MTK. 1981. Breeding systems and pollination biology in Leguminosae, In: Polhill RM, Raven PH (eds.). Advances in Legume Systematics, Part 2. Kew, Royal Botanic Garden. p. 723-769.
- Banks H, Forest F, Lewis G. 2014. Evolution and diversity of pollen morphology in tribe Cercideae (Leguminosae). Taxon 63: 299-314.
- Barneby RC. 1991. Sensitivae Censitae: A description of the genus Mimosa L. (Mimosaceae) in the New World. Memoirs of the New York Botanical Garden 65: 1-835.
- Barros MM. 1963. Contribuição ao estudo das políades polínicas em Leguminosae-Mimosoideae. Atlas da Sociedade de Biologia do Rio de Janeiro 7: 1-11.
- Barth OM, Silva SAF. 1963. Catálogo Sistemático dos Pólens das Plantas Arbóreas do Brasil Meridional. IV - Cunoniaceae, Rosacede e Connaraceae. Memórias do Instituto Oswaldo Cruz 61: 411-427.
- Barth MO, Yoneshigue Y. 1966. Catálogo sistemático dos pólens das plantas arbóreas do Brasil meridional: VIII - Leguminosae (Mimosoideae). Memórias do Instituto Oswaldo Cruz 64: 79-111.
- Bentham G. 1840. Enumeration of Plants Collected by Mr. Schomburgk in British Guiana. Journal of Botany 2: 134-135.
- Bentham G. 1875. Revision of Suborder Mimoseae. Transactions of the Linnaean Society of London 30: 335-664.
- Brenan JPM. 1955. Notes on Mimosoideae: I. Kew Bulletin 10: 161-192.
- Brenan JPM. 1965. A Further Note on Piptadenia. Kew Bulletin 17: 227-228.
- Buril MT, Santos FAR. 2010. Diversidade polínica das Mimosoideae (Leguminosae) ocorrentes em uma área de caatinga, Pernambuco, Brasil. Acta Botanica Brasilica 24: 53-64.
- Caccavari MA. 1987. Ultraestructura del polen de Mimosa (Leguminosae-Mimosoideae). Pollen Spores 30: 275-296.
- Caccavari MA. 2002. Pollen morphology and structure of Tropical and Subtropical American genera of the Piptadenia-group (Leguminosae: Mimosoideae). Grana 41: 130-141.
- Cowan RS, Brenan JPM. 1960. Typification of Piptadenia Bentham. Taxon 9: 56.
- Cruz PF, Mendonça CBF, Gonçalves-Esteves V. 2017. Pollen morphology of selected species of tribes Ingeae and Mimoseae (Mimosoideae - Leguminosae) occurring in the Atlantic Forest of the state of Rio de Janeiro, Brazil. Brazilian Journal of Botany 41: 197-208.
- Du Bocage AL, Souza MA, Miotto STS, Gonçalves-Esteves V. 2008. Palinotaxonomia de espécies de Acacia (Leguminosae - Mimosoideae) no semi-árido brasileiro. Rodriguésia 59: 587-596.
- Duarte AFT, Ribeiro PG, Moreira ALC, Queiroz LP, Santos FARS. 2021. Pollen morphology of Senegalia Raf. species and related genera (Leguminosae: Caesalpinioideae: mimosoid clade). Palynology 45: 1-13.
- Erdtman G. 1952. Pollen Morphology and Plant Taxonomy - Angiosperms. Stockholm, Almqvisit & Wiksell.
- Erdtman G. 1960. The acetolysis method. A revised description. Svensk Botanisk Tidskrift 39: 561-564.
- Guinet PH. 1969. Lês Mimosacées - Étude de palynologie fondamentale corrélations, évolution. Travaux de la Section Scientifique et Technique 9: 32-261.
- Halbritter H, Ulrich S, Grímsson F et al 2018. Illustrated Pollen Terminology. 2nd edn. New York, SpringerOpen.
- Jiménez LCB. 1996. Atlas Palinológico de la Amazonia Colombiana I. Familias Caesalpinaceae, Fabaceae y Mimosaceae. Caldasia 18: 295-327.
- Jobson RW, Luckow M. 2007. Phylogenetic study of the genus Piptadenia (Mimosoideae: Leguminosae) using plastid trnL˗F and trnK/matK sequence data. Systematic Botany 32: 569-575.
- Kenrick J, Knox RB. 1982. Function of the polyad in the reproduction of Acacia Annals of Botany 50: 721-727.
- Lewis GP, Elias TS. 1981. Mimosae. In: Polhill RM, Raven PH (eds.). Advances in Legume Systematics. Kew, Royal Botanic Gardens . p. 155-168.
- Lima LCL, Silva FHM, Santos FAR. 2008. Palinologia de espécies de Mimosa L. (Leguminosae - Mimosoideae) do Semi-Árido brasileiro. Acta Botanica Brasilica 22: 794-805.
- Luckow M, Miller JT, Murphy DJ, Livshultz T. 2003. A phylogenetic analysis of the Mimosoideae (Leguminosae) based on chloroplast DNA sequence data. In: Klitgaard B, Bruneau A (eds.). Advances in Legume Systematics, Part 10, Higher Level Systematics. Kew, Royal Botanic Gardens . p. 197-220.
- Medina-Acosta M, Grether R, Martínez-Bernal A., Ramírez-Arriaga E. 2018. Comparative study of pollen morphology and exine ultrastructure in tetrads, octads and polyads of the genus Mimosa (Leguminosae). Palynology 43: 188-212.
- Punt W. 1986. The Northwest European Pollen Flora, III. Review of Palaeobotany and Palynology 49: 1-96.
- Punt W, Hoen PP, Blackmore S, Nilson SLE, Thomas A. 2007. Glossary of pollen and spore terminology. Review of Palaeobotany and Palynology 143: 1-81.
- Ribeiro PG. 2017. Filogenia molecular do grupo Piptadenia (Leguminosae, Caesalpinioideae, Clado Mimosoide) e taxonomia do gênero Piptadenia Benth. PhD Thesis, Universidade Estadual de Feira de Santana, Brasil.
- Ribeiro PG, Luckow M, Lewis GP et al 2018. Lachesiodendron, a new monospecific genus segregated from Piptadenia (Leguminosae: Caesalpinioideae: mimosoid clade): Evidence from morphology and molecules. Taxon 67: 37-54.
- Santos FAR, Romão CO. 2008. Pollen morphology of some species of Calliandra Benth. (Leguminosae - Mimosoideae) from Bahia, Brazil. Grana 47: 101-116.
- Santos-Silva J, Simon MF, Tozzi AMGA. 2013. Pollen diversity and its phylogenetic implications in Mimosa ser. Leiocarpae Benth. (Leguminosae, Mimosoideae). Grana 52: 15-25.
- Silvestre-Capelato MSF, Melhem TS. 1997. Flora polínica da reserva do Parque Estadual das Fontes do Ipiranga (São Paulo, Brasil) família: Leguminosae. Hoehnea 24: 115-163.
- Soares EL, Landi LADC, Souza CN, Gasparino EC. 2022. Polyads types of the mimosoid clade (Caesalpinioideae, Fabaceae): Size and pollen numbers variations. Grana 61: 45-66.
- Sorsa P. 1969. Pollen morphological studies on the Mimosaceae. Annales Botanici Fennici 6: 13.
-
Thiers B. 2023 [continuously updated]. Index herbariorum: A global directory of public herbaria and associated staff. New York Botanical Garden’s Virtual Herbarium. https://sweetgum.nybg.org/ih. 13 Feb. 2024.
» https://sweetgum.nybg.org/ih. - Van Campo M, Guinet PH. 1961. Les pollens composés. L’ exemple des Mimosacees. Pollen et spores 8: 201-218.
- Wyatt R, Broyles SB, Lipow SR. 2000. Pollen ovules rations in milkweeds (Asclepiadaceae): An exception that proves the rules. Systematic Botany 25: 171-180.
- Ybert JP, Scheel-Ybert R, Carvalho MA. 2017. Grãos de pólen de plantas vasculares do Estado do Rio de Janeiro, Brasil. 3rd ed. Rio de Janeiro: Museu Nacional, Universidade Federal do Rio de Janeiro.



