Abstract
Terpenes are a very large and structurally diverse group of secondary metabolites which are abundant in many essential oils, resins and floral scents. Additionally, some terpenes have roles as phytoalexins in plant-pathogen relationships, allelopathic inhibitors in plant-plant interactions, or as airborne molecules of plant-herbivore multitrophic signaling. Thus the elucidation of the biochemistry and molecular genetics of terpenoid biosynthesis has paramount importance in any crop species. With this aim, we searched the CitEST database for clusters of expressed sequence tags (ESTs) coding for terpene synthases. Herein is a report on the identification and in silico characterization of 49 putative members of the terpene synthase family in diverse Citrus species. The expression patterns and the possible physiological roles of the identified sequences are also discussed.
secondary metabolism; flavor; aroma; plant protection; terpene synthesis
BIOCHEMICAL PROCESSES
RESEARCH ARTICLE
A genomic approach to characterization of the Citrus terpene synthase gene family
Marcelo Carnier Dornelas; Paulo Mazzafera
Departamento de Fisiologia Vegetal, Instituto de Biologia, Universidade Estadual de Campinas, Campinas, SP, Brazil
ABSTRACT
Terpenes are a very large and structurally diverse group of secondary metabolites which are abundant in many essential oils, resins and floral scents. Additionally, some terpenes have roles as phytoalexins in plant-pathogen relationships, allelopathic inhibitors in plant-plant interactions, or as airborne molecules of plant-herbivore multitrophic signaling. Thus the elucidation of the biochemistry and molecular genetics of terpenoid biosynthesis has paramount importance in any crop species. With this aim, we searched the CitEST database for clusters of expressed sequence tags (ESTs) coding for terpene synthases. Herein is a report on the identification and in silico characterization of 49 putative members of the terpene synthase family in diverse Citrus species. The expression patterns and the possible physiological roles of the identified sequences are also discussed.
Key words: secondary metabolism, flavor, aroma, plant protection, terpene synthesis.
Introduction
Terpenes are found widely distributed in the plant kingdom, from lichens and algae to higher plants. Although terpenes cover a wide range of compounds with diverse structure, the word terpene has been frequently associated with essential oils, which are volatile compounds belonging to some terpene classes. Terpenes have several applications in the food and cosmetic industry in the production of flavors and fragrances (Croteau et al., 2000; Phillips et al., 2006; Tholl, 2006).
Terpenes are classified according to the following classes: hemiterpenes (C5H8); monoterpenes (C10H16); sesquiterpenes (C15H24); diterpenes (C20H32); sesterterpenes (C25H40); triterpenes (C30H48); tetraterpenes (C40H64) and polyterpenes [(C5H8)n]. Hemiterpenes are not found in appreciable amounts as free compounds but are usually bound to other non-terpene compounds, such as the purine base adesonine and the plant hormone zeatin (Goodwin, 1967; Croteau et al., 2000; Phillips et al., 2006; Tholl, 2006). Monoterpenes consist of two condensed basic units of isopentenyl pyrophosphate (IPP). Accordingly, sesquiterpenes have three, diterpenes four, sesterterpenes five, triterpenes six and tetraterpenes eight IPP molecules, respectively. Polyterpenes are all terpenes containing more than eight isoprene units, which include all natural rubbers (Goodwin, 1967; Croteau et al., 2000).
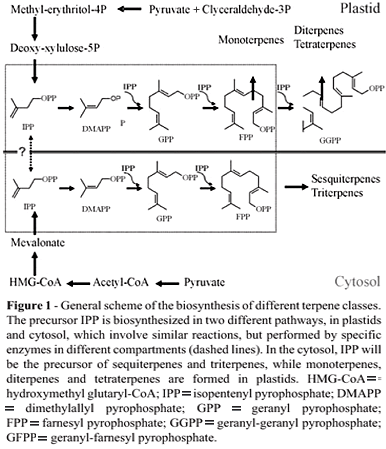
Elucidation of the biochemistry and molecular genetics of terpenoid biosynthesis has made rapid progress in recent years (Rohdich et al., 2005). The genes coding for the main enzymes involved in this biological process are being identified, and all the members of the Arabidopsis terpene synthase gene family have been characterized following the sequencing of the whole genome of this model plant (Aubourg et al., 2002). Figure 1 shows the general scheme for the synthesis of the precursor molecules of the main terpene classes. The starting reaction is the isomerization of IPP to dimethylallyl pyrophosphate. Both compounds condense to form the first parental structure, geranyl-PP. Subsequently, the other structures are formed by sequential addition of IPP (Phillips et al., 2006; Tholl, 2006). Two farnesyl-PP are condensed to form the parent molecule squalene, the precursor of triterpenes, and two geranyl-PP are condensed to form phytoene, the parent molecule for tetraterpenes (Goodwin, 1967; Croteau et al., 2000; Rohdich et al., 2005; Phillips et al., 2006; Tholl, 2006).
The biological functions of the many terpene molecules in plants are linked not only to the biosynthesis of hormones, but also to protection against UV radiation and photo-oxidative stress. Additionally, terpenes are also related to thermal protection, pollinator attraction, membrane stabilization, resistance against insects and microorganisms, plant-plant signaling, etc. (Steele et al., 1998; Trapp and Croteau, 2001; Copolovici et al., 2005; Baldwin et al., 2006; Keeling and Bohlmann, 2006).
In Citrus spp, terpene molecules belonging to different classes are produced especially in leaves, fruit epidermis (flavedo) and fruit juice. These terpenes have special economic interest, as they are the main components of Citrus essential oils and some of them (carotenoids) give the Citrus juice its special color. Additionally, carotenoids are well known to be important to human health. Several biotechnological approaches have been taken to increase such compounds in food (Botella-Pavia and Rodriguez-Concepcion, 2006). There are several reports on the composition of terpenes in several Citrus species, mainly regarding the essential oil composition (Ruberto and Rapisarda, 2002; Sawamura et al., 2005; Verzera et al., 2005). The aromatic components of citrus are classified in two categories: those present in the oil from the flavedo and juice, and those soluble in the water and components of the juice. Monoterpene d-limonene is the main component of oil from the flavedo, with concentrations over 85% of the oil fraction. In the Pêra variety of orange, the concentration may reach up to 93% while in the Tahiti variety of lime it ranges from 50 to 60%. In addition to d-limonene, other terpenes found in the flavedo oil fraction are linalool, geraniol, citronellol, a-terpineol, valencene, mircene, a-pinene, etc. (Ruberto and Rapisarda, 2002; Sawamura et al., 2005; Verzera et al., 2005).
Thus, due to the importance of Citrus terpenes to the Brazilian economy (Boteon and Neves, 2005), we undertook a genomic approach for the characterization of the Citrus terpene synthase gene family by analyzing the CitEST database of Citrus expressed sequence tags. We have identified 49 putative members of the terpene synthase family in diverse Citrus species and we suggest their possible biological roles based on their expression patterns and on sequence comparisons with other terpene synthases that have already been functionally characterized in other plant species.
Material and Methods
Searching Citrus EST homologs for terpene synthases
The clustered expressed sequence tags (ESTs) from the CitEST project database were used as a primary source of data for our analyses. These sequences were assembled from ESTs obtained from the sequencing of several Citrus spp. cDNA libraries, made from different tissues and various physiological states (see other papers in this issue for details on library construction and sequencing). Nucleotide and amino acid sequences from other terpene synthase genes were obtained from The National Center for Biotechnology Information (NCBI). Searches for terpene synthase sequences in the CitEST database were conducted using the tBLASTN module that compares the consensus amino acid sequence with a translated nucleotide sequences database (Altschul et al., 1997). We used as a query a consensus terpene synthase sequence obtained by aligning all Arabidopsis thaliana terpene synthase protein sequences (Aubourg et al., 2002). All sequences that exhibit a significant alignment (e-value lower than 10-5) with the consensus were retrieved from the CitEST database. All retrieved sequences were then re-inspected for occurrence of terpene synthase conserved motives using the InterProScan.
Phylogenetic analysis and expression patterns of putative Citrus terpene synthases
Amino acid sequences were used for all the phylogenetic analyses. Sequence alignments were performed With ClustalX (Thompson et al., 1994) using default parameters, but the final alignment was visually inspected and manually corrected. The MEGA software, version 2.0 (Kumar et al., 2000) was used for the phylogenetic analysis. Average p-distances were high so the Poisson model was used to provide unbiased estimates of the number of substitutions between sequences. Phylogenetic trees were obtained using parsimony and/or genetic distance calculations. Neighbor-joining (Saitou and Nei, 1987) and Bootstrap (with 1000 replicates) trees were also constructed.
For each EST-contig, the frequency of reads in the selected libraries was calculated. This procedure requires a normalization that is accomplished by dividing the total number of reads in the specific library by the total number of reads in all libraries and then dividing the number of reads of each EST-contig by the ratio found for each library. The results were cast in a matrix and a hierarchical clustering was performed, using the Cluster and Tree View programs (Eisen et al., 1998). The pattern of gene expression was displayed as color-coded arrays of EST-contigs, using a color scale representing the number of reads from a specific library in each EST-contig.
Results
Identifying Citrus spp putative terpene synthase genes
Despite the fact that the whole genome of the model plant Arabidopsis thaliana had been completely sequenced four years ago, the function of only three, out of the 32 Arabidopsis terpene synthase genes (Aubourg et al., 2002), has been described to date. These are the AtTPSGA1 gene for copalyl diphosphate synthase (Sun and Kamiya, 1994) and the AtTPSGA2 transcript which encodes kaurene synthase (Yamaguchi et al., 1998), both of which are involved in the formation of gibberellins, and the AtTPS10 transcript for myrcene/ocimene synthase (Bohlmann et al., 2000) required for the formation of acyclic monoterpenes. In an initial attempt to identify the members of the Citrus terpene synthase gene family, we have performed an in silico screen of the CitEST database for putative terpene synthase sequences. This exhaustive sequence search detected 49 unique assembled Citrus spp EST sequences.
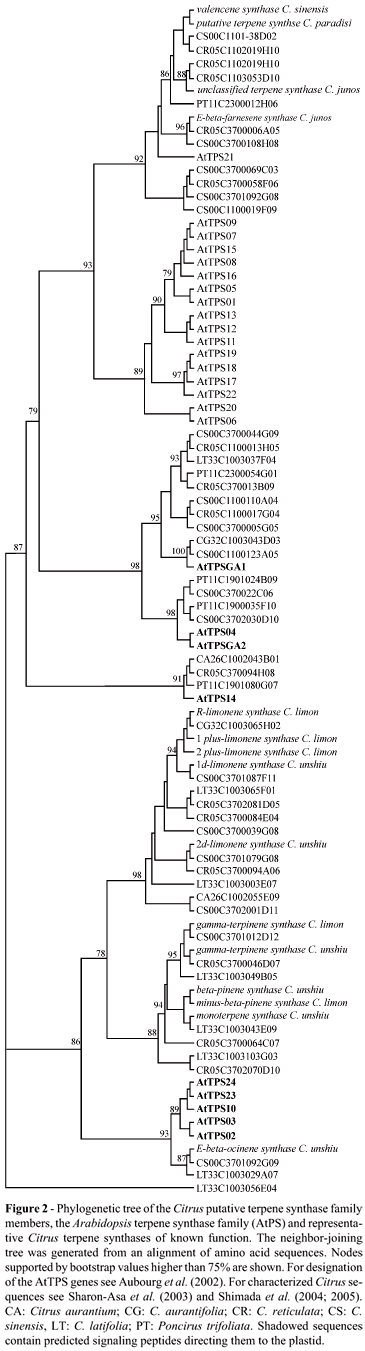
We started our analysis by detecting conserved sequence motifs within the putative Citrus terpene synthase proteins. In pairwise comparisons of all predicted Citrus terpene synthases to all Arabidopsis AtTPS proteins, overall sequence identity varies widely from 18% (LT33-C1-003-056-E04 and AtTPS21) to 91% (CS00-C1-100-123-A05 and AtTPGA1). These sequence comparisons allowed the construction of the distance-based tree shown in Figure 2. For this analysis, we considered all Citrus spp EST clusters found within the CitEST database (we used the name of the founder EST sequence to name the entire sequence cluster), all Arabidopsis TPS and all publicly available Citrus terpene synthase sequences available at GenBank (as of May 2006). We adopted the separation of the terpene synthases into classes, as suggested by Aubourg et al. (2002).
When the putative Citrus terpene synthase EST contigs were long enough to allow the complete encoded protein sequences to be deduced, their size ranged from 547 to 617 amino acids, which corresponds to the size of known monoterpene synthases, sesquiterpene synthases and diterpene synthases of secondary metabolism (Bohlmann et al., 1998; Aubourg et al., 2002). Variation in length within this class of terpene synthase could be attributed to the presence or absence of putative plastid transit peptides (Figure 1).
Most terpene synthases encoded by class-III genes contain variations of a conserved motif, RR(x)8 W, close to the N-terminus (Bohlmann et al., 1998; Aubourg et al., 2002). The RR(x)8 W motif is absolutely conserved in most Citrus sequences that resemble typical monoterpene synthases. There was a clade within this group that contained only Arabidopsis-derived sequences (AtTPS02, AtTPS 03, AtTPS 10, AtTPS 23 and AtTPS 24; see Figures 2 and 3) and it has been reported that these proteins contain variations on the RR(x)8 W motif; however, these variations have as yet unknown biological implications (Aubourg et al., 2002).
Attributing putative functions to Citrus putative terpene synthases by sequence comparisons
Comparison of the predicted Citrus terpene synthase proteins with homologs of known function from other species allowed the identification of putative orthologues that evolved from a common ancestral gene by speciation and retained the same or similar biological function in different species during the course of evolution (Tatusov et al., 1997; Huynen and Bork, 1998). The deduced Citrus terpene synthase proteins were compared with more than 40 terpene synthases from over 20 different species, including monocotyledonous and dicotyledonous species and gymnosperms, to determine sequence identity. A neighbor-joining tree was constructed based on multiple sequence alignment (Figure 3). Six subfamilies of the plant terpene synthase family, designated TPS-a through TPS-f, were previously defined based on clusters identified in the phylogeny (Bohlmann et al., 1997; 1998; Aubourg et al., 2002). Sequence relatedness places all Citrus putative terpene synthases into the previously defined angiosperm terpene synthase subfamilies (Figure 3). Most Citrus sequences cluster in the class III. The terpene synthases from this group contain all known sesquiterpene and diterpene synthases of the secondary metabolism from angiosperms. It is therefore most likely that the Citrus members of this group are also sesquiterpene synthases or diterpene synthases, rather than monoterpene synthases. The lack of transit peptides in some of the Citrus proteins from this group is reminiscent of previously characterized sesquiterpene synthases of the TPS-a group (Bohlmann et al., 1998; Aubourg et al., 2002).
Class I terpene synthases include AtTPS GA1 (Sun and Kamiya, 1994), which is a member of the TPS-c group of angiosperm copalyl diphosphate synthases. This class also includes the AtTPS GA2 enzyme (Yamaguchi et al., 1998), which is a diterpene synthase of the TPS-e subfamily of kaurene synthases. These terpene synthases are involved in the biosynthesis of gibberellic acid and putative Citrus ortologues were found for both of them. The Class I sub-clade, which includes the highly divergent AtTPS14, also contains terpene synthases from three different Citrus species that share the conserved DDxxD motif. Finally, Class I also includes AtTPS04 of which the primary structure is reminiscent of that of linalool synthase from Clarkia breweri (Dudareva et al., 1996).
Expression patterns of Citrus putative terpene synthases
To investigate whether the expression patterns of putative Citrus terpene synthases were biased towards a certain organ and/or tissue, we performed an in silico Northern in order to determine the relative abundance of putative terpene synthase-coding transcripts among different CitEST cDNA libraries. The results, shown in Figure 4, indicate a preferential accumulation of transcripts in leaves, and in the fruit flavedo, especially during the early stages of fruit development. These results are in agreement with observations of terpene accumulation and essential oil production by these tissues in Citrus plants (Ruberto and Rapisarda, 2002; Sawamura et al., 2005; Verzera et al., 2005).
Citrus putative terpene synthase sequences were present at extremely low frequencies (a single EST among all CitEST sequences, as in the case of PT11-C1-901-080-G07) or at relatively high proportions (more than one hundred ESTs, in the case of CR05-C3-700-046-D07), indicating that the terpene synthase family members are differentially expressed among Citrus species, as well as in different tissues and developmental stages (Figure 4).
Discussion
The duplication followed by divergence of terpene synthase genes are central to the biosynthesis of hundreds of basic terpenoid skeletons derived from only four prenyl-diphosphate intermediates of the isoprenoid pathway (Figure 1; Davis and Croteau, 2000). Evolution of a large terpene synthase family reflects, at the genetic level, the structural diversity of terpenoid natural products and their roles in ecological plant interactions. Earlier phylogenies of plant terpene synthases established characteristic features for this family: clustering of terpene synthases into at least six subfamilies; independent evolution of specific catalytic functions of terpene synthases in gymnosperms and angiosperms; the presence of a 200 amino acid motif in an ancestor of angiosperm and gymnosperm terpene synthases and divergence of terpene synthase genes involved in secondary metabolism as the consequence of gene duplication and functional diversification (Bohlmann et al., 1998; Aubourg et al., 2002).
In the present analysis, a protein phylogeny was combined with novel genomic information generated from mining the data provided by the CitEST database of Citrus expressed sequence tags. Due to the importance of Citrus terpenes in aroma, flavor and the juice industries, we set out to characterize the Citrus spp terpene synthase gene family. We successfully identified 49 putative Citrus terpene synthase coding sequences derived from six different species (Figure 2). In most cases, sequence comparison analyses revealed that some terpene synthases showed higher similarity among different Citrus species, indicating a possible conservation of their biological roles within these species. This observation also suggests that the different classes of terpene synthases diverged before Citrus speciation events and that the last ancestor of all Citrus species analyzed possessed at least one member of each known terpene synthases classes. For instance, CS00-C1-100-123-A05 showed higher similarity to CG32-C1-003-043-D03 from C. aurantifolia than to any other terpene synthase from C. sinensis.
The position of three conifer terpene synthase sequences within the present protein phylogeny (Figure 3) is in agreement with previous analyses of terpene synthase family evolution (Bohlmann et al., 1998; Aubourg et al., 2002). Nevertheless, more terpene synthase sequences from gymnosperms and lower nonvascular plants are required for a better understanding of their phylogenetic location relative to the large number of known angiosperm tepene synthases. On the other hand, the placement of all Citrus spp putative terpene synthases in the protein phylogeny is well supported and might be indicative of their potential biological roles (Figure 3). For instance, the Class III sub-clade containing well-characterized limonene synthases also includes many clusters from five different Citrus species, which might indicate that these clusters code for putative limonene synthase orthologs from these species.
Sequence comparisons of all Citrus putative terpene synthases found in the CitEST database with terpene synthases from other plant species allowed the prediction of their putative preferential substrates and thus, suggest their potential biological roles (Figure 3). Among the Citrus putative terpene synthase coding sequences that we have found, most of them were preferentially expressed in the leaves and in the fruit flavedo, in agreement with their expected putative roles in terpene biosynthesis (Figure 4). Apart from being an important constituent of Citrus essential oils, some reports have described the activation of terpene synthase gene expression in defense responses against insects and pathogens. For example, in cotton, sesquiterpene phytoalexins are elicited in response to bacterial or fungal infection. Chen et al. (1995) observed that Gossypium arboreum cell suspension culture showed an increase of transcripts of a (+)-delta-cadinene synthase when challenged by a preparation from Verticillium dahliae. The authors concluded that such observation was consistent with a role for this enzyme as the first step in the pathways leading to the biosynthesis of phytoalexins gossypol and lacinilene C in cotton. Accordingly, tent caterpillars feeding on leaves of hybrid poplar induced local and systemic emissions of (-)-germacrene D, (E)-b-ocimene, linalool, (E)-4,8-dimethyl-1,3,7-nonatriene, benzene cyanide, and (E,E)-a-farnesene (Arimura et al., 2004b). This emission of volatile terpene compounds was correlated with an increase in transcription levels of a sesquiterpene synthase greatly induced in response to herbivory (Arimura et al., 2004a). We found putative Citrus homologs to terpene synthases expressed preferentially in leaves (Figure 4). However, it remains speculative whether their biological function is related to the plant signaling system induced by pest and/or pathogen attack.
Transcripts of a valencene synthase were found to accumulate in fruits of C. sinensis only towards maturation, contributing to the accumulation of valencene (Sharon-Asa et al., 2003). Curiously, Citrus fruits are non-climacteric but valencene and its synthase were induced by ethylene, indicating that this hormone may play a role at the final stages of Citrus fruit maturation.
Within the CitEST frame, six libraries were made from the flavedo tissue of fruits at different developmental stages, ranging from 1 to 9 cm diameter. Only two C. reticulata EST clusters, CR05-C3-700-084-E04 and CR05-C3-700-046-D07), showed a significant increase in EST abundance correlated with developmental progress up to the third developmental stage, fruits of 5 cm in diameter (Figure 4). The putative orthologs of these sequences in other Citrus species analyzed did not show this behavior, indicating that this up-regulation during fruit development may be a characteristic of C. reticulata fruit.
Cluster CS00-C3-700-022-C06 showed very high similarity to cycloartenol synthase, which converts oxidosqualene to cycloartenol, a pentacyclic isomer of the animal and fungal sterol precursor lanosterol. Plants cyclize oxidosqualene to cycloartenol as the initial sterol (Corey et al., 1993). Other clusters/genes closely related within this same clade (Figure 3) are also apparently involved with steroid biosynthesis. Steroids are structural components of membranes and are very important for membrane fluidity. Brassinosteroids, a class of hormones, are also exclusively formed by the terpene metabolism (Croteau et al., 2000). Thus it would be interesting to test whether the protein predicted to be coded by cluster CS00-C3-700-022-C06 is indeed involved with steroid biosynthesis in Citrus.
The cluster CA26-C1-002-055-E09 showed high similarity to an isoprene synthase from Pueraria montana (Sharkey et al., 2005). Isoprene is formed from dimethylallyl-PP and its emission from leaves can significantly influence the surrounding atmosphere. Claeys et al. (2004) showed that photo-oxidation of isoprene emitted from plants in the Amazon was sufficient to influence the rain regime in the region. Additionally, isoprene emission has been reported to protect plants against high temperatures particularly during rapid temperature fluctuation periods (Velikova and Loreto, 2005). Future work on the putative substrate of the enzyme coded by cluster CA26-C1-002-055-E09 might help with the elucidation of its biochemical function.
We found many Citrus spp clusters showing high levels of similarity with limonene synthases and g-terpinene synthases (Figure 3). We speculate that these transcripts might represent Citrus orthologs for limonene synthases, as limonene is the most abundant terpene in Citrus spp essential oils. Figure 4 shows that contigs CR05C3700084E04 and CR05C3700046D07, which showed high similarity to both limonene and g-terpinene synthases, are highly expressed in the flavedo, especially during the first three stages of fruit development. This was also observed for d-limonene synthase (Shimada et al., 2005) and g-terpinene synthase (Shimada et al., 2004) genes isolated from C. unshiu. The transcripts of these genes were reported to accumulate in the fruit peel at the early stages of fruit development (Shimada et al., 2004; 2005).
It was surprising to discover that some putative Citrus terpene synthases such as those coded by Poncirus sequences PT11-C2-300-054-G01 and PT11-C2-300-012-H06 were apparently exclusively expressed in bark tissue (Figure 4). Nevertheless, it has been recently suggested that expression of limonene synthase may be related with induced oleoresinosis response against the white pine weevil attacking Sitka spruce trees (Byun-McKay et al., 2006). Oleoresin is a mixture of turpentine (85% monoterpenes and 15% sesquiterpenes) and rosin (diterpene resin acids) that seal wounds and is toxic to both invading insects and their pathogenic fungal symbionts (Steele et al., 1998). Of course, this attempt to attribute a putative function to the proteins predicted to be coded by sequences PT11-C2-300-054-G01 and PT11-C2-300-012-H06 remains speculative, but it would be interesting to find out whether their expression is indeed bark-specific.
Conclusions and Perspectives
This initial characterization of a large number of putative members of the Citrus spp terpene synthase gene family provides novel resources for research on terpene secondary metabolism in Citrus species. We report here the largest number of putative terpene synthase sequences ever published for a single plant genus. We have characterized 49 sequence contigs that might represent 49 different genes, although the exact number of members of the Citrus terpene synthase gene family will be established only when a complete genome sequence is available for Citrus. Together with the recent characterization of the complete terpene synthase gene family for the model plant Arabidopsis (Aubourg et al., 2002) and the functional characterization of some of its members (Bohlmann et al., 2000; van Poecke et al., 2001), our findings add exciting new aspects to our concept of secondary metabolism in Citrus and open several new avenues for natural product research directed by genome analysis. The expression patterns of some of the Citrus putative terpene synthases presented here will be tested in future work by using a combination of in situ hybridization and functional experiments.
Acknowledgments
We would like to thank Marcos Machado for coordinating our efforts on the analysis of the data generated by the CITEST Project, Luiz Humberto Gomez for help in early stages of this work and CNPq and FAPESP (Brazil) for financial support. The authors would also like to thank CNPq for research fellowships.
Internet Resources
Received: July 21, 2006; Accepted: April 2, 2007.
Associate Editor: Marco Aurélio Takita
References
- Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W and Lipman DJ (1997) Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res 25:3389-3402.
- Arimura G, Huber DP and Bohlmann J (2004a) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides): cDNA cloning, functional characterization, and patterns of gene expression of (-)-germacrene D synthase, PtdTPS1. Plant J 37:603-616.
- Arimura G, Ozawa R, Kugimiya S, Takabayashi J and Bohlmann J (2004b). Herbivore-induced defense response in a model legume: Two-spotted spider mites induce emission of (E)- b-ocimene and transcript accumulation of (E)-b-ocimene synthase in Lotus japonicus Plant Physiol 135:1976-1983.
- Aubourg S, Lecharny A and Bohlmann J (2002) Genomic analysis of the terpenoid synthase (AtTPS) gene family of Arabidopsis thaliana Mol Genet Genomics 267:730-745.
- Baldwin IT, Halitschke R, Paschold A, Von Dahl CC and Preston CA (2006) Volatile signaling in plant-plant interactions: "Talking trees" in the genomics era. Science 311:812-815.
- Bohlmann J, Meyer-Gauen G and Croteau R (1998) Plant terpenoid synthases: Molecular biology and phylogenetic analysis. Proc Natl Acad Sci USA 95:4126-4133.
- Bohlmann J, Steele CL and Croteau R (1997) Monoterpene synthases from grand fir (Abies grandis): cDNA isolation, characterization, and functional expression of myrcene synthase, (-)-(4S)-limonene synthase, and (-)-(1S,5S)-pinene synthase. J Biol Chem 272:21784-21792.
- Bohlmann J, Meyer-Gauen G and Croteau R (1998) Plant terpenoid synthases: Molecular biology and phylogenetic analysis. Proc Natl Acad Sci USA 95:4126-4133.
- Bohlmann J, Martin D, Oldham NJ and Gershenzon J (2000) Terpenoid secondary metabolism in Arabidopsis thaliana: cDNA cloning, characterization, and functional expression of a myrcene/(E)-b-ocimene synthase. Arch Biochem Biophys 375:261-269.
- Botella-Pavia P and Rodriguez-Concepcion M (2006) Carotenoid biotechnology in plants for nutritionally improved foods. Physiol Plant 126:369-381.
- Boteon M and Neves EM (2005). Citricultura brasileira: Aspectos econômicos. In: Mattos Jr D, De Negri JD and Pompeu Jr J (eds) Citrus. Instituto Agronômico de Campinas, Campinas, pp 19-36.
- Byun-McKay A, Godard KA, Toudefallah M, Martin DM, Alfaro R, King J, Bohlmann J and Plant AL (2006) Wound-induced terpene synthase gene expression in sitka spruce that exhibit resistance or susceptibility to attack by the white pine weevil. Plant Physiol 140:1009-1021.
- Chen XY, Chen Y, Heinstein P and Davisson VJ (1995) Cloning, expression, and characterization of (+)-delta-cadinene synthase: A catalyst for cotton phytoalexin biosynthesis. Arch Biochem Biophys 324:255-266.
- Claeys M, Graham B, Vas G, Wang W, Vermeylen R, Pashynska V, Cafmeyer J, Guyon P, Andreae MO, Artaxo P, et al. (2004) Formation of secondary organic aerosols through photo-oxidation of isoprene. Science 303:1173-1176.
- Copolovici LO, Filella I, Llusia J, Niinemets U and Penuelas J (2005) The capacity for thermal protection of photosynthetic electron transport varies for different monoterpenes in Quercus ilex Plant Physiology 139:485-496.
- Corey EJ, Matsuda SPT and Bartel B (1993) Isolation of an Arabidopsis thaliana gene encoding cycloartenol synthase by functional expression in a yeast mutant lacking lanosterol synthase by the use of a chromatographic screen. Proc Natl Acad Sci USA 90:11628-11632.
- Croteau R, Kutchan TM and Lewis NG (2000) Natural products In: Buchanan B, Gruissem W and Jones R (eds) Biochemistry and Molecular Biology of Plants. American Society of Plant Physiology, New York, pp 1250-1318.
- Davis EM and Croteau R (2000) Cyclization enzymes in the biosynthesis of monoterpenes, sesquiterpenes, and diterpenes. Curr Topics Chem 209:53-95.
- Dudareva N, Cseke L, Blanc VM and Pichersky E (1996) Evolution of floral scent in Clarkia: Novel patterns of S-linalool synthase gene expression in the C. breweri flower. Plant Cell 8:1137-1148.
- Eisen MB, Spellman PT, Brown PO and Botstein D (1998) Cluster analysis and display of genome-wide expression patterns. Proc Natl Acad Sci USA 95:14863-14868.
- Goodwin TW (1967) Terpenoids in Plants. Academic Press, New York, 326 pp.
- Huynen MA and Bork P (1998) Measuring genome evolution. Proc Natl Acad Sci USA 95:5849-5856.
- Keeling CI and Bohlmann J (2006) Genes, enzymes and chemicals of terpenoid diversity in the constitutive and induced defence of conifers against insects and pathogens. New Phytol 170:657-675.
- Kumar S, Tamura K, Jacobsen I and Nei N (2000) MEGA: Molecular Evolutionary Genetics Analysis, version 2.0.Pennsylvania and Arizona State University, University Park, Pennsylvania and Tempe, Arizona.
- Phillips DR, Rasbery JR, Bartel B and Matsuda SPT (2006) Biosynthetic diversity in plant triterpene cyclization. Curr Opin Plant Biol 9:305-314.
- Rohdich F, Bacher A and Eisenreich W (2005) Isoprenoid biosynthetic pathways as anti-infective drug targets. Biochem Soc Trans 33:785-791.
- Ruberto G and Rapisarda P (2002) Essential oils of new pigmented citrus hybrids: Citrus sinensis L. Osbeck x C. clementina Hort. ex Tanaka. J Food Sci 67:2778-2780.
- Saitou N and Nei M (1987) The neighbour joining method: A new method for reconstructing phylogenetic trees. Molec Biol Evol 4:406-425.
- Sawamura M, Tu NTM, Yu XL and Xu BQ (2005) Volatile constituents of the peel oils of several sweet oranges in China. J Essen Oil Res 17:2-6.
- Sharkey TD, Yeh S, Wiberley AE, Falbel TG, Gong D and Fernandez DE (2005) Evolution of the isoprene biosynthetic pathway in kudzu. Plant Physiol 137:700-712.
- Sharon-Asa L, Shalit M, Frydman A, Bar E, Holland D, Or E, Lavi U, Lewinsohn E and Eyal Y (2003) Citrus fruit flavor and aroma biosynthesis: Isolation, functional characterization, and developmental regulation of Cstps1, a key gene in the production of the sesquiterpene aroma compound valencene. Plant J 36:664-674.
- Shimada T, Endo T, Fujii H, Hara M, Ueda T, Kita M and Omura M (2004) Molecular cloning and functional characterization of four monoterpene synthase genes from Citrus unshiu Marc. Plant Sci 166:49-58.
- Shimada T, Endo T, Fujii H and Omura M (2005) Isolation and characterization of a new d-limonene synthase gene with a different expression pattern in Citrus unshiu Marc. Sci Hortic 105:507-512.
- Steele CL, Katoh S, Bohlmann J and Croteau R (1998) Regulation of oleoresinosis in grand fir (Abies grandis) - Differential transcriptional control of monoterpene, sesquiterpene, and diterpene synthase genes in response to wounding. Plant Physiol 116:1497-1504.
- Sun T and Kamiya Y (1994) The Arabidopsis GA1 locus encodes the cyclase ent-kaurene synthetase A of gibberellin biosynthesis. Plant Cell 6:1509-1518.
- Tatusov RL, Koonin EV and Lipman DJ (1997) A genomic perspective on protein families. Science 278:631-637.
- Tholl D (2006) Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Curr Opin Plant Biol 9:297-304.
- Thompson JD, Higgins DG and Gibson, TJ (1994) CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, positions-specific gap penalties and weight matrix choice. Nucleic Acids Res 22:4673-4680.
- Trapp S and Croteau R (2001) Defensive resin biosynthesis in conifers. Annu Rev Plant Physiol Plant Molec Biol 52:689-724.
- Van Poecke RMP, Posthumus MA and Dicke M (2001) Herbivore-induced volatile production by Arabidopsis thaliana leads to attraction of the parasitoid Cotesia rubecula: Chemical, behavioural and gene-expression analysis. J Chem Ecol 27:1911-1928.
- Velikova V and Loreto F (2005) On the relationship between isoprene emission and thermotolerance in Phragmites australis leaves exposed to high temperatures and during the recovery from heat stress. Plant Cell Environ 28:318-327.
- Verzera A, Trozzi A, Zappala M, Condurso C and Cotroneo A (2005) Essential oil composition of Citrus meyerii Y. Tan. and Citrus medica L. cv. Diamante and their lemon hybrids. J Agric Food Chem 53:4890-4894.
- Yamaguchi S, Sun T-P, Kawaide H and Kamiya Y (1998) The GA2 locus of Arabidopsis thaliana encodes ent-kaurene synthase of gibberellin biosynthesis. Plant Physiol 116:1271-1278.
- CitEST Database, http://citest.centrodecitricultura.br/ (March 25, 2006).
- National Center for Biotechnology Information (NCBI), http:// www.ncbi.nlm.nih.gov/ (March 25, 2006).
- BLAST, http://www.ncbi.nlm.nih.gov/BLAST/ (March 25, 2006).
- InterProScan, http://www.ebi.ac.uk/InterProScan/ (March 28, 2006).