Resumos
A extensa área coberta pelo domínio amazônico e os recursos financeiros limitados para estudos da biodiversidade exigem programas de pesquisa que obtenham o máximo de informação com os menores custos. Com o objetivo de avaliar as conseqüências da redução do esforço amostral sobre a retenção da informação ecológica, testou-se como diferentes larguras de parcela afetam os resultados da relação entre variação ambiental e composição da comunidade de pteridófitas da Amazônia Central. Foram feitas medidas da inclinação do terreno, abertura do dossel e do teor de argila do solo, em 37 parcelas de floresta de terra-firme. Todos os indivíduos de pteridófitas foram identificados e mapeados com relação a três faixas de amostragem. Foram testados os efeitos das variáveis ambientais sobre a composição de pteridófitas em parcelas de 250 x 2,5 m e em sub-amostras com a largura da parcela reduzida. A redução de 2,5 para 1 m na largura da parcela (60% de redução no esforço amostral) correspondeu a uma redução de 24% no custo do projeto, considerado os custos das excursões à campo, e a uma redução no número de espécies amostradas de 52 para 44. A redução 2,5 para 2 m na largura da parcela causou redução de apenas duas (4%) nas espécies amostradas e 8% no custo do projeto. Para todas as larguras testadas, a composição da comunidade esteve associada ao teor de argila e não esteve associada à inclinação do terreno. O efeito de abertura de dossel sobre a composição de espécies de pteridófitas não foi coerente entre os diferentes tamanhos de amostra. Esta incoerência deve estar relacionada à importância relativamente menor da luz sobre a estruturação da comunidade. Concluímos que as amostras com menor tamanho foram suficientemente informativas para detectar os principais gradientes de composição e sua associação aos fatores ambientais. Isso permitiria a redução nos custos totais ou alocação de mais parcelas pelo mesmo custo, o que poderia aumentar o poder dos testes estatísticos, reduzir o intervalo de confiança e aumentar a probabilidade de detecção de mais espécies.
comunidade; gradiente ambiental; floresta tropical; protocolo de amostragem; riqueza de espécies; samambaia; textura do solo
The large area covered by the Amazonian domain and the limited financial support for biodiversity studies demand efficient research programs. Aiming to evaluate the consequences of reduced sampling effort on the retention of ecological information, we tested how differences in plot width affect the perceived relationship between environmental variation and the composition of the pteridophyte community in Central Amazonia. Measures of slope, canopy openness and soil clay content were taken in 37 terra-firme forest plots. All pteridophyte individuals were identified and mapped according to three sample strips. We tested the effects of environmental variables on pteridophyte composition of 250 x 2.5 meters-plots and on sub-samples of reduced width. The reduction of plot width from 2.5 to 1 m (60% reduction in sampling effort) corresponded to a reduction of 24% of the project's costs concerning field work expenses. This would reduce the number of sampled species from 52 to 44. The reduction of 20% plot width (from 2.5 to 2 m) would cause a reduction of two (4%) sampled species and a reduction of 8% of the costs. For all tested plot widths, community composition was associated with soil clay content and was not associated with the terrain slope. The effect of canopy openness on pteridophyte species composition was not consistent among sample sizes. These may be related to the relatively lower importance of light availability in determining community structure at the studied spatial scale. We concluded that the smallest plots were informative enough to detect the main gradients of composition and their association to environmental factors. This would allow a reduction in total costs, or the allocation of available budget to more plots, which could increase the power of the statistical analyses, reduce the confidence intervals and increase probability of detecting more species.
community; environmental gradients; ferns; sampling protocol; species richness; soil texture; tropical forest
ARTIGOS
Redução de esforço amostral vs. retenção de informação em inventários de pteridófitas na Amazônia Central
Sample effort reduction vs. information retention in inventories of pteridophytes in Central Amazonia
Gabriela ZuquimI, 1 1 Autor para correspondência: Gabriela Zuquim, e-mail: zuquim@inpa.gov.br ; Flávia Regina Capellotto CostaI; Jefferson PradoII
IInstituto Nacional de Pesquisas da Amazônia, Av. André Araújo, CEP 69083-000, Manaus, AM, Brasil
IIInstituto de Botânica de São Paulo, Av. Miguel Stéfano, 3687, CEP 04301-902, São Paulo, SP, Brasil
RESUMO
A extensa área coberta pelo domínio amazônico e os recursos financeiros limitados para estudos da biodiversidade exigem programas de pesquisa que obtenham o máximo de informação com os menores custos. Com o objetivo de avaliar as conseqüências da redução do esforço amostral sobre a retenção da informação ecológica, testou-se como diferentes larguras de parcela afetam os resultados da relação entre variação ambiental e composição da comunidade de pteridófitas da Amazônia Central. Foram feitas medidas da inclinação do terreno, abertura do dossel e do teor de argila do solo, em 37 parcelas de floresta de terra-firme. Todos os indivíduos de pteridófitas foram identificados e mapeados com relação a três faixas de amostragem. Foram testados os efeitos das variáveis ambientais sobre a composição de pteridófitas em parcelas de 250 x 2,5 m e em sub-amostras com a largura da parcela reduzida. A redução de 2,5 para 1 m na largura da parcela (60% de redução no esforço amostral) correspondeu a uma redução de 24% no custo do projeto, considerado os custos das excursões à campo, e a uma redução no número de espécies amostradas de 52 para 44. A redução 2,5 para 2 m na largura da parcela causou redução de apenas duas (4%) nas espécies amostradas e 8% no custo do projeto. Para todas as larguras testadas, a composição da comunidade esteve associada ao teor de argila e não esteve associada à inclinação do terreno. O efeito de abertura de dossel sobre a composição de espécies de pteridófitas não foi coerente entre os diferentes tamanhos de amostra. Esta incoerência deve estar relacionada à importância relativamente menor da luz sobre a estruturação da comunidade. Concluímos que as amostras com menor tamanho foram suficientemente informativas para detectar os principais gradientes de composição e sua associação aos fatores ambientais. Isso permitiria a redução nos custos totais ou alocação de mais parcelas pelo mesmo custo, o que poderia aumentar o poder dos testes estatísticos, reduzir o intervalo de confiança e aumentar a probabilidade de detecção de mais espécies.
Palavras-chave: comunidade, gradiente ambiental, floresta tropical, protocolo de amostragem, riqueza de espécies, samambaia, textura do solo.
ABSTRACT
The large area covered by the Amazonian domain and the limited financial support for biodiversity studies demand efficient research programs. Aiming to evaluate the consequences of reduced sampling effort on the retention of ecological information, we tested how differences in plot width affect the perceived relationship between environmental variation and the composition of the pteridophyte community in Central Amazonia. Measures of slope, canopy openness and soil clay content were taken in 37 terra-firme forest plots. All pteridophyte individuals were identified and mapped according to three sample strips. We tested the effects of environmental variables on pteridophyte composition of 250 x 2.5 meters-plots and on sub-samples of reduced width. The reduction of plot width from 2.5 to 1 m (60% reduction in sampling effort) corresponded to a reduction of 24% of the project's costs concerning field work expenses. This would reduce the number of sampled species from 52 to 44. The reduction of 20% plot width (from 2.5 to 2 m) would cause a reduction of two (4%) sampled species and a reduction of 8% of the costs. For all tested plot widths, community composition was associated with soil clay content and was not associated with the terrain slope. The effect of canopy openness on pteridophyte species composition was not consistent among sample sizes. These may be related to the relatively lower importance of light availability in determining community structure at the studied spatial scale. We concluded that the smallest plots were informative enough to detect the main gradients of composition and their association to environmental factors. This would allow a reduction in total costs, or the allocation of available budget to more plots, which could increase the power of the statistical analyses, reduce the confidence intervals and increase probability of detecting more species.
Keywords: community, environmental gradients, ferns, sampling protocol, species richness, soil texture, tropical forest.
Introdução
A extensa área do domínio amazônico e os recursos financeiros limitados para estudos da biodiversidade exigem programas de pesquisa que obtenham o máximo de informação com os menores custos, baseados em amostragens informativas e facilmente replicáveis. Amostragens em parcelas possuem área e esforço amostral bem determinados e contribuem para melhor compreensão de relações ecológicas (Phillips et al. 2003). Surpreendentemente, nos trópicos, onde as limitações financeiras e de taxonomistas são ainda maiores, pouco esforço tem sido feito para a avaliação da qualidade dos protocolos de amostragem utilizados em inventários da biodiversidade (Phillips et al. 2003).
Para assegurar que réplicas de estudos serão feitas de maneira comparável, deve haver um protocolo detalhado (Condit 1998). A padronização das amostragens permite que estudos com os mesmos objetivos, mas em áreas amostrais diferentes sejam comparáveis (Magnusson et al. 2005). Apesar disso, a importância de protocolos de amostragem gerais, que se adequem a diferentes grupos e sistemas biológicos é freqüentemente negligenciada por ecólogos. Belovsky et al. (2004) apontam que a incompatibilidade de metodologia e a falta de rigor na coleta de dados são fatores que prejudicam o progresso da ecologia.
Inventários biológicos geralmente pretendem responder três questões principais: 1) qual o número de espécies (riqueza) da área, 2) qual a composição de espécies e 3) como as espécies se associam ao ambiente. As respostas a estas questões dependem de características da amostragem, já que o tamanho, formato e orientação das unidades amostrais podem determinar quais questões serão respondidas (Caughley & Sinclair 1994, Magnusson & Mourão 2003). Parcelas de 1 ha quadradas são amplamente utilizadas em inventários de árvores na Amazônia (Prance et al. 1976, Gentry 1988a, b, Oliveira & Mori 1999, Pitman et al. 2002), mas possuem utilidade limitada em estudos de padrões de distribuição espacial de organismos no ambiente por serem internamente heterogêneas quanto às características edáficas e topográficas (Phillips et al. 2003).
Inventários da biodiversidade feitos em unidades amostrais comuns são a única forma de obter dados comparáveis no espaço e no tempo (Belovsky et al. 2004). Porém, amostrar todos os tipos de organismos em parcelas de mesma dimensão é pouco eficiente. De maneira geral, em estudos sobre organismos grandes são necessárias unidades amostrais grandes, enquanto para organismos pequenos ou muito abundantes, a área da unidade amostral pode ser menor. Tipicamente, o número de indivíduos de qualquer grupo biológico necessário para que se tenha uma boa estimativa da diversidade regional varia entre 200 e 500 por unidade amostral (Hayek & Buzas 1997). Em inventários de árvores (DAP > 10 cm) na Amazônia, estes valores são atingidos com cerca de 1 ha de área amostrada (Gentry 1998b, Oliveira & Mori 1999, Pitman et al. 2002). Entretanto, 1 ha de floresta na Amazônia Central pode conter cerca de 8.700 indivíduos de espécies herbáceas (Costa 2006), ou 6.750 pteridófitas e 720 melastomatáceas na Amazônia Equatoriana (Tuomisto et al. 2002). Fica claro que não é viável amostrar muitas parcelas de 1 ha para plantas pequenas como a maior parte das plantas de subosque.
O tamanho da unidade amostral determina o número de indivíduos nela contidos, e, portanto, quanto da comunidade local está representada. Entretanto, conforme o tamanho da unidade amostral aumenta, a heterogeneidade interna tende a aumentar, o que causa problemas para determinar as associações entre espécies e ambiente (Magnusson et al. 2005). Além disso, o aumento no número de indivíduos conforme aumenta o tamanho da unidade amostral pode ser informativo ou não. O aumento no número de indivíduos pode melhorar a representação da distribuição de abundâncias das espécies na comunidade (Hubbell 2001) o que é informativo, ou apenas repetir o padrão que pode ser observado com menor número de indivíduos. Portanto, determinar o tamanho ideal de uma unidade amostral para um determinado grupo de organismos envolve maximizar a representação da estrutura da comunidade, ao mesmo tempo em que se minimiza a heterogeneidade ambiental interna. Como o tempo e o custo da amostragem aumentam (embora não necessariamente de forma linear) com o tamanho da unidade amostral, a diminuição do tamanho das unidades permite que um número maior de unidades possa ser amostrado por área com o mesmo custo total. Gentry (1982) mostrou que várias parcelas pequenas são mais eficientes para capturar a diversidade de uma região do que uma parcela grande. É possível reduzir o tamanho ou número de sub-amostras considerados ideais nos programas de inventário tradicionais, com pouca perda de informação. Pesquisadores envolvidos no PPBio mostraram que mais de 75% da informação sobre a composição de espécies é retida utilizando apenas 12,5% das amostras dos ácaros que foram coletados em uma savana Amazônica. Essa redução tornaria o projeto US$ 5.000 (88%) mais barato (Santos 2001).
O delineamento amostral utilizado neste trabalho segue o método RAPELD, sigla que une as duas escalas temporais que o método contempla: inventários rápidos, RAP e Projetos Ecológicos de Longa-Duração, PELD (Magnusson et al. 2005). O RAPELD foi adotado pelo Programa de Pesquisas em Biodiversidade (PPBio), que é financiado pelo Ministério de Ciência e Tecnologia. Um dos objetivos do PPBio para o monitoramento da biodiversidade brasileira é implementar um delineamento padronizado que permita a comparação do mesmo grupo biológico entre regiões, e de grupos diferentes na mesma região (Magnusson et al. 2005). O delineamento para inventários do PPBio busca ajustar as necessidades amostrais de diversos grupos biológicos dentro das mesmas unidades amostrais (parcelas), propiciando a integração dos dados bióticos e abióticos, bem como a integração de dados de diferentes grupos biológicos.
Em investigações de padrões de distribuição espacial de organismos, os fatores ambientais precisam ser bem controlados estatisticamente (Ricklefs 2004). Para as comunidades de plantas neotropicais, os fatores ambientais que têm maior influência sobre a estrutura das comunidades, desde a mesoescala até a escala da paisagem, são as características associadas ao solo (como textura e disponibilidade de nutrientes) e à topografia (Pitman et al. 2001, Condit et al. 2002, Tuomisto et al. 2003a, b, Vormisto et al. 2004, Svenning et al. 2006). No método RAPELD, a variação interna da textura do solo na parcela é minimizada, pois as parcelas seguem a curva de nível do terreno (Magnusson et al. 2005). Na Amazônia Central, há uma forte correlação entre textura do solo e altitude (Chauvel et al. 1987) e assim, uma parcela posicionada ao longo da curva de nível minimiza a variação na altitude e, portanto, nas condições de textura do solo.
Para a maior parte dos estudos ecológicos e para propósitos aplicados, tais como planejamento de conservação e uso da terra, medidas de riqueza têm pouca utilidade quando isoladas. Por outro lado, informações sobre a composição e complementaridade das assembléias de espécies podem ser usadas para planejamento, mesmo na ausência de informação sobre o número total de espécies (Margules et al. 2002). Muitas vezes, características grosseiras do hábitat, como estrutura da vegetação ou a geomorfologia, podem ser usados como substitutos para informações sobre a distribuição de cada espécie (Margules et al. 2002), mas somente se a forma como as variações no ambiente afetam cada grupo de espécies é conhecida.
As parcelas terrestres sugeridas pelo método RAPELD possuem comprimento fixo de 250 m, mas a largura é determinada pelo pesquisador, de acordo com seu organismo de interesse. Por exemplo, na Reserva Florestal Adolpho Ducke, Amazônia Central, Castilho (2005) utilizou 40 m de largura para levantamentos de árvores acima de 30 cm de diâmetro à altura do peito (DAP), mas Costa et al. (2005) e Costa (2006) utilizaram 2 m para a amostragem de plantas herbáceas. Braga-Neto et al. (2007) utilizou 25 cm para fungos de serapilheira e Nogueira (2006) usou 10 m para lianas lenhosas acima de 1 cm de DAP.
Este trabalho teve por objetivo avaliar as conseqüências de uma das maneiras de se reduzir o esforço amostral em inventários, sobre a retenção da informação ecológica. Para isso, utilizou-se as pteridófitas como um modelo para testar como os diferentes tamanhos de parcela afetam as relações entre a variação ambiental e a variação na composição de uma comunidade. Dentre as características ambientais mais comumente utilizadas para modelagem da distribuição dos organismos estão o solo, a topografia e a estrutura da vegetação. Portanto, variáveis que representam cada uma destas características foram selecionadas para este estudo.
O ajuste do tamanho da parcela será importante para os futuros inventários de pteridófitas nos sítios do PPBio atualmente implantados no Parque Nacional do Viruá (RR), Estação Ecológica Maracá (RR), Caxiuanã (PA), Pantanal (MT) e Reserva Biológica do Uatumã (AM), bem como aqueles a serem implantados em toda a região amazônica.
Material e Métodos
1. Área de estudo
O estudo foi feito em três reservas de cerca de 12 km2 de floresta pluvial tropical de terra-firme da Amazônia Central. A pluviosidade média anual na região é de 2200 mm e a temperatura média anual é de 26,7 °C (RADAMBRASIL 1978). Os dados foram coletados nas Reservas do Km 41 (02° 25' S e 59° 45' O), Cabo Frio (02° 23' S e 59° 54' O) e Gavião (02° 24' S e 59° 50' O), que fazem parte do Projeto Dinâmica Biológica de Fragmentos Florestais (PDBFF), 80 km ao norte de Manaus (Figura 1). Nas áreas do PDBFF, os solos são tipicamente ácidos, com elevada concentração de alumínio e pobres em fósforo, cobre e potássio, arenosos a argilosos, e fortemente lixiviados (Chauvel et al. 1987).
2. Delineamento amostral
Foram estabelecidas 12 a 14 parcelas de 250 x 2,5 m por reserva, totalizando 38 parcelas georreferenciadas (Figura 1). O número de parcelas foi determinado de maneira a cobrir de maneira sistemática toda a extensão de cada uma das reservas, mantendo-se uma distância mínima de 1 km entre as parcelas. Cada parcela acompanhou uma curva de nível, para minimizar a variação do solo dentro de uma mesma parcela (Magnusson et al. 2005).
3. Coleta de dados
Em cada parcela registrou-se o número de indivíduos de cada espécie de pteridófita terrestre, hemiepífita ou epífitas estabelecidas até 2 m de altura no forófito. Foram contados apenas indivíduos com frondes maiores que 5 cm de comprimento. Foram excluídas hemiepífitas sem folhas abaixo de 2 m de altura e indivíduos pertencentes às espécies do gênero Selaginella. A reprodução por brotamento em Selaginella spp. resulta na formação de grandes manchas que impossibilitaram o reconhecimento de indivíduos neste estudo. Todos os indivíduos foram mapeados com relação às seguintes faixas de amostragem: de 0 a 1 m de largura, de 1 a 2 m de largura e de 2 a 2,5 m de largura da parcela ( ). Estas faixas foram usadas para gerar os diferentes tamanhos de amostragem analisados. As identificações foram feitas no Herbário do Instituto de Botânica do Estado de São Paulo (IBt-SP). O material testemunho encontra-se depositado no Herbário do INPA (G. Zuquim 1 a G. Zuquim 65).
As variáveis ambientais foram medidas a cada 50 m ao longo do maior eixo da parcela. A inclinação do terreno foi obtida através da média das seis medidas feitas perpendicularmente ao maior eixo da parcela, com um clinômetro Suunto. As estimativas da abertura de dossel e as coletas de solo foram feitas a 90 cm de uma das laterais do maior eixo da parcela (Figura 2). A abertura do dossel foi estimada através de fotos hemisféricas digitais obtidas a 50 cm de altura do solo, entre 5:30 e 8:30 horas e entre 16:00 e 18:00 horas. As imagens foram analisadas com o programa Gap Light Analyzer (Frazer et al. 1999) e a média dos seis valores obtidos por parcela foi utilizada para caracterizar a abertura de dossel de toda a parcela. As coletas de solo foram feitas superficialmente (até 5 cm de profundidade, após remoção da camada de serapilheira) em seis pontos da parcela e misturadas para compor uma única amostra por parcela, independentemente da largura. As análises de granulometria (teor de areia, argila e silte) foram feitas segundo protocolo do Laboratório Temático de Solos e Plantas do Instituto Nacional de Pesquisas da Amazônia, que se baseia nos métodos aplicados pela Empresa Brasileira de Pesquisa Agropecuária (Embrapa). O controle dos procedimentos laboratoriais foi feito através da análise simultânea de solo padrão com características físico-químicas conhecidas que foi fornecido pela Embrapa. Como as variáveis texturais do solo estiveram correlacionadas, apenas a porcentagem de argila foi usada para representar a textura do solo.
4. Obtenção de sub-amostras
A partir dos dados coletados nas 38 parcelas de 250 x 2,5 m de largura, foram feitas sub-amostragens, para determinar os resultados que seriam obtidos utilizando-se parcelas reduzidas a 1 e 2 m de largura. Para tanto, foram criados dois sub-grupos que compreendem sub-amostras do levantamento total. O primeiro sub-grupo inclui somente as plantas amostradas no primeiro metro de largura da parcela. O segundo sub-grupo inclui as plantas amostradas nos dois primeiros metros da largura da parcela. Cada sub-grupo simula diferentes larguras de parcela que podem ser usadas em estudos futuros (Figura 2).
5. Análises estatísticas
A composição de espécies terrestres e hemiepífitas em cada uma das três larguras de parcela foi resumida através de análise multivariada, com a técnica de Análise de Coordenadas Principais (PCoA). Os eixos 1 e 2 da PCoA foram utilizados como variável dependente, em modelos de regressão linear. Regressões múltiplas foram usadas para avaliar a influência das variáveis ambientais (textura do solo, abertura do dossel e inclinação do terreno) sobre a riqueza (número de espécies por parcela), e composição (eixos PCoA1 e PCoA2) de espécies nas parcelas, para a amostra total e para cada uma das amostras com esforço reduzido. A ordenação foi baseada na similaridade entre amostras calculada pelo índice de Bray-Curtis, após relativização das abundâncias das espécies por unidade amostral. Para esta análise da relação entre a composição de espécies e as variações ambientais, excluiu-se as espécies epífitas, pois estas estão em condições ambientais diferentes das espécies que possuem contato com o solo.
O teste de Mantel foi usado para determinar as correlações entre a dissimilaridade florística baseada na amostra total e nas amostras com esforço reduzido. Uma das parcelas foi excluída das análises que envolviam abertura de dossel, por ausência de dados, desta forma o número de parcelas nestas análises foi 37.
Para determinar a forma do aumento no número de espécies e de indivíduos terrestres, epífitos e hemiepífitos à medida que aumenta o número de parcelas amostradas, foram feitas curvas de acumulação de espécies (Rarefação baseada nas Unidades Amostrais, Gotelli & Colwell 2001) para cada sub-amostra e para a amostra total.
As ordenações por PCoA e os testes de Mantel foram feitos no pacote estatístico PATN (Belbin 1992), as regressões no Systat 8.0 (Wilkinson 1998) e as curvas de acumulação no EstimateS 7.5 (Colwell 1997).
Resultados
1. Redução de esforço x retenção de informação
Foram avaliados os resultados que seriam obtidos caso a largura da parcela fosse reduzida para 2 ou para 1 metro, o que representa uma redução de 20 ou 60% da área total amostrada. Considerando todas as espécies terrestres, epífitas e hemiepífitas (exceto Selaginella spp.), não ocorreu estabilização da curva de rarefação, mesmo para a amostra total (2,5 m) (Figura 3). A redução de 60% no tamanho da parcela representou uma diminuição de 52 para 44 espécies. A redução de 20% na largura da parcela representou uma redução de duas espécies, porém, os intervalos de confiança das estimativas do número de espécies para os diferentes tamanhos de amostra se sobrepõem (Figura 3).
As correlações entre as matrizes de distância florística da amostra total (2,5 m) e das sub-amostras contendo 80 e 40% da amostragem total foram altas e significativas (Tabela 1).
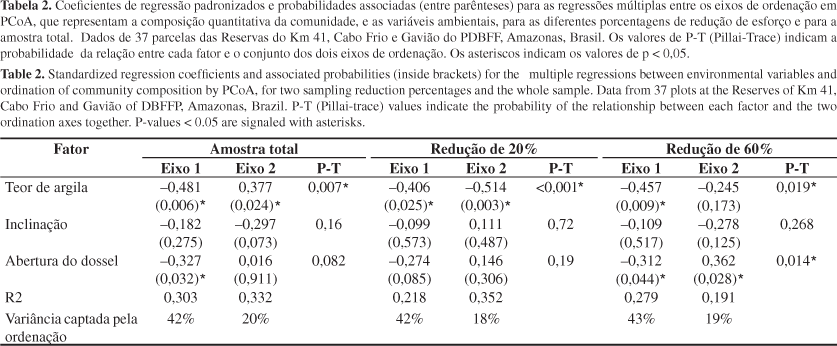
O efeito global (valores de Pillai-Trace) da textura do solo e da inclinação do terreno sobre a comunidade de pteridófitas foi consistente entre todas as larguras de parcela consideradas. O efeito de abertura do dossel sobre a composição de espécies não foi coerente entre as diferentes larguras de parcela. Utilizando um metro como largura de parcela, a abertura do dossel esteve relacionada aos dois eixos da PCoA. Para dois metros de largura, a abertura de dossel não esteve relacionada a qualquer eixo. Para a largura de 2,5 m, a abertura de dossel esteve relacionada apenas ao eixo 1. Para todas as larguras de parcela, o eixo 1 da PCoA captou, aproximadamente o dobro da variação captada pelo eixo 2 (Tabela 2). A composição de espécies, quando representada apenas pelo eixo 1 da PCoA, esteve coerentemente relacionada ao teor de argila do solo em todas as larguras de parcela testadas. A inclinação do terreno não esteve associada à composição de espécies mesmo considerando cada eixo separadamente.
2. Redução de esforço x redução de custo
Reduzir o esforço amostral acarreta em redução de tempo em campo. Considerando apenas a coleta dos dados bióticos, as parcelas de 2,5 m de largura foram amostradas em duas excursões de oito dias e em uma de nove dias, a uma taxa de duas parcelas por dia, que não inclui os dois dias de deslocamento necessários por excursão. Baseado na experiência em campo dos autores, estimou-se que a redução em 60% na largura da parcela possibilitaria a amostragem de uma parcela a mais por dia em campo, reduzindo o tempo total em campo de 25 para 19 dias. As parcelas poderiam ser amostradas em duas excursões de seis dias e em uma de sete, a uma taxa de três parcelas por dia. As parcelas reduzidas em 20% poderiam ser amostradas em 23 dias, a uma taxa de 2,2 parcelas por dia (exceto dias de deslocamento da cidade até a reserva). Cada dia em campo de um pesquisador acompanhado por um auxiliar nas áreas do PDBFF custa US$ 45 (Tabela 3). Portanto, essa redução de 24% no número de dias em excursões a campo causaria uma redução de custo de US$ 125 para US$ 855.
Discussão e Conclusões
A forte correlação da dissimilaridade florística entre as parcelas da amostra total e de sub-amostras das mesmas, indica que mesmo com a redução de esforço amostral, a composição de espécies e a estrutura (distribuição relativa das abundâncias entre espécies) da comunidade continuariam sendo bem representadas. Porém, além de determinar a composição de espécies das comunidades, o delineamento amostral adotado por este trabalho e pelo PPBio foi elaborado para atingir outros objetivos. Um dos principais é a determinação dos fatores ambientais que condicionam a distribuição dos organismos e a variação na composição de espécies de diferentes locais.
O efeito global de argila sobre a composição de espécies de pteridófitas foi coerente para todos os tamanhos de parcela considerados, o que mostra que amostras pequenas são suficientes para captar um efeito ambiental forte. O segundo eixo de PCoA não esteve associado ao teor de argila para a sub-amostra de 1 m, indicando que houve alguma perda de informação com a redução de 60% da amostra. Entretanto, o segundo eixo representa apenas metade da variação da comunidade que foi captada pelo primeiro eixo. Assim, a maior parte da variação na estrutura e composição de espécies foi captada no primeiro eixo de ordenação para todos os tamanhos de amostra, e esta esteve consistentemente relacionada ao solo. O teor de argila é um gradiente ambiental composto que pode representar diversas características do solo, como disponibilidade de nutrientes, pH e capacidade de drenagem (Luizão et al. 2004). Apesar do efeito global do teor de argila ter sido coerente, não é possível descartar a possibilidade de parcelas menores estarem subestimando outros efeitos associados ao solo que não foram investigados no presente trabalho.
Por outro lado, o efeito da abertura de dossel não foi consistente entre diferentes larguras de parcelas. Apesar de significativo para algumas larguras de parcela, a magnitude do efeito de abertura do dossel foi sempre menor do que a do teor de argila. Como a contribuição da abertura do dossel para a estruturação da comunidade é relativamente pequena, a detecção deste efeito deve sofrer mais com o ruído inerente a qualquer amostragem. A comparação entre as parcelas realizada neste estudo situa-se na escala regional, onde se espera que a luz não seja um fator estruturador, por dois motivos: 1) a amplitude de variação da abertura do dossel em florestas densas tropicais é pequena (Chazdon & Fetcher 1984), e 2) a distribuição temporal e espacial da luz é heterogênea mesmo em pequena escala (Clark 2002). Neste trabalho, a média da abertura de dossel (medida a cada 50 m) representa a variação de luz em toda a parcela, mascarando efeitos locais de radiação direta no interior da floresta causada por clareiras ou interrupções no dossel. Portanto há heterogeneidade interna nas parcelas, em qualquer uma das larguras utilizadas. A inconsistência dos resultados na relação entre a composição de espécies de pteridófitas e a abertura de dossel deve estar relacionada à importância relativamente menor da luz na estruturação da comunidade em escala regional.
A redução na largura da parcela para um metro teria reduzido a representação da riqueza de espécies. Porém, o aumento no tamanho da parcela é menos eficiente para captar a riqueza de uma região do que aumento no número de parcelas (Gentry 1982, Phillips et al. 2003). Conforme o tamanho da parcela aumenta, é preciso aumentar muito o número de indivíduos amostrados para acrescentar uma nova espécie à lista. Isso acarreta em maiores custos em campo e em laboratório, construindo planilhas e analisando dados e pode ser pouco produtivo. A curva de acumulação de espécies de uma região raramente estabiliza (Magurran 2004). Magurran & Henderson (2003) observaram após 21 anos de coletas em um canal no Reino Unido, que 61% das espécies eram pouco freqüentes e, detectadas em no máximo um terço dos anos. Além disso, para a compreensão dos fatores que determinam a distribuição de organismos no ambiente, visando o monitoramento da biodiversidade e o planejamento do uso da terra e conservação, o número de espécies não é a medida mais relevante, e sim a identidade associada às abundâncias relativas de cada uma das espécies (Feinsinger 2001, Margules et al. 2002).
Nossos resultados sugerem que amostras com tamanho reduzido, mesmo contendo um número menor de espécies, ainda representam bem a estrutura da comunidade de pteridófitas. Sendo assim, o custo de levantamento pode ser reduzido ou os recursos realocados para a amostragem de um maior número de parcelas.
Em conclusão, a redução de esforço não afeta a capacidade de detectar os principais efeitos ambientais que determinam a estrutura da comunidade de pteridófitas. Porém, a capacidade de detecção dos efeitos ambientais de menor magnitude sobre a composição de espécies pode ser afetada, o que ocorreu com o efeito da abertura de dossel. Dependendo dos objetivos do estudo, parcelas estreitas podem ser muito informativas, especialmente para a descrição dos padrões gerais de composição. Utilizando uma largura de dois metros, estudos futuros na região possivelmente serão capazes de detectar os principais efeitos que determinam a distribuição de pteridófitas em mesoescala.
Agradecimentos
Agradecemos à Gonçalo Ferraz, William E. Magnusson, Ricardo Braga-Neto e Thayná J. Mello, pelas valiosas sugestões. A primeira autora recebeu bolsa da CAPES durante a execução do projeto. Apoio financeiro foi concedido pelo Projeto Dinâmica Biológica de Fragmentos Florestais (PDBFF), Fundação O Boticário de Proteção à Natureza e Projetos Ecológicos de Longa Duração. Esta é a publicação número 487 da série técnica do PDBFF.
Recebido 04/10/06
Versão reformulada recebida em 26/07/07
Publicado em: 25/10/07
ISSN 1676-0603.
- BELBIN, L. 1992. PATN: Pattern Analysis Package. CSIRO, Camberra, Australia.
- BELOVSKY, G.E., BOTKIN, D.B., CROWL, T.A., CUMMINS, K.W., FRANKLIN, J.F., HUNTER JR., M.L., JOERN, A., LINDENMAYER, D.B., MacMAHON, J.A., MARGULES C.R. & SCOTT, J.M. 2004. Ten suggestions to strengthen the science of ecology. Biosc. 54:345-351.
- BRAGA-NETO, R., LUIZÃO, R.C.C.L., MAGNUSSON, W.E., ZUQUIM, G. & CASTILHO, C.V. 2007. Leaf litter fungi in a Central Amazonian forest: the influence of rainfall, soil and topography on the distribution of fruiting bodies. Biod. Cons. XX: Online first. DOI 10.1007/s10531-007-9247-6
- CASTILHO, C. 2005. Variação espacial e temporal da biomassa arbórea viva em 64 km2 de floresta de terra firme da Amazônia Central. Tese de Doutorado. INPA.
- CAUGHLEY, G. & SINCLAIR, A.R.E. 1994. Wildlife management and ecology. Blackwell Science, Cambridge.
- CHAUVEL, A., LUCAS, Y. & BOULET, R. 1987. On the genesis of the soil mantle of the region of Manaus, Central Amazonia, Brasil. Experientia 43:234-240.
- CHAZDON, R.L. & FETCHER, N. 1984. Photoshynthetic light environments in a lowland tropical rainforest in Costa Rica. J. Ecol. 72:553-564.
- CLARK, D.B. 2002. Los factores edáficos y la distribución de las plantas. In Ecología y Conservación de Bosques Neotropicales (eds M. Guariguata & G. Kattan), p. 193-224. Ediciones LUR, Cartago.
- COLWELL, R.K. 1997. EstimateS: Statistical estimation of species richness and shared species from sample. Version 5. Users guide and application. http://viceroy.eeb.uconn.edu/estimates
- CONDIT, R. 1998. Tropical forest census plots: Methods and results from Barro Colorado Island, Panama and a comparison with other plots. Springer-Verlag and R.G. Landes Company. 211p.
- CONDIT, R., PITMAN, N., LEIGH Jr., E.G., CHAVE, J., TERBORGH, J., FOSTER, R.B., NUNEZ, P., AGUILAR, V.S., VALENCIA, R., VILLA, G., MULLER-LANDAU, H.C., LOSOS, H. & HUBBELL, S.P. 2002 Beta diversity in tropical forest trees. Science 295:666-669.
- COSTA, F.R.C., MAGNUSSON, W.E. & LUIZÃO, R.C. 2005 Mesoscale distribution patterns of Amazonian understorey herbs in relation to topography, soil and watersheds. J. Ecol. 93:863-878.
- COSTA, F.R.C. 2006. Mesoscale Gradients of Herb Richness and Abundance in Central Amazonia. Biotropica. 23:171-177
- FRAZER, G.W., CANHAM, C.D. & LERTZMAN, K.P. 1999. Gap Light Analyzer (GLA), Version 2.0: Imaging software to extract canopy structure and gap light transmission indices from true-colour fisheye photographs, users manual and program documentation. British Columbia and New York, USA. www.ecostudies.org/gla
- FEINSINGER, P. 2001. Designing field studies for biodiversity conservation. Island Press. Washington.
- GENTRY, A.H. 1982. Patterns of neotropical plant species diversity. Evol. Biol. 15:1-84.
- GENTRY, A. H. 1988a. Tree species richness of upper Amazonian forests. Proc. Natl. Acad. U.S.A. 85:156-159.
- GENTRY, A. H. 1988b. Changes in plant community diversity and floristic composition of environmental and geographical gradients. Ann. Missouri Bot. Gard. 75:1-34
- GOTELLI, N.J. & COLWELL, R.C. 2001. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 4:379-391.
- HAYEK, L. A.C. & BUZAS, M.A. 1997. Surveying natural populations. Columbia University Press. NY.
- HUBBELL, S.P. 2001. The unified neutral theory of biodiversity and biogeography. Princeton Univ. Press, New Jersey.
- JONES, M.M., TUOMISTO, H., CLARK, D.B. & OLIVAS, P. 2006. Effects of mesoscale environmental heterogeneity and dispersal limitation on floristic variation in rain forest ferns. Journal of Ecology 94:181-195.
- LUIZÃO, R.C.C., LUIZÃO, F.J., PAIVA, R.Q., MONTEIRO, T.F., SOUSA, L.S. & KRUIJ, B. 2004. Variation of carbon and nitrogen cycling processes along a topographic gradient in a central Amazonian forest. Global Change Biology, 10, 592-600.
- MAGNUSSON, W.E. & MOURÃO, G. 2003. Estatística sem matemática. Editora Planta, Londrina.
- MAGNUSSON, W.E., LIMA, A.P., LUIZÃO, R., LUIZÃO, F., COSTA, F.R.C., de CASTILHO, C.V. & KINUPP, V.P. 2005. RAPELD: a modification of the Gentry method for biodiversity surveys in long-term ecological research sites. Biota Neotropica 5(2). http://www.biotaneotropica.org.br/v5n2/pt/abstract?point-of-view+bn01005022005
- MAGURRAN, A.E. 2004. Measuring biological diversity. Blackwell Publishing. United Kingdon.
- MAGURRAN, A.E. & HENDERSON, P.A. 2003. Explaining the excess of rare species in natural species abundance distributions. Nature 422:714716.
- MARGULES, C.R., PRESSEY, R.L. & WILLIAMS, P.H. 2002. Representing biodiversity: data and procedures for identifying priority areas for conservation. J. Bios. 27:309-326.
- NOGUEIRA, A. 2006. Variação da densidade, área basal e biomassa de lianas em 64 km2 de floresta de Terra-Firma da Amazônia Central. Dissertação de Mestrado. INPA.
- OLIVEIRA, A.A. & MORI, S. 1999. A central Amazonian terra firme forest I. High tree species richness on poor soils. Biod. Cons. 8:1219-1244.
- PHILLIPS, O.L., MARTÍNEZ, R.V., VARGAS, P.N., MONTEAGUDO, A.L., ZANS, M-E.C., SÁNCHEZ, W.G., CRUZ, A.P., TIMANÁ, M., YLI-HALLA, M. & ROSE, S. 2003. Efficient plot-based floristic assessment of tropical forests. J. Trop. Ecol. 19:629-645.
- PITMAN, N.C.A., TERBORGH, J.W., MILES, R. SILMAN, P.N.V., NEILL, D.A., CERÓN, C.E., PALACIOS, W.A. & AULESTIA, M. 2001. Dominance and distribution of tree species in upper Amazonian terra firme forests. Ecol. 82:2101-2117
- PITMAN, N. C.A., TERBORGH, J.H., SILMAN, M.R., NUNEZ, P., NEILL, D.A., CERON, C.E., PALACIOS, W.A. & AULESTIA, M. 2002. A comparison of tree species diversity in two upper Amazonian Rain Forests. Ecol. 83:3210-3224.
- PRANCE, A.H., RODRIGUEZ, W.A. & SILVA, M.F. 1976. Inventário florestal de um hectare de mata de terra firme km. 30 da estrada Manaus-Itacoatiara. Acta Amazonica 6:9-35.
- RADAMBRASIL. 1978. Levantamento de Recursos Naturais. Vols.1-18. Ministério das Minas e Energia. Departamento Nacional de Produção Mineral, Rio de Janeiro.
- RICKLEFS, R.E. 2004. A comprehensive framework for global patterns on biodiversity. Ecol. Lett. 7:1-15.
- SANTOS, E.M. 2001. Densidade, diversidade e biomassa da fauna do solo em serrapilheira manipulada numa floresta secundária na Amazônia Central. Dissertação de Mestrado. INPA.
- SVENNING, J-C, ENGELBRECHT, B.M.J., KINNER, D.A., KURSAR, T.A., STALLARD, R.F. & WRIGHT, S.J. 2006. The relative roles of environment, history and local dispersal in controlling the distributions of common tree and shrub species in a tropical forest landscape, Panama. J. Trop. Ecol. 22:575-586.
- TUOMISTO, H., RUOKOLAINEN, K., POULSEN, A.D.; MORAN, R.C., QUINTANA, C., CAÑAS, G. & CELI, J. 2002. Distribution and diversity of pteridophytes and Melastomataceae along edaphic gradients in Yasuní national park, Ecuadorian amazonia. Biotropica 34:516-533.
- TUOMISTO, H., RUOKOLAINEN K. & YLI-HALLA, M. 2003a. Dispersal, environmental, and floristic variation of Western Amazonian forests. Science 299:241-244.
- TUOMISTO, H., RUOKOLAINEN, K., AGUILAR. M. & SARMIENTO, A. 2003b. Floristic patterns along a 43-km long transect in an Amazonian rain forest. J. Ecol. 91:43-756.
- VORMISTO, J., TUOMISTO, H. & OKSANEN, J. 2004. Palm distribution patterns in Amazonian rainforests: What is the role of topographic variation? J. Veg. Sci. 15:485-494.
- WILKINSON, L. 1998. Systat: the system for statistics. Systat Inc. Evanston, Illinois, USA.
Datas de Publicação
-
Publicação nesta coleção
12 Mar 2008 -
Data do Fascículo
2007
Histórico
-
Aceito
25 Out 2007 -
Recebido
04 Out 2006 -
Revisado
26 Jul 2007