Abstract
Penicillum janthinellum SDX7 was isolated from aged petroleum hydrocarbon-affected soil at the site of Anand, Gujarat, India, and was tested for different pH, temperature, agitation and concentrations for optimal growth of the isolate that was capable of degrading upto 95%, 63% and 58% of 1%, 3% and 5% kerosene, respectively, after a period of 16 days, at optimal growth conditions of pH 6.0, 30 °C and 180 rpm agitation. The GC/MS chromatograms revealed that then-alkane fractions are easily degraded; however, the rate might be lower for branched alkanes, n-alkylaromatics, cyclic alkanes and polynuclear aromatics. The test doses caused a concentration-dependent depletion of carbohydrates of P. janthinellum SDX7 by 3% to 80%, proteins by 4% to 81% and amino acids by 8% to 95% upto 16 days of treatment. The optimal concentration of 3% kerosene resulted in the least reduction of the metabolites of P. janthinellum such as carbohydrates, proteins and amino acids with optimal growth compared to 5% and 1% (v/v) kerosene doses on the 12th and 16th day of exposure. Phenols were found to be mounted by 43% to 66% at lower and higher concentrations during the experimental period. Fungal isolate P. janthinellum SDX7 was also tested for growth on various xenobiotic compounds.
Penicillum janthinellum SDX7; optimal growth; metabolites; kerosene degradation
Introduction
Kerosene currently has several uses such as aircraft gas turbine and jet fuel for both commercial airlines and the military activities, as heating oil, and as a spray oil to combat insects on agricultural plants. Because of its availability compared to gasoline during wartime, commercial illuminating kerosene was the fuel chosen for the early jet engines. Consequently, the development of commercial jet aircraft following World War II focused primarily on the use of kerosene-type fuels (Irwin et al., 1997Irwin RJ, Van Mouwerik M, Stevens L et al. (1997) Environmental Contaminants Encyclopedia. Kerosene Entry. National Park Service, Water Resources Division, Water Operating Branch. Oakridge Drive, Suite Fort Collins, 42 p.). According to the US Coast Guard Emergency Response Notification System, kerosene is one of the most commonly spilled petroleum products containing paraffins (alkanes), cycloparaffins (cycloalkanes), aromatics, and olefins with carbon numbers betweenC9–C20 (Irwin et al., 1997Irwin RJ, Van Mouwerik M, Stevens L et al. (1997) Environmental Contaminants Encyclopedia. Kerosene Entry. National Park Service, Water Resources Division, Water Operating Branch. Oakridge Drive, Suite Fort Collins, 42 p.). Large amounts of spills and leaks of petroleum products such as gasoline, diesel, kerosene, and similar compounds have been refined and handled on land every year. Despite careful handling, there is a possibility of introduction into the soil environment. The penetration of hydrocarbons from the top of the soil into the subsoil leads to a direct risk of ground water contamination (Schinner and Margesin, 1997Margesin R, Schinner F (1997) Bioremediation of diesel-oil contaminated alpine soils at low temperatures. Appl Microbiol Biot 47:462–468.; Watkinson and Morgan, 1989Morgan P, Watkinson RJ (1989) Hydrocarbon degradation and methods for soil biotreatment. CRC Crit Rev Biotechnol 8:305–333.). Although a significant proportion of the compounds in petroleum products are relatively innocuous, a number of lower molecular weight compounds are toxic or mutagenic and require remedial action to restrict environmental damage (Greer et al., 2003Greer CW, Fortin N, Roy R et al. (2003) Indigenous sediment microbial activity in response to nutrient enrichment and plant growth following a controlled oil spill on a fresh water wetland. Bioremed J 7:69–80.)
Microorganisms are powerful alternatives to conventional methods in resolving environmental problems (Bento et al., 2003Bento FM, Camargo FA, Okeke B et al. (2003) Bioremediation of soil contaminated by diesel oil. Braz J Microbiol 34:65–68.). Bacteria, yeast and filamentous fungi have been reported as transforming agents due to their ability to degrade a wide range of pollutants because of their ubiquitous nature. Various bacterial genera that have been reported to contain hydrocarbon degrading species include Pseudomonas, Vibrio, Arthrobacter and Bacillus (Zajic, 1972Zajic E, Supplisson B (1972) Emulsification and degradation of "Bunker C" fuel oil by microorganisms. Biotechnol Bioeng 14:31–43.; Snape et al., 2001Snape I, Riddle MJ, Stark JS et al. (2001) Management andremediation of contaminated sites at Casey station. Antart Polar Rec 37:199–214.). Fungal degradation of defined single hydrocarbons as well as petroleum products has been performed by Penicillium, Candida, Fusarium, Aspergillus, Articulosporium (Sugiura et al., 1997Sugiura K, Ishihara M, Shimauchi T et al. (1997) Physicochemical properties and biodegradability of crude oil. Environ Sci Technol 31:45–51.; Mukherji et al., 2004Mukherji S, Jagadevan S, Mohapatra G et al. (2004) Biodegradation of diesel oil by an Arabian Sea sediment culture isolated from the vicinity of an oil field. Bioresour Technol 95:281–286.). Kumar et al. (2013)Nirmal Kumar JI, Megha B, Shamiyan RK (2013) Some intermediate bio-transformants during biodegradation of high molecular weight phenanthrene and fluoranthene by cyanobacterial species - Aulosira fertilissima ghose. Int J App Sci Biotechnol 1:97–105. performed biodegradation of high molecular weight PAHs such asphenanthrene and fluoranthene using cyanobacterial species - Aulosira fertilissima ghose. These organisms have been isolated in large numbers from many oil-polluted waters and soils but are found in lesser numbers in uncontaminated environments (Okoh, 2003Okoh AI (2003) Biodegradation of bonny light crude oil in soil microcosm by some bacterial strains isolated from crude flow stations saver pits in Nigeria. Afr J Biotechnol 2:104–108.). Biodegradation is considered to be a major mechanism for the removal of spilled petroleum hydrocarbons from the environment (Braddock and Lindstrom, 2002Lindstrom JE, Braddock JF (2002) Biodegradation of petroleum hydrocarbons at low temperature in the presence of the dispersant Corexit 9500. Mar Pollut Bull 44:739–747.). Therefore, it is necessary to perform laboratory feasibility tests to determine the effectiveness of biodegradation of kerosene due to the significant effect of the inherent capabilities of the microorganisms, by their ability to overcome the bioavailability limitations in multiphase environmental scenarios and by environmental factors such as pH, temperature, nutrients and electron acceptor availability (Mukherji et al., 2004Mukherji S, Jagadevan S, Mohapatra G et al. (2004) Biodegradation of diesel oil by an Arabian Sea sediment culture isolated from the vicinity of an oil field. Bioresour Technol 95:281–286.; Trindade et al., 2005Trindade PVO, Sobral LG, Rizzo ACL et al. (2005) Bioremediation of a weathered and a recently oil-contaminated soils from Brazil: a comparison study. Chemosphere 58:515–522.). Less is known about the biodegradability of petroleum commercial products such as kerosene (Gouda et al., 2007Gouda MK, Omar SH, Zohra A et al. (2007) Nour Eldin Bioremediation of kerosene I: A case study in liquid media. Chemosphere 69:1807–1814.). Shamiyan et al. (2013aShamiyan RK, Nirmal KJI, Rita NK et al. (2013a) Physico-chemical properties, heavy metal enrichment and fungal characterization of aged Gasoline contaminated soil site in Anand, Gujarat, India. Environ Exp Biol 11:137–143., b)Shamiyan RK, Nirmal KJI, Rita NK et al. (2013b) An assessment of physicochemical properties, heavy metal enrichment and fungal characterization of refined kerosene impacted soil in Anand, Gujarat, India. Int J Environ (In press). investigated the interactions of soil nutrients with Total Petroleum Hydrocarbons (TPH), with isolation and characterization of different petroleum-degrading fungal isolates. Moreover, no attempt has been made in the proposed work to establish the differential effects of various concentrations of kerosene on biochemical constituents of the fungal isolate Penicillium janthinellum SDX7 in liquid media during the biodegradation process. Hence, in the present investigation, an attempt has been made to optimize the fungal isolate at different conditions and its biochemical response during the petroleum hydrocarbon biodegradation of kerosene.
Materials and Methods
Fungal Isolate, Media and Culture Conditions
A fungal strain P. janthinellum SDX7 was isolated and screened from aged petroleum hydrocarbon-infected soil at the site of Anand, Gujarat, India, supporting the data of the TPH in the contaminated soil compared to the uncontaminated soil (garden soil). The isolate was identified as P. janthinellum SDX7 based on morphological and molecular (18S rRNA) methods, and the sequences were submitted to the National Center for Biotechnology Information (NCBI) gene bank under accession no. KC545842-KC545843 (Shamiyan et al., 2013bShamiyan RK, Nirmal KJI, Rita NK et al. (2013b) An assessment of physicochemical properties, heavy metal enrichment and fungal characterization of refined kerosene impacted soil in Anand, Gujarat, India. Int J Environ (In press).). Two types of media were used for cultivation, Potato Dextrose Agar (PDA) media and for the optimization and degradation studies, Mineral Salt Medium (MSM) with the addition of various concentrations of kerosene as the sole carbon source. The liquid medium was sterilized at 121°C for 15 min before addition of the kerosene (Zajic and Supplisson, 1972Zajic E, Supplisson B (1972) Emulsification and degradation of "Bunker C" fuel oil by microorganisms. Biotechnol Bioeng 14:31–43.).
Preparation of standardized inocula
Axenic spore suspensions of P. janthinellum SDX7 of approximately 105 spores/mLwere grown by adding 15 mL of sterile distilled water to mature fungal colonies on PDA plates (4–5 days) to dislodge the spores from the mycelium. These suspensions were then used to inoculate 100 mL MSM containing 3% (w/v) glucose in 500 mL Erlenmeyer flasks (Kendrick and Ratledge, 1996Kendrick A, Ratledge C (1996) Cessation of polyunsaturated fatty acid formation in four selected filamentous fungi when grown on plant oil. J Am Oil Chem Soc 73:431–435.). The cultures were incubated at 30 °C in an incubator shaker operating at 180 rpm for 48 h. The resultant active growing cultures were aseptically washed three times with 300 mL of sterile distilled water to remove remaining glucose fractions. This resulting culture was then used as the standard inoculum for further experiments.
Determination of optimal growth
The optimization study addresses different pH values, temperature, speed of agitation and kerosene concentration on P. janthinellum SDX7. A total of 10% (v/v) of standard inoculum was inoculated in each experiment and performed in triplicate. Biomass production (g/L) was used as an indicator for growth after 7 days of incubation. MSM medium with kerosene (without inoculation) was used as a control (Hamzah et al., 2012Hamzah A, Zarin MA, Hamid AA et al. (2012) Optimal Physical and Nutrient Parameters for Growth of Trichoderma virens UKMP-1 M for Heavy Crude Oil Degradation. Sains Malaysiana 41:71–79.).
Growth parameters
pH
The influence of the pH of the initial medium on the fungal growth of P. janthinellum SDX7 was determined at pH 4.0, 5.0, 6.0, 7.0, 8.0 and 9.0. Ten percent (v/v) standard inoculum was inoculated in a 500 mL Erlenmeyer flask containing 100 mL of MSM with the addition of 3% (v/v) kerosene and incubated at 30 °C in an orbital shaker at 180 rpm for 7 days. The pH that promoted the highest biomass production (in terms of dry weight) was used for subsequent steps of the investigation.
Temperature
The effect of temperature on P. janthinellum SDX7 growth was studied at 20, 30 and 40 °C in the MSM medium with 3% (v/v) kerosene at the determined optimum pH and incubated in an orbital shaker at 180 rpm for 7 days. The temperature that promoted the highest biomass production was used for the subsequent steps of the investigation.
Speed of agitation
The effect of different agitation at 130, 180 and 230 rpm during incubation on growth of P. janthinellum SDX7 was performed in the MSM medium with 3% (v/v) kerosene at optimum pH using an orbital shaker. Incubation was conducted at the determined optimum pH and temperature. The agitation speed that promoted the highest biomass production was used for the subsequent steps of the investigation.
Concentration of kerosene
The isolate of P. janthinellum SDX7 was grown in MSM prepared in accordance with the optimum nutrient parameters but supplemented with 1%, 3%, 5%, 10% and 20% (v/v) concentrations of kerosene and incubated at optimum pH, temperature and agitation speed for 7 days.
Dry weight measurement
The biomass of P. janthinellum SDX7 recovered by filtration using Whatman filter paper (No. 4) was washed with 100 mL chloroform to remove residual kerosene, then dried in the oven at 60 °C overnight, and cooled in a desiccators for 10–20 min prior to weighing.
Determination of biodegradation activity
The determination of the biodegradation activity of Penicillum janthinellum SDX7 was performed in 100 mL of the MSM medium treated by lower 1%, optimal 3% and higher 5% concentrations of kerosene in a 500 mL Erlenmeyer flask and incubated at 30 °C and agitated at 180 rpm for 16 days. The residual petroleum hydrocarbon was recovered by chloroform extraction at a ratio of 1:1 MSM medium: chloroform (Chaillan et al., 2004Chaillan F, Fleche AL, Bury E et al. (2004) Identification and biodegradation potential of tropical aerobic hydrocarbon-degrading microorganisms. Res Microbiol 155:587–595.). MSM without fungal inoculation was used as the control. Analysis of the fungal biodegradation activity was made using a computerized capillary gas chromatograph with flame ionization detector (GC/FID, Perkin Elmer-Auto System, SICART, V.V. Nagar) equipped with HP 3390A Integrator, split injector (split ratio 20/1) and flame ionization detector set at 300 °C. The carrier gas was nitrogen at a flow rate of 1.5 mL min−1. The column was polydimethylsiloxane (length 30 m, internal diameter 0.32 mm, film thickness 0.25 μm). The temperature was programmed to increase from 60 to 320 °C at 4 °C min−1. The total petroleum hydrocarbon (TPH) degradation by P. janthinellum SDX7 isolate was calculated according to the following equation:
where B is biodegradation, TPHC is the total petroleum hydrocarbon in the abiotic control (without fungal inoculation) and TPHI is the total petroleum hydrocarbon with inoculation (in this case with P. janthinellum SDX7).
GC/MS analysis
GC/MS spectra were acquired in the electron ionization mode (70 eV, nominal) scanning from m/z 30 to 650 s−1 and detected using an auto system XL GC apparatus (Perkin Elmer, SICART, V.V. Nagar). The column temperature was initially 80 °C, held for 5 min, then ramped from 80 °C- 290 °C at 10 °C min−1. Helium (1.0 mL min−1) was used as the carrier gas. Both line and injector temperatures were set at 250 °C. Each methanolic extract (1 μL) was injected in the split mode (1:40). MS conditions were EI + through a Perkin Elmer Turbo mass spectrometer as follows: ionization energy −70 e V, nominal; scan rate, 1.6 scans/s; inter-scan delay, 0.01 s; source temperature, 250 °C; mass range, 30 to 650 Daltons; solvent delay, 3.00 min. The gas chromatogram as reproduced by the mass spectrometer identified the mass spectrum scanned at each GC peak maximum. Data were thus obtained by comparing the mass spectra to those in the Wiley NIST/EPA/NIH Mass Spectral Library 2005.
Determination of metabolites
The variation in the metabolites was determined on the 4th, 8th, 12th and 16th days by cultivating the P. janthinellum SDX7 culture under optimal conditions such as pH 6, temperature of 30 °C, speed of agitation at 180 rpm and addition of optimal 3% (v/v) with higher 5% and lower 1% (v/v) dose of kerosene. Total carbohydrate release was determined spectrophotometrically by the anthrone method using glucose as the standard (Hedge and Hofreitte, 1991Hedge JE, Hofreitte BT (1991) Carbohydrates Chemistry. In: Sadasivam S, Manickam A (eds). Biochemical Methods for Agriculture Science. Wiley Eastern Ltd, p 8.). The protein content of the crude cell-free extract was estimated (Lowry et al., 1951Lowry OH, Rosenbrough NH, Farr AL et al. (1951) Protein measurements with Folin Phenol Reagent. J Biol Chem1 93:265–275.) using bovine serum albumin as the standard. An improved colorimetric determination of amino acids by the use of ninhydrin was performed (Lee and Takahashi, 1966Lee K, Weise AM, St-Pierre S (1996) Enhanced oil biodegradation with mineral fine interaction. Spill Sci Technol B 3:263–267.). Phenol stress metabolites were estimated using Folin-Ciocalteu reagent (Malick and Singh, 1980Malick CP, Singh MB (1980) Plant enzymology and histo enzymology. Kalyani publishers, New Delhi.). Each experiment was conducted in replicates of three and their ± SE values were calculated. Multivariate analysis was performed using KY plot (2.0 beta) to estimate the correlation between metabolites.
Growth of P. janthinellum SDX7 on other xenobiotic compounds
Fungal isolate P. janthinellum SDX7 was also tested for its growth on various xenobiotic compounds, namely, diesel fuel, gasoline fuel, toluene, benzene, xylene, naphthalene, pyrene and phenanthrene at a concentration of 0.1% [(w/v)/ (v/v)] as a sole carbon source in MSM and incubated under optimized conditions (at 30 °C temperature and shaking conditions of 180 rpm). The samples were withdrawn after 7 days, and the biomass was estimated in the form of the dry weight (Gojgic et al., 2012Gojgic-Cvijovic GD, Milic JS, Solevic TM et al. (2012) Biodegradation of petroleum sludge and petroleum polluted soil by a bacterial consortium: a laboratory study. Biodegradation 23:1–14.).
Results and Discussion
Optimization of growth parameters
P. janthinellum SDX7 grew better under acidic conditions, showing optimal growth (0.38 g/L) at pH 6. Previous studies also reported that several fungal isolates such as Fusariumsolani, F. oxysporum, Trichodermaviride (Verdin et al., 2004Verdin A, Sahraoui AL, Durand R (2004) Degradation of benzo[a]pyrene by mitosporic fungi and extracellular oxidative enzymes. Int Biodeterior Biodegradation 53:65–70.) and Aspergillus niger (Srivastava and Thakur, 2006Srivastava S, Thakur IS (2006) Evaluation of bioremediation and detoxification potentiality of Aspergillus niger for removal of hexavalent chromium in soil microcosm. Soil Biol Biochem 38:1904–1911.) cultured in acidic MSM medium provided good growth. Although its growth was highest under acidic conditions, isolate P. janthinellum SDX7 was able to grow sparsely in a relatively wide range of pH values from 4.0 to 9.0, suggesting that this isolate could degrade kerosene under not only acidic but also under neutral and alkaline conditions (Figure 1a). Among the parameters that could affect biomass production, temperature was generally considered to be the most important and limiting factor (Delille, 2004Delille D, Coulan F, Pelletier E (2004) Effects of temperature warming during a bioremediation study of natural and nutrient-amended hydrocarbon-contaminated sub-Antarctic soils. Cold Reg Sci Technol 40:61–70.). The common incubation temperature for the growth of fungi such as A. niger, Fusarium sp., Penicillium sp. and Graphium sp. is taken to be 30 °C (Santos and Linardi, 2004Santos VL, Linardi VR (2004) Biodegradation of phenol by a filamentous fungi isolated from industrial effluents-identification and degradation potential. Process Biochem 39:1001–1006.). Moreover, in this study, P. janthinellum SDX7 cultured at different temperatures produced maximum biomass (0.38 g/L) when incubated at 30 °C compared to 20 °C and 40 °C, temperatures that resulted in the production of a maximum of 0.31 g/L and 0.33 g/L biomass, respectively (Figure 1b). This difference may result from a greater production of enzymes and optimal growth conditions of the isolate for its kerosene degradation (Rohilla et al., 2012Rohilla SK, Salar RK, Kumar J (2012) Optimization of Physiochemical Parameters for Decolorization of Reactive Black HFGR Using Soil Fungus, Aspergillus allhabadii MTCC 9988. Int Biodeterior Biodegradation 3:1–5.). This range of temperatures makes this isolate suitable for use in bioremediation in tropical climates. P. janthinellum SDX7 showed an increase of biomass as the rate of agitation increased up to 180 rpm; biomass production was then reduced when the speed of agitation accelerated to 230 rpm (Figure 1c). Agitation influenced the tested fungi to absorb more nutrients by not only increasing the surface area of the microorganism for degradation of the kerosene oil hydrocarbons (Lee et al., 1996Lee K, Weise AM, St-Pierre S (1996) Enhanced oil biodegradation with mineral fine interaction. Spill Sci Technol B 3:263–267.) but also by booming the amount of dissolved oxygen in the cultivation medium (Purwanto et al., 2009Purwanto LA, Ibrahim D, Sudrajat H (2009) Effect of agitation speed on morphological changes in Aspergillus niger hyphae during production of tannase. World J Chem 4:34–38.). Agitation speed has also been proven to be a critical factor influencing mycelial biomass (Hamzah et al., 2012Hamzah A, Zarin MA, Hamid AA et al. (2012) Optimal Physical and Nutrient Parameters for Growth of Trichoderma virens UKMP-1 M for Heavy Crude Oil Degradation. Sains Malaysiana 41:71–79.). P. janthinellum was found to grow in a wide range of kerosene concentrations. The production of biomass rose to 0.38 g/L in the presence of 3% (v/v) of kerosene on the 5th day. However, the production of biomass declined when the concentration of kerosene escalated beyond 3% (v/v) (Figure 1d). Notably, minimal growth was found when the medium contained 10% and 20% kerosene doses.
Biodegradation of kerosene
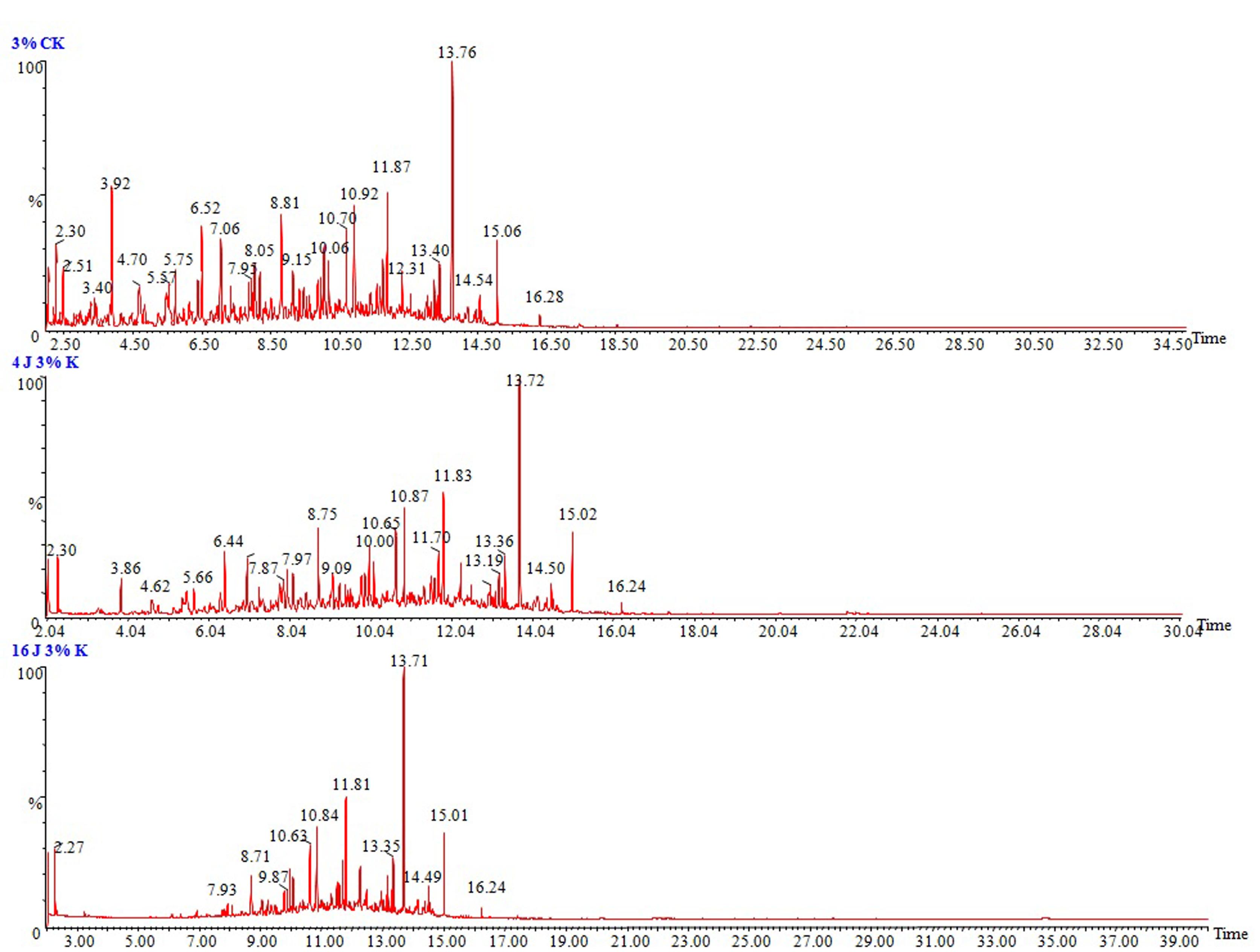
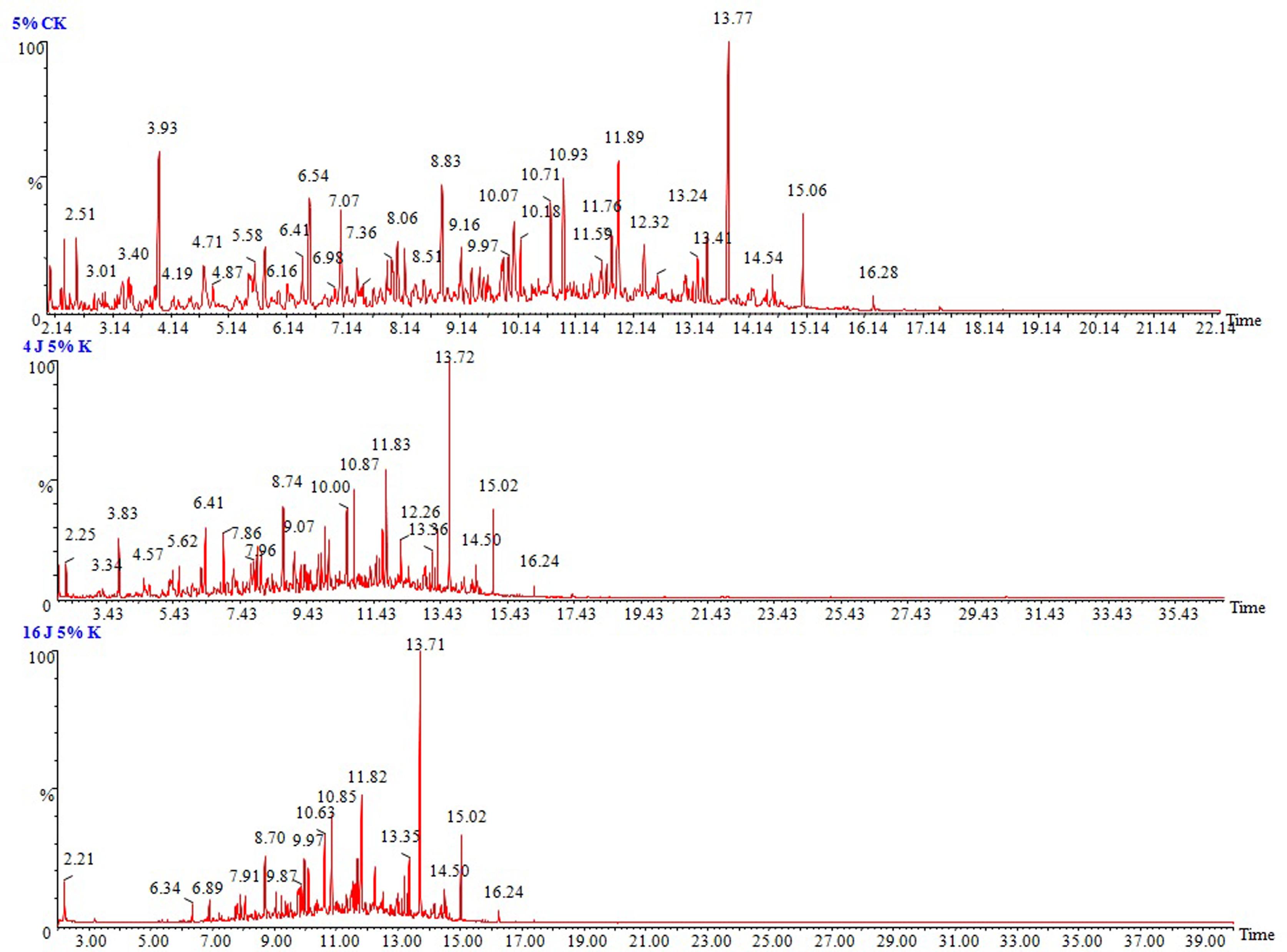
To determine the ability of P. janthinellum SDX7 to degrade kerosene, the total petroleum hydrocarbons were estimated at the end of the 4th and the 16th day of treatment at 1%, 3% and 5% kerosene treatments. Where 1% v/v kerosene showed the highest degradation at 85% and 95%, followed by 3% kerosene treatment at 32% and 63% degradation and a 5% kerosene dose by 28% and 58% reduction on 4th and 16th day, respectively (Figure 2). The biodegradation of kerosene was confirmed by the reduction in the area under the hydrocarbon peaks of the chromatograms when compared to that of the abiotic control (without organism), suggesting that the removal of kerosene hydrocarbon components ranged from 8 to 18 carbon atoms. In the chromatographic images of 1%, 3% and 5% (v/v) kerosene doses in a liquid medium, the sharp and highest peak stands for the n-alkanes, and the peaks between them comprise the naphthenes and aromatics. P. janthinellum SDX7 showed a reduction in the area under the hydrocarbon peaks corresponding to the carbon atoms (C8 to C18) when compared to the abiotic control in the case of the 1%, 3% and 5% kerosene, where the lowest concentration of 1% displayed the highest reduction (Figure 3) followed by 3% (Figure 4) and 5% (Figure 5). The chromatograms showed that the n-alkane fractions are easily degraded by the tested fungal isolate as the days progress. However, the rate might be lower for branched alkanes followed by n-alkylaromatics, cyclic alkanes and polynuclear aromatics. The present results are in agreement with the findings obtained by Wang et al. (1998)Wang Z, Fingas M, Blenkinsopp S et al. (1998) Comparsion of oil composition changes due to biodegradation and physical weathering in different oils. J Chromatogr A 809:89–107., who studied the comparison of crude oil composition changes due to biodegradation and physical weathering using different fungal and bacterial isolates. Moreover, Nocentini et al. (2000)Nocentini M, Nocentini D, Pinelli F et al. (2000) Bioremediation of a soil contaminated by hydrocarbon mixtures: the residual concentration problem. Chemosphere 41:1115–1123. studied the biotreatability and feasibility of a bioremediation process by a bacterial species for a kerosene-contaminated soil. The results indicated that the P. janthinellum SDX7 isolate in this study is extremely efficient in degrading kerosene hydrocarbons. Our results also agreed very well with the findings of Mancera-López et al. (2008)Mancera-López ME, Esparza-García F, Chávez-Gómez B et al. (2008) Bioremediation of an aged hydrocarbon-contaminated soil by a combined system of biostimulation-bioaugmentation with filamentous fungi. Int Biodeterior Biodegradation 61:151–160., who reported that the Penicillum genus was one of the major hydrocarbon-degrading groups.
Degradation of different concentrations of kerosene after 4th and 16th days by P. janthinellum SDX7.
GC/MS chromatograms of 1% (v/v) kerosene in MSM medium. (a) Abiotic control, (b) After 4 days of incubation with P. janthinellum SDX7 strain, and (c) After 16 days incubation with P. janthinellum SDX7 strain. The sharp and highest peaks stand for the n-alkanes, and the peaks between them comprise the naphthenes and aromatics.
GC/MS chromatograms of 3% (v/v) kerosene in MSM medium. (a) Abiotic control, (b) After 4 days of incubation with P. janthinellum SDX7 strain, and (c) After 16 days incubation with P. janthinellum SDX7 strain.
GC/MS chromatograms of 5% (v/v) kerosene in MSM medium. (a) Abiotic control, (b) After 4 days of incubation with P. janthinellum SDX7 strain, and (c) After 16 days of incubation with P. janthinellum SDX7 strain.
Metabolites
Based on the inhibitory effects and growth arrest, the release of certain metabolic products such as carbohydrates, amino acids and proteins was affected at the earlier stage of kerosene applications, a result that is in agreement with the study of Kumar et al. (2013)Nirmal Kumar JI, Megha B, Shamiyan RK (2013) Some intermediate bio-transformants during biodegradation of high molecular weight phenanthrene and fluoranthene by cyanobacterial species - Aulosira fertilissima ghose. Int J App Sci Biotechnol 1:97–105. who studied the impact of PAH exposure on certain cyanobacterial species. The level of total carbohydrates decreased after 12 to 16 days in all the concentrations of kerosene-treated isolate. After the 12th day of exposure, carbohydrate content ranged from 0.6 mgmL−15 to 2.24 mgmL−15 and was significantly reduced by 50%, 11% and 76%, whereas a higher level of reduction was encountered after the 16th day by 59%, 24% and 80% at 1%, 3% and 5% kerosene exposures to P. janthinellum SDX7, respectively (Figure 6a). A similar reduction in the carbohydrate content was recorded by Kumar et al. (2008)Kumar S, Khalid H, Tasneem F (2008) Endosulfan induced biochemical changes in nitrogen-fixing cyanobacteria. Sci Total Environ 403:130–138. in nitrogen-fixing cyanobacteria when treated with endosulfan. Protein content fluctuated from 1.44 mgmL−15 to 3.78 mgmL−15 and was inhibited by 46%, 13% and 67% on the 12th day (Figure 6b) and 70%, 48% and 81% after the 16th day, significantly at 1%, 3% and 5%, respectively, of kerosene-treated P. janthinellum SDX7 isolate as shown by Babu et al. (2001)Babu SG, Hans RK, Singh J et al. (2001) Effect of lindane on the growth and metabolic activities of cyanobacteria. Ecotoxicol Environ Saf 48:219–221. and Laxmi et al. (2007)Lakshmi PTV, Annamalai A (2007) Biochemical studies on response of organo-phosphorus insecticide and release of extra cellular products by cyanobacteria. Res J Fisheries Hydrobiol 2:13–17. in response to lindane and organophosphorus on certain cyanobacterial species. At the end of the experiment after the 16th day, the highest reduction of amino acids in P. janthinellum SDX7 (by 95%) was observed when treated with 5% kerosene relative to the control, which ranged from 0.05 mgmL−15 to 0.4 mgmL−15 (Figure 6c). The optimal concentration of 3% kerosene showed less impact on the metabolite reduction. However, the greatest reduction observed at the higher concentration of 5% followed by the lower concentration of 1% kerosene showed results quite well correlated with the findings of Standyk et al. (1971)Standyk V, Camphell RS, Johnsin T (1971) Pesticide effect on growth and C-14 assimilation in fresh water algae. Bullet Environ Cont Toxicol 6:1–8., who depicted concentration-dependent inhibition of amino acids and proteins at an earlier stage based on inhibitory effects and growth arrest in fresh water algae in response to pesticide treatments.
Metabolic content in P. janthinellum SDX7 treated with kerosene at different daily intervals.
A more pronounced effect of kerosene on phenol content was observed on the tested fungal isolate. The release of the stress metabolite phenol was significantly stimulated after 4days of exposure, varying from 0.4 mgmL−15 to 0.64 mgmL−15 and higher in all treated cultures compared to untreated isolate (Figure 6d). The highest level of phenol was observed in 5% treated isolates, showing 61%, 63%, 65% and 66% increase on the 4th, 8th, 12th and 16th days, respectively. Elevated levels of phenols might be due to the liberation of the phenolic compounds during gradual degradation of kerosene by hydrolysis or oxidation processes and release under stress conditions because of the catabolic activity of primary metabolites (Mostafa and Helling, 2002Mostafa FI, Helling CS (2002) Impact of four Pesticides on the Growthand Metabolic Activities of Two Photosynthetic Algae. J Environ Sci Health 37:417–444.). Kumar and Kumar (2002)Kumar NJI, Kumar RN (2002) Some metabolic observations of Nostocmuscorum to herbicide Fluchloralin. Plant Arch 2:289–293. also suggested that the release of phenols due to applications of different concentrations of fungicide was higher than untreated cultures, possibly due to the accumulation of phenolic compounds from the larger polycyclic aromatic compounds.
A significant positive correlation between carbohydrates, proteins and amino acids (r = 0.88 to 0.94) was encountered, whereas a highly negative correlation was registered with phenols (r= −0.48 to −0.72) after 16 days of exposure to kerosene (Table 1).
Growth of P. janthinellum SDX7 on various xenobiotic compounds
The efficiency of the fungal isolate in utilizing other xenobiotic compounds as the sole source of carbon is represented in Table 2. P. janthinellum SDX7 displayed a very high growth in the presence of kerosene, diesel and gasoline fuels. However, a moderate growth was encountered in case of naphthalene and poor growth was registered for phenanthrene, toluene, benzene and phenol. The isolate was not able to utilize the high molecular weight PAHs such as pyrene. Our results have been further substantiated by earlier studies of Okerentugba and Ezeronye (2003)Okerentugba PD, Ezeronye OU (2003) Petroleum degrading potentials of single and mixed microbial cultures isolated from rivers and refinery effluent in Nigeria. Afr J Biotechnol 2:288–292., revealing the degradation of various xenobiotic compounds by P. janthinellum SDX7 based on the ability of utilizing these xenobiotic compounds as the sole source of carbon.
Growth of kerosene-degrading P. janthinellum SDX7 in the presence of different xenobiotic compounds (0.1% [(v/v)/ (w/v)].
The present investigation suggests that the fungal isolate P. janthinellum SDX7 employed in this study is extremely efficient in degrading kerosene, displaying a maximum degradation of 95% after 16 days under optimal growth conditions based on its ability to utilize kerosene. The optimal kerosene concentration of 3% showed the least reduction of the metabolites- carbohydrates, proteins and amino acids compared to the 1% and 5% (v/v) kerosene doses on the 12th and 16th days of exposure. Stress metabolite phenol was found to rise at lower and higher concentrations due to the inhibitory effect and growth retardation of the test organism in response to kerosene treatments. The highly efficient degradative ability of P. janthinellum SDX7 proved to be suitable for mycoremediation of kerosene-contaminated soil environments.
Acknowledgments
One of the authors, Ms. Shamiyan Rahat Khan, is highly grateful to the University Grants Commission (UGC) for financial support for receiving the Maulan Azad Fellowship, and the authors are also thankful to the Sophisticated Instrumentation Centre for Advanced Research and Testing (SICART) for analysis of samples by GC.
References
- Babu SG, Hans RK, Singh J et al. (2001) Effect of lindane on the growth and metabolic activities of cyanobacteria. Ecotoxicol Environ Saf 48:219–221.
- Bento FM, Camargo FA, Okeke B et al. (2003) Bioremediation of soil contaminated by diesel oil. Braz J Microbiol 34:65–68.
- Chaillan F, Fleche AL, Bury E et al. (2004) Identification and biodegradation potential of tropical aerobic hydrocarbon-degrading microorganisms. Res Microbiol 155:587–595.
- Delille D, Coulan F, Pelletier E (2004) Effects of temperature warming during a bioremediation study of natural and nutrient-amended hydrocarbon-contaminated sub-Antarctic soils. Cold Reg Sci Technol 40:61–70.
- Gojgic-Cvijovic GD, Milic JS, Solevic TM et al. (2012) Biodegradation of petroleum sludge and petroleum polluted soil by a bacterial consortium: a laboratory study. Biodegradation 23:1–14.
- Gouda MK, Omar SH, Zohra A et al. (2007) Nour Eldin Bioremediation of kerosene I: A case study in liquid media. Chemosphere 69:1807–1814.
- Greer CW, Fortin N, Roy R et al. (2003) Indigenous sediment microbial activity in response to nutrient enrichment and plant growth following a controlled oil spill on a fresh water wetland. Bioremed J 7:69–80.
- Hamzah A, Zarin MA, Hamid AA et al. (2012) Optimal Physical and Nutrient Parameters for Growth of Trichoderma virens UKMP-1 M for Heavy Crude Oil Degradation. Sains Malaysiana 41:71–79.
- Hedge JE, Hofreitte BT (1991) Carbohydrates Chemistry. In: Sadasivam S, Manickam A (eds). Biochemical Methods for Agriculture Science. Wiley Eastern Ltd, p 8.
- Irwin RJ, Van Mouwerik M, Stevens L et al. (1997) Environmental Contaminants Encyclopedia. Kerosene Entry. National Park Service, Water Resources Division, Water Operating Branch. Oakridge Drive, Suite Fort Collins, 42 p.
- Kendrick A, Ratledge C (1996) Cessation of polyunsaturated fatty acid formation in four selected filamentous fungi when grown on plant oil. J Am Oil Chem Soc 73:431–435.
- Kumar NJI, Kumar RN (2002) Some metabolic observations of Nostocmuscorum to herbicide Fluchloralin. Plant Arch 2:289–293.
- Kumar S, Khalid H, Tasneem F (2008) Endosulfan induced biochemical changes in nitrogen-fixing cyanobacteria. Sci Total Environ 403:130–138.
- Lakshmi PTV, Annamalai A (2007) Biochemical studies on response of organo-phosphorus insecticide and release of extra cellular products by cyanobacteria. Res J Fisheries Hydrobiol 2:13–17.
- Lee K, Weise AM, St-Pierre S (1996) Enhanced oil biodegradation with mineral fine interaction. Spill Sci Technol B 3:263–267.
- Lee Y, Takahasi T (1966) An Imported Colorimetric Determination of Amino acids with the use of Ninhydrin. Anal Biochem 14:71–77.
- Lindstrom JE, Braddock JF (2002) Biodegradation of petroleum hydrocarbons at low temperature in the presence of the dispersant Corexit 9500. Mar Pollut Bull 44:739–747.
- Lowry OH, Rosenbrough NH, Farr AL et al. (1951) Protein measurements with Folin Phenol Reagent. J Biol Chem1 93:265–275.
- Malick CP, Singh MB (1980) Plant enzymology and histo enzymology. Kalyani publishers, New Delhi.
- Mancera-López ME, Esparza-García F, Chávez-Gómez B et al. (2008) Bioremediation of an aged hydrocarbon-contaminated soil by a combined system of biostimulation-bioaugmentation with filamentous fungi. Int Biodeterior Biodegradation 61:151–160.
- Margesin R, Schinner F (1997) Bioremediation of diesel-oil contaminated alpine soils at low temperatures. Appl Microbiol Biot 47:462–468.
- Morgan P, Watkinson RJ (1989) Hydrocarbon degradation and methods for soil biotreatment. CRC Crit Rev Biotechnol 8:305–333.
- Mostafa FI, Helling CS (2002) Impact of four Pesticides on the Growthand Metabolic Activities of Two Photosynthetic Algae. J Environ Sci Health 37:417–444.
- Mukherji S, Jagadevan S, Mohapatra G et al. (2004) Biodegradation of diesel oil by an Arabian Sea sediment culture isolated from the vicinity of an oil field. Bioresour Technol 95:281–286.
- Nocentini M, Nocentini D, Pinelli F et al. (2000) Bioremediation of a soil contaminated by hydrocarbon mixtures: the residual concentration problem. Chemosphere 41:1115–1123.
- Nirmal Kumar JI, Megha B, Shamiyan RK (2013) Some intermediate bio-transformants during biodegradation of high molecular weight phenanthrene and fluoranthene by cyanobacterial species - Aulosira fertilissima ghose. Int J App Sci Biotechnol 1:97–105.
- Nirmal Kumar JI, Jignasha GP, Rita NK et al. (2013) Chronic response of three different Cyanobacterial species on growth, pigment and metabolic variations to the high molecular weight Polycyclic Aromatic Hydrocarbon- Pyrene. Poly Aro Hydro (In Press).
- Okerentugba PD, Ezeronye OU (2003) Petroleum degrading potentials of single and mixed microbial cultures isolated from rivers and refinery effluent in Nigeria. Afr J Biotechnol 2:288–292.
- Okoh AI (2003) Biodegradation of bonny light crude oil in soil microcosm by some bacterial strains isolated from crude flow stations saver pits in Nigeria. Afr J Biotechnol 2:104–108.
- Purwanto LA, Ibrahim D, Sudrajat H (2009) Effect of agitation speed on morphological changes in Aspergillus niger hyphae during production of tannase. World J Chem 4:34–38.
- Rohilla SK, Salar RK, Kumar J (2012) Optimization of Physiochemical Parameters for Decolorization of Reactive Black HFGR Using Soil Fungus, Aspergillus allhabadii MTCC 9988. Int Biodeterior Biodegradation 3:1–5.
- Santos VL, Linardi VR (2004) Biodegradation of phenol by a filamentous fungi isolated from industrial effluents-identification and degradation potential. Process Biochem 39:1001–1006.
- Shamiyan RK, Nirmal KJI, Rita NK et al. (2013a) Physico-chemical properties, heavy metal enrichment and fungal characterization of aged Gasoline contaminated soil site in Anand, Gujarat, India. Environ Exp Biol 11:137–143.
- Shamiyan RK, Nirmal KJI, Rita NK et al. (2013b) An assessment of physicochemical properties, heavy metal enrichment and fungal characterization of refined kerosene impacted soil in Anand, Gujarat, India. Int J Environ (In press).
- Snape I, Riddle MJ, Stark JS et al. (2001) Management andremediation of contaminated sites at Casey station. Antart Polar Rec 37:199–214.
- Srivastava S, Thakur IS (2006) Evaluation of bioremediation and detoxification potentiality of Aspergillus niger for removal of hexavalent chromium in soil microcosm. Soil Biol Biochem 38:1904–1911.
- Standyk V, Camphell RS, Johnsin T (1971) Pesticide effect on growth and C-14 assimilation in fresh water algae. Bullet Environ Cont Toxicol 6:1–8.
- Sugiura K, Ishihara M, Shimauchi T et al. (1997) Physicochemical properties and biodegradability of crude oil. Environ Sci Technol 31:45–51.
- Trindade PVO, Sobral LG, Rizzo ACL et al. (2005) Bioremediation of a weathered and a recently oil-contaminated soils from Brazil: a comparison study. Chemosphere 58:515–522.
- Verdin A, Sahraoui AL, Durand R (2004) Degradation of benzo[a]pyrene by mitosporic fungi and extracellular oxidative enzymes. Int Biodeterior Biodegradation 53:65–70.
- Wang Z, Fingas M, Blenkinsopp S et al. (1998) Comparsion of oil composition changes due to biodegradation and physical weathering in different oils. J Chromatogr A 809:89–107.
- Zajic E, Supplisson B (1972) Emulsification and degradation of "Bunker C" fuel oil by microorganisms. Biotechnol Bioeng 14:31–43.
Publication Dates
-
Publication in this collection
Apr-Jun 2015
History
-
Received
06 Feb 2014 -
Accepted
07 Aug 2014