Abstracts
The aim of this study was to evaluate the foliar nutrient content in Eucalyptus benthamii mini-stumps treated with different Zn and B concentrations during successive shoot collections. Shoots were collected from mini-stumps fertigated with nutrient solutions containing different concentrations of Zn and B (S1 - free of Zn and B; S2 - 0.5 mg L-1 Zn; S3 - 0.5 mg L-1 B; S4 - 0.5 mg L-1 Zn and B; S5 - 1.0 mg L-1 Zn and B; S6 - 2.0 mg L-1 Zn and B). Eight shoot collections from the clonal mini-garden were performed. For each shoot collection, the nutritional content of the leaf was determined (N, P, K, Ca, Mg, S, B, Cu, Fe, Mn and Zn). The experiment was conducted using a randomized design with a split plot and included three replications. The foliar macro and micronutrient contents varied significantly in relation to the treatments, and the S5 and S6 nutritive solutions presented the best responses.
nutritive solution; fertigation; mini-cutting technique; clonal mini-garden
Objetivou-se avaliar os teores foliares nutricionais de minicepas de Eucalyptus benthamii em relação a diferentes concentrações de Zn e B ao longo de sucessivas coletas de brotações. As brotações foram oriundas de minicepas fertirrigadas com soluções nutritivas variando nas concentrações de Zn e B (S1 - isento de Zn e B, S2 - 0,5 mg L-1 de Zn, S3 - 0,5 mg L-1 de B, S4 - 0,5 mg L-1 de Zn e B, S5 - 1,0 mg L-1 de Zn e B, S6 - 2,0 mg L-1 de Zn e B), sendo realizadas oito coletas de brotações do minijardim clonal. Ao longo do experimento foram determinados o teor nutricional foliar de N, P, K, Ca, Mg, S, B, Cu, Fe, Mn e Zn. Considerou-se o delineamento inteiramente casualizado em esquema de parcelas subdivididas, com três repetições. Os teores foliares de macro e micronutrientes variaram significativamente em relação aos tratamentos avaliados, sendo que as soluções S5 e S6 apresentaram as melhores respostas.
solução nutritiva; fertirrigação; miniestaquia; minijardim clonal
PRODUÇÃO VEGETAL/CROP PRODUCTION
Nutritional content in Eucalyptus benthamii mini-stump leaves
Teor nutricional foliar em minicepas de Eucalyptus benthamii
Gilvano Ebling BrondaniI, * * Author for correspondence. E-mail: gebrondani@yahoo.com.br ; Francisco José Benedini BaccarinII; Antônio Natal GonçalvesII; Marcílio de AlmeidaIII
IFaculdade de Engenharia Florestal, Programa de Pós-graduação em Ciências Florestais e Ambientais, Universidade Federal de Mato Grosso, Avenida Fernando Correa da Costa, 2367, 78060-900, Cuiabá, Mato Grosso, Brazil
IILaboratório de Fisiologia das Árvores, Programa de Pós-graduação em Recursos Florestais, Escola Superior de Agricultura "Luiz de Queiroz", Universidade de São Paulo, Piracicaba, São Paulo, Brazil
IIILaboratório de Morfogênese e Biologia Reprodutiva de Plantas, Programa de Pós-graduação em Fisiologia e Bioquímica de Plantas, Escola Superior de Agricultura "Luiz de Queiroz", Universidade de São Paulo, Piracicaba, São Paulo, Brazil
ABSTRACT
The aim of this study was to evaluate the foliar nutrient content in Eucalyptus benthamii mini-stumps treated with different Zn and B concentrations during successive shoot collections. Shoots were collected from mini-stumps fertigated with nutrient solutions containing different concentrations of Zn and B (S1 - free of Zn and B; S2 - 0.5 mg L-1 Zn; S3 - 0.5 mg L-1 B; S4 - 0.5 mg L-1 Zn and B; S5 - 1.0 mg L-1 Zn and B; S6 - 2.0 mg L-1 Zn and B). Eight shoot collections from the clonal mini-garden were performed. For each shoot collection, the nutritional content of the leaf was determined (N, P, K, Ca, Mg, S, B, Cu, Fe, Mn and Zn). The experiment was conducted using a randomized design with a split plot and included three replications. The foliar macro and micronutrient contents varied significantly in relation to the treatments, and the S5 and S6 nutritive solutions presented the best responses.
Keywords: nutritive solution, fertigation, mini-cutting technique, clonal mini-garden.
RESUMO
Objetivou-se avaliar os teores foliares nutricionais de minicepas de Eucalyptus benthamii em relação a diferentes concentrações de Zn e B ao longo de sucessivas coletas de brotações. As brotações foram oriundas de minicepas fertirrigadas com soluções nutritivas variando nas concentrações de Zn e B (S1 - isento de Zn e B, S2 - 0,5 mg L-1 de Zn, S3 - 0,5 mg L-1 de B, S4 - 0,5 mg L-1 de Zn e B, S5 - 1,0 mg L-1 de Zn e B, S6 - 2,0 mg L-1 de Zn e B), sendo realizadas oito coletas de brotações do minijardim clonal. Ao longo do experimento foram determinados o teor nutricional foliar de N, P, K, Ca, Mg, S, B, Cu, Fe, Mn e Zn. Considerou-se o delineamento inteiramente casualizado em esquema de parcelas subdivididas, com três repetições. Os teores foliares de macro e micronutrientes variaram significativamente em relação aos tratamentos avaliados, sendo que as soluções S5 e S6 apresentaram as melhores respostas.
Palavras-chave: solução nutritiva, fertirrigação, miniestaquia, minijardim clonal.
Introduction
Eucalyptus benthamii is an alternative component of forest plantations for industrial purposes. However, because this species was recently introduced to forest plantations, seed production is limited, and when seeds are available, the prices are high. Vegetative propagation is an alternative for reproduction of the species and included pre-selected matrices for each region. However, adventitious rooting of E. benthamii is low for the cutting technique (GRAÇA et al., 1999).
Some progress can be achieved by adjusting the nutritional content of clonal mini-gardens (HARTMANN et al., 2011), which may lead to the production of shoots with a predisposition to adventitious rooting (ALFENAS et al., 2004; SCHWAMBACH et al., 2008; WENDLING et al., 2010). The nutritional status of the mother plant used as a shoot donor (mini-stump) for mini-cuttings can significantly influence the process of rhizogenesis (HARTMANN et al., 2011).
In practical terms, semi-hydroponic mini-gardens on sand beds or in conical plastic tubes are used more often in Brazil (HIGASHI et al., 2005). In this system, nutrients are supplied via fertirrigation in adequate quantities to support the growth and development of the plant and may result in increased adventitious rooting. However, the optimal concentrations of nutrients have not been established for Eucalyptus spp. mini-stumps (ALFENAS et al., 2004) considering the influences of shoot collections and their interactions with the environment (e.g., temperature, light, photoperiod and relative humidity) during different seasons (BHARDWAJ; MISHRA, 2005; TRUEMAN et al., 2012). Although the nutrients needed for development are the same, the quantities required are different between and within species (different clones), and there is no standard nutrient solution (HIGASHI et al., 2005), which highlights the need for more detailed studies.
Considering the theoretical rules, mineral nutrition can influence the adventitious rooting of propagules in two ways: (i) due to the vigor of the mother plant shoot donor and (ii) the proper nutritional status of the collected material (shoot and sprout). According to Alfenas et al. (2004), nutritional problems in clonal mini-gardens may be related to a deficiency or excess of elements. In a way, any nutrient involved in the metabolic processes associated with adventitious root differentiation is considered essential for rhizogenic initiation (CUNHA et al., 2009a; TRUEMAN et al., 2012). Among the micro-elements that affect adventitious rooting, zinc (Zn) and boron (B) are highlighted. Several studies have reported increased adventitious rooting in propagules when subjected to various forms and concentrations of Zn and B (KERSTEN et al., 1993; MALTA et al., 2002) considering that these nutrients are essential for plant growth and development (TAIZ; ZEIGER, 2009). However, there are no reports in the literature concerning the effects of these nutrients (Zn and B) in terms of the nutritional quality of shoots collected from Eucalyptus benthamii mini-stumps to be used as mini-cuttings.
The aim of the present study was to determine the nutritional content of leaves (N, P, K, Ca, Mg, S, B, Cu, Fe, Mn and Zn) from Eucalyptus benthamii mini-stumps from different shoot collections and with varying Zn and B concentrations.
Material and methods
Constitution of the clonal mini-garden
The experimental units were composed of Eucalyptus benthamii Maiden and Cambage clones propagated through mini-cutting techniques with the application of 2,000 mg L-1 IBA (indole-3-butyric acid) and cultivated in plastic pots (18 x 16 x 45 cm) with two openings in the lower portion (Figure 1A). The substrate was composed of sand (0.10 mm < particle diameter < 0.25 mm), which was sieved (mesh sieve of 3 mm), washed with deionized water five times and autoclaved for 20 min. at 121°C (≈ 1.0 kg cm-2) to eradicate weeds and pathogens, such as fungi and bacteria. Prior to planting the clones in the plastic pots, the roots were washed with deionized water for 10 min. to leach nutrients and other components. The pots were suspended by metal bars (85 cm of soil), and six clones were planted per pot in a 10 x 8 cm space, consisting of six seedlings arranged sequentially. After seven days, the stems of the clones were broken at 10 cm above the basal portion to reduce the apical dominance and favor the growth of axillary buds, which were issued 21 days after the stems were broken (Figure 1B). To form the mini-stumps, the shoots were pruned to 7 cm above the base of the clone 15 days after the stem was broken, and the clonal mini-garden was formed in a pot system (Figure 1C and D).

Management of mini-stumps
The clonal mini-garden was installed in a greenhouse covered with transparent plastic and with two fan systems. Air temperature data were collected daily to investigate a possible correlation with the response variables. Every seven days, 100 mL of basic nutrient solution was added to the mini-stump (Table 1). Irrigation was performed with deionized water daily to find excessive loss by evapotranspiration. Each week, the sand was washed with deionized water to prevent excess salination.
Treatments and experimental design
Various concentrations of zinc (Zn) and boron (B) were provided by nutritive solution via fertigation (Table 2). Every seven days, 100 mL of the nutrient solution (Tables 1 and 2) was added per mini-stump (Figure 1C and D). The experiment was conducted using a randomized design with an arrangement split plot in time. There were eight shoot collections (conducted for 12 months and performed through eight successive shoot collections at intervals ranging from 21 to 35 days) and six combinations of Zn and B concentrations (Table 2), which were tested in triplicate.
Nutrient content of leaves
The samples were composed of fully expanded leaves from the upper third of the shoot. After the collection, all the leaves were washed with deionized water and then dried at 60°C until they reached a constant weight. The leaves were then ground into a powder. Macro and micronutrients were determined according to the method described by Malavolta et al. (1989). Total N was determined by digestion with sulfuric acid (Semi-Micro Kjeldahl). The P, S, K, Ca, Mg, Fe, Cu, Zn and Mn contents were determined by digestion with nitric-perchloric acid. The P content was determined by colorimetry with ammonium metavanadate, and the S content was determined by turbidimetry of barium sulphate with a spectrophotometer. The K content was determined by atomic absorption spectrophotometry in a flame photometer with a K hollow lamp cathode. The Ca, Mg, Fe, Cu, Zn and Mn contents were determined by atomic absorption spectrophotometry. The B content was determined by digesting the dry matter by burning it in an alkaline medium using azomethine colorimetry, followed by measurement in a spectrophotometer.
Statistical analysis
The data measured in all experiments were analyzed by the Hartley test (p < 0.05) and Lilliefors test (p < 0.05) and transformed when necessary. Then, analysis of variance (ANOVA, p < 0.01 and p < 0.05) was performed. In accordance with the significance of the ANOVA, the data of qualitative factors were compared by Tukey's test (p < 0.05). Pearson correlation analysis (p < 0.01 and p < 0.05) between the environmental data and the measured variables was performed. The packages SOC (EMBRAPA, 1990) and statistical program (MSTAT, 1994) were used for statistical analysis.
Results and discussion
The nutritional content of the leaves of Eucalyptus benthamii mini-stumps varied significantly between the shoot collections and nutrient solutions (p < 0.01). In the 8th shoot collection, there was a significant difference in N content between the S2 (27.8 g kg-1) and S6 (22.2 g kg-1) solutions. The foliar N content ranged from 34.0 to 17.6 g kg-1 over the eight shoot collections (Table 3).
The highest P content occurred in the 3rd, 5th, 6th and 7th shoot collections. In general, the foliar P content varied from 3.4 to 1.8 g kg-1 over the eight shoot collections (Table 3). The lowest values for K content were recorded in the 1st, 2nd, 3rd, and 4th shoot collections. In the 8th shoot collection, the nutrient solution S1 presented the lowest foliar K content in relation to the nutrient solutions S3, S4, S5, and S6. The foliar K content ranged from 22.1 to 7.1 g kg-1 over the eight shoot collections (Table 3). In the 1st, 3rd, and 8th shoot collections, the foliar Ca content was the highest. In general, the foliar Ca content ranged from 6.9 to 3.0 g kg-1 over the eight shoot collections (Table 3). The foliar Mg content ranged from 2.3 to 1.5 g kg-1 over the eight shoot collections and presented little variation (Table 3). The foliar S content was higher in the 1st, 2nd and 3rd shoot collections and then showed a decrease from the 4th collection onward. The foliar S content ranged from 3.2 to 1.5 g kg-1 over the eight shoot collections (Table 3).
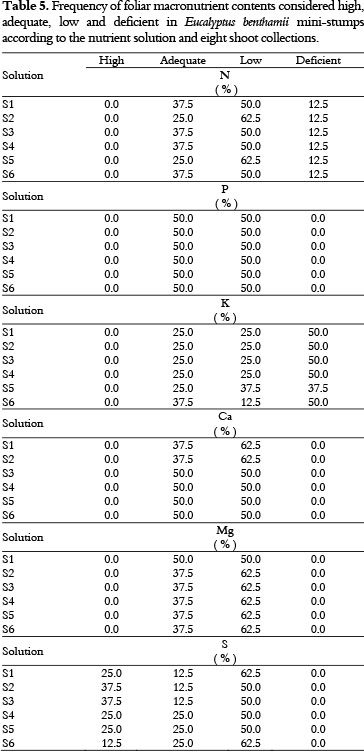
Based on the interpretation of Table 4 and the significant variation in the foliar micronutrient contents of Eucalyptus benthamii mini-stumps in relation to the nutrient solutions and shoot collections, only 12.5% of the leaves were considered N-deficient (i.e., only in the 1st shoot collection) (Table 5). The other values for foliar N content were classified as adequate and low for all nutrient solutions (Table 5). The foliar P, Ca, and Mg contents were considered adequate and low. The foliar S content has not presented deficiency, and the values were high, adequate and low (Table 5). Approximately 50% of the foliar K contents measured from the 1st to the 4th shoot collection were deficient. The remaining measured foliar K contents were adequate and low (i.e., 5th to 8th shoots collection) (Table 5).
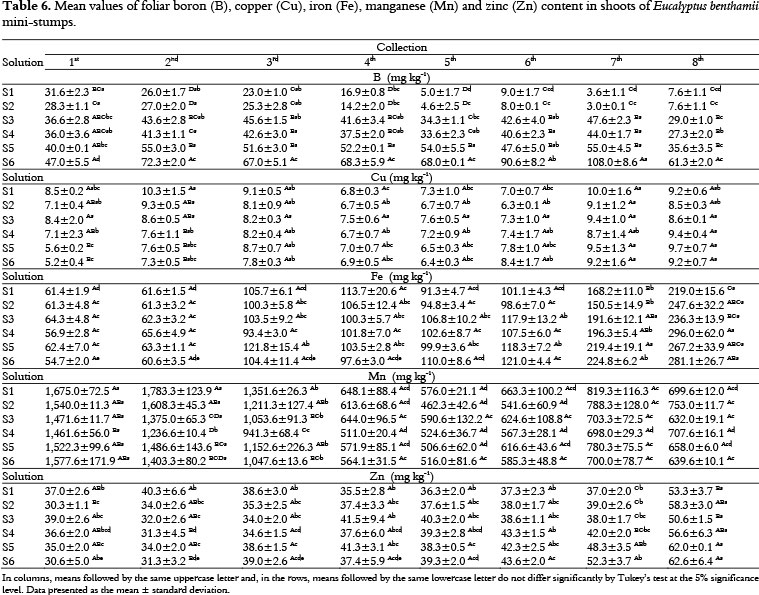
The leaf B content showed variation during the experimental period and the lowest accumulations were measured for the S1 and S2 solutions (i.e., free of B) (Table 6). The leaf B content ranged from 108.0 to 3.0 mg kg-1 over the eight shoot collections (Table 6) and symptoms of nutritional deficiency were observed. The lowest values of leaf Cu content occurred in the 4th and 5th shoot collections. The leaf Cu content ranged from 10.3 to 5.2 mg kg-1 over the eight shoot collections (Table 6). The foliar Fe content increased until the 8th shoot collection; the highest Fe values were recorded in the 7th and 8th collection. The leaf Fe content ranged from 296.0 to 54.7 mg kg-1 over the eight shoot collections (Table 6). The foliar Mn content decreased through the 8th shoot collection, and the highest accumulations were recorded in the 1st, 2nd and 3rd collections. The foliar Mn content ranged from 1,783.3 to 462.3 mg kg-1 over the eight shoot collections (Table 6). The foliar Zn content ranged from 62.6 to 30.3 mg kg-1 over the eight shoot collections (Table 6).
Considering the S1 and S2 solutions (i.e., free of B), 62.5% of the B concentrations were rated as deficient (5th at 8th shoot collections), and 37.5% were categorized as low (1st at 4th shoot collections), which shows the Eucalyptus benthamii mini-stumps strong response to treatments that did not include B (Table 7). The foliar B content of the mini-stumps that received the S3 (0.5 mg L-1 B) and S4 (0.5 mg L-1 Zn and B) solutions were classified as adequate and low.
The mini-stumps that received the S5 (1.0 mg L-1 Zn and B) solution presented adequate B concentrations in all shoot collections. However, mini-stumps that were fertigated with the S6 (2 mg L-1 Zn and B) solution presented B concentrations in the adequate and high range, which may cause toxicity (Table 7). The foliar Cu content values were classified as adequate and low and the Mn contents were between high and adequate. Despite the variations in the Zn concentrations recorded for the different nutrient solutions that were evaluated, Zn was the only nutrient that presented adequate foliar content for 100% of the shoot collections (Table 7). Iron was the micronutrient that presented the greatest variation in foliar content among the different shoot collections; iron content values were classified as high, adequate, low and deficient (Table 7). Thus, Fe requires additional attention when managing the nutrient solutions used for the fertigation of Eucalyptus benthamii clonal mini-gardens, and this management should take into account the successive shoots collections throughout the year as well as temperature variations by season.
Analysis of the Pearson Correlation Matrix verifies a high correlation between maximum (TMAX), medium (TMED) and minimum (TMIN) air temperature and foliar N, P, K, Mg, S, Fe and Mn contents (Table 8). The higher the temperature recorded inside the greenhouse containing the Eucalyptus benthamii clonal mini-garden, the lower the leaf N, P, K and Fe content (i.e., the correlation was negative). However, foliar Mg, S and Mn content increased with increasing temperature, showing a positive correlation (Table 8). There was a positive correlation, with the highest value of 0.60, for P*N, Mg*Ca, Fe*K and Mn*S, which indicates increased foliar content of these nutrients. However, there was a negative correlation, with a value at -0.60, for Mn*N, which indicates that the increased foliar content of one nutrient (Mn or N) was followed by the reduction of the other nutrients (Table 8).
The nutritional status of donor plant shoots is one of the determining factors for vegetative propagation success through adventitious rooting (CUNHA et al., 2009c; HARTMANN et al., 2011). The use of balanced nutrient solutions in clonal mini-garden systems ensures the nutritional equilibrium of mini-stumps. However, ideal nutrient concentration patterns have not been established for Eucalyptus mini-stumps (ALFENAS et al., 2004), and adjustments are required in accordance with the management conditions and genetic material.
N concentrations were classified as deficient in 12.5% of the observations (i.e., during the summer), regardless of the nutrient solution. This result indicates that the N concentration in the basic nutrient solution may be increased according to the nutritional needs of the genetic material and the time of year. N is a component of amino acids, amines, amides, nitrogenous bases, alkalis, chlorophylls and many coenzymes (TAIZ; ZEIGER, 2009) and is correlated with adventitious rooting (CUNHA et al., 2009c). The deficiency in Eucalyptus leaves is characterized by gradual and generalized chlorosis of older leaves and etiolation (DELL et al., 1995)
The influence of N on the production system of Eucalyptus mini-cuttings has been reported in several studies and controlling N concentrations in nutritive solutions requires great care. Rosa et al. (2009) observed that N significantly influenced the productivity of shoots in Eucalyptus dunnii mini-stumps; N is essential for adequate plant metabolism and therefore affects the quantity and quality of shoot production. Carvalho Neto et al. (2011) verified a significant effect of a fertigation solution's N concentration on dry matter in Eucalyptus urophylla mini-cuttings; they observed a linear decrease in dry matter with increasing N concentrations. Cunha et al. (2008) observed a positive correlation between nitrogen content and the number of mini-cuttings in a sand bed clonal mini-garden of Eucalyptus grandis × Eucalyptus urophylla, which indicates that it is necessary to increase the availability of nitrogen to the genetic material to increase the mini-cuttings' productivity. Nevertheless, Higashi et al. (2005) commented that the excessive increase of the nutrient solutions N concentrations can reduce the concentrations of B, Cu, Fe, Mn, and Zn among a clonal mini-garden of Eucalyptus and demonstrated the importance of knowing the critical levels and nutrient interactions for each genetic material.
Among the observations for all nutrient solutions, 50% the K content was deficient (Table 5), which suggests that it may be necessary to increase the K concentration in the nutrient solution (Table 1). The element K activates many enzymes and is involved in stomatal control and carbohydrate transport (TAIZ; ZEIGER, 2009); the K participates in three inter-related functions: (i) transport through the membrane, (ii) neutralization of anions and (iii) control of the osmotic potential (DELL et al., 1995). A K deficiency is characterized by green leaves or blue-green leaves with marginal necrosis and wilting and may cause lateral bud and terminal mortality under severe conditions (DELL et al., 1995).
Cunha et al. (2008) reported a positive correlation of the K foliar content for a Eucalyptus grandis hybrid (clone 57) and for a genotype of Eucalyptus grandis (clone 7074) and the adequate and ideal K concentrations proposed in this study were not the same between the different clones. However, in the same study, the authors found negative effects of the nutrient solutions' K concentrations on a clone of Eucalyptus grandis × Eucalyptus urophylla and recommended reducing the fertigation solution's K concentration. These effects may be associated with the high mobility of K (TAIZ; ZEIGER, 2009), which is easily absorbed and redistributed in plant tissues when it is available (DELL et al., 1995), which may also have influenced the growth and development of Eucalyptus benthamii mini-stumps.
Nutritional deficiencies in Fe were present for all nutrient solutions, and 25% of the total observations were classified as deficient, which suggests that it is necessary to increase the nutrient solutions' Fe concentration according to the amount required by the genetic material and the time of year. The Fe activates enzymes and participates in oxidation-reduction reactions in homeoproteins (cytochromes, leghemoglobin, catalase, peroxidase and superoxide dismutase) and non-hemic proteins with F-S binding, such as ferredoxin and reductase enzymes, nitrogenase and sulfate reductase. In addition, Fe catalyzes chlorophyll biosynthesis (TAIZ; ZEIGER, 2009). An iron deficiency is characterized by the inability of young leaves to synthesize chlorophyll; the leaves show interveinal paling in which only the conductive vessels remain green, fine and curved stems, necrotic areas on the edges of the limbus, premature falling of leaves and reduction in growth (DELL et al., 1995).
Cunha et al. (2008) reported negative correlations between Fe content and shoot production in Eucalyptus grandis × Eucalyptus urophylla mini-stumps and for Eucalyptus grandis in experiments conducted with clonal mini-gardens in sand beds. Similar to the present study's observations for Eucalyptus benthamii mini-stumps, Cunha et al. (2008) observed variations in the frequency of cases where the Fe concentration was considered adequate, low and deficient; this fact indicates the need for further studies related to Fe nutrition in Eucalyptus clonal mini-gardens. These studies are also essential to generate further knowledge about the nutritional quality of shoots to be used as mini-cuttings.
With regard to the omission of Zn and B in the nutrient solutions, B deficiency symptoms were more frequent in Eucalyptus benthamii mini-stumps, which demonstrated the efficiency of the methodology applied to study the B's effects. The Zn nutritional content was not affected by the different nutrient solutions, the foliar content was classified as adequate in all treatments (Table 7), which is similar to what was reported by Sgarbi et al. (1999) for Eucalyptus grandis × Eucalyptus urophylla. According to the literature, B deficiency in Eucalyptus is often characterized by death of apical meristems followed by over budding of the lateral buds, deformed leaves of reduced size, marginal chlorosis and the presence of salient veins in the middle of the intermediate leaves (SGARBI et al., 1999; SILVEIRA et al., 1998; SILVEIRA et al., 2000; SILVEIRA et al., 2002). These symptoms were observed in mini-stumps subjected to treatment with the S1 (omission of Zn and B) and S2 (omission of B) solutions from the 5th shoot collection.
The Zn content of the water used to formulate the nutrient solution is another important factor to consider and, in this case, was 0.01 mg L-1. The daily irrigations (ranging from 600 to 1,000 mL per pot according to the season) and weekly washings to reduce the salinity in the substrate may have supplied additional Zn for the mini-stumps, although this element was present in low concentrations in the water. This factor may have influenced the lack of Zn deficiency symptoms (DELL et al., 1995) in the Eucalyptus benthamii mini-stumps. Future experiments using the same type of management system adopted in this study should consider these factors and control them more precisely to study the effect of Zn on mini-stump growth characteristics, especially in relation to substrate type and irrigation water and fertigation purity control (e.g., hydroponic systems).
Another important effect in relation to nutritional aspects includes the high correlations between N (< -0.76), K (< -0.78) and Fe (< -0.61) and maximum, mean and minimum air temperatures in the greenhouse (Table 8). This observation indicates that the increase in temperature may have induced deficiencies of these nutrients in Eucalyptus benthamii mini-stumps. The effect of temperature on the nutritional characteristics of shoots has been reported by several studies (HUANG et al., 1999; RAMAGE; WILLIAMS, 2002; SCHWAMBACH et al., 2005; TRUEMAN et al., 2012), including studies on Eucalyptus clonal mini-garden systems (ALFENAS et al., 2004; CUNHA et al., 2009a). Temperature influences the mobility of nutrients in the nutrient solution, absorption sites of the plasma membrane, absorption speed, translocation and redistribution in the tissues (DELL et al., 1995) and interferes with enzymatic activity (TAIZ; ZEIGER, 2009). These observations emphasize that the air temperature in the Eucalyptus benthamii clone production system should be more tightly controlled in consideration of the subtropical species' characteristics, especially during the warmer seasons (BRONDANI et al., 2010; BRONDANI et al., 2012; SCHWAMBACH et al., 005; TRUEMAN et al., 2012).
Conclusion
The foliar macro and micronutrient contents varied significantly according to the shoot collections and nutrient solutions.
The foliar B, N, K, and Fe contents were the most affected during the mini-stump shoot collections.
In general, the S5 (1.0 mg L-1 Zn and B) and S6 (2.0 mg L-1 Zn and B) solutions favored the foliar accumulation of macro and micronutrients in Eucalyptus benthamii mini-stumps.
Acknowledgements
We thank FAPESP (São Paulo Research Foundation, Brazil) and CAPES (Coordination for the Improvement of Higher Level, Brazil) for their financial support. We also thank Planflora for providing selected genotypes of Eucalyptus benthamii. Finally, we are grateful for the technical support provided by José Roberto Romanini.
Received on April 16, 2012
Accepted on June 25, 2012
License information: This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
- ALFENAS, A. C.; ZAUZA, E. A. V.; MAFIA, R. G.; ASSIS, T. F. Clonagem e doenças do eucalipto Viçosa: UFV, 2004.
- BHARDWAJ, D. R.; MISHRA, V. K. Vegetative propagation of Ulmus villosa: effects of plant growth regulators, collection time, type of donor and position of shoot on adventitious root formation in stem cuttings. New Forests, v. 29, n. 2, p. 105-116, 2005.
- BRONDANI, G. E.; GROSSI, F.; WENDLING, I.; DUTRA, L. F.; ARAUJO, M. A. Aplicação de IBA para o enraizamento de miniestacas de Eucalyptus benthamii Maiden and Cambage × Eucalyptus dunnii Maiden. Acta Scientiarum. Agronomy, v. 32, n. 4, p. 667-674, 2010.
- BRONDANI, G. E.; WENDLING, I.; BRONDANI, A. E.; ARAUJO, M. A.; SILVA, A. L. L.; GONÇALVES, A. N. Dynamics of adventitious rooting in mini-cuttings of Eucalyptus benthamii × Eucalyptus dunnii Acta Scientiarum. Agronomy, v. 34, n. 2, p. 169-178, 2012.
- CARVALHO NETO, J. P.; SILVA, E. B.; SANTANA, R. C.; GRAZZIOTTI, P. H. Effect of NPK fertilization on production and leaf nutrient content of Eucalyptus minicuttings in nutrient solution. Revista Brasileira de Ciência do Solo, v. 35, n. 1, p. 249-254, 2011.
- CUNHA, A. C. M. C. M.; PAIVA, H. N.; BARROS, N. F.; LEITE, H. G.; LEITE, F. P. Relação do estado nutricional de miniestacas com o número de miniestacas de eucalipto. Scientia Forestalis, v. 36, n. 79, p. 203-213, 2008.
- CUNHA, A. C. M. C. M.; PAIVA, H. N.; LEITE, H. G.; BARROS, N. F.; LEITE, F. P. Relações entre variáveis climáticas com produção e enraizamento de miniestacas de eucalipto. Revista Árvore, v. 33, n. 2, p. 195-203, 2009a.
- CUNHA, A. C. M. C. M.; PAIVA, H. N.; LEITE, H. G.; BARROS, N. F.; LEITE, F. P. Influência do estado nutricional de minicepas no enraizamento de miniestacas de eucalipto. Revista Árvore, v. 33, n. 4, p. 607-615, 2009b.
- CUNHA, A. C. M. C. M.; PAIVA, H. N.; BARROS, N. F.; LEITE, H. G.; LEITE, F. P. Relação do estado nutricional de minicepas com o enraizamento de miniestacas de eucalipto. Revista Brasileira de Ciência do Solo, v. 33, n. 3, p. 591-599, 2009c.
- DELL, B.; MALAJCZUK, N.; GROVE, T. S. Nutrient disorders in plantation eucalypts Canberra: Australian Centre for International Agricultural Research, 1995.
- EMBRAPA-Empresa Brasileira de Pesquisa Agropecuária. Programa SOC - software científico: versão 2.1. Campinas: Embrapa Informática Agropecuária, 1990.
- GRAÇA, M. E. C.; SHIMIZU, J. Y.; TAVARES, F. R. Capacidade de rebrota e de enraizamento de Eucalyptus benthamii Boletim de Pesquisa Florestal, n. 39, p. 135-138, 1999.
- HARTMANN, H. T.; KESTER, D. E.; DAVIES JR., F. T.; GENEVE, R. L. Plant propagation: principles and practices. 8th ed. São Paulo: Prentice-Hall, 2011.
- HIGASHI, E. N.; SILVEIRA, R. L. V. A.; GONÇALVES, A. N. Monitoramento nutricional e fertilização em macro, mini e microjardim clonal de Eucalyptus In: GONÇALVES, J. L. M.; BENEDETTI, V. (Ed.). Nutrição e fertilização florestal Piracicaba: IPEF, 2005. p. 191-217.
- HUANG, L.; BELL, R. W.; DELL, B. Factors controlling equilibrium boron (B) concentration in nutrient solution buffered with B-specific resin (Amberlite IRA-743). Plant and Soil, v. 208, n. 2, p. 233-241, 1999.
- KERSTEN, E.; LUCCHESI, A. A.; GUTIERREZ, L. E. Efeitos do boro e zinco no teor de carboidratos solúveis, aminoácidos totais e no enraizamento de estacas de ramos de ameixeira (Prunus salicina Lindl.). Scientia Agricola, v. 50, n. 1, p. 13-18, 1993.
- MALAVOLTA, E.; VITTI, G. C.; OLIVEIRA, S. A. Avaliação do estado nutricional das plantas: princípios e aplicações. Piracicaba: Potafos, 1989.
- MALTA, M. R.; FURTINI NETO, A. E.; ALVES, J. D.; GUIMARÃES, P. T. G. Efeito da aplicação de zinco via foliar na síntese de triptofano, aminoácidos e proteínas solúveis em mudas de cafeeiro. Brazilian Journal of Plant Physiology, v. 14, n. 1, p. 31-37, 2002.
- MSTAT. Statistical program Michigan: Michigan State University, Crop and Soil Department Science, 1994.
- RAMAGE, C. M.; WILLIAMS, R. R. Mineral nutrition and plant morphogenesis. In Vitro Cellular and Developmental Biology - Plant, v. 38, n. 2, p. 116-124, 2002.
- ROSA, L. S.; WENDLING, I.; GROSSI, F.; REISSMANN, C. B. Efeito da dose de nitrogênio e de formulações de substratos na miniestaquia de Eucalyptus dunnii Maiden. Revista Árvore, v. 33, n. 6, p. 1025-1035, 2009.
- SCHWAMBACH, J.; FADANELLI, C.; FETT-NETO, A. G. Mineral nutrition and adventitious rooting in microcuttings of Eucalyptus globulus Tree Physiology, v. 25, n. 4, p. 487-494, 2005.
- SCHWAMBACH, J.; RUEDELL, C. M.; ALMEIDA, M. R.; PENCHEL, R. M.; ARAÚJO, E. F.; FETT-NETO, A. G. Adventitious rooting of Eucalyptus globulus × maidennii mini-cuttings derived from mini-stumps grown in sand bed and intermittent flooding trays: a comparative study. New Forests, v. 36, n. 3, p. 261-271, 2008.
- SGARBI, F.; SILVEIRA, R. L. V. A.; TAKAHASHI, E. N.; CAMARGO, M. A. F. Crescimento e produção de biomassa de clone de Eucalyptus grandis × Eucalyptus urophylla em condições de deficiência de macronutrientes, B e Zn. Scientia Forestalis, n. 56, p. 69-82, 1999.
- SILVEIRA, R. L. V. A.; GONÇALVES, A. N.; KRÜGNER, T. L. Estado nutricional de Eucalyptus citriodora Hook cultivado sob diferentes doses de boro e sua relação com a agressividade de Botryosphaeria ribis Scientia Forestalis, n. 53, p. 57-70, 1998.
- SILVEIRA, R. L. V. A.; MOREIRA, A.; TAKASHI, E. N.; SGARBI, F.; BRANCO, E. F. Sintomas de deficiência de macronutrientes e de boro em clones híbridos de Eucalyptus grandis com Eucalyptus urophylla Cerne, v. 8, n. 2, p. 107-116, 2002.
- SILVEIRA, R. L. V. A.; TAKAHASHI, E. N.; SGARBI, F.; CAMARGO, M. A. F.; MOREIRA, A. Crescimento e estado nutricional de brotações de Eucalyptus citriodora sob doses de boro em solução nutritiva. Scientia Forestalis, n. 57, p. 53-67, 2000.
- TAIZ, L.; ZEIGER, E. Fisiologia vegetal 4. ed. Porto Alegre: Artmed, 2009.
- TRUEMAN, S. J.; MCMAHON, T. V.; BRISTOW, M. Production of cuttings in response to stock plant temperature in the subtropical eucalypts, Corymbia citriodora and Eucalyptus dunnii New Forests, v. 44, n. 2, p. 265-279, 2012.
- WENDLING, I.; BRONDANI, G. E.; DUTRA, L. F.; HANSEL, F. A. Mini-cuttings technique: a new ex vitro method for clonal propagation of sweetgum. New Forests, v. 39, n. 3, p. 343-353, 2010.
Publication Dates
-
Publication in this collection
07 Oct 2014 -
Date of issue
Dec 2014
History
-
Accepted
25 June 2012 -
Received
16 Apr 2012