Resumos
Com o objetivo de caracterizar o padrão de nervação foliar na subfamília Myrtoideae Nied. (Myrtaceae) bem como de reconhecer caracteres particulares a cada subtribo, foram estudadas as folhas de 44 espécies, pertencentes às subtribos Myrtinae (nove gêneros), Myrciinae (cinco gêneros) e Eugeniinae (oito gêneros). Foi observado que, embora o padrão camptódromo-broquidódromo seja o dominante entre as espécies, o padrão misto acródromo-broquidódromo também se encontra presente. Para cada uma das espécies foi descrito o padrão secundário de nervação, o de nervação última marginal, o de nervação terciária, o tipo de aréola, bem como a presença ou ausência de nervura intramarginal.
Myrtaceae; morfologia foliar; nervação foliar; Brasil
Leaf venation in the subfamily Myrtoideae Nied. (Myrtaceae) was examined and the unique features in each subtribe were defined using leaves of 44 species belonging to subtribes Myrtinae (9 genera), Myrciinae (5 genera), and Eugeniinae (8 genera). Although the camptodromous-brochidodromous pattern is dominant among the species, the mixed acrodromous-brochidodromous pattern is also present. For each species, secondary venation pattern, ultimate marginal venation, tertiary venation and areolation, as well as presence or absence of the intramarginal vein were described.
Myrtaceae; leaf morphology; leaf venation; Brazil
Nervação foliar em espécies brasileiras de Myrtaceae Adans.1 1 Parte da Tese de Doutorado do primeiro Autor
Leaf venation in Brazilian species of Myrtaceae Adans.
Cristiano Marcelo Viana Cardoso; Maria das Graças Sajo2 2 Autor para correspondência: mgsajo@rc.unesp.br
Universidade Estadual Paulista, Instituto de Biociências, Departamento de Botânica, UNESP, Av. 24ª, 1515, Bela Vista, C. Postal 199, 13506-900 Rio Claro, SP, Brasil
RESUMO
Com o objetivo de caracterizar o padrão de nervação foliar na subfamília Myrtoideae Nied. (Myrtaceae) bem como de reconhecer caracteres particulares a cada subtribo, foram estudadas as folhas de 44 espécies, pertencentes às subtribos Myrtinae (nove gêneros), Myrciinae (cinco gêneros) e Eugeniinae (oito gêneros). Foi observado que, embora o padrão camptódromo-broquidódromo seja o dominante entre as espécies, o padrão misto acródromo-broquidódromo também se encontra presente. Para cada uma das espécies foi descrito o padrão secundário de nervação, o de nervação última marginal, o de nervação terciária, o tipo de aréola, bem como a presença ou ausência de nervura intramarginal.
Palavras-chave: Myrtaceae, morfologia foliar, nervação foliar, Brasil
ABSTRACT
Leaf venation in the subfamily Myrtoideae Nied. (Myrtaceae) was examined and the unique features in each subtribe were defined using leaves of 44 species belonging to subtribes Myrtinae (9 genera), Myrciinae (5 genera), and Eugeniinae (8 genera). Although the camptodromous-brochidodromous pattern is dominant among the species, the mixed acrodromous-brochidodromous pattern is also present. For each species, secondary venation pattern, ultimate marginal venation, tertiary venation and areolation, as well as presence or absence of the intramarginal vein were described
Key words: Myrtaceae, leaf morphology, leaf venation, Brazil
Introdução
As Myrtaceae Adans. agrupam cerca de 3.100 espécies em aproximadamente 140 gêneros, separados em duas subfamílias: Leptospermoideae e Myrtoideae (Judd et al. 1999). A primeira ocorre predominantemente na Austrália e Polinésia e reúne espécies com fruto seco, geralmente cápsulas loculicidas e folhas alternas. A outra, Myrtoideae, predomina na América tropical e subtropical e compreende espécies com frutos carnosos baciformes e folhas opostas (Cronquist 1981; Heywood 1993). À subfamília Myrtoideae, pertencem todas as Myrtaceae americanas, com exceção de Tepualia stipularis (Hook. & Arn.) Griseb. que, embora ocorra no Chile, pertence à subfamília Lepstopermoideae. Myrtoideae compreende uma única tribo, Myrteae DC., dividida em três subtribos, de acordo com características do embrião (Berg 1855-1856): subtribos Eugeniinae, Myrciinae e Myrtinae. Segundo Landrum & Kawasaki (1997), no Brasil ocorrem 23 gêneros e aproximadamente 1.000 espécies de Myrtaceae. Apesar da importância dessa família nos ecossistemas brasileiros (Mori et al. 1993; Freire 1990; Oliveira Filho & Carvalho 1993; Fabris & Cesar 1996; Soares-Silva, dados não publicados), a morfoanatomia foliar de seus representantes é pouco estudada, destacando-se os trabalhos de Behar (1971), Backes (1971), Du Plessis & Van Wik (1982); Khatijah et al. (1992), Haron & Moore (1996), Fontenelle et al. (1993; 1994), Arruda & Fontenelle (1994), Callado (1997) e Gomes & Neves (1997) sobre anatomia e os Klucking (1988), Costa et al. (1995) e Cardoso & Sajo (2004) sobre venação foliar.
O presente estudo descreve a venação foliar de 44 espécies, pertencentes a 22 dos 23 gêneros, que compõem a subfamília Myrtoideae, no Brasil, com o objetivo principal de contribuir para o conhecimento morfológico da família bem como reconhecer caracteres particulares a cada subtribo.
Material e métodos
O material foi coletado nos municípios de Itirapina, SP (Cerrado), Botucatu, SP (Cerrado), Sete Barras, SP (Mata Atlântica), Londrina, PR (Floresta Estacional Semidecidual) e Brasília, DF (Cerrado). Parte dele foi fixado em FAA 50 e estocado em etanol 50% e parte herborizada e incluída nos herbários HRCB (Herbário Rioclarense), UB (Herbário da Universidade de Brasília) e FUEL (Herbário da Universidade Estadual de Londrina). O material botânico foi identificado por comparação e enviado à Dra. Lucia Helena Soares-Silva, especialista na família, para confirmação.
Os herbários UB e HUEFS (Herbário da Universidade Estadual de Feira de Santana) doaram, respectivamente, material herborizado de Accara elegans (DC.) Landrum, proveniente dos campos rupestres de Minas Gerais e Calycolpus legrandii Mattos, proveniente do nordeste do país, ambas são espécies de distribuição restrita.
A Tabela 1 relaciona os gêneros/número de espécies ocorrentes no Brasil (Landrum & Kawasaki 1997) e o material estudado com o respectivo número de coletor e herbário no qual se encontra depositado. A Tabela 2 contém dados referentes a nervação foliar das espécies estudadas e de outras obtidas na literatura, devidamente citadas.
Foram selecionadas folhas totalmente expandidas, preferencialmente do quarto ou quinto nós. Para cada espécie, as folhas foram diafanizadas a partir de modificações da metodologia proposta por Shobe & Lersten (1967), e montadas entre placas de vidro, tendo como meio de montagem resina sintética.
A rede menor de nervuras foi observada a partir de fragmentos, do terço mediano de folhas diafanizadas, montados em lâminas permanentes e fotografados em fotomicroscópio Olympus BX40 com projeção de escala micrométrica, nas mesmas condições ópticas. As ampliações para a análise do padrão de venação foram confeccionadas em ampliador fotográfico (Durst M601) a partir das próprias placas de vidro, que serviram como "negativos". Para a análise e descrição dos padrões de venação foliar foram utilizados os tipos básicos definidos por Hickey (1974).
Resultados
O padrão de venação observado para a maioria das espécies, das três subtribos, é o camptódromo-broquidódromo; esse padrão se caracteriza por apresentar nervuras secundárias que se anastomosam entre si, desde a base da folha, formando uma série de arcos próximos ao bordo, sendo que em alguns casos esses arcos podem formar outros menores de segunda, terceira ou quarta ordem (Fig. 1-5, 7-18 ). Dependendo do ângulo de fechamento (reto ou próximo a reto), os arcos (laços) broquidódromos podem ser mais ou menos proeminentes (Fig. 1-5, 7-18 ). Em alguns casos (Tab. 2), entre os arcos broquidódromos e a margem da folha, ocorre a formação de uma nervura intramarginal como observado em Calyptranthes concinna DC. (Fig. 11), Gomidesia spectabilis (DC.) O. Berg, Marlierea obscura O. Berg e Myrcia rostrata DC., da subtribo Myrciinae, e em Plinia trunciflora (O. Berg) Kausel (Fig. 14) e Calycorectes psidiiflorus (O. Berg) Sobral e C. australis D. Legrand, da subtribo Eugeniinae. Das espécies estudadas, da subtribo Myrtinae, nenhuma apresentou nervura intramarginal.


Outro padrão de venação, observado para as folhas estudadas, é o tipo misto, acródromo-broquidódromo que aparece apenas em Campomanesia pubescens (DC.) O. Berg (Fig. 6), Campomanesia sp. e Psidium australe Cambess., da subtribo Myrtinae (Tab. 2). Tal padrão se caracteriza pela presença de um a quatro pares de nervuras secundárias laterais, de origem basal ou supra-basal, que tendem ao padrão acródromo e diminuem gradualmente de calibre, geralmente até o terço mediano da folha. A partir desse ponto as nervuras se unem formando arcos broquidódromos.
As nervuras terciárias são ramificadas, percorrentes ou reticuladas, dependendo da espécie considerada, encontrando-se, às vezes, mais de um padrão numa única folha (Tab. 2). As nervuras terciárias são ramificadas quando não se fundem às nervuras secundárias, embora ramificações derivadas delas possam fazê-lo; esse tipo está presente na maioria dos representantes estudados (Tab. 2). As nervuras terciárias ramificadas são admediais, quando as suas ramificações se dirigem à nervura central (Fig. 1-4, 8-12 , 15-18), e exmediais quando elas se dirigem à margem foliar (Fig. 17). No padrão percorrente, que aparece em representantes das três subtribos (Tab. 2), nervuras terciárias originadas de nervuras secundárias opostas, se fundem formando outra nervura paralela à secundária (Fig. 1, 3, 9 , 10-13, 16-18). Quando as nervuras terciárias se fundem umas com as outras ou com nervuras secundárias, elas determinam o padrão reticulado. Se os ângulos de anastomose são predominantemente retos, como observado em Campomanesia guazulmifolia (Cambess.) O. Berg (Fig. 5) e C. pubescens (Fig. 6) o padrão reticulado é chamado ortogonal e quando os ângulos são variáveis, trata-se do padrão reticulado ao acaso, observado, por exemplo, em Myrcia bella Cambess. (Fig. 7 ) e Plinia trunciflora (Fig. 14).
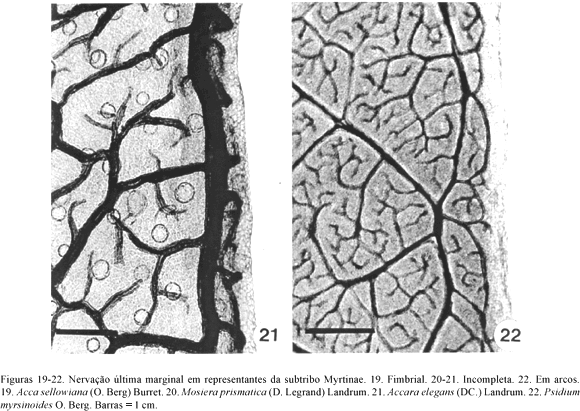
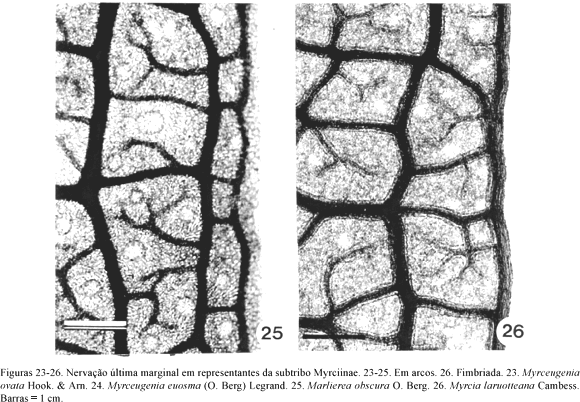
A venação última marginal pode ser: a) fimbriada, quando as nervuras próximas ao bordo se fundem e originam uma nervura contínua (nervura fimbrial), que corre junto à margem, como observado em Acca sellowiana (O. Berg) Burret (Fig. 19), Calycolpus legrandii e Psidium firmum O. Berg, da subtribo Myrtinae, em Myrcia laruotteana Cambess. (Fig. 26), Gomidesia lindeniana O. Berg e Myrcia lingua (O. Berg) Mattos & D. Legrand, da subtribo Myrciinae, e em Myrcianthes pungens (O. Berg) D. Legrand (Fig. 30), Eugenia punicifolia (Kunth.) DC. e Hexachlamys itatiaiensis Mattos, da subtribo Eugeniinae; b) incompleta onde as nervuras, em sua maioria, terminam livres adjacentes à margem como nas folhas de Mosiera prismatica (D. Legrand) Landrum (Fig. 20), Accara elegans (Fig. 21) e Blepharocalyx salicifolius (Kunth.) O. Berg, da subtribo Myrtinae, nas de Marlierea eugeniopsoides (D. Legrand & Kausel) D. Legrand, da subtribo Myrciinae, e nas de Eugenia klotzschiana O. Berg (Fig. 27), Siphoneugena densiflora O. Berg (Fig. 28), Myrciaria delicatula (DC.) O. Berg e Plinia rivularis (Cambess.) A.D. Rotman, da subtribo Eugeniinae e c) em arco, que predomina nos representantes das três subtribos (Tab. 2), como observado em Psidium myrsinoides O. Berg (Fig. 22), Myrceugenia euosma (O. Berg) Legrand e M. ovata Hook. & Arn. (Fig. 23-24), Marlierea obscura (Fig. 25) e Eugenia pyriformis Cambess. (Fig. 29). As nervuras quaternárias e de ordem superior determinam aréolas incompletas, de formato irregular e distribuídas ao acaso. Numa mesma folha, as terminações vasculares podem ser simples ou ramificadas.



Discussão
O padrão de venação mais comum nas folhas dos representantes estudados de Myrtinae, Myrciinae e Eugeniinae é o camptódromo-broquidódromo, embora Klucking (1988) considere o padrão acródromo como predominante para Myrtaceae. Essa diferença nos resultados encontrados talvez se explique porque nos representantes de Myrtoideae estudados pelo referido autor, nota-se que a nervura acródroma é na verdade a nervura marginal resultante da fusão dos arcos broquidódromos.
Nenhuma das folhas estudadas apresenta venação acródroma típica que, segundo descrição de Hickey (1974), se caracteriza pela presença de um ou mais pares de nervuras secundárias de origem basal, ou supra-basal, que se dirigem ao ápice da folha, formando arcos convergentes. Tal padrão, encontrado nas Melastomataceae (Reis et al. 2004) e nas Myrtaceae da subfamília Leptospermoideae (Klucking 1988), representa uma apomorfia para a família segundo Johnson & Briggs (1984) e Wilson et al. (2001). O padrão camptódromo-broquidódromo também predomina nas espécies de Eugenia estudadas por Costa et al. (1995) e por Cardoso & Sajo (2004).
Embora o padrão de venação foliar se mostre bastante uniforme, nas três subtribos, especialmente quanto ao número e ângulo de divergência das nervuras secundárias, e ao ângulo de fechamento dos arcos broquidódromos, determinados aspectos parecem predominar numa subtribo ou outra. Por exemplo, quase todas as espécies estudadas de Eugeniinae apresentam nervuras terciárias percorrentes enquanto nervuras intramarginais estão ausentes nas espécies estudadas de Myrtinae.
Com relação à nervação última marginal, o padrão em arco é o mais freqüente nas três subtribos, seguido do padrão fimbriado e do padrão incompleto. Para as espécies de Eugenia estudadas por Costa et al. (1995) e por Cardoso & Sajo (2004), o padrão fimbriado foi o mais freqüente. Para a nervação terciária observa-se um predomínio do padrão ramificado admedial (RFA), sendo o padrão ramificado exmedial (RFE) exclusivo de alguns representantes de Eugeniinae e o padrão reticulado ortogonal (RTO) restrito a algumas espécies de Myrtinae. Como observado para outras Myrtaceae (Klucking 1988; Costa et al. 1995; Cardoso & Sajo 2004), as nervuras de menor calibre determinam aréolas incompletas, de formato irregular e distribuídas ao acaso; as terminações vasculares podem ser simples ou ramificadas.
Embora as diferenças no padrão de venação foliar tenham permitido a separação das espécies de Eugenia estudadas por Costa et al. (1995) e por Cardoso & Sajo (2004), ele se mostra bastante uniforme nas subtribos de Myrtaceae. Portanto, conforme observado no presente estudo, esse único caráter não deve ser utilizado como distintivo para subtribos, mas pode auxiliar principalmente a identificação de espécies.
Agradecimentos
Os autores agradecem à Dra. Lucia Helena Soares-Silva, pela identificação do material botânico; à Coordenadoria de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), pela concessão da bolsa de Doutorado.
Recebido em 13/06/2005. Aceito em 20/03/2006
- Arruda, R.C.O. & Fontenelle, G.B. 1994. Anatomia foliar de Psidium catleyanun Revista Brasileira de Botânica 17(1): 25-35.
- Backes, A. 1971. Contribuição ao estudo da anatomia foliar e da fisiologia de Psidium multiflorum Camb. Ciência e Cultura 23(3): 297-303.
- Berg, O.C. 1855-1856. Revisio Myrtacearum Americae. Linnaea 27: 1-472.
- Behar, L. 1971. Dados sobre transpiração e anatomia foliar de Eugenia uniflora L. Ciência e Cultura 23(3): 273284.
- Boeger, M.R.T & Wisniewski, C. 2003. Morfologia foliar de espécies arbóreas da Floresta Atlântica. Revista Brasileira de Botânica 26(1): 61-72.
- Callado, C.H. 1997. Anatomia foliar de Eugenia cuprea (Berg) NDZ. e Marlierea suaveolens Gard. (Myrtaceae). Rodriguesia 45/49(71/75): 25-37.
- Cardoso, C.M.V. & Sajo, M.G. 2004. Vascularização foliar e a identificação de espécies de Eugenia L. (Myrtaceae) da bacia hidrográfica do Rio Tibagi, PR. Revista Brasileira de Botânica 27(1): 47-54.
- Costa, C.G.; Machado, R.D. & Fontenelle, J.B. 1995. Sistema vascular em folhas de Eugenia L.(Myrtaceae). Bradea 6(42): 345-356.
- Cronquist, A. 1981. An integrated system of classification of flowering plants New York, Columbia University Press.
- Du Plessis, E. & Van Wik, A.E. 1982. The genus Eugenia (Myrtaceae) in southern Africa: Taxometrics of foliar organography. South African Journal of Botany 1: 147157.
- Fabris, L.C. & Cesar, O. 1996. Estudos florísticos em uma mata litorânea no sul do Estado do Espírito Santo. Boletim do Museu de Biologia Mello Leitão 5: 15-46.
- Ferri, M.G. 1971. Informações sobre transpiração e anatomia foliar de diversas mirtáceas. Ciência e Cultura 23(3): 313-316.
- Fontenelle, G.B.; Costa, C.G. & Machado, R.D. 1994. Foliar anatomy and micromorphology of eleven species of Eugenia L.(Myrtaceae). Botanical Journal of the Linnean Society 115: 111-133.
- Fontenelle, G.B.; Gomes, D.M.S. & Machado, R.D. 1993. Anatomia foliar de Gomidesia martiana Berg e Gomidesia fenzliana Berg (Myrtaceae). Revista Brasileira de Botânica 16(1): 17-30.
- Freire, M.S.B. 1990. Levantamento florístico do Parque Estadual das Dunas de Natal. Acta Botanica Brasilica 4: 41-59.
- Gomes, D.M.S. & Neves, L.J. 1997. Anatomia foliar de Gomidesia spectabilis (DC) Berg. e Gomidesia nitida (Vell.) Legr. (Myrtaceae). Rodriguésia 45/49(71/75): 5170.
- Haron, N.W. & Moore, D.M. 1996. The taxonomic significance of micromorphology in the genus Eugenia L. (Myrtaceae). Botanical Journal of the Linnean Society 120: 265-277.
- Heywood, V.H. 1993. Flowering plants of the world London, B.T. Barsford Ltd.
- Hickey, L.J. 1974. Classificacion de la arquitetura de las hojas de dicotiledoneas. Boletín de la Sociedad Argentina de Botánica 16: 1-26.
- Johnson, L.A.S. & Briggs, B.G. 1984. Myrtales and Myrtaceae - a phylogenetic analysis. Annals of the Missouri Botanical Garden 71: 700-756.
- Judd, W.S.; Campbell, C.S.; Kellogg, E.A. & Stevens, P.F. 1999. Plant systematics a phylogenetic approach. Sunderland, Sinauer Associates.
- Khatijah, H.H.; Cutler, D.R. & Moore, D.M. 1992. Leaf anatomical studies of Eugenia L. (Myrtaceae) species from Malay Peninsula. Botanical Journal of the Linnean Society 110: 137-156.
- Klucking, E.P. 1988. Leaf venation patterns. v. III. Stuttgart, Myrtaceae. J. Cramer.
- Landrum, L.R. & Kawasaki, M.L. 1997. The genera of Myrtaceae in Brazil: an illustrated synoptic treatment and identifications keys. Brittonia 49(4): 508-536.
- Machado, R.; Costa, C.G. & Fontenelle, G.B. 1988. Anatomia foliar de Eugenia sulcata Spring ex Mart. (Myrtaceae). Acta Botanica Brasilica 1(2): 275-285.
- Mori, S.A.; Boom, B.M.; Carvalho, A.M. & Santos, T.S. 1993. Ecological importance of Myrtaceae in an eastern brazilian wet forest. Biotropica 15(1): 68-70.
- Oliveira Filho, A.T. & Carvalho, D.A. 1993. Florística e fisionomia da vegetação do extremo norte do litoral da Paraíba. Revista Brasileira de Botânica 16: 115130.
- Reis, C.; Proença, S.L. & Sajo, M.G. 2004. Vascularização foliar e anatomia do pecíolo de Melastomataceae do cerrado do Estado de São Paulo, Brasil. Acta Botanica Brasilica 18(4): 987-999.
- Shobe, W.R. & Lersten, N.R. 1967. A technique for clearing and staining gymnosperm leaves. Botanical Gazette 127(2): 150-152.
- Souza, A.V.G. 1971. Contribuição ao estudo da anatomia foliar e da fisiologia de Psidium guayava Raddi. Ciência e Cultura 23(3): 373-382.
- Wilson, P.G.; O'Brien, M.M.; Gadek, P.A. & Quinn, C.J. 2001. Myrtaceae revisited: a reassessment of infrafamilial groups. American Journal of Botany 88(11): 2013-2025.

Datas de Publicação
-
Publicação nesta coleção
27 Fev 2007 -
Data do Fascículo
Set 2006
Histórico
-
Recebido
13 Jun 2006 -
Aceito
20 Mar 2006