Abstracts
AIM: Beta diversity is defined as the change in species composition along environmental gradients, and in the present study, we investigated the influence of local (i.e., environmental) and regional (i.e., dispersal) factors in community structure. The aims of this study were to evaluate the beta diversity of phytoplankton communities in the Curuaí floodplain and to determine the relative importance of environmental and spatial processes in shaping phytoplankton community structure; METHOD: The phytoplankton communities were sampled in 16 lakes of the Curuaí floodplain (Amazon Basin) during high-water periods in 2002 and 2003. We used partial redundancy analysis (pRDA) to evaluate the pure effect of environmental (six variables) and spatial (spatial filter) variability on phytoplankton community composition; RESULTS: There were 156 taxa recorded in the two study years, including 122 algae species in 2002 and 66 algae species in 2003. The beta diversity that we measured (βSIM index) was 0.889 in 2002 and 0.789 in 2003. The partitioning variation demonstrated that the majority of variation in phytoplankton community structure was not significantly explained by pure environmental and pure spatial components. However, environmental variables presented a larger coefficient of determination than the spatial variable; CONCLUSION: Other factors than those we measured in this study, such as local variables (i.e., biotic interactions, hydrology, etc.) and stochastic events, affected the absence of significant results in our data. Therefore, we suggest that additional variables, such as biological interactions and other local factors, should be considered in this type of analysis to increase its explanatory power for understanding the variation of diversity in these communities.
metacommunity; partitioning variation; rainy season; Amazon
OBJETIVO: A diversidade beta é definida como as mudanças na composição de espécies ao longo de um gradiente ambiental, e atualmente, ecólogos têm investigado a influência de fatores locais (i.e. ambiente) e regionais (i.e. dispersão) na estrutura de comunidades. Os objetivos desse trabalho foram avaliar a diversidade beta da comunidade fitoplanctônica da planície de inundação do Curuaí e determinar a importância relativa dos processos ambientais e espaciais em sua estrutura; MÉTODO: A comunidade fitoplanctônica foi amostrada em 16 lagos da planície de inundação do Curuaí (Bacia Amazônica) durante o período de águas altas em 2002 e 2003. Usou-se a análise de redundância parcial (pRDA) para avaliar o efeito do ambiente (seis variáveis) e espacial (filtros espaciais) na variabilidade da composição da comunidade fitoplanctônica; RESULTADO: Foram registradas 156 espécies em dois anos de estudos, 122 espécies em 2002 e 66 espécies em 2003. A beta diversidade (índice βSIM) foi 0.889 em 2002 e 0.789 em 2003. A análise de partição da variância revelou que a maior parte da variação da comunidade fitoplanctônica não foi significativamente explicada pelos únicos efeitos ambientais e espaciais. No entanto, as variáveis ambientais no presente estudo apresentaram maior coeficiente de determinação do que as variáveis espaciais; CONCLUSÃO: Outros fatores afetaram a ausência de resultados significativos nos presentes dados, como variáveis locais não mensuradas (i.e. interações biológicas, hidrologia e outros) e eventos estocásticos. Dessa forma, sugerimos que variáveis como interações biológicas e outras variáveis locais sejam consideradas nesse tipo de análise para aumentar o poder de explicação e elucidar a variação da diversidade biológica.
metacomunidade; partição da variância; estação chuvosa; Amazônia
BIOLOGICAL LIMNOLOGY
Ina de Souza NogueiraI; João Carlos NaboutII, III; Maria do Socorro Rodrigues IbañezIV;Laurence Maurice BourgoinV
ILaboratório de Limnologia, Department of Biologia Geral, Instituto de Ciências Biológicas - ICB I,Universidade Federal de Goiás - UFG, CP 131, Campus II, CEP 74001-970, Goiânia, GO, Brazil e-mail: nogueira@icb.ufg.br
IIUnidade Universitária de Ciências Exatas e Tecnologicas - UnUCET, Universidade Estadual de Goiás - UFG, BR-153, CEP 75132-903, Anápolis, GO, Brazil e-mail: naboutjc@hotmail.com
IIIPrograma Nacional de Pós-Doutorado - PNPD/CNPq, Universidade Federal de Goiás - UFG e-mail: naboutjc@hotmail.com
IVDepartamento de Ecologia, Universidade de Brasília - UnB, Darcy Ribeiro Campus Darcy Ribeiro, Asa Norte, CEP 70910-900, Brasília, DF, Brazil e-mail: marias@unb.br
VIRD, Observatoire Midi Pyrénéss, LMTG-UMR 5563, 14 Avenue Edouard Belin, 31400 Toulouse, France e-mail: maurice@lmtg.obs-mip.fr
ABSTRACT
AIM: Beta diversity is defined as the change in species composition along environmental gradients, and in the present study, we investigated the influence of local (i.e., environmental) and regional (i.e., dispersal) factors in community structure. The aims of this study were to evaluate the beta diversity of phytoplankton communities in the Curuaí floodplain and to determine the relative importance of environmental and spatial processes in shaping phytoplankton community structure;
METHOD: The phytoplankton communities were sampled in 16 lakes of the Curuaí floodplain (Amazon Basin) during high-water periods in 2002 and 2003. We used partial redundancy analysis (pRDA) to evaluate the pure effect of environmental (six variables) and spatial (spatial filter) variability on phytoplankton community composition;
RESULTS: There were 156 taxa recorded in the two study years, including 122 algae species in 2002 and 66 algae species in 2003. The beta diversity that we measured (βSIM index) was 0.889 in 2002 and 0.789 in 2003. The partitioning variation demonstrated that the majority of variation in phytoplankton community structure was not significantly explained by pure environmental and pure spatial components. However, environmental variables presented a larger coefficient of determination than the spatial variable;
CONCLUSION: Other factors than those we measured in this study, such as local variables (i.e., biotic interactions, hydrology, etc.) and stochastic events, affected the absence of significant results in our data. Therefore, we suggest that additional variables, such as biological interactions and other local factors, should be considered in this type of analysis to increase its explanatory power for understanding the variation of diversity in these communities.
Keywords: metacommunity, partitioning variation, rainy season, Amazon.
RESUMO
OBJETIVO: A diversidade beta é definida como as mudanças na composição de espécies ao longo de um gradiente ambiental, e atualmente, ecólogos têm investigado a influência de fatores locais (i.e. ambiente) e regionais (i.e. dispersão) na estrutura de comunidades. Os objetivos desse trabalho foram avaliar a diversidade beta da comunidade fitoplanctônica da planície de inundação do Curuaí e determinar a importância relativa dos processos ambientais e espaciais em sua estrutura;
MÉTODO: A comunidade fitoplanctônica foi amostrada em 16 lagos da planície de inundação do Curuaí (Bacia Amazônica) durante o período de águas altas em 2002 e 2003. Usou-se a análise de redundância parcial (pRDA) para avaliar o efeito do ambiente (seis variáveis) e espacial (filtros espaciais) na variabilidade da composição da comunidade fitoplanctônica;
RESULTADO: Foram registradas 156 espécies em dois anos de estudos, 122 espécies em 2002 e 66 espécies em 2003. A beta diversidade (índice βSIM) foi 0.889 em 2002 e 0.789 em 2003. A análise de partição da variância revelou que a maior parte da variação da comunidade fitoplanctônica não foi significativamente explicada pelos únicos efeitos ambientais e espaciais. No entanto, as variáveis ambientais no presente estudo apresentaram maior coeficiente de determinação do que as variáveis espaciais;
CONCLUSÃO: Outros fatores afetaram a ausência de resultados significativos nos presentes dados, como variáveis locais não mensuradas (i.e. interações biológicas, hidrologia e outros) e eventos estocásticos. Dessa forma, sugerimos que variáveis como interações biológicas e outras variáveis locais sejam consideradas nesse tipo de análise para aumentar o poder de explicação e elucidar a variação da diversidade biológica.
Palavras-chaves: metacomunidade, partição da variância, estação chuvosa, Amazônia.
1. Introduction
Species richness is the simpler measurement for quantification and expression of the presence of taxa in a region. However, there are three types of diversity to be considered in biodiversity analyses: alpha, beta and gamma (Magurran, 2004). Beta diversity is an important concept for understanding the functioning of ecosystems and for the conservation of biodiversity. Moreover, ecologists are currently interested not only in analyzing beta diversity, but in interpreting the causes of beta diversity (Legendre et al., 2005). For decades, patterns in the variation of community structure (i.e., beta diversity) were assumed to be mainly controlled by environmental factors and niche differences among species (Tokeshi, 1999). The current neutral theory of biodiversity (Hubbell, 2001) has presented a different point of view, as it assumes that interacting species are equivalent and that population dynamics are driven by random variation in births, deaths, and stochastic, but spatially restricted dispersal.
The Brazilian floodplain lakes provide a good system to test the influence of local (i.e., environmental) and regional factors (i.e., dispersal) in determining metacommunity structure. A metacommunity is a set of local communities within a landscape that are linked by the dispersal of multiple potentially interacting species (Holt, 1991; Wilson, 1992; Cottenie and De Meester, 2004). Currently, four main theoretical paradigms of metacommunity ecology have been proposed (species-sorting, mass-effects, patch-dynamics and neutral), and their difference is based in the importance of local and/or regional factors that affect community structure (Leibold et al., 2004; Thompson and Townsend, 2006).
There have been a number of phycological studies on the Amazon basin (e.g., Ibañez, 1997, 1998; Huszar, 2000; Huszar and Reynolds, 1997; Huszar, 1996a, b; Melo and Huszar, 2000). However, most of these studies only explored phytoplankton composition in a descriptive way (i.e., evaluated the species richness, abundance or dominance of phytoplankton species).
The aims of this study were to evaluate the beta diversity of the phytoplankton community in the Curuaí floodplain and to determine the relative importance of environmental and spatial processes in shaping phytoplankton community structure. Thus, considering the importance of the flood pulse on the homogenization of phytoplankton and limnological attributes (Thomaz et al., 2007), our expectation was that no difference in beta diversity would be found between the years 2002 and 2003 as precipitation levels were similar in both years (see Figure 1 in Materials and Methods), especially when taking into consideration the cumulative effects of this variable. Furthermore, because the samples were taken during the rainy season in both years, we expected that this environmental component would be observed to be shaping the structure of the phytoplankton community; in other words, in the rainy season, all of the lakes in the floodplains in this region are connected, and it is expected that there is a high dispersal capacity of small aquatic organisms (Finlay, 2002; Figuerola and Green, 2002; Padisák, 2004). Thus, we believe that L. G. M. Baas-Becking´s premise can be used to explain the distribution of phytoplankton in the rainy season: "everything is everywhere, but the environment selects" (Green et al., 2008).
2. Materials and Methods
2.1. Study area
The Lago Grande de Curuaí floodplain (01° 50' S and 02° 15' S and 55° 00' W and 56° 05' W, Pará State) is a complex system with more than 16 interconnected lakes that are linked to the Amazon River by nine channels on the south-eastern side (see map in Maurice-Bourgoin et al., 2005). It drains a total area of 3,610 km2, which is covered in the south dense forest.
The Curuaí floodplain is very extensive and shallow, and it is vegetated during the low water season by savannah, shrubs and alluvial forest (RADAMBRASIL, 1976). The inundated area varies from 700 km2 during the dry season to 2,300 km2 at the time of the flood peak (Martinez et al., 2003), representing 0.8% of the total flooded area of the Central Amazon basin. The low water phase usually occurs in November, and the water levels rise from December to May-June (Maurice-Bourgoin et al., 2005).
The Lago Grande de Curuaí lake system is approximately 100-m wide and 3-km long during the low water period; its depth varies between 6 m, during the low water period, to 12 m, during the high water period. As the river level rises, several other channels progressively connect different parts of the floodplain to the main river (Bonnet et al., 2008).
The main lakes in the region are classified as "white-water" lakes, including Lago do Poção, Lago do Salé, Lago de Irateua, Lago da Porta, Lago Santa Ana and Lago Grande de Curuaí, because they are mainly fed by waters from Amazon, while Lago Curumucuri, Lago Açaí and, to a lesser extent, Lago Miuá, are considered to be "black water" lakes, in which the main water input comes from the local runoff from an extensive forest drainage area (Sioli, 1984).
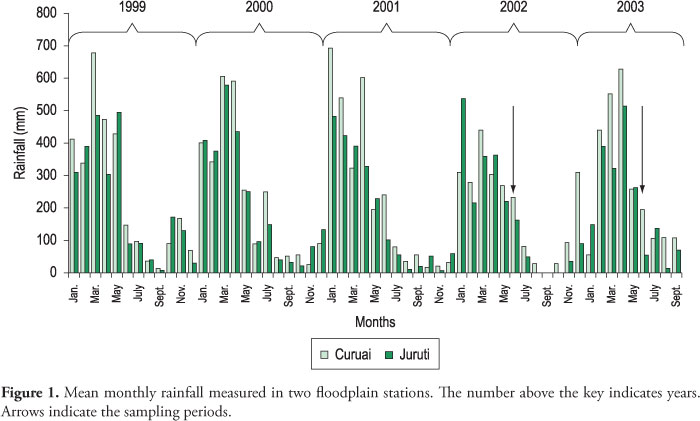
Precipitation at Curuaí is influenced by an orographic effect in the south and differs from rainfall received by lakes in the northern part of the system. Precipitation over the floodplain was higher in 2003 than in 2002 (Figure 1), with maximum levels over 500 mm. In the sampling months for both years, rainfall was approximately 200 mm. The highest rainfall in the Amazon Basin is generally from January to March. However, in 2003 the highest precipitation was recorded in April.
2.2. Biological, limnological and spatial data
The data in this study were collected in two years (2002 and 2003). In 2002 11 lakes in the Curuaí floodplain were sampled. In 2003, 14 lakes were sampled (Table 1). Phytoplankton richness was estimated from subsurface samples collected during high water periods (23-28 June, 2002 and 20-25 June, 2003) by a selective method using a plankton net of 25-μm mesh size. In this study, we collected only qualitative data, though 50 aliquots were analyzed for each sample.
Samples were preserved with Transeau solution (Bicudo and Menezes, 2005). Taxonomic classes were identified according to the classification system of Van Den Hoeck et al. (1993).
Beta diversity index applied to quantify species composition turnover across all lakes was Simpson Index (βSIM; Lennon et al., 2001). Beta diversity can be caused by nestedness and species turnover, thus, the βSIM is adequate to separate turnover from nestedness processes (Baselga, 2010). The βSIM varies from zero (totally similar lakes) to 1 (dissimilar lakes). We performed the βSIM index using R software (function beta.SIM; developed by Baselga, 2010).
Limnological characterization was conducted according to the HiBAM project (Hidrologia e Geoquímica da Amazônia) sampling protocol that followed APHA (1992) methodology. The main aim of this project was to understand the hydrological, sediment and geochemistry processes of the Amazon basin at the continental scale based on a hydrodynamic and geochemistry model of large fluvial systems. The limnological variables analyzed were water temperature (ºC), electrical conductivity (μS/cm), pH, water transparency (m), seston (mg/L) and alkalinity (mg/L).
Spatial variables based on the matrices of Euclidean distance were created using eigenfunction analyses (Borcard and Legendre, 2002; Borcard et al., 2004; Diniz-Filho and Bini, 2005; Dray et al., 2006; Griffith and Peres-Neto, 2006). In these analyses, geographical coordinates were first used to create distance. Then, by following the procedures described in detail by Griffith and Peres-Neto (2006), it was possible to generate distance-based or topology-based eigenvector maps. The eigenvectors (spatial variables, also known as filters) allow the representation of spatial relationships among the sampling units at different spatial scales. In our study, they can be considered to be different and independent propositions of how lakes are geographically related. Positive and negative spatial autocorrelations are represented, respectively, by eigenvectors associated with positive and negative eigenvalues. Small eigenvalues (in absolute terms) indicate the absence of spatial autocorrelation, and, therefore, they are not suitable for defining spatial structures (Griffith and Peres-Neto, 2006; Dray et al., 2006). Here, all eigenvectors with Moran's I coefficients higher than 0.1 were included in the matrices of spatial predictors. For instance, the results of the eigenvector maps derived from the spatial coordinates of the lakes studied indicate the flexibility of this method in creating spatial variables, where the eigenvector maps indicated different levels of complexity in terms of spatial relationships among the lakes. For both years, two spatial filters were selected. Eigenfunction spatial analyses were performed using the freely available Spatial Analysis in Macroecology v. 4.0, SAM package (Rangel et al., 2010).
2.3. Variation partitioning
To estimate the relative importance of environmental and spatial components in shaping phytoplankton community structure, we used a partial redundancy analysis (pRDA) (Legendre and Legendre, 1998). The percentages of the total variation in the phytoplankton matrix that can be attributed to the different components of variation (pure environmental [described by letter a], pure spatial [c], shared by environment and space [b], and unexplained [d]) were based on the adjusted (unbiased) fractions. The significance of each fraction was tested by permutation tests using 999 randomizations (Peres-Neto et al., 2006). All analyses were conducted using the complete dataset. All environmental variables (except pH) were log-transformed before analysis. We performed the above analyses using R software (function Varpart of Vegan Package).
3. Results
The limnological characteristics in the lakes of the Curuaí floodplain were very similar in both years (2002 and 2003), with neutral pH, high concentration of suspended material and low transparency (Table 2).
There were 156 taxa recorded in the two years of the study. These included 122 algae species in 2002 belonging to the following classes: 46 Chlorophyceae, 28 Cyanophyceae and 17 Bacillariophyceae. In 2003, there were 66 algae species, including 21 Bacillariophyceae, 12 Cyanophyceae and 11 Zygnemaphyceae.
Of the lakes studied in 2002 (Figure 2a), Lago Grande Curuaí and Lago Açaí, which are the only two lakes with black water (fed by local rains and drainage from elevated terrain) in the Curuaí floodplain, had the highest richness with 38 and 31 species, respectively. Lake Vermelho had the lowest richness with only two species. In 2003 (Figure 2b), Lago Grande de Curuaí had the highest richness with 41 species, whereas the richness of Lago da Preguiça was extremely species-poor with only two species.
Lago Grande Curuaí presented the highest species richness in both years. Additionally, there was a high frequency of occurrence of some Cyanophyceae in this lake, including Dolichospermum spiroides (Klebahn) Wacklin et al., Dolichospermum circinalis (Rabenh.ex Bornet & Flah.) Wacklin et al., Microcystis aeruginosa (Kütz.) Kütz, Microcystis wesenbergii (Kom.) Kom. in Kondr., Microcysitis panniformis Kom. et al., Radiocystis fernandoi Kom. et Kom.-Legn. and Planktothrix agardhii (Gom.) Kom. et Anagn.
In both 2002 and 2003, none of the species we observed were present in all lakes; however, the most frequent taxa in 2002 were Aulacoseira italica (Ehr.) Simonsen¸ A. herzogii (Lemm.) Simonsen and Ulnaria ulna (Nitz.) Compère, which were found in four lakes. The most frequent species in 2003 was Aulacoseira granulata (Ehr.) Simonsen var. granulata, which was found in 13 lakes.
The Simpson index (βSIM) for the Curuaí floodplain was 0.889 in 2002 and 0.789 in 2003, showing that the lakes in 2002 were more dissimilar in their phytoplankton communities than in 2003.
In both sampling years, we applied the analysis of variation partitioning (using a partial redundancy analysis) to decompose the pure importance of environmental and spatial components on phytoplankton community variation, thus, we observed, in both years, that phytoplankton community variation in phytoplankton community structure was not significantly explained by pure environmental and pure spatial components (Table 3).
4. Discussion
Floodplains store water during the rising stage of the Amazon River and during the rainy season and release it when the river level is decreasing. It has been estimated that approximately 30% of the water in the Amazon River passes through the connected floodplains (Richey et al., 1989). The limnological variables analyzed in this studied did not present great variation among years, probably because the samples were obtained during high water in both years. Moreover, in the limnological literature, it is very common to analyze and investigate seasonal variation (e.g., Loverde-Oliveira and Huszar, 2007); however, other authors that analyzed interannual variation registered distinct results, such as great interannual variation (Nabout et al., 2007) or no interannual variation (Shoup and Wahl, 2009).
We registered high beta diversity values (βSIM index) in the phytoplankton community of the Curuai floodplain in both years. However, in 2002 beta diversity was higher than in 2003. In 2002, most environmental variables had higher variation coefficients, indicating higher environmental variability. Some authors had observed that more variation in environmental gradients (i.e., a larger coefficient of variation) can promote high beta diversity (Harrison et al., 1992; Bini et al., 2001; Nabout et al., 2007).
In the field of phycology, Huszar et al. (1990) discussed beta diversity in Brazil, focusing on the lower Doce River lakes. They found elevated beta diversity and attributed it to elevated environmental heterogeneity due mainly to anthropic effects. Nogueira et al. (2008) registered elevated beta diversity in artificial lakes (urban lakes), varying from 0.4 to 0.65 using the other index (β-1; Harrinson et al., 1992). For floodplain lakes along the Paraná River (Borges and Train, 2009) and Araguaia River (Nabout et al., 2007), using β-1 index, registered lower beta diversity than we observed in this study.
Currently, a large number of papers have discussed the determinants of beta diversity utilizing different groups of organisms as models (e.g., Beisner et al., 2006; Soininen et al., 2007; Nabout et al., 2009; Heino et al., 2010). The proposal of the neutral theory (Hubbel, 2001) and statistical advances (i.e., partial RDA, partial Mantel and others) have promoted a new debate about the relative importance of regional (i.e., dispersal) or local (i.e., environmental) components in structuring metacommunities. Despite the great amount of discussion in the scientific literature, the answer to this question is far from final. Clearly, answering questions such as which environmental or local variable (Beisner et al., 2006), temporal and spatial scale (Borcard et al., 2004), and spatial data (Legendre et al., 2005) is important to understand the determinants of beta diversity. The study we present here contributes to this discussion by analyzing the floodplain model and the relative role of environmental and spatial factors in the phytoplankton structure in a floodplain in the Amazon River Basin.
We expected that environmental factors would be the spatial factors that played the largest role in explaining the structure of phytoplankton communities in the Curuaí floodplain in both years. We found that neither environmental nor spatial factors explained a significant proportion of the total variation in phytoplankton community structure, though the results were similar in both years. This result is very similar to the ones found in phytoplankton communities in lakes in Canada (Beisner et al., 2006) and the Araguaia river floodplain (Nabout et al., 2009).
Others authors found significance of spatial or environmental variables utilizing distinct communities. However, in most of these studies, a large portion of the total variation remained unexplained, such as the diatom community in one lake of Scotland (Yang et al., 2009), diatom communities in rivers in Finland (Heino et al., 2010), phytoplankton communities in Canada (Beisner et al., 2006) and Brazil (Nabout et al., 2009), and macroinvertebrates in streams in Brazil (Roque et al., 2008). These papers all discussed the importance of local or regional effect, despite the fact that all of them have one similarity: a large part of the variance was not accounted for. Thus, the high amount of unexplained variation (i.e., residual) observed in our data and in phytoplankton data from other studies causes us to inquire about which local variables are important in order to explain phytoplankton beta diversity.
Other local environmental variables can be relevant to the structure of phytoplankton communities, and these will contribute to reducing the portion of the total variation that is not explained by the environmental variables we utilized. For example, variables such as hydrological aspects, water residence time, lake geomorphology, mixing patterns and soil characteristics can all be drivers of phytoplankton community structure (Padisák, 2004; Gruberts et al., 2007; Cowell and Dawes, 2008). Additionally, investigating biological interactions among phytoplankton, such as zooplankton herbivory or other top-down effects could increase our understanding of the factors structuring phytoplankton communities (Søndergaard et al., 1990; Jeppesen et al., 1997; Padisák, 2004). Besides biological interactions, the phytoplankton community could be regulated by stochastic events, resulting in communities that are randomly assembled (Sloan et al., 2006).
Finally, our analyses agree with other phytoplankton studies (Beisner et al., 2006; Nabout et al., 2009) and provide evidence of the importance of negative results (Young et al., 2008) in discussions of conceptual models of metacommunities. Additionally, we believe that the inclusion of other local variables, such as biological interactions and fine-scale environmental variation, should increase the explanatory power of these models.
Acknowledgements
This study was funded by the HiBAm Program of IRD, the French Research Development Institute and ANA (Brazilian Water Agency), in a joint cooperation agreement between IRD and CNPq (Brazil). The authors are grateful to CPRM (Manaus), ANA (Brasilia) and Geraldo Resende Boaventura from Brasilia University for their cooperation. We thank the crews who accompanied us during the surveys and assisted us at various stages in the field work. João C. Nabout received a Pos-Doc scholarship from CNPq (151531/2009-9). The referee was outstandingly helpful with this paper, and we extend our sincere gratitude for your efforts.
Received: 05 October 2009
Accepted: 27 September 2010
- American Public Health Association - APHA, American Water Works Association - AWWA and Water Environment Federation - WEF. 1992. Standard Methods for the Examination of Water and Wastewater 18th ed. Washington, D.C.: American Public Health Association.
- BASELGA, A. 2010. Partitioning the turnover and nestedness components of beta diversity. Global Ecology and Biogeography, vol. 19, p. 134-143.
- BEISNER, BE., PERES-NETO, PR., LINDSTROM, ES., BARNETT, A. and LONGHI, ML. 2006. The role of environmental and spatial processes in structuring lake communities from bacteria to fish. Ecology, vol. 87, p. 2985-2991.
- BICUDO, CE. and MENEZES, M. 2005. Gêneros de algas de águas continentais do Brasil. Chave para identificação e descrições. São Carlos. RiMa. 508 p.
- BINI, LM., THOMAZ, SM. and SOUZA, DC. 2001. Species richness and beta diversity of aquatic macrophytes in the Upper Paraná River floodplain. Archiv für Hydrobiologie, vol. 151, no. 3, p. 511-525.
- BONNET, MP., BARROUX, G., MARTINEZ, JM., SEYLER, F., MOREIRA-TURCQ, P., COCHONNEAU, G., MELACK, JM., BOAVENTURA, G., MAURICE-BOURGOIN, L., LEON, JG., ROUX, E., CALMANT, S., KOSUTH, P., GUYOT, JL. and SEYLER, P. 2008. Floodplain hydrology in an Amazon floodplain lake (Lago Grande de Curuai). Journal of Hydrology, vol. 349, no. 1-2, p. 18-30.
- BORCARD, D. and LEGENDRE, P. 2002. All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices. Ecological Modelling, vol. 153, p. 51-68.
- BORCARD, D., LEGENDRE P., AVOIS-JACQUET, V. and TUOMISTO, H. 2004. Dissecting the spatial structure of ecological data at multiple scales. Ecology, vol. 85, no. 7, p. 1826-1832.
- BORGES, PAF. and TRAIN, S. 2009. Phytoplankton diversity in the Upper Paraná River floodplain during two years of drought (2000 and 2001). Brazilian Journal of Biology, vol. 69, no. 2, p. 637-647. Suppl.
- COTTENIE, K. and DE MEESTER, L. 2004. Metacommunity structure: synergy of biotic interactions as selective agents and dispersal as fuel. Ecology, vol. 85, p. 114-119.
- COWELL, BC. and DAWES, CJ. 2008. Seasonal comparisons of the phytoplankton and decline in water transparency in the spring-fed Rainbow River, Florida. Journal of Freshwater Ecology, vol. 23, p. 169-177.
- DINIZ-FILHO, JAF. and BINI, LM. 2005. Modelling geographical patterns in species richness using eigenvector based spatial filters. Global Ecology Biogeography, vol. 14, p. 177-185.
- DRAY, S., LEGENDRE, P. and PERES-NETO, PR., 2006. Spatial modeling: a comprehensive framework for principal coordinate analysis of neighbor matrices (PCNM). Ecological Modelling, vol. 196, p. 483-493.
- FIGUEROLA, J. and GREEN, AJ. 2002. Dispersal of aquatic organisms by waterbirds: a review of past research and priorities for future studies. Freshwater Biology, vol. 47, p. 483-494.
- FINLAY, BJ., 2002. Global dispersal of free-living microbial eukaryote species. Science, vol. 296, p. 1061-1063.
- GREEN, JL., BOHANNAN, BJM. and WHITAKER, RJ., 2008. Microbial Biogeography: From Taxonomy to Traits. Science, vol. 320, p. 1039-1043.
- GRIFFITH, DA. and PERES-NETO, PR., 2006. Spatial modeling in ecology: the flexibility of eigenfunction spatial analyses. Ecology, vol. 87, p. 2603-2613.
- GRUBERTS, D., DRUVIETIS, I., PARELE, E., PAIDERE, J., POPPELS, A., PRIEDITIS, J. and SKUTE, A. 2007. Impact of hydrology on aquatic communities of floodplain lakes along the Daugava River (Latvia). Hydrobiologia, vol. 584, p. 223-237.
- HARRINSON, S., ROSS, SJ. and LAWTON, JH. 1992. Beta diversity on geographic gradients in Britain. Journal of Animal Ecology, vol. 62, p. 151-158.
- HEINO, J., BINI, LM., KARJALAINEN, SM., MYKRÃ, H., SOININEN, J., VIEIRA, LCG. and DINIZ-FILHO, JAF. 2010. Geographical patterns of micro-organismal community structure: are diatoms ubiquitously distributed across boreal streams? Oikos, vol. 119, p. 129-137.
- HOLT, RD., 1991. From metapopulation dynamics to community structure: some consequences of spatial heterogeneity. In GILPIN, ME. and HANSKI, I., ed. Metapopulation dynamics: empirical and theoretical investigations. London: Academic Press. p. 149-165.
- HUBBELL, SP. 2001. A Unified Neutral Theory of Biodiversity and Biogeography Princeton: Princeton University Press. 382 p.
- HUSZAR, VLM. 1996a. Planktonic algae, other than desmids, of three Amazonian systems (Lake Mussurá and Trombetas River), Pará, Brazil. Amazoniana, vol. 14, p. 37-73.
- HUSZAR, VLM. 1996b. Floristic composition and biogeographical aspects of the phytoplankton of an Amazonian flood-plain lake (Lago Batata, Pará, Brazil). Acta Limnologica Brasiliensia, vol. 8, p. 127-136.
- HUSZAR, VLM. 2000. Fitoplâncton. In BOZELLI, RL., ESTEVES, FA. and ROLAND, F., ed. Lago Batata: impacto e recuperação de um ecossistema amazônico. Rio de Janeiro: IB-UFRJ/SBL. p. 91-104.
- HUSZAR, VLM. and REYNOLDS CS. 1997. Phytoplankton periodicity and sequences of dominance in an Amazonian flood-plain lake (Lago Batata, Pará, Brazil): responses to gradual environmental change. Hydrobiologia, vol. 346, p. 169-181.
- HUSZAR, VLM., SILVA, LH. and ESTEVES, F. 1990. Estrutura das comunidades fitoplanctônicas de 18 lagoas da região do Baixo Rio Doce, Linhares, Espírito Santo, Brasil. Revista Brasileira de Biologia, vol. 50, p. 585-598.
- IBAÑEZ, MSR., 1997. Phytoplankton biomass of a central Amazonian flood-plain lake. Verhandlungen des Internationalen Verein Limnologie, vol. 26, p. 605-609.
- IBAÑEZ, MSR. 1998. Phytoplankton composition and abundance of a Central Amazonian flood plain lake. Hydrobiologia, vol. 362, p. 79-83.
- JEPPESEN, E., JENSEN, JP., SONDERGAARD, M., LAURIDSEN, T., PEDERSEN, LJ and JENSEN, L., 1997. Top-down control in freshwater lakes: The role of nutrient state, submerged macrophytes and water depth. Hydrobiologia, vol. 342, p. 151-164.
- LEGENDRE, P. and LEGENDRE, L., 1998. Numerical ecology Elsevier, Amsterdam. 853 p.
- LEGENDRE, P., BORCARD, D. and PERES-NETO, PR., 2005. Analyzing beta diversity: partitioning the spatial variation of community composition data. Ecological Monographs, vol. 75, no. 4, p. 435-450.
- LEGENDRE, P., 2008. Studying beta diversity: ecological variation partitioning by multiple regression and canonical analysis. Journal of Plant Ecology, vol. 1, p. 3-8.
- LEIBOLD, M.A., HOLYOAK, M., MOUQUET, N., AMARASEKARE, P., CHASE, JM., HOOPES, MF., HOLT, R D., SHURIN, J B, LAW, R., TILMAN, D., LOREAU, M. and GONZALEZ, A., 2004. The metacommunity concept: a framework for multi-scale community ecology. Ecology Letters, vol. 7, p. 601-613.
- LENNON, JJ., KOLEFF, P., GREENWOOD, JJD. and GASTON, KJ., 2001 The geographical structure of British bird distributions: diversity, spatial turnover and scale. Journal of Animal Ecology, vol. 70, p. 966-979.
- LOVERDE-OLIVEIRA, S. and HUSZAR, VLM., 2007. Phytoplankton ecological responses to the flood pulse in a Pantanal lake, Central Brazil. Brazil. Acta Limnologica Brasiliensia, vol. 19, p. 5-19.
- MAGURRAN, AF., 2004. Measuring Biological diversity Blackwell Publishing, 256 p.
- MARTINEZ, JM., KOSUTH, P., COCHONNEAU, G., MAURICE-BOURGOIN, L., SEYLER, F., BOURREL, L., and GUYOT, JL., 2003. Application of remote sensing data for the quantification of an Amazon floodplain extension, dynamics and water storage EGS-AGU-EUG Joint Assembly, Nice, April, France.
- MAURICE-BOURGOIN, L., MARTINEZ, J.M., GRÉLAUD, J., FILIZOLA, N., and BOAVENTURA, GR., 2005. The role of flood plains in the hydrology and sediment dynamics of the Amazon River, Brazil, in "Sediment budgets 1&2" (D. E. WALLING and A. J. HOROWITZ, Eds.), IAHS Publication, vol. 291, p. 310-322.
- MELO, S. and HUSZAR, VLM., 2000. Phytoplankton in an Amazonian flood-plain lake (Lago Batata, Brasil): diel variation and species strategies. Journal of Plankton Research, vol. 22, no.1, p. 63-76.
- NABOUT, JC., NOGUEIRA, IS., OLIVEIRA, LG. and MORAIS, RR., 2007. Phytoplankton diversity (alpha, beta and gamma) from the Araguaia River tropical floodplain lakes (central Brazil). Hydrobiologia, vol. 575, no. 1, p. 455-461.
- NABOUT, JC., SIQUEIRA, T., BINI, LM. and NOGUEIRA, IS., 2009. No evidence for environmental and spatial processes in structuring phytoplankton communities. Acta Oecologica, vol. 35, p. 720-726.
- NOGUEIRA, IS., NABOUT, JC., OLIVEIRA, JE. and SILVA, KD., 2008. Diversidade (alfa, beta, gama) da comunidade fitoplanctônica de quatro lagos artificiais urbanos do município de Goiânia, GO. Hoehnea, vol. 35, p. 219-233.
- PADISÁK, J., 2004. Phytoplankton. In: O'Sullivan, P.E., Reynolds, C.S. (Eds.), The Lakes Handbook 1. Limnology and Limnetic Ecology, Blackwell Science Ltd. Oxford, p. 251-308.
- PERES-NETO, P., LEGENDRE, P., DRAY, S. and BORCARD, D., 2006. Variation partitioning of species data matrices: estimation and comparison of fractions. Ecology, vol. 87, p. 2614-2625.
- RADAMBRASIL. Projeto RADAMBRASIL., 1976. Levantamento de Recursos Naturais, 10 (Folha SA.21 - Santarém). Departamento Nacional da Produção Mineral, Rio de Janeiro, 522 p.
- RANGEL, TFLVB., DINIZ-FILHO, JAF., BINI, and LM., 2006. SAM: a comprehensive application for Spatial Analysis in Macroecology. Ecography, vol. 31, no. 1, p. 46-50.
- RICHEY, JE., MERTES, LAK., DUNNE, T., VICTORIA, R., FORSBERG, BR., TANCREDI, ACFNS. and OLIVEIRA, E., 1989. Source and routing of the Amazon river flood wave. Global biogeochemical cycles, vol. 3, p. 191-204.
- ROQUE, FO., LECCI, L S., SIQUEIRA, T. and FROEHLICH, CG., 2008. Using environmental and spatial filters to explain stonefly occurrences in Southeastern Brazilian streams: implications for biomonitoring. Acta Limnologica Brasiliensia, vol. 20, p. 117-130.
- SHOUP, DE. and WAHL, DH., 2009. Fish Diversity and Abundance in Relation to Interannual and Lake-Specific Variation in Abiotic Characteristics of Floodplain Lakes of the Lower Kaskaskia River, Illinois. Transactions of the American Fisheries Society, vol.138, p.1076-1092.
- SIOLI, H., 1984. The Amazon and its main affluents: Hydrography morphology of the rivers course, and tivers types. In: SIOLI, H., (Ed.), The Amazon: limnology and landscape of a mighty tropical river and its basin Dordrecht, Dr,. W. Junk.., p. 127-165.
- SLOAN, WT., LUNN, M., WOODCOCK, S., HEAD, IM, NEE, S. and CURTIS, TP., 2006. Quantifying the roles of immigration and chance in shaping prokaryote community structure. Environmental Microbiology, vol. 8, p. 732-740.
- SOININEN, J., KOKOCINSKI, M., ESTLANDER, S., KOTANEN, J. and HEINO, J., 2007. Neutrality, niches, and determinants of plankton metacommunity structure across boreal wetland ponds. Ecoscience, vol. 14, no 2, p. 146-154.
- SØNDERGAARD, M., JEPPESEN, E., MORTENSEN, E., DALL, E., KRISTENSEN, P. and SORTKJÆR, O., 1990. Phytoplankton biomass reduction after planktivorous fish reduction in a shallow, eutrophic lake: A combined effect of reduced internal P-loading and increased zooplankton grazing. Hydrobiologia, vol. 200/201, p. 229-240.
- THOMAZ, SM., BINI, LM. and BOZELLI, RL., 2007. Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia, vol. 579, p. 1-13.
- THOMPSON, R. and TOWNSEND, C., 2006. A truce with neutral theory: local deterministic factors, species traits and dispersal limitation together determine patterns of diversity in stream invertebrates. Journal of Animal Ecology, vol. 75, p. 476-484.
- TOKESHI, M., 1999. Species Coexistence: Ecological and Evolutionary Perspectives Blackwell Publishing, Oxford. 454 p.
- VAN DEN HOECK, C., MANN, DG. and JAHNS, HM., 1993. Algae: an introduction to phycology. Cambridge University Press. Cambridge, 627 p.
- WILSON, DS., 1992. Complex interactions in metacommunities, with implications for biodiversity and higher levels of selection. Ecology, vol. 73, p. 1984-2000.
- YANG, H., ROGER, J.F. and BATTARBEE, RW., 2009. Influence of environmental and spatial variables on the distribution of surface sediment diatoms in an upland loch, Scotland. Acta Botanica Croatica, vol. 68 no.2, p. 367-380.
- YOUNG, NS., IOANNIDIS, JPA. and AL-UBAYDLI, O., 2008. Why Current Publication Practices May Distort Science. PLoS Medicine, vol.5, no. 10, e201.
Determinants of beta diversity: the relative importance of environmental and spatial processes in structuring phytoplankton communities in an Amazonian floodplain
Publication Dates
-
Publication in this collection
11 Apr 2011 -
Date of issue
Sept 2010
History
-
Accepted
27 Sept 2010 -
Received
05 Oct 2009