Abstract
Aim
Our objective was to describe the structure and spatial patterns of the zooplankton community in a small Brazilian river system (Paranaíba River sub-basin) and evaluate the role of an oxbow lake in the dynamics of this community.
Methods
Zooplankton samples were collected from sites located in an oxbow lake and the Aporé River bed during the rainy and dry seasons. We collected these samples from the surface and bottom levels of two different zones (littoral and channel). For each hydrological season, analysis of variance was conducted to determine the differences in species richness and abundance of zooplankton among different sites (longitudinal), zones (lateral), and depths (vertical). Cluster analysis was performed to assess the similarity in species composition. In addition, a canonical correspondence analysis was used to select the environmental variables that best explained the variation in zooplankton abundance data.
Results
We recorded 152 species: 70 rotifers, 53 testate amoebae, 20 cladocerans, and 9 copepods. We observed changes in the species richness and abundance of zooplankton along the longitudinal axis of the river in both the rainy and dry seasons. However, we did not detect any lateral or vertical changes in these attributes. Cluster analysis also indicated that the greatest differences in species composition were in the longitudinal axis of the river. The longitudinal variability of zooplankton was mainly associated with the influence of an upstream oxbow lake, as well as the changes in water flow velocity, turbidity, and concentrations of suspended solids downstream.
Conclusions
Our results demonstrate the importance of a heterogeneity of hydrological characteristics as a structuring factor of the zooplankton community in the Aporé River. Thus, we highlight the relevance of actions to protect and maintain the natural characteristics of these environments.
Keywords:
Aporé river; meander; microhabitat; potamoplankton; taxonomic composition
Resumo
Objetivo
Nosso objetivo foi descrever a estrutura e os padrões espaciais da comunidade zooplanctônica em um pequeno sistema fluvial brasileiro (sub-bacia do rio Paranaíba) e avaliar o papel de um meandro na dinâmica dessa comunidade.
Métodos
Amostras de zooplâncton foram coletadas em pontos localizados em um meandro e no leito do rio Aporé durante os períodos chuvoso e seco. Coletamos essas amostras na superfície e fundo de duas zonas diferentes (litorânea e canal). Para cada período hidrológico, a análise de variância foi conduzida para determinar as diferenças na riqueza de espécies e abundância do zooplâncton entre os diferentes pontos (longitudinal), zonas (lateral) e profundidades (vertical). A análise de cluster foi realizada para avaliar a similaridade na composição de espécies. Além disso, uma análise de correspondência canônica foi usada para selecionar as variáveis ambientais que melhor explicaram a variação nos dados de abundância do zooplâncton.
Resultados
Registramos 152 espécies: 70 de rotíferos, 53 de amebas testáceas, 20 de cladóceros e 9 de copépodes. Observamos mudanças na riqueza de espécies e abundância do zooplâncton ao longo do eixo longitudinal do rio tanto no período chuvoso quanto no seco. No entanto, não detectamos nenhuma mudança lateral ou vertical nesses atributos. A análise de cluster também indicou que as maiores diferenças na composição de espécies estavam no eixo longitudinal do rio. A variabilidade longitudinal do zooplâncton foi associada principalmente a influência de um meandro a montante, bem como a mudanças na velocidade de fluxo d’água, turbidez e concentrações de sólidos suspensos à jusante.
Conclusões
Nossos resultados demonstraram a importância de uma heterogeneidade de características hidrológicas como fator estruturante da comunidade zooplanctônica no rio Aporé. Assim, destacamos a relevância de ações para proteger e manter as características naturais desses ambientes.
Palavras-chave:
rio Aporé; meandro; micro-habitat; potamoplâncton; composição taxonômica
1. Introduction
In river systems, hydrology, connectivity, and geomorphology create a variety of habitats with different environmental conditions. These characteristics reveal that rivers are highly dynamic and heterogeneous environments, exhibiting gradual changes in physical (flow, turbidity, slope, substrate) and chemical (nutrients, dissolved oxygen) variables, which induce habitat preferences and influence the migration and dispersion of aquatic organisms (Pace et al., 1992Pace, M.L., Findlay, S.E.G., & Lints, D., 1992. Zooplankton in advective environments: The Hudson River community and a comparative analysis. Can. J. Fish. Aquat. Sci. 49(5), 1060-1069. http://dx.doi.org/10.1139/f92-117.
http://dx.doi.org/10.1139/f92-117...
; Baranyi et al., 2002Baranyi, C., Hein, T., Holarek, C., Keckeis, S., & Schiemer, F., 2002. Zooplankton biomass and community structure in a Danube River floodplain system: effects of hydrology. Freshw. Biol. 47(3), 473-482. http://dx.doi.org/10.1046/j.1365-2427.2002.00822.x.
http://dx.doi.org/10.1046/j.1365-2427.20...
; Bomfim et al., 2017Bomfim, F.F., Mantovano, T., Amaral, D.C., Palhiarini, W.S., Bonecker, C.C., & Lansac-Tôha, F.A., 2017. Adjacent environments contribute to the increase of zooplankton species in a neotropical river. Acta Limnol. Bras. 29(0), e103. http://dx.doi.org/10.1590/s2179-975x10316.
http://dx.doi.org/10.1590/s2179-975x1031...
; Ramos et al., 2021Ramos, E.A., Okumura, A.T.R., Silva, A.G., Pereira, T.L., & Simões, N.R., 2021. Alpha and beta diversity of planktonic microcrustaceans are associated with environmental heterogeneity in the Frades River Basin, Brazil. Stud. Neotrop. Fauna Environ. 56(3), 1-12. http://dx.doi.org/10.1080/01650521.2021.1933702.
http://dx.doi.org/10.1080/01650521.2021....
).
Zooplankton in river systems occupy an important level of the food web in which they contribute to secondary production and allow for the flow of energy from primary producers to higher organisms (Thorp & Casper, 2003Thorp, J.H., & Casper, A.F., 2003. Importance of biotic interactions in large rivers: an experiment with planktivorous fish, dreissenid mussels and zooplankton in the St Lawrence River. River Res. Appl. 19(3), 265-279. http://dx.doi.org/10.1002/rra.703.
http://dx.doi.org/10.1002/rra.703...
; Sluss et al., 2011Sluss, T.D., Jack, J.D., & Thorp, J.H., 2011. A comparison of sampling methods for riverine zooplankton. River Syst. 19(4), 315-326. http://dx.doi.org/10.1127/1868-5749/2011/0048.
http://dx.doi.org/10.1127/1868-5749/2011...
). Rivers are generally considered inhospitable for the development of zooplankton because of the challenge of swimming against the currents (Hynes, 1970Hynes, H.B.N., 1970. The ecology of running waters. Liverpool: University of Liverpool Press.; Kobayashi et al., 1998Kobayashi, T., Shiel, R.J., & Gibbs, P., 1998. Size structure of river zooplankton: seasonal variation, overall pattern and functional aspect. Mar. Freshw. Res. 49(6), 547-552. http://dx.doi.org/10.1071/MF97254.
http://dx.doi.org/10.1071/MF97254...
; Viroux, 2002Viroux, L., 2002. Seasonal and longitudinal aspects of microcrustacean (Cladocera, Copepoda) dynamics in a lowland river. J. Plankton Res. 4(4), 281-292. http://dx.doi.org/10.1093/plankt/24.4.281.
http://dx.doi.org/10.1093/plankt/24.4.28...
). However, some studies have reported considerable development of zooplanktonic populations in river parts with reduced water flow caused by irregularities in channel morphology, such as anastomosis, backwaters, or depth variations (Basu et al., 2000aBasu, B.K., Kalff, J., & Pinel-Alloul, B., 2000a. Midsummer plankton development along a large temperate river: the St. Lawrence River. Can. J. Fish. Aquat. Sci. 57(S1), 7-15. http://dx.doi.org/10.1139/f99-249.
http://dx.doi.org/10.1139/f99-249...
; Thorp & Mantovani, 2005Thorp, J.H., & Mantovani, S., 2005. Zooplankton of turbid and hydrologically dynamic prairie rivers. Freshw. Biol. 50(9), 1474-1491. http://dx.doi.org/10.1111/j.1365-2427.2005.01422.x.
http://dx.doi.org/10.1111/j.1365-2427.20...
; Zimmermann-Timm et al., 2007Zimmermann-Timm, H., Holst, H., & Kausch, H., 2007. Spatial dynamics of rotifers in a large lowland river, the Elbe, Germany: how important are retentive shoreline habitats for the plankton community? Hydrobiologia 593(1), 49-58. http://dx.doi.org/10.1007/s10750-007-9046-9.
http://dx.doi.org/10.1007/s10750-007-904...
; Picapedra et al., 2018Picapedra, P.H.S., Fernandes, C., Baumgartner, G., & Lansac-Tôha, F.A., 2018. Effect of slackwater areas on the establishment of plankton communities (testate amoebae and rotifers) in a large river in the semi-arid region of northeastern Brazil. Limnetica 37(1), 19-31. https://doi.org/10.23818/limn.37.03.
https://doi.org/10.23818/limn.37.03...
). Furthermore, the “extreme expression” of irregularities in river channel can be found in marginal environments, such as meanders (oxbow lakes). These parts recreate true lentic conditions that provide zooplankton populations for inoculation of riverine stretches downstream (Spaink et al., 1998Spaink, P.A., Ietswaart, T., & Roijackers, R., 1998. Plankton dynamics in a dead arm of the River Waal: a comparison with the main channel. J. Plankton Res. 20(10), 1997-2007. http://dx.doi.org/10.1093/plankt/20.10.1997.
http://dx.doi.org/10.1093/plankt/20.10.1...
; Viroux, 2002Viroux, L., 2002. Seasonal and longitudinal aspects of microcrustacean (Cladocera, Copepoda) dynamics in a lowland river. J. Plankton Res. 4(4), 281-292. http://dx.doi.org/10.1093/plankt/24.4.281.
http://dx.doi.org/10.1093/plankt/24.4.28...
).
However, understanding of factors controlling the structure of riverine zooplankton and their ecological importance is lacking when compared with that of lentic systems (Picapedra et al., 2019Picapedra, P.H.S., Fernandes, C., & Baumgartner, G., 2019. Structure and ecological aspects of zooplankton (Testate amoebae, Rotifera, Cladocera and Copepoda) in highland streams in southern Brazil. Acta Limnol. Bras. 31, e5. http://dx.doi.org/10.1590/s2179-975x2917.
http://dx.doi.org/10.1590/s2179-975x2917...
; Padovesi-Fonseca et al., 2021Padovesi-Fonseca, C., Rezende, R.S., Costa, D.F., & Martins-Silva, M.J., 2021. Spatial scales drive zooplankton diversity in savanna Cerrado streams. Community Ecol. 22(2), 249-259. http://dx.doi.org/10.1007/s42974-021-00052-9.
http://dx.doi.org/10.1007/s42974-021-000...
). Previous studies characterized the zooplankton from streams and small rivers (e.g. Picapedra et al., 2019Picapedra, P.H.S., Fernandes, C., & Baumgartner, G., 2019. Structure and ecological aspects of zooplankton (Testate amoebae, Rotifera, Cladocera and Copepoda) in highland streams in southern Brazil. Acta Limnol. Bras. 31, e5. http://dx.doi.org/10.1590/s2179-975x2917.
http://dx.doi.org/10.1590/s2179-975x2917...
; Gomes et al., 2020Gomes, L.F., Barbosa, J.C., Barbosa, H.O., Vieira, M.C., & Vieira, L.C.G., 2020. Environmental and spatial influences on stream zooplankton communities of the Brazilian Cerrado. Community Ecol. 21(1), 25-31. http://dx.doi.org/10.1007/s42974-020-00008-5.
http://dx.doi.org/10.1007/s42974-020-000...
; Godfrey et al., 2020Godfrey, P.C., Pearson, R.G., Pusey, B.J., & Arthington, A.H., 2020. Drivers of zooplankton dynamics in a small tropical lowland river. Mar. Freshw. Res. 72(2), 173-185. http://dx.doi.org/10.1071/MF20067.
http://dx.doi.org/10.1071/MF20067...
), as well as large rivers (e.g. Bum & Pick, 1996Bum, B.K., & Pick, F.R., 1996. Factors regulating phytoplankton and zooplankton biomass in temperate rivers. Limnol. Oceanogr. 41(7), 1572-1577. http://dx.doi.org/10.4319/lo.1996.41.7.1572.
http://dx.doi.org/10.4319/lo.1996.41.7.1...
; Basu & Pick, 1997Basu, B.K., & Pick, F.R., 1997. Phytoplankton and zooplankton development in a lowland, temperate river. J. Plankton Res. 19(2), 237-253. http://dx.doi.org/10.1093/plankt/19.2.237.
http://dx.doi.org/10.1093/plankt/19.2.23...
; Paggi et al., 2014Paggi, S.B.J., Devercelli, M., & Molina, F.R., 2014. Zooplankton and their driving factors in a large subtropical river during low water periods. Fundam. Appl. Limnol. 184(2), 125-139. http://dx.doi.org/10.1127/1863-9135/2014/0520.
http://dx.doi.org/10.1127/1863-9135/2014...
; Portinho et al., 2016Portinho, J.L., Perbiche-Neves, G., & Nogueira, M.G., 2016. Zooplankton community and tributary effects in free-flowing section downstream a large tropical reservoir. Int. Rev. Hydrobiol. 101(1–2), 48-56. http://dx.doi.org/10.1002/iroh.201501798.
http://dx.doi.org/10.1002/iroh.201501798...
), as having a low density and a predominance of protozoa or rotifers and small-sized microcrustaceans. Moreover, they identified downstream advection and spatiotemporal changes in turbulence, water residence time, turbidity, and food availability as the main factors that regulate these organisms.
Furthermore, most studies describe zooplankton dynamics based only on point samples, normally taken from the middle of the channel, with the lateral and vertical dimensions largely neglected (Viroux, 1999Viroux, L., 1999. Zooplankton distribution in flowing waters and its implications for sampling: case studies in the River Meuse (Belgium) and the River Moselle (France, Luxembourg). J. Plankton Res. 21(7), 1231-1248. http://dx.doi.org/10.1093/plankt/21.7.1231.
http://dx.doi.org/10.1093/plankt/21.7.12...
; Sluss et al., 2011Sluss, T.D., Jack, J.D., & Thorp, J.H., 2011. A comparison of sampling methods for riverine zooplankton. River Syst. 19(4), 315-326. http://dx.doi.org/10.1127/1868-5749/2011/0048.
http://dx.doi.org/10.1127/1868-5749/2011...
; Appel et al., 2019Appel, D.S., Gerrish, G.A., Fisher, E.J., & Fritts, M.W., 2019. Zooplankton sampling in large riverine systems: A gear comparison. River Res. Appl. 36(1), 102-114. http://dx.doi.org/10.1002/rra.3539.
http://dx.doi.org/10.1002/rra.3539...
). The rare studies that have focused on analyzing the possible heterogeneity in zooplankton distribution, such as laterally and vertically, have indicated the existence of some spatial variations in one or all dimensions (e.g. Viroux, 1999Viroux, L., 1999. Zooplankton distribution in flowing waters and its implications for sampling: case studies in the River Meuse (Belgium) and the River Moselle (France, Luxembourg). J. Plankton Res. 21(7), 1231-1248. http://dx.doi.org/10.1093/plankt/21.7.1231.
http://dx.doi.org/10.1093/plankt/21.7.12...
; Thorp et al., 1994Thorp, J.H., Black, A.R., Haag, K.H., & Wehr, J.D., 1994. Zooplankton assemblages in the Ohio River: seasonal, tributary, and navigation dam effects. Can. J. Fish. Aquat. Sci. 51(7), 1634-1643. http://dx.doi.org/10.1139/f94-164.
http://dx.doi.org/10.1139/f94-164...
; Jack et al., 2006Jack, J.D., Fang, W., & Thorp, J.H., 2006. Vertical, lateral and longitudinal movement of zooplankton in a large river. Freshw. Biol. 51(9), 1646-1654. http://dx.doi.org/10.1111/j.1365-2427.2006.01600.x.
http://dx.doi.org/10.1111/j.1365-2427.20...
; Casper & Thorp, 2007Casper, A.F., & Thorp, J.H., 2007. Diel and lateral patterns of zooplankton distribution in the St. Lawrence River. River Res. Appl. 23(1), 73-85. http://dx.doi.org/10.1002/rra.966.
http://dx.doi.org/10.1002/rra.966...
), although no consistent pattern has been observed. However, it should be noted that the zooplankton community of large rivers is better understood than that of small rivers or streams, especially in the tropical region. Investigating the factors that control the zooplankton community structure can be an important step toward establishing protection and conservation measures for the natural characteristics of these environments.
This study aimed to determine the structure and describe the spatial patterns (longitudinal, lateral, and vertical) of the zooplankton community in a small neotropical river during different hydrological seasons (rainy and dry), as well as to assess the role of an oxbow lake on the zooplankton dynamics of the main river channel. We hypothesized the following: (i) Zooplankton of the main river channel will essentially consist of testate amoebae and rotifers, which can develop in dynamic fluvial environments; in the oxbow lake, the zooplankton will consist of planktonic species and/or individuals of microcrustaceans (cladocerans and copepods), owing to their typical lacustrine conditions; (ii) Because of the natural advection process of riverine zooplankton, higher abundances will be observed in the oxbow lake; the higher flow velocities in the main river channel will be related to greater species richness, facilitated by stronger lateral and vertical mixing of water; (iii) Changes in zooplankton structure will be clearer for longitudinal dimension in comparison to lateral and vertical dimensions in the river, as a result of greater heterogeneity of environmental characteristics; (iv) Seasonal differences in spatial distribution patterns of riverine zooplankton will be evident, owing to the variation in the rainfall regime that affects water flow, turbidity, and nutrient input into environments.
2. Material and Methods
2.1. Study area
The Aporé River, located in the Midwest region of Brazil, is a tributary of the Paranaíba River sub-basin. The course of the river flows west/east and stretches between the states of Goiás and Mato Grosso do Sul as a natural border. Finally, the Aporé River flows into the Paranaíba River, which together with the Grande River form the Paraná River.
The region has a tropical climate (Aw, Köppen climate classification), with an average annual temperature of approximately 22.5 °C and rainfall ranging from 60 to 1,700 mm, with rainy and dry seasons. The phytophysiognomy of the basin is located in an area of intersection between two biomes, the Cerrado and the Atlantic Forest (Ramalho et al., 2014Ramalho, W.P., Batista, V.G., & Lozi, L.R.P., 2014. Anfíbios e répteis do médio rio Aporé, estados de Mato Grosso do Sul e Goiás, Brasil. Neotrop. Biol. Conserv. 9(3), 147-160. http://dx.doi.org/10.4013/nbc.2014.93.04.
http://dx.doi.org/10.4013/nbc.2014.93.04...
). This study was conducted in the lower stretch of the Aporé River (Figure 1). This stretch presents a great heterogeneity of hydrological characteristics (e.g. flow and depth), provided by the high sinuosity and the presence of oxbow lakes, waterfalls, rapids, boulders, and littoral vegetation (Table 1; Figure 2).

Map showing the location of the sampling sites and a schematic view of where microhabitats were sampled at each site on the Aporé River, Paranaíba River basin.
Environmental characteristics and physicochemical water variables (mean ± standard deviation) measured along the Aporé River, Paranaíba River basin, during the different hydrological seasons.

Collection sites in the Aporé River, Paranaíba River basin. Lentic (Site 1) and lotic (Sites 2-9) environments.
2.2. Sampling
Diurnal collections were conducted during the rainy (December 2018) and dry (July 2019) seasons. Nine sampling sites were established, one of which was located in an oxbow lake (Site 1) and the other eight were on the Aporé River bed (Sites 2-9). Four samples were taken from each site, comprising two zones: littoral (sublittoral) and channel (middle of the river), and two depths: surface and bottom (Figure 1). A total of 72 samples were collected in the two seasons.
Zooplankton samples were obtained using a motor pump. We filtered 200 L of water per sample using a conical plankton net with a mesh size of 68 µm. Subsequently, the collected material was placed in polyethylene bottles (500 mL), labeled, and fixed in 4% formaldehyde buffered with sodium borate (Na3BO3).
The following environmental variables were also measured: electrical conductivity (μS cm−1; portable conductivity meter, Digimed® DM‐3P), dissolved oxygen (mg L−1; portable oximeter, YSI 550A), pH (portable pH meter, Digimed® DM-2P), water temperature (°C; mercury bulb thermometer), total phosphorus (mg L−1; APHA, 2005American Public Health Association – APHA, 2005. Standard methods for the examination water and wastewater. Washington: APHA.), total suspended solids (mg L−1; APHA, 2005American Public Health Association – APHA, 2005. Standard methods for the examination water and wastewater. Washington: APHA.), turbidity (NTU; portable turbidimeter, LaMotte® 2020i), and water flow velocity (m s−1; mechanical flow meter, General OceanicsTM).
2.3. Data processing and statistical analysis
In the laboratory, samples were concentrated to 50 mL, 10 subsamples were taken with a Hensen-Stempell pipette (2.5 mL), and 25 mL of each sample was evaluated. Sedgewick-Rafter chambers were used to quantify zooplankton under an optical microscope at magnifications from ×10 to ×100. Total density was expressed as individuals per m−3. The species were identified using: Koste (1978)Koste, W., 1978. Rotatoria Die Rädertiere Mitteleuropas begründet von Max Voigt- Monogononta. Berlin: Gebrüder Borntraeger., Elmoor-Loureiro (1997Elmoor-Loureiro, L.M.A., 1997. Manual de identificação de cladóceros límnicos do Brasil. Brasília: Universa., 2010Elmoor-Loureiro, L.M.A. 2010. Cladóceros do Brasil: Famílias Chydoridae e Eurycercidae [online]. Brasília. Retrieved in 2019, January 20, from https://www.cladocera.wordpress.com/.
https://www.cladocera.wordpress.com/...
), Lansac-Tôha et al. (2002)Lansac-Tôha, F.A., Velho, L.F.M., Higuti, J., & Takahashi, E.M., 2002. Cyclopidae (Crustacea, Copepoda) from the upper Paraná River floodplain, Brazil. Braz. J. Biol. 62(1), 125-133. PMid:12185912. http://dx.doi.org/10.1590/S1519-69842002000100015.
http://dx.doi.org/10.1590/S1519-69842002...
, Souza (2008)Souza, M.B.G., 2008. Guia das tecamebas – Bacia do rio Peruaçu – Minas Gerais: subsídio para conservação e monitoramento da bacia do rio São Francisco. Belo Horizonte: Editora da UFMG., and Perbiche-Neves et al. (2015)Perbiche-Neves, G., Boxshall, G., Previatelli, D., Rocha, C.E.F., & Nogueira, M.G., 2015. Identification guide to some Diaptomid species (Crustacea, Copepoda, Calanoida, Diaptomidae) of de la Plata-River Basin (South America). ZooKeys 497(497), 1-111. PMid:25931959. http://dx.doi.org/10.3897/zookeys.497.8091.
http://dx.doi.org/10.3897/zookeys.497.80...
.
For each hydrological season (rainy and dry), we performed principal component analysis (PCA) with a correlation matrix to characterize the environmental conditions of the Aporé River. The data used for this analysis were transformed into log(x+1), except for pH. Subsequently, we applied a one-way permutational multivariate analysis of variance (PERMANOVA), based on Euclidean distance, to determine the differences between the environments, i.e., sites (longitudinal), zones (lateral), and depths (vertical). Additionally, we used two-way PERMANOVA to determine the differences in environments between seasons (site × season, zone × season, and depth × season).
We performed one-way analysis of variance (ANOVA) to compare the changes in species richness (number of species) and abundance of zooplankton (Total, Testate amoebae, Rotifera, Cladocera, and Copepoda) among sites, zones, and depths for each season. We used two-way ANOVA to verify spatial changes in these attributes between seasons. Normality and homoscedasticity (homogeneity of variance) were verified using the Shapiro-Wilk and Levene tests, respectively. When ANOVA was significant (p < 0.05), we used Tukey's post-hoc test to investigate the differences between pairs. A cluster analysis was performed using the Jaccard similarity coefficient (presence/absence) to analyze the spatial similarity in species composition for each season.
The relationship between environmental variables and zooplankton species was explored using canonical correspondence analysis (CCA) (Ter Braak, 1986Ter Braak, C.J.F., 1986. Canonical correspondence analysis: a new eigenvector technique for multivariate direct gradient analysis. Ecology 67(5), 1167-1179. http://dx.doi.org/10.2307/1938672.
http://dx.doi.org/10.2307/1938672...
). For this analysis, the most common species (≥ 25% occurrence) in the samples from each season were used to avoid the effects of rare species. The data used for this analysis were transformed into log(x+1). Additionally, the effect of multicollinearity among environmental variables was analyzed using variance inflation factors. The statistical significance of the eigenvalues and species-environment correlations for the axes generated by the CCA were tested using the Monte Carlo method, based on 999 permutations and a significance level of p < 0.05 (Legendre et al., 2011Legendre, P., Oksanen, J., & Ter-Braak, C.J.F., 2011. Testing the significance of canonical axes in redundancy analysis. Methods Ecol. Evol. 2(3), 269-277. http://dx.doi.org/10.1111/j.2041-210X.2010.00078.x.
http://dx.doi.org/10.1111/j.2041-210X.20...
).
All analyses were performed using the statistical environment R version 3.5.0 (R Development Core Team, 2018R Development Core Team, 2018. R: a language and environment for statistical computing [online]. Vienna: R Foundation for Statistical Computing. Retrieved in 2021, May 25, from https://www.R-project.org/
https://www.R-project.org/...
).
3. Results
3.1. Environmental variables
Environmental heterogeneity was observed only between the Aporé River sites during the rainy (one-way PERMANOVA, df = 8, pseudo-F = 10.9, p < 0.001) and dry (one-way PERMANOVA, df = 8, pseudo-F = 34.5, p < 0.001) seasons. Moreover, environmental changes were noticed at the sites in relation to the two hydrological seasons (two-way PERMANOVA, df = 1, pseudo-F = 13.8, p < 0.001). The environmental characteristics of the Aporé River sites during the different hydrological seasons are described in Table 1.
The first two PCA axes generated for the rainy season explained 68.1% of environmental variability. PC 1 showed that Sites 5-9 had high dissolved oxygen. PC 2 showed that Sites 2, 3, 4, 7, and 9 had high turbidity and total suspended solids. For the dry season, the first two PCA axes explained 67.2% of environmental variability. PC 1 showed that Sites 2-9 had high dissolved oxygen, turbidity, and water flow velocity. PC 2 showed that Sites 3, 7, 8, and 9 had high electrical conductivity and total phosphorus contents. Sites 1, 2, 4, 5, and 6 were associated with low temperature values (Figure 3).

Principal component analysis (PCA) ordination showing the environmental differences for each site in the Aporé River, Paranaíba River basin, during the different hydrological seasons. Abbreviations: Cond, electrical conductivity; O2, dissolved oxygen; Temp, temperature; TP, total phosphorus; TSS, total suspended solids; Turb, turbidity; WV, water flow velocity.
3.2. General scenario of the zooplankton community
The zooplankton community comprised 152 taxa. Rotifera was the richest group in number of species (70 species), followed by testate amoebae (53 species), Cladocera (20 species), and Copepoda (9 species). Testate amoebae were represented by nine families, with Difflugiidae being the richest, with 24 species. The rotifers were distributed in 17 families, of which Lecanidae (17 species) and Brachionidae (15 species) were the most representative in terms of number of species. Among microcrustaceans, cladocerans were represented by seven families, with Chydoridae having the greatest number of species (11). In turn, copepods were distributed into two families, with Cyclopidae being the most representative, with six species. Some taxa occurred in all studied sites: Arcella costata, A. discoides, A. vulgaris, Centropyxis aculeata, Difflugia sp., Cephalodella sp., Collotheca sp., Filinia terminalis, Lecane bulla, L. lunaris, Notommata sp., and nauplii of cyclopoid and calanoid copepods. In addition, we observed that 28 taxa were frequent (≥ 25% occurrence) in the samples, as shown in Table 2.
Taxonomic composition, site occurrence and frequency of occurrence (FO%) of zooplankton in the Aporé River, Paranaíba River basin.
The density of the zooplankton community was mainly driven by rotifers, with Cephalodella sp., F. terminalis, and Notommata sp. as the most abundant taxa. Testate amoebae represented the second most abundant group in the study, with species of the genus Centropyxis dominating. In turn, Bosminopsis deitersi and Alonella dadayi were the most abundant species among cladocerans, and nauplii among copepods.
3.3. Spatial distribution of zooplankton community
We only observed significant differences (ANOVA, p < 0.05) in the attributes (species richness and abundance) of the zooplankton community among the Aporé River sites. We also observed that the richness of cladocerans and abundance of testate amoebae and rotifers showed significant differences in their spatial distribution (sites) between the two hydrological seasons (Table 3).
Analysis of variance (ANOVA) of zooplankton richness and abundance by sites (1-9), zones (littoral and channel), and depths (surface and bottom) within and between the hydrological seasons (rainy and dry).
In terms of species richness, the rainy season was characterized by higher values of total zooplankton and rotifers at Sites 9 and 4, testate amoebae at Sites 6, 9, and 4, and cladocerans and copepods at Sites 4 and 1, respectively. In turn, the dry season was characterized by higher values of total zooplankton and rotifers at Site 4, testate amoebae at Sites 7 and 9, and cladocerans and copepods at Sites 1 and 2. Notably, the distribution of zooplankton species richness among different zones and depths were homogeneous during the two hydrological seasons (Figure 4).

Zooplankton species richness of sites (A and B), zones (C and D), and depths (E and F) of the Aporé River, Paranaíba River basin, during the different hydrological seasons. Data were represented as mean ± standard error.
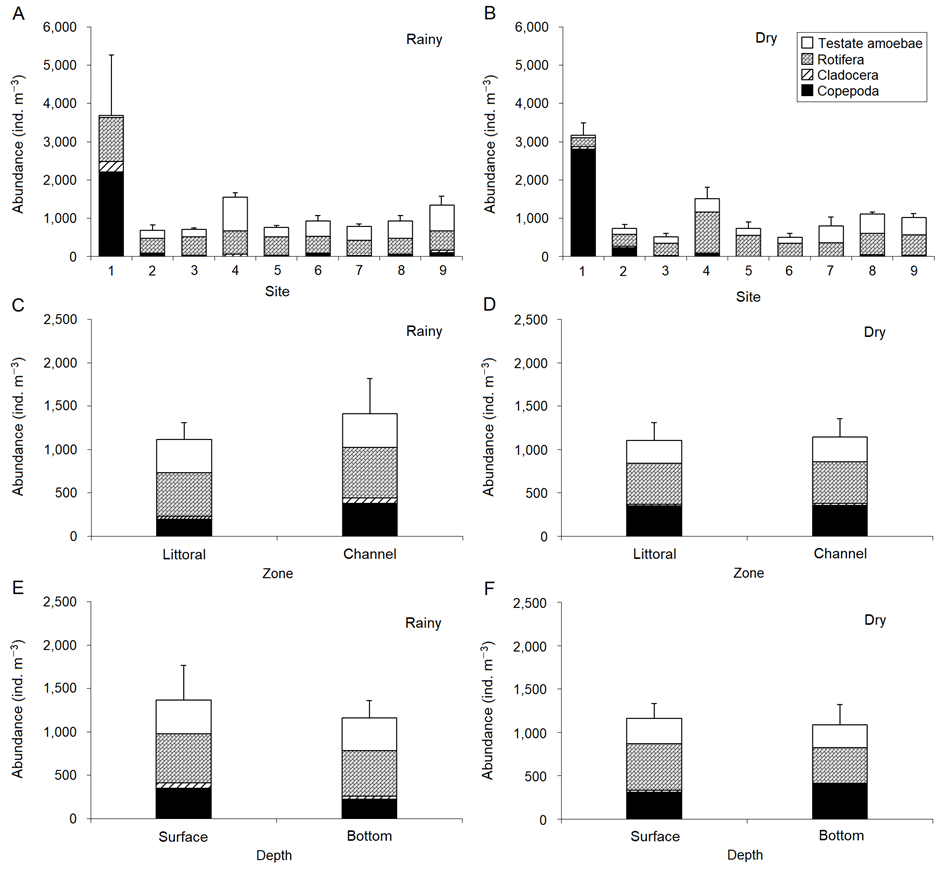
In terms of abundance, the rainy season showed higher values of total zooplankton and copepods at Site 1, whereas testate amoebae showed higher values at Site 4. The distribution of rotifers and cladocerans was found to be homogenous in the sites. In turn, the dry season exhibited higher values of total zooplankton, cladocerans, and copepods at Site 1, whereas testate amoebae and rotifers showed higher values at Sites 8 and 4, respectively. We observed homogeneity in the distribution of zooplankton abundance among different zones and depths of the Aporé River during the two hydrological seasons (Figure 5).

Zooplankton abundance of sites (A and B), zones (C and D) and depths (E and F) of the Aporé River, Paranaíba River basin, during the different hydrological seasons. Data were represented as mean ± standard error.
Cluster analysis based on species occurrence indicated a distinct environmental separation of the Aporé River during different hydrological seasons. During the rainy season, we observed a greater similarity in zooplankton species composition according to the lentic and lotic characteristics of the environments, separating the oxbow lake (A) from the river (B). However, a smaller similarity was noticed between environments during the dry season, as the first group (A) was formed by the oxbow lake and the upstream site of the river (Site 2, channel zone), whereas the other groups (B, C, and D) were formed by the river sites (Figure 6).

Cluster analysis dendrogram showing the similarity between zooplankton-sampled environments in the Aporé River, Paranaíba River basin, during the different hydrological seasons. Abbreviations: CS, channel-surface; CB, channel-bottom; LS, littoral-surface; LB, littoral-bottom. Different letters (A-D) indicate the formation of different groups.
3.4. Relationship between environmental variables and zooplankton
According to the CCAs, the first two axes explained 64.4% and 69.0% of data variability during the rainy and dry seasons, respectively. Furthermore, the species-environment correlation in the first two axes was significant for the rainy (CCA 1, p = 0.001; CCA 2, p = 0.013) and dry (CCA 1, p = 0.001; CCA 2, p = 0.01) seasons, according to the Monte Carlo permutation test (Figure 7).

Canonical correspondence analysis (CCA) ordination showing the relationships between the environmental variables and the environments, and zooplankton taxa of the Aporé River, Paranaíba River basin, during the different hydrological seasons. Abbreviations: Cond, electrical conductivity; O2, dissolved oxygen; Temp, temperature; TP, total phosphorus; TSS, total suspended solids; Turb, turbidity; WV, water flow velocity.
For the rainy season, we found a positive correlation of dissolved oxygen, pH, turbidity, and water flow velocity with the first axis of the CCA, showing that these variables were correlated with the distribution of some species of testate amoebae (e.g. Arcella rotundata, A. discoides, Difflugia acuminata, and D. corona) and rotifers (Cephalodella sp., Notommata sp., and Trichocerca similis) at Sites 3, 5, 6, and 7. The second axis represented the electrical conductivity and total phosphorus. The conductivity was found to be negatively correlated with the abundance of some species of testate amoebae (e.g. Arcella costata, Difflugia sp.), rotifers (e.g. Filinia terminalis, Keratella tropica, and Polyarthra dolichoptera), and copepodites of calanoid copepods at Sites 1 (channel-bottom and littoral-surface) and 2. Total phosphorus was negatively correlated with the abundance of some species of testate amoebae (e.g., Arcella vulgaris, A. megastoma, Centropyxis ecornis, Lesquereusia spiralis), rotifers (e.g., Brachionus angularis, Lecane bulla, and Lepadella ovalis), and the cladoceran Alonella dadayi at sites 4, 8, and 9.
For the dry season, we observed a positive correlation of electrical conductivity, dissolved oxygen, pH, turbidity, and water flow velocity with the first axis of the CCA, indicating their influence on the abundance of some species of testate amoebae (e.g. Arcella vulgaris, Centropyxis cassis, C. constricta, and Cyclopyxis kahli) and rotifers (e.g. Dissotrocha aculeata, Macrochaetus sericus, Tripleuchlanis plicata, and Lepadella ovalis) on Sites 3-9. On the first axis, we found a positive correlation between temperature and the abundance of some species of testate amoebae of the genus Arcella and rotifers (Lecane bulla, L. luna, and Ptygura sp.) at Site 2. The second axis showed a positive correlation between the concentration of total suspended solids and the abundance of some species of rotifers of the genus Lecane and microcrustaceans (nauplii and copepodites of copepods, and cladoceran Alonella dadayi) at Site 1.
4. Discussion
In our study, we found a high species richness of zooplankton when compared to studies in large rivers (e.g. Matsumura-Tundisi et al., 2015Matsumura-Tundisi, T., Tundisi, J.G., Souza-Soares, F., & Tundisi, J.E.M., 2015. Zooplankton community structure of the lower Xingu River (PA) related to the hydrological cycle. Braz. J. Biol. 75(3, Suppl.1), 47-54. PMid:26691075. http://dx.doi.org/10.1590/1519-6984.03814BM.
http://dx.doi.org/10.1590/1519-6984.0381...
; Picapedra et al., 2017Picapedra, P.H.S., Fernandes, C., & Lansac-Tôha, F.A., 2017. Zooplankton community in the Upper Parnaíba River (Northeastern Brazil). Braz. J. Biol. 77(2), 402-412. PMid:27533733. http://dx.doi.org/10.1590/1519-6984.20215.
http://dx.doi.org/10.1590/1519-6984.2021...
; Branco et al., 2018Branco, C.W.C., Silveira, R.M.L., & Marinho, M.M., 2018. Flood pulse acting on a zooplankton community in a tropical river (Upper Paraguay River, Northern Pantanal, Brazil). Fundam. Appl. Limnol. 192(1), 23-42. http://dx.doi.org/10.1127/fal/2018/1155.
http://dx.doi.org/10.1127/fal/2018/1155...
). This fact can be explained by our sampling effort, which sampled different microhabitats, and by the heterogeneity of habitats found along the Aporé River, which differed in their abiotic characteristics. Allied to this, changes in the surrounding characteristics (presence of rural, urban, and preserved areas) along the studied section of the river may also have contributed to the occurrence of a large number of taxa. Topographic features such as stability of the margins and sinuosity of the bed, in addition to the use and occupation of the surroundings, can contribute to a greater or lesser entry of allochthonous material into the bodies of small rivers, imposing different environmental pressures on the organisms that live there (Hepp et al., 2010Hepp, L.U., Milesi, S.V., Biasi, C., & Restello, R.M., 2010. Effects of agricultural and urban impacts on macroinvertebrates assemblages in streams (Rio Grande do Sul, Brazil). Zoologia 27(1), 106-113. http://dx.doi.org/10.1590/S1984-46702010000100016.
http://dx.doi.org/10.1590/S1984-46702010...
).
When pooling the data from our study, rotifers clearly dominated the zooplankton community, both in terms of species richness and density. This predominance of rotifers in rivers has been explained by their lower susceptibility to turbidity, short life cycle, and low predation by fish (Pace et al., 1992Pace, M.L., Findlay, S.E.G., & Lints, D., 1992. Zooplankton in advective environments: The Hudson River community and a comparative analysis. Can. J. Fish. Aquat. Sci. 49(5), 1060-1069. http://dx.doi.org/10.1139/f92-117.
http://dx.doi.org/10.1139/f92-117...
; Sluss et al., 2008Sluss, T.D., Cobbs, G.A., & Thorp, J.H., 2008. Impact of turbulence on riverine zooplankton: a mesocosm experiment. Freshw. Biol. 53(10), 1999-2010. http://dx.doi.org/10.1111/j.1365-2427.2008.02023.x.
http://dx.doi.org/10.1111/j.1365-2427.20...
; Bomfim et al., 2017Bomfim, F.F., Mantovano, T., Amaral, D.C., Palhiarini, W.S., Bonecker, C.C., & Lansac-Tôha, F.A., 2017. Adjacent environments contribute to the increase of zooplankton species in a neotropical river. Acta Limnol. Bras. 29(0), e103. http://dx.doi.org/10.1590/s2179-975x10316.
http://dx.doi.org/10.1590/s2179-975x1031...
). In terms of species richness, the Lecanidae and Brachionidae families were the most relevant, while in terms of abundance, the species of the Notommatidae (Genera Cephalodella and Notommata) and Trochosphaeridae (Filinia terminalis) families predominated. This fact is commonly observed in rivers in South America (Paggi et al., 2014Paggi, S.B.J., Devercelli, M., & Molina, F.R., 2014. Zooplankton and their driving factors in a large subtropical river during low water periods. Fundam. Appl. Limnol. 184(2), 125-139. http://dx.doi.org/10.1127/1863-9135/2014/0520.
http://dx.doi.org/10.1127/1863-9135/2014...
; Picapedra et al., 2017Picapedra, P.H.S., Fernandes, C., & Lansac-Tôha, F.A., 2017. Zooplankton community in the Upper Parnaíba River (Northeastern Brazil). Braz. J. Biol. 77(2), 402-412. PMid:27533733. http://dx.doi.org/10.1590/1519-6984.20215.
http://dx.doi.org/10.1590/1519-6984.2021...
; Branco et al., 2018Branco, C.W.C., Silveira, R.M.L., & Marinho, M.M., 2018. Flood pulse acting on a zooplankton community in a tropical river (Upper Paraguay River, Northern Pantanal, Brazil). Fundam. Appl. Limnol. 192(1), 23-42. http://dx.doi.org/10.1127/fal/2018/1155.
http://dx.doi.org/10.1127/fal/2018/1155...
), where there is a mix of good swimmers from true plankton and poor swimmers from the littoral zone, and combined with the opportunistic characteristic of these organisms of having a large food spectrum, favors the simultaneous presence of many species in the same environment or the same species in environments with different characteristics (Lansac-Tôha et al., 2009Lansac-Tôha, F.A., Bonecker, C.C., Velho, L.F.M., Simões, N.R., Dias, J.D., Alves, G.M., & Takahashi, E.M., 2009. Biodiversity of zooplankton communities in the Upper Paraná River floodplain: interannual variation from long-term studies. Braz. J. Biol. 69(2, Suppl.), 539-549. PMid:19738961. http://dx.doi.org/10.1590/S1519-69842009000300009.
http://dx.doi.org/10.1590/S1519-69842009...
). The testate amoebae were also important in terms of richness and abundance, particularly the Difflugidae, Arcellidae, and Centropyxidae families, which are predominant in the plankton of other tropical rivers (e.g. Picapedra et al., 2018Picapedra, P.H.S., Fernandes, C., Baumgartner, G., & Lansac-Tôha, F.A., 2018. Effect of slackwater areas on the establishment of plankton communities (testate amoebae and rotifers) in a large river in the semi-arid region of northeastern Brazil. Limnetica 37(1), 19-31. https://doi.org/10.23818/limn.37.03.
https://doi.org/10.23818/limn.37.03...
; 2019Picapedra, P.H.S., Fernandes, C., & Baumgartner, G., 2019. Structure and ecological aspects of zooplankton (Testate amoebae, Rotifera, Cladocera and Copepoda) in highland streams in southern Brazil. Acta Limnol. Bras. 31, e5. http://dx.doi.org/10.1590/s2179-975x2917.
http://dx.doi.org/10.1590/s2179-975x2917...
; Gomes et al., 2020Gomes, L.F., Barbosa, J.C., Barbosa, H.O., Vieira, M.C., & Vieira, L.C.G., 2020. Environmental and spatial influences on stream zooplankton communities of the Brazilian Cerrado. Community Ecol. 21(1), 25-31. http://dx.doi.org/10.1007/s42974-020-00008-5.
http://dx.doi.org/10.1007/s42974-020-000...
), mainly due to the high water turbulence that cause the suspension of these organisms from the sediment into the water column. Furthermore, tolerance to physical stress and competitive capacity seem to be key factors that allow these protozoa to remain in riverine plankton (Branco et a., 2018Branco, C.W.C., Silveira, R.M.L., & Marinho, M.M., 2018. Flood pulse acting on a zooplankton community in a tropical river (Upper Paraguay River, Northern Pantanal, Brazil). Fundam. Appl. Limnol. 192(1), 23-42. http://dx.doi.org/10.1127/fal/2018/1155.
http://dx.doi.org/10.1127/fal/2018/1155...
).
Among microcrustaceans, Chydoridae was especially important for the high degree of richness of cladocerans, with most representatives of this family normally associated with littoral vegetation and the benthic interstitial environment (Lansac-Tôha et al., 2009Lansac-Tôha, F.A., Bonecker, C.C., Velho, L.F.M., Simões, N.R., Dias, J.D., Alves, G.M., & Takahashi, E.M., 2009. Biodiversity of zooplankton communities in the Upper Paraná River floodplain: interannual variation from long-term studies. Braz. J. Biol. 69(2, Suppl.), 539-549. PMid:19738961. http://dx.doi.org/10.1590/S1519-69842009000300009.
http://dx.doi.org/10.1590/S1519-69842009...
). In this study, a greater abundance of the littoral cladoceran Alonella dadayi was observed in the lentic water of the oxbow lake during the dry season and downstream of the river during the rainy season, indicating that some chydorids prefer lentic water and are probably transported away from their habitat by the increase in river flow during the rainy season. In turn, the planktonic cladoceran Bosminopsis deitersi (Bosminidae) was the most abundant species, occurring in both lentic and lotic waters. The presence of bosminids in the main river channel can be attributed to the inoculation of individuals from low-flow environments (Zimmermann-Timm et al., 2007Zimmermann-Timm, H., Holst, H., & Kausch, H., 2007. Spatial dynamics of rotifers in a large lowland river, the Elbe, Germany: how important are retentive shoreline habitats for the plankton community? Hydrobiologia 593(1), 49-58. http://dx.doi.org/10.1007/s10750-007-9046-9.
http://dx.doi.org/10.1007/s10750-007-904...
), such as the oxbow lake. In addition, bosminids are favored in river environments because of their shorter generation time, ability to withstand a high sediment rate, and adaptability to feed on detritus and bacteria, which reduces the impact of advection loss (Pace et al., 1992Pace, M.L., Findlay, S.E.G., & Lints, D., 1992. Zooplankton in advective environments: The Hudson River community and a comparative analysis. Can. J. Fish. Aquat. Sci. 49(5), 1060-1069. http://dx.doi.org/10.1139/f92-117.
http://dx.doi.org/10.1139/f92-117...
; Casper & Thorp, 2007Casper, A.F., & Thorp, J.H., 2007. Diel and lateral patterns of zooplankton distribution in the St. Lawrence River. River Res. Appl. 23(1), 73-85. http://dx.doi.org/10.1002/rra.966.
http://dx.doi.org/10.1002/rra.966...
; Paggi et al., 2014Paggi, S.B.J., Devercelli, M., & Molina, F.R., 2014. Zooplankton and their driving factors in a large subtropical river during low water periods. Fundam. Appl. Limnol. 184(2), 125-139. http://dx.doi.org/10.1127/1863-9135/2014/0520.
http://dx.doi.org/10.1127/1863-9135/2014...
; Matsumura-Tundisi et al., 2015Matsumura-Tundisi, T., Tundisi, J.G., Souza-Soares, F., & Tundisi, J.E.M., 2015. Zooplankton community structure of the lower Xingu River (PA) related to the hydrological cycle. Braz. J. Biol. 75(3, Suppl.1), 47-54. PMid:26691075. http://dx.doi.org/10.1590/1519-6984.03814BM.
http://dx.doi.org/10.1590/1519-6984.0381...
). Copepods, represented mainly by nauplii, were the most abundant group in the oxbow lake, owing to their favorable lentic conditions. The scarcity of copepods, especially adults, in river channels can be explained by greater water turbulence and downstream advection, which makes it difficult to search for food and partners for reproduction (Dole-Olivier et al., 2001Dole-Olivier, M.J., Galassi, D.M.P., Marmonier, P., & Creuzé Des Châtelliers, M., 2001. The biology and ecology of lotic microcrustaceans. Freshw. Biol. 44(1), 63-91. http://dx.doi.org/10.1046/j.1365-2427.2000.00590.x.
http://dx.doi.org/10.1046/j.1365-2427.20...
).
Statistical analyses highlighted that the richness, abundance, and composition of zooplankton differed more clearly in the longitudinal axis of the Aporé River, both in the rainy and dry seasons. In our study, there was a predominance of cladoceran and copepod taxa in the oxbow lake, and mainly rotifer and testate amoebae taxa were found along the river. In river systems, different communities appear to have different responses to environmental variables, such as water residence time, turbidity, and flow velocity (Thorp & Mantovani, 2005Thorp, J.H., & Mantovani, S., 2005. Zooplankton of turbid and hydrologically dynamic prairie rivers. Freshw. Biol. 50(9), 1474-1491. http://dx.doi.org/10.1111/j.1365-2427.2005.01422.x.
http://dx.doi.org/10.1111/j.1365-2427.20...
; Lair, 2006Lair, N., 2006. A review of regulation mechanisms of metazoan plankton in riverine ecosystems: aquatic habitat versus biota. River Res. Appl. 22(5), 567-593. http://dx.doi.org/10.1002/rra.923.
http://dx.doi.org/10.1002/rra.923...
). For example, when the water residence time was short, the rotifers dominated the zooplankton, and when the residence time was long, microcrustaceans were more prevalent (Baranyi et al., 2002Baranyi, C., Hein, T., Holarek, C., Keckeis, S., & Schiemer, F., 2002. Zooplankton biomass and community structure in a Danube River floodplain system: effects of hydrology. Freshw. Biol. 47(3), 473-482. http://dx.doi.org/10.1046/j.1365-2427.2002.00822.x.
http://dx.doi.org/10.1046/j.1365-2427.20...
). A similar pattern was also observed by Spaink et al. (1998)Spaink, P.A., Ietswaart, T., & Roijackers, R., 1998. Plankton dynamics in a dead arm of the River Waal: a comparison with the main channel. J. Plankton Res. 20(10), 1997-2007. http://dx.doi.org/10.1093/plankt/20.10.1997.
http://dx.doi.org/10.1093/plankt/20.10.1...
in the Waal River (Netherlands), with a predominance of microcrustaceans in the oxbow lake and rotifers in the main channel. Furthermore, rotifers are better adapted than microcrustaceans to waters with greater turbidity, as they are less negatively affected by high concentrations of suspended material (Kirk & Gilbert, 1990Kirk, K.L., & Gilbert, J.J., 1990. Suspended clay and the population dynamics of planktonic rotifers and cladocerans. Ecology 71(5), 1741-1755. http://dx.doi.org/10.2307/1937582.
http://dx.doi.org/10.2307/1937582...
). High flow velocities are also known to inhibit the reproduction and growth rate of zooplankton populations in rivers. Zooplankton population growth is rarely observed at velocities greater than 0.4 m s−1 (Rzoska, 1978Rzoska, J., 1978. Zooplankton of the Nile system. In: Junk, W., ed. On the nature of rivers with case stories of Nile, Zaire and Amazon. The Hague: Dr Junk Publishers, 333-343.). In the Aporé River, the lowest abundances were recorded at sites that exhibited flow velocities greater than 0.5 m s−1. Finally, the increase in species richness in the river channel may be related to drift or resuspension of organisms, especially rotifers and testate amoebae, from littoral and benthic zones. The microfauna that live at the bottom are under constant threat of being dislodged from their microhabitat due to water turbulence and becoming trapped by the flow of the main river channel (Viroux, 2002Viroux, L., 2002. Seasonal and longitudinal aspects of microcrustacean (Cladocera, Copepoda) dynamics in a lowland river. J. Plankton Res. 4(4), 281-292. http://dx.doi.org/10.1093/plankt/24.4.281.
http://dx.doi.org/10.1093/plankt/24.4.28...
).
The oxbow lake (Site 1) may have played an important role in structuring the zooplankton of the Aporé River. At the downstream site of the oxbow lake (Site 2), we observed planktonic species of rotifers and microcrustaceans, which might have originated from the lake. Because of their low turbulence, shallow depths, and long retention times, fluvial lakes can act as plankton sources for rivers (Basu et al., 2000bBasu, B.K., Kalff, J., & Pinel-Alloul, B., 2000b. The influence of macrophyte beds on plankton communities and their export from fluvial lakes in the St Lawrence River. Freshw. Biol. 45(4), 373-382. http://dx.doi.org/10.1046/j.1365-2427.2000.00635.x.
http://dx.doi.org/10.1046/j.1365-2427.20...
; Viroux, 2002Viroux, L., 2002. Seasonal and longitudinal aspects of microcrustacean (Cladocera, Copepoda) dynamics in a lowland river. J. Plankton Res. 4(4), 281-292. http://dx.doi.org/10.1093/plankt/24.4.281.
http://dx.doi.org/10.1093/plankt/24.4.28...
). However, the influence of adjacent lentic environments may be spatially limited and no longer observable at a distance downstream (Viroux, 1999Viroux, L., 1999. Zooplankton distribution in flowing waters and its implications for sampling: case studies in the River Meuse (Belgium) and the River Moselle (France, Luxembourg). J. Plankton Res. 21(7), 1231-1248. http://dx.doi.org/10.1093/plankt/21.7.1231.
http://dx.doi.org/10.1093/plankt/21.7.12...
), as verified in the other sites of the Aporé River channel. In turn, the predominance of nauplii and bosminids over rotifers in the oxbow lake may be linked to the higher grazing rates of these organisms. Although grazing rates are higher in copepod adults than in nauplii (Casper & Thorp, 2007Casper, A.F., & Thorp, J.H., 2007. Diel and lateral patterns of zooplankton distribution in the St. Lawrence River. River Res. Appl. 23(1), 73-85. http://dx.doi.org/10.1002/rra.966.
http://dx.doi.org/10.1002/rra.966...
), the high densities of nauplii (approximately 85% of the total copepod abundance), should exert a stronger cumulative grazing effect on phytoplankton in low-flow areas, which are generally more productive. Finally, the scarcity of adult copepods in the oxbow lake may be related to the higher concentration of suspended solids that can cause mechanical disturbances in the filtering apparatus (Jönsson et al., 2011Jönsson, M., Ranåker, L., Nicolle, A., Ljungberg, P., Fagerberg, T., Hylander, S., Jephson, T., Lebret, K., von Einem, J., Hansson, L.-A., Nilsson, P.A., Balseiro, E., & Modenutti, B., 2011. Glacial clay affects foraging performance in a Patagonian fish and cladocerans. Hydrobiologia 663(1), 101-108. http://dx.doi.org/10.1007/s10750-010-0557-4.
http://dx.doi.org/10.1007/s10750-010-055...
), and to the greater risk of predation in standing water from the higher density of larvae and adult planktivorous fish (Czerniawski & Domagała, 2012Czerniawski, R., & Domagała, J., 2012. Potamozooplankton communities in three different outlets from mesotrophic lakes located in lake-river system. Oceanol. Hydrobiol. Stud. 41(1), 46-56. http://dx.doi.org/10.2478/s13545-012-0006-2.
http://dx.doi.org/10.2478/s13545-012-000...
; Godfrey et al., 2020Godfrey, P.C., Pearson, R.G., Pusey, B.J., & Arthington, A.H., 2020. Drivers of zooplankton dynamics in a small tropical lowland river. Mar. Freshw. Res. 72(2), 173-185. http://dx.doi.org/10.1071/MF20067.
http://dx.doi.org/10.1071/MF20067...
).
We did not identify significant differences in zooplankton structure between the littoral zone and the main river channel and between different depths, while other studies have recorded zooplankton variability among these microhabitats. For example, greater species richness and abundance of zooplankton may occur in the littoral zone because of the presence of macrophyte beds or other structures (e.g. rocks, tree trunks, and sandbanks), which reduce the flow velocity and create adequate conditions (backwaters) for the development of planktonic populations (Thorp et al., 1994Thorp, J.H., Black, A.R., Haag, K.H., & Wehr, J.D., 1994. Zooplankton assemblages in the Ohio River: seasonal, tributary, and navigation dam effects. Can. J. Fish. Aquat. Sci. 51(7), 1634-1643. http://dx.doi.org/10.1139/f94-164.
http://dx.doi.org/10.1139/f94-164...
, Basu et al., 2000bBasu, B.K., Kalff, J., & Pinel-Alloul, B., 2000b. The influence of macrophyte beds on plankton communities and their export from fluvial lakes in the St Lawrence River. Freshw. Biol. 45(4), 373-382. http://dx.doi.org/10.1046/j.1365-2427.2000.00635.x.
http://dx.doi.org/10.1046/j.1365-2427.20...
; Sluss et al., 2008Sluss, T.D., Cobbs, G.A., & Thorp, J.H., 2008. Impact of turbulence on riverine zooplankton: a mesocosm experiment. Freshw. Biol. 53(10), 1999-2010. http://dx.doi.org/10.1111/j.1365-2427.2008.02023.x.
http://dx.doi.org/10.1111/j.1365-2427.20...
). A greater abundance of zooplankton can also be observed at greater depths during the day in the river channel because of daily vertical migration that occurs to avoid predation by fish (Casper & Thorp, 2007Casper, A.F., & Thorp, J.H., 2007. Diel and lateral patterns of zooplankton distribution in the St. Lawrence River. River Res. Appl. 23(1), 73-85. http://dx.doi.org/10.1002/rra.966.
http://dx.doi.org/10.1002/rra.966...
). Although shoreline macrophyte banks were present at some sites in our study, the banks were especially small and sparse and did not contribute to hydraulic retention. In addition, we found boulder formations both in the littoral zone and in the river channel, which provided high turbulence. Furthermore, because it is a small river, the lateral and vertical dimensions are closer and allow for a greater mixing of water; therefore, thermal or chemical stratification rarely occurs, with abiotic changes being more evident in large rivers (Baranyi et al., 2002Baranyi, C., Hein, T., Holarek, C., Keckeis, S., & Schiemer, F., 2002. Zooplankton biomass and community structure in a Danube River floodplain system: effects of hydrology. Freshw. Biol. 47(3), 473-482. http://dx.doi.org/10.1046/j.1365-2427.2002.00822.x.
http://dx.doi.org/10.1046/j.1365-2427.20...
; Thorp et al., 2006Thorp, J.H., Thoms, M.C., & Delong, M.D., 2006. The riverine ecosystem synthesis: biocomplexity in river networks across space and time. River Res. Appl. 22(2), 123-147. http://dx.doi.org/10.1002/rra.901.
http://dx.doi.org/10.1002/rra.901...
). Thus, hydraulic forces probably affected the distribution of organisms in the lateral and vertical dimensions of our study, causing a mix of species and preventing the spatial heterogeneity of zooplankton.
The seasonal changes in spatial distribution of zooplankton were more significant for the species composition, cladoceran richness, and testate amoebae and rotifer abundances. The smaller similarity in species composition during the dry season may be related to a greater variation in abiotic characteristics (e.g., water flow velocity, electrical conductivity, dissolved oxygen, turbidity) along the longitudinal axis of the river, as indicated by the PCA and CCA. Each season has environmental conditions that influence the distribution of species and that select those with the most similar characteristics, promoting the coexistence of those able to adapt to these conditions (Arrieira et al., 2015Arrieira, R.L., Alves, G.M., Schwind, L.T.F., & Lansac-Tôha, F.A., 2015. Local factors affecting the testate amoebae community (Protozoa: Arcellinida; Euglyphida) in a Neotropical floodplain. J. Limnol. 74(3), 444-452. https://doi.org/10.4081/jlimnol.2015.1078.
https://doi.org/10.4081/jlimnol.2015.107...
). In turn, the increase in the abundance of testate amoebae and the richness of cladocerans in the rainy season may have been induced by their resuspension in river water because of the increase in flow and turbulence. Finally, the increase in rotifer density in the oxbow lake during the rainy season, mainly of lecanid species (e.g. Lecane luna, L. lunaris), may be associated with an increase in the concentration of suspended solids caused by the introduction of allochthonous material. Previous studies have shown that bacteria or organic compounds can adhere to the surface of suspended solids, which may be beneficial for zooplankton species as a food source (Lind & Davalos-Lind, 1991Lind, O.T., & Davalos-Lind, L., 1991. Association of turbidity and organic carbon with bacterial abundance and cell size in a large, turbid, tropical lake. Limnol. Oceanogr. 36(6), 1200-1208. http://dx.doi.org/10.4319/lo.1991.36.6.1200.
http://dx.doi.org/10.4319/lo.1991.36.6.1...
; Melão & Rocha, 2006Melão, M.G.G., & Rocha, O., 2006. Life history, population dynamics, standing biomass and production of Bosminopsis deitersi (Cladocera) in a shallow tropical reservoir. Acta Limnol. Bras. 18(4), 433-450.).
In conclusion, the predominance of rotifers and testate amoebae in the main channel and microcrustaceans in the oxbow lake indicates a distinct adaptive capacity of these organisms to live in these environments. Higher abundance of zooplankton in the oxbow lake was resulted by low water flow velocity and high concentration of suspended material. On the other hand, higher species richness of zooplankton in the river resulted from a high contribution of species with littoral characteristics, owing to considerable mixing of water in the channel. Furthermore, the changes in zooplankton structure were more evident along the longitudinal axis of the river, which can be attributed to a greater influence of the oxbow lake on sites upstream of the river, as well as a greater heterogeneity of abiotic factors in the longitudinal dimension than in the lateral and vertical dimensions. Finally, the spatial and seasonal changes in flow, turbidity, and the concentration of suspended solids seemed to exert an important effect on the attributes, especially the composition and abundance, of riverine zooplankton. Thus, we emphasize the importance of studies in river environments, especially those focused on a high degree of sampling, both spatially and temporally, which allows for a more accurate assessment and understanding of the structure and dynamics of riverine zooplankton, which are currently lacking.
Acknowledgements
We thank the Grupo de Pesquisas em Recursos Pesqueiros e Limnologia (GERPEL) of the Universidade Estadual do Oeste do Paraná (UNIOESTE) for providing logistical support during collection. We are also grateful to the anonymous reviewers, whose detailed comments and constructive suggestions improved the quality of the manuscript.
-
Cite as: Picapedra, P.H.S. Drivers of zooplankton spatial dynamics in a small neotropical river. Acta Limnologica Brasiliensia, 2022, vol. 43, e13.
References
- American Public Health Association – APHA, 2005. Standard methods for the examination water and wastewater. Washington: APHA.
- Appel, D.S., Gerrish, G.A., Fisher, E.J., & Fritts, M.W., 2019. Zooplankton sampling in large riverine systems: A gear comparison. River Res. Appl. 36(1), 102-114. http://dx.doi.org/10.1002/rra.3539
» http://dx.doi.org/10.1002/rra.3539 - Arrieira, R.L., Alves, G.M., Schwind, L.T.F., & Lansac-Tôha, F.A., 2015. Local factors affecting the testate amoebae community (Protozoa: Arcellinida; Euglyphida) in a Neotropical floodplain. J. Limnol. 74(3), 444-452. https://doi.org/10.4081/jlimnol.2015.1078
» https://doi.org/10.4081/jlimnol.2015.1078 - Baranyi, C., Hein, T., Holarek, C., Keckeis, S., & Schiemer, F., 2002. Zooplankton biomass and community structure in a Danube River floodplain system: effects of hydrology. Freshw. Biol. 47(3), 473-482. http://dx.doi.org/10.1046/j.1365-2427.2002.00822.x
» http://dx.doi.org/10.1046/j.1365-2427.2002.00822.x - Basu, B.K., & Pick, F.R., 1997. Phytoplankton and zooplankton development in a lowland, temperate river. J. Plankton Res. 19(2), 237-253. http://dx.doi.org/10.1093/plankt/19.2.237
» http://dx.doi.org/10.1093/plankt/19.2.237 - Basu, B.K., Kalff, J., & Pinel-Alloul, B., 2000a. Midsummer plankton development along a large temperate river: the St. Lawrence River. Can. J. Fish. Aquat. Sci. 57(S1), 7-15. http://dx.doi.org/10.1139/f99-249
» http://dx.doi.org/10.1139/f99-249 - Basu, B.K., Kalff, J., & Pinel-Alloul, B., 2000b. The influence of macrophyte beds on plankton communities and their export from fluvial lakes in the St Lawrence River. Freshw. Biol. 45(4), 373-382. http://dx.doi.org/10.1046/j.1365-2427.2000.00635.x
» http://dx.doi.org/10.1046/j.1365-2427.2000.00635.x - Bomfim, F.F., Mantovano, T., Amaral, D.C., Palhiarini, W.S., Bonecker, C.C., & Lansac-Tôha, F.A., 2017. Adjacent environments contribute to the increase of zooplankton species in a neotropical river. Acta Limnol. Bras. 29(0), e103. http://dx.doi.org/10.1590/s2179-975x10316
» http://dx.doi.org/10.1590/s2179-975x10316 - Branco, C.W.C., Silveira, R.M.L., & Marinho, M.M., 2018. Flood pulse acting on a zooplankton community in a tropical river (Upper Paraguay River, Northern Pantanal, Brazil). Fundam. Appl. Limnol. 192(1), 23-42. http://dx.doi.org/10.1127/fal/2018/1155
» http://dx.doi.org/10.1127/fal/2018/1155 - Bum, B.K., & Pick, F.R., 1996. Factors regulating phytoplankton and zooplankton biomass in temperate rivers. Limnol. Oceanogr. 41(7), 1572-1577. http://dx.doi.org/10.4319/lo.1996.41.7.1572
» http://dx.doi.org/10.4319/lo.1996.41.7.1572 - Casper, A.F., & Thorp, J.H., 2007. Diel and lateral patterns of zooplankton distribution in the St. Lawrence River. River Res. Appl. 23(1), 73-85. http://dx.doi.org/10.1002/rra.966
» http://dx.doi.org/10.1002/rra.966 - Czerniawski, R., & Domagała, J., 2012. Potamozooplankton communities in three different outlets from mesotrophic lakes located in lake-river system. Oceanol. Hydrobiol. Stud. 41(1), 46-56. http://dx.doi.org/10.2478/s13545-012-0006-2
» http://dx.doi.org/10.2478/s13545-012-0006-2 - Dole-Olivier, M.J., Galassi, D.M.P., Marmonier, P., & Creuzé Des Châtelliers, M., 2001. The biology and ecology of lotic microcrustaceans. Freshw. Biol. 44(1), 63-91. http://dx.doi.org/10.1046/j.1365-2427.2000.00590.x
» http://dx.doi.org/10.1046/j.1365-2427.2000.00590.x - Elmoor-Loureiro, L.M.A., 1997. Manual de identificação de cladóceros límnicos do Brasil. Brasília: Universa.
- Elmoor-Loureiro, L.M.A. 2010. Cladóceros do Brasil: Famílias Chydoridae e Eurycercidae [online]. Brasília. Retrieved in 2019, January 20, from https://www.cladocera.wordpress.com/
» https://www.cladocera.wordpress.com/ - Godfrey, P.C., Pearson, R.G., Pusey, B.J., & Arthington, A.H., 2020. Drivers of zooplankton dynamics in a small tropical lowland river. Mar. Freshw. Res. 72(2), 173-185. http://dx.doi.org/10.1071/MF20067
» http://dx.doi.org/10.1071/MF20067 - Gomes, L.F., Barbosa, J.C., Barbosa, H.O., Vieira, M.C., & Vieira, L.C.G., 2020. Environmental and spatial influences on stream zooplankton communities of the Brazilian Cerrado. Community Ecol. 21(1), 25-31. http://dx.doi.org/10.1007/s42974-020-00008-5
» http://dx.doi.org/10.1007/s42974-020-00008-5 - Hepp, L.U., Milesi, S.V., Biasi, C., & Restello, R.M., 2010. Effects of agricultural and urban impacts on macroinvertebrates assemblages in streams (Rio Grande do Sul, Brazil). Zoologia 27(1), 106-113. http://dx.doi.org/10.1590/S1984-46702010000100016
» http://dx.doi.org/10.1590/S1984-46702010000100016 - Hynes, H.B.N., 1970. The ecology of running waters. Liverpool: University of Liverpool Press.
- Jack, J.D., Fang, W., & Thorp, J.H., 2006. Vertical, lateral and longitudinal movement of zooplankton in a large river. Freshw. Biol. 51(9), 1646-1654. http://dx.doi.org/10.1111/j.1365-2427.2006.01600.x
» http://dx.doi.org/10.1111/j.1365-2427.2006.01600.x - Jönsson, M., Ranåker, L., Nicolle, A., Ljungberg, P., Fagerberg, T., Hylander, S., Jephson, T., Lebret, K., von Einem, J., Hansson, L.-A., Nilsson, P.A., Balseiro, E., & Modenutti, B., 2011. Glacial clay affects foraging performance in a Patagonian fish and cladocerans. Hydrobiologia 663(1), 101-108. http://dx.doi.org/10.1007/s10750-010-0557-4
» http://dx.doi.org/10.1007/s10750-010-0557-4 - Kirk, K.L., & Gilbert, J.J., 1990. Suspended clay and the population dynamics of planktonic rotifers and cladocerans. Ecology 71(5), 1741-1755. http://dx.doi.org/10.2307/1937582
» http://dx.doi.org/10.2307/1937582 - Kobayashi, T., Shiel, R.J., & Gibbs, P., 1998. Size structure of river zooplankton: seasonal variation, overall pattern and functional aspect. Mar. Freshw. Res. 49(6), 547-552. http://dx.doi.org/10.1071/MF97254
» http://dx.doi.org/10.1071/MF97254 - Koste, W., 1978. Rotatoria Die Rädertiere Mitteleuropas begründet von Max Voigt- Monogononta. Berlin: Gebrüder Borntraeger.
- Lair, N., 2006. A review of regulation mechanisms of metazoan plankton in riverine ecosystems: aquatic habitat versus biota. River Res. Appl. 22(5), 567-593. http://dx.doi.org/10.1002/rra.923
» http://dx.doi.org/10.1002/rra.923 - Lansac-Tôha, F.A., Bonecker, C.C., Velho, L.F.M., Simões, N.R., Dias, J.D., Alves, G.M., & Takahashi, E.M., 2009. Biodiversity of zooplankton communities in the Upper Paraná River floodplain: interannual variation from long-term studies. Braz. J. Biol. 69(2, Suppl.), 539-549. PMid:19738961. http://dx.doi.org/10.1590/S1519-69842009000300009
» http://dx.doi.org/10.1590/S1519-69842009000300009 - Lansac-Tôha, F.A., Velho, L.F.M., Higuti, J., & Takahashi, E.M., 2002. Cyclopidae (Crustacea, Copepoda) from the upper Paraná River floodplain, Brazil. Braz. J. Biol. 62(1), 125-133. PMid:12185912. http://dx.doi.org/10.1590/S1519-69842002000100015
» http://dx.doi.org/10.1590/S1519-69842002000100015 - Legendre, P., Oksanen, J., & Ter-Braak, C.J.F., 2011. Testing the significance of canonical axes in redundancy analysis. Methods Ecol. Evol. 2(3), 269-277. http://dx.doi.org/10.1111/j.2041-210X.2010.00078.x
» http://dx.doi.org/10.1111/j.2041-210X.2010.00078.x - Lind, O.T., & Davalos-Lind, L., 1991. Association of turbidity and organic carbon with bacterial abundance and cell size in a large, turbid, tropical lake. Limnol. Oceanogr. 36(6), 1200-1208. http://dx.doi.org/10.4319/lo.1991.36.6.1200
» http://dx.doi.org/10.4319/lo.1991.36.6.1200 - Matsumura-Tundisi, T., Tundisi, J.G., Souza-Soares, F., & Tundisi, J.E.M., 2015. Zooplankton community structure of the lower Xingu River (PA) related to the hydrological cycle. Braz. J. Biol. 75(3, Suppl.1), 47-54. PMid:26691075. http://dx.doi.org/10.1590/1519-6984.03814BM
» http://dx.doi.org/10.1590/1519-6984.03814BM - Melão, M.G.G., & Rocha, O., 2006. Life history, population dynamics, standing biomass and production of Bosminopsis deitersi (Cladocera) in a shallow tropical reservoir. Acta Limnol. Bras. 18(4), 433-450.
- Pace, M.L., Findlay, S.E.G., & Lints, D., 1992. Zooplankton in advective environments: The Hudson River community and a comparative analysis. Can. J. Fish. Aquat. Sci. 49(5), 1060-1069. http://dx.doi.org/10.1139/f92-117
» http://dx.doi.org/10.1139/f92-117 - Padovesi-Fonseca, C., Rezende, R.S., Costa, D.F., & Martins-Silva, M.J., 2021. Spatial scales drive zooplankton diversity in savanna Cerrado streams. Community Ecol. 22(2), 249-259. http://dx.doi.org/10.1007/s42974-021-00052-9
» http://dx.doi.org/10.1007/s42974-021-00052-9 - Paggi, S.B.J., Devercelli, M., & Molina, F.R., 2014. Zooplankton and their driving factors in a large subtropical river during low water periods. Fundam. Appl. Limnol. 184(2), 125-139. http://dx.doi.org/10.1127/1863-9135/2014/0520
» http://dx.doi.org/10.1127/1863-9135/2014/0520 - Perbiche-Neves, G., Boxshall, G., Previatelli, D., Rocha, C.E.F., & Nogueira, M.G., 2015. Identification guide to some Diaptomid species (Crustacea, Copepoda, Calanoida, Diaptomidae) of de la Plata-River Basin (South America). ZooKeys 497(497), 1-111. PMid:25931959. http://dx.doi.org/10.3897/zookeys.497.8091
» http://dx.doi.org/10.3897/zookeys.497.8091 - Picapedra, P.H.S., Fernandes, C., & Baumgartner, G., 2019. Structure and ecological aspects of zooplankton (Testate amoebae, Rotifera, Cladocera and Copepoda) in highland streams in southern Brazil. Acta Limnol. Bras. 31, e5. http://dx.doi.org/10.1590/s2179-975x2917
» http://dx.doi.org/10.1590/s2179-975x2917 - Picapedra, P.H.S., Fernandes, C., & Lansac-Tôha, F.A., 2017. Zooplankton community in the Upper Parnaíba River (Northeastern Brazil). Braz. J. Biol. 77(2), 402-412. PMid:27533733. http://dx.doi.org/10.1590/1519-6984.20215
» http://dx.doi.org/10.1590/1519-6984.20215 - Picapedra, P.H.S., Fernandes, C., Baumgartner, G., & Lansac-Tôha, F.A., 2018. Effect of slackwater areas on the establishment of plankton communities (testate amoebae and rotifers) in a large river in the semi-arid region of northeastern Brazil. Limnetica 37(1), 19-31. https://doi.org/10.23818/limn.37.03
» https://doi.org/10.23818/limn.37.03 - Portinho, J.L., Perbiche-Neves, G., & Nogueira, M.G., 2016. Zooplankton community and tributary effects in free-flowing section downstream a large tropical reservoir. Int. Rev. Hydrobiol. 101(1–2), 48-56. http://dx.doi.org/10.1002/iroh.201501798
» http://dx.doi.org/10.1002/iroh.201501798 - R Development Core Team, 2018. R: a language and environment for statistical computing [online]. Vienna: R Foundation for Statistical Computing. Retrieved in 2021, May 25, from https://www.R-project.org/
» https://www.R-project.org/ - Ramalho, W.P., Batista, V.G., & Lozi, L.R.P., 2014. Anfíbios e répteis do médio rio Aporé, estados de Mato Grosso do Sul e Goiás, Brasil. Neotrop. Biol. Conserv. 9(3), 147-160. http://dx.doi.org/10.4013/nbc.2014.93.04
» http://dx.doi.org/10.4013/nbc.2014.93.04 - Ramos, E.A., Okumura, A.T.R., Silva, A.G., Pereira, T.L., & Simões, N.R., 2021. Alpha and beta diversity of planktonic microcrustaceans are associated with environmental heterogeneity in the Frades River Basin, Brazil. Stud. Neotrop. Fauna Environ. 56(3), 1-12. http://dx.doi.org/10.1080/01650521.2021.1933702
» http://dx.doi.org/10.1080/01650521.2021.1933702 - Rzoska, J., 1978. Zooplankton of the Nile system. In: Junk, W., ed. On the nature of rivers with case stories of Nile, Zaire and Amazon. The Hague: Dr Junk Publishers, 333-343.
- Sluss, T.D., Cobbs, G.A., & Thorp, J.H., 2008. Impact of turbulence on riverine zooplankton: a mesocosm experiment. Freshw. Biol. 53(10), 1999-2010. http://dx.doi.org/10.1111/j.1365-2427.2008.02023.x
» http://dx.doi.org/10.1111/j.1365-2427.2008.02023.x - Sluss, T.D., Jack, J.D., & Thorp, J.H., 2011. A comparison of sampling methods for riverine zooplankton. River Syst. 19(4), 315-326. http://dx.doi.org/10.1127/1868-5749/2011/0048
» http://dx.doi.org/10.1127/1868-5749/2011/0048 - Souza, M.B.G., 2008. Guia das tecamebas – Bacia do rio Peruaçu – Minas Gerais: subsídio para conservação e monitoramento da bacia do rio São Francisco. Belo Horizonte: Editora da UFMG.
- Spaink, P.A., Ietswaart, T., & Roijackers, R., 1998. Plankton dynamics in a dead arm of the River Waal: a comparison with the main channel. J. Plankton Res. 20(10), 1997-2007. http://dx.doi.org/10.1093/plankt/20.10.1997
» http://dx.doi.org/10.1093/plankt/20.10.1997 - Ter Braak, C.J.F., 1986. Canonical correspondence analysis: a new eigenvector technique for multivariate direct gradient analysis. Ecology 67(5), 1167-1179. http://dx.doi.org/10.2307/1938672
» http://dx.doi.org/10.2307/1938672 - Thorp, J.H., & Casper, A.F., 2003. Importance of biotic interactions in large rivers: an experiment with planktivorous fish, dreissenid mussels and zooplankton in the St Lawrence River. River Res. Appl. 19(3), 265-279. http://dx.doi.org/10.1002/rra.703
» http://dx.doi.org/10.1002/rra.703 - Thorp, J.H., & Mantovani, S., 2005. Zooplankton of turbid and hydrologically dynamic prairie rivers. Freshw. Biol. 50(9), 1474-1491. http://dx.doi.org/10.1111/j.1365-2427.2005.01422.x
» http://dx.doi.org/10.1111/j.1365-2427.2005.01422.x - Thorp, J.H., Black, A.R., Haag, K.H., & Wehr, J.D., 1994. Zooplankton assemblages in the Ohio River: seasonal, tributary, and navigation dam effects. Can. J. Fish. Aquat. Sci. 51(7), 1634-1643. http://dx.doi.org/10.1139/f94-164
» http://dx.doi.org/10.1139/f94-164 - Thorp, J.H., Thoms, M.C., & Delong, M.D., 2006. The riverine ecosystem synthesis: biocomplexity in river networks across space and time. River Res. Appl. 22(2), 123-147. http://dx.doi.org/10.1002/rra.901
» http://dx.doi.org/10.1002/rra.901 - Viroux, L., 1999. Zooplankton distribution in flowing waters and its implications for sampling: case studies in the River Meuse (Belgium) and the River Moselle (France, Luxembourg). J. Plankton Res. 21(7), 1231-1248. http://dx.doi.org/10.1093/plankt/21.7.1231
» http://dx.doi.org/10.1093/plankt/21.7.1231 - Viroux, L., 2002. Seasonal and longitudinal aspects of microcrustacean (Cladocera, Copepoda) dynamics in a lowland river. J. Plankton Res. 4(4), 281-292. http://dx.doi.org/10.1093/plankt/24.4.281
» http://dx.doi.org/10.1093/plankt/24.4.281 - Zimmermann-Timm, H., Holst, H., & Kausch, H., 2007. Spatial dynamics of rotifers in a large lowland river, the Elbe, Germany: how important are retentive shoreline habitats for the plankton community? Hydrobiologia 593(1), 49-58. http://dx.doi.org/10.1007/s10750-007-9046-9
» http://dx.doi.org/10.1007/s10750-007-9046-9
Edited by
Publication Dates
-
Publication in this collection
13 May 2022 -
Date of issue
2022
History
-
Received
20 Oct 2021 -
Accepted
09 Apr 2022