Abstract
Electrophorus spp. generate high-voltage electric discharges for defense and hunting, and low-voltage electric discharges (as other Gymnotiformes) for electrolocation and communication. Despite intense interest in the unusual electrogenic and electroreceptive capacities of electric eels, the other sensory systems of Electrophorus spp. are relatively poorly known. Here we describe the ontogenetic development and organization of the cephalic lateral-line canals in the lowland electric eel, Electrophorus varii. Preserved specimens of larvae, juveniles, and adults were examined to describe the spatial distribution of the canals and pores. Ontogenetic shifts of the cephalic lateral line formation were observed for each canal and support a hypothesis of non-synchronized development. The morphogenesis of cephalic canals in larvae and juveniles begins just before the onset of exogenous feeding. In adults, the cephalic sensory canals are formed separately from the skull and overlay cranial and mandibular bones and muscles. This study provides the first detailed description of the development and organization of the cephalic lateral-line system in Electrophorus varii.
Keywords:
Electric eel; Life history; Mechanoreception; Ontogenetic shifts; Sensory system
Resumo
Electrophorus spp. geram descargas elétricas de alta voltagem, usadas para defesa e caça, e (como os demais Gymnotiformes) descargas elétricas de baixa voltagem para eletrolocalização e comunicação. Apesar do grande interesse nas capacidades eletrogênicas e eletrorreceptivas incomuns dos poraquês, os outros sistemas sensoriais de Electrophorus spp. são relativamente pouco conhecidos. Aqui nós descrevemos o desenvolvimento ontogenético e a organização dos canais da linha lateral cefálica no poraquê da planície Electrophorus varii. Exemplares preservados de larvas, juvenis e adultos foram analisados para descrever a distribuição espacial dos canais e poros. Variações ontogenéticas na formação da linha lateral cefálica foram observadas para cada canal, apresentando um desenvolvimento não-sincronizado. Variações ontogenéticas da formação da linha lateral cefálica foram observadas para cada canal e suportam uma hipótese de desenvolvimento assincrônico. A morfogênese dos canais cefálicos em larvas e juvenis precede o início da alimentação exógena. Nos adultos, os canais sensoriais cefálicos se formam separadamente do crânio e sobrepõem os ossos e músculos cranianos e mandibulares. Este estudo fornece a primeira descrição detalhada do desenvolvimento e organização do sistema da linha lateral cefálica de Electrophorus varii.
Palavras-chave:
História de vida; Mecanorrecepção; Mudanças ontogenéticas; Poraquê; Sistema sensorial
INTRODUCTION
The genus Electrophorus comprises freshwater electric eels (known as poraquês in the Brazilian Amazon) that are able to generate high-voltage discharges of up to 860V (Crampton, 2019Crampton WGR. Electroreception, electrogenesis and electrical signal evolution. J Fish Biol. 2019; 95(1):92–134. https://doi.org/10.1111/jfb.13922
https://doi.org/10.1111/jfb.13922...
; de Santana et al., 2019de Santana CD, Crampton WGR, Dillman CB, Frederico RG, Sabaj MH, Covain R et al. Unexpected species diversity in electric eels with a description of the strongest living bioelectricity generator. Nat Commun. 2019; 10:4000. https://doi.org/10.1038/s41467-019-11690-z
https://doi.org/10.1038/s41467-019-11690...
) for defense and hunting (Catania, 2019Catania KC. The astonishing behavior of electric eels. Front Integr Neurosci. 2019; 13(1):23. https://doi.org/10.3389/fnint.2019.00023
https://doi.org/10.3389/fnint.2019.00023...
). Like other Gymnotiformes, they also generate low-voltage electric discharges for electrolocation, prey detection, and communication with conspecifics (Moller, 1995Moller P. Electric fishes: history and behavior. Springer; 1995.; Crampton, Albert, 2006Crampton WGR, Albert JS. Evolution of electric signal diversity in gymnotiform fishes. In: Ladich F, Collin SP, Moller P, Kapoor BG, editors. Communication in fishes. Gwalior: Science Publishers, 2006. p.647–731.). During early life stages, however, electric eels are unable to generate high discharges to stun prey. This, in part, may drive ontogenetic changes in diet (Assunção, Schwassmann, 1995Assunção MIS, Schwassmann HO. Reproduction and larval development of Electrophorus electricus on Marajó Island (Pará, Brazil). Ichthyol Explor Fres. 1995; 6(2):175–84. ; Schwassmann et al., 2014Schwassmann HO, Assunção MIS, Kirschbaum F. Ontogeny of the electric organs in the electric eel, Electrophorus electricus: Physiological, histological and fine structural investigations. Brain Behav Evol. 2014; 84(4):288–302. https://doi.org/10.1159/000367884
https://doi.org/10.1159/000367884...
). Electrophorus larvae are known to feed on eggs provided by an adult while still in the nest, and, as they grow up, the small eels start to eat small aquatic invertebrates (Assunção, Schwassmann, 1995Assunção MIS, Schwassmann HO. Reproduction and larval development of Electrophorus electricus on Marajó Island (Pará, Brazil). Ichthyol Explor Fres. 1995; 6(2):175–84. ).
Recently, de Santana et al., (2019)de Santana CD, Crampton WGR, Dillman CB, Frederico RG, Sabaj MH, Covain R et al. Unexpected species diversity in electric eels with a description of the strongest living bioelectricity generator. Nat Commun. 2019; 10:4000. https://doi.org/10.1038/s41467-019-11690-z
https://doi.org/10.1038/s41467-019-11690...
described two new species of Electrophorus, E. varii de Santana, Wosiacki, Crampton, Sabaj, Dillman, Mendes-Júnior & Castro e Castro, 2019, and E. voltai de Santana, Wosiacki, Crampton, Sabaj, Dillman, Castro e Castro, Bastos & Vari, 2019, based on molecular, anatomical and ecological evidence. This unexpected diversity suggests that, although electric eels have been known for almost 250 years, their basic biology may exhibit interspecific variation that is still poorly comprehended. Also, while many studies have focused on the electrogenic-electrosensory attributes of electric eels, the anatomy, physiology, and ecological relevance of their non-electrical sensory systems and their functional relevance have been overlooked (e.g., Hagiwara et al., 1965Hagiwara S, Szabo T, Enger PS. Physiological properties of electroreceptors in the electric eel, Electrophorus electricus. J Neurophysiol. 1965; 28(5):775–83. https://doi.org/10.1152/jn.1965.28.5.775
https://doi.org/10.1152/jn.1965.28.5.775...
; Szabo, 1974Szabo T. Anatomy of the specialized lateral line organs of electroreception. In: Fessard A, editor. Electroreceptors and other specialized receptors in lower vertebrates. Handbook of Sensory Physiology, vol. 3. Berlin: Springer, Heidelberg; 1974. p.13–58. ; Gotter et al., 1998Gotter AL, Kaetzel MA, Dedman JR. Electrophorus electricus as a model system for the study of membrane excitability. Comp Biochem Physiol A Mol Integr Physiol. 1998; 119(1):225–41. https://doi.org/10.1016/S1095-6433(97)00414-5
https://doi.org/10.1016/S1095-6433(97)00...
).
The lateral line system is responsible for detecting water movements generated by potential predators, prey and conspecifics (e.g., Coombs, Braun, 2003Coombs S, Braun CB. Information processing by the lateral line system. In: Collin SP, Marshall NJ, editors. Sensory processing in aquatic environments. New York: Springer; 2003. p.122–38.). In electric eels, it is presumed to play an important role in early life stages, before the electrogenic system is fully functional (Schwassmann et al., 2014Schwassmann HO, Assunção MIS, Kirschbaum F. Ontogeny of the electric organs in the electric eel, Electrophorus electricus: Physiological, histological and fine structural investigations. Brain Behav Evol. 2014; 84(4):288–302. https://doi.org/10.1159/000367884
https://doi.org/10.1159/000367884...
). The distribution of the lateral line canals is also important as a morphological character used for taxonomic studies and phylogenetic relationships inferences (e.g., Arratia, Huaquín, 1995Arratia G, Huaquín L. Morphology of the lateral line system and of the skin of diplomystid and certain primitive loricarioid catfishes and systematic and ecological considerations. Bonn Zool Monogr. 1995; 36:1–110.; Di Dario, de Pinna, 2006Di Dario F, de Pinna MC. The supratemporal system and the pattern of ramification of cephalic sensory canals in Denticeps clupeoides (Denticipitoidei, Teleostei): additional evidence for monophyly of Clupeiformes and Clupeoidei. Pap Avulsos Zool. 2006; 46(10):107–23. https://doi.org/10.1590/S0031-10492006001000001
https://doi.org/10.1590/S0031-1049200600...
; de Santana, Crampton, 2010de Santana CD, Crampton WGR. A review of the South American electric fish genus Porotergus (Gymnotiformes: Apteronotidae) with the description of a new species. Copeia. 2010; 2010(1):165–75. https://doi.org/10.1643/CI-05-136
https://doi.org/10.1643/CI-05-136...
; Stephens, 2010Stephens RR. A description of the cephalic lateralis system of Anchoa mitchilli (Valenciennes) (Clupeomorpha: Engraulidae) with identification of synapomorphies for the Engraulidae. P Biol Soc Wash. 2010; 123(1):8–16. https://doi.org/10.2988/09-13.1
https://doi.org/10.2988/09-13.1...
; Crampton et al., 2016Crampton WGR, de Santana CD, Wadell JC, Lovejoy NR. A taxonomic revision of theNeotropical electric fish genus Brachyhypopomus (Ostariophysi: Gymnotiformes: Hypopomidae), with descriptions of 15 new species. Neotrop Ichthyol. 2016, 14(4):e150146. http://dx.doi.org/10.1590/1982-0224-20150146
http://dx.doi.org/10.1590/1982-0224-2015...
).
Despite its evolutionary and adaptive importance, little is known about the lateral line structure in Electrophorus species, with the main studies on the topic dating back to the 1970s and focusing on the electroreceptive organs associated with the mechanosensory system (e.g., Szabo, 1965Szabo T. Sense organs of the lateral line system in some electric fish of the Gymnotidae, Mormyridae and Gymnarchidae. J Morphol. 1965; 117(2):229–49. https://doi.org/10.1002/jmor.1051170208
https://doi.org/10.1002/jmor.1051170208...
, 1974Szabo T. Anatomy of the specialized lateral line organs of electroreception. In: Fessard A, editor. Electroreceptors and other specialized receptors in lower vertebrates. Handbook of Sensory Physiology, vol. 3. Berlin: Springer, Heidelberg; 1974. p.13–58. ). The only detailed anatomical description of the lateral-line canals of Electrophorus is provided by Pastana (2014)Pastana MNL. Canais e poros do sistema látero-sensorial cefálico de Characiformes (Ostariophysi): anatomia e seu significado filogenético. [Master Dissertation] Ribeirão Preto: Universidade de São Paulo; 2014. Available from: https://www.teses.usp.br/teses/disponiveis/59/59139/tde-06012015-123252/pt-br.php
https://www.teses.usp.br/teses/disponive...
who included cephalic lateral-line features as characters in a study of characiform phylogeny (the study included Electrophorus electricus and some other gymnotiform species as outgroups).
The spatial distribution of lateral-line canals and pores on the cephalic surface has been broadly related to the habitats use and feeding habits in fishes (e.g., Arratia, Huaquín, 1995Arratia G, Huaquín L. Morphology of the lateral line system and of the skin of diplomystid and certain primitive loricarioid catfishes and systematic and ecological considerations. Bonn Zool Monogr. 1995; 36:1–110.). Nonetheless, despite numerous investigations of the importance and function of their electric organs (Szabo, 1966Szabo T. The origin of electric organs of Electrophorus electricus. Anat Rec. 1966; 155(1):103–10. https://doi.org/10.1002/ar.1091550112
https://doi.org/10.1002/ar.1091550112...
; Schwassmann et al., 2014Schwassmann HO, Assunção MIS, Kirschbaum F. Ontogeny of the electric organs in the electric eel, Electrophorus electricus: Physiological, histological and fine structural investigations. Brain Behav Evol. 2014; 84(4):288–302. https://doi.org/10.1159/000367884
https://doi.org/10.1159/000367884...
) the mechanoreceptive and other non-electrical sensory systems of Electrophorus species are poorly known. Likewise, most investigations of predatory behavior in Electrophorus species have focused on the strength of the high-voltage electric discharges for striking prey, mostly fishes and invertebrates (Catania, 2014Catania K. The shocking predatory strike of the electric eel. Science. 2014; 346(6214):1231–34. https://doi.org/10.1126/science.1260807
https://doi.org/10.1126/science.1260807...
; Oliveira et al., 2020Oliveira MSB, Mendes-Júnior RNG, Tavares-Dias M. Diet composition of the electric eel Electrophorus voltai (Pisces: Gymnotidae) in the Brazilian Amazon region. J Fish Biol. 2020. https://doi.org/10.1111/jfb.14413
https://doi.org/10.1111/jfb.14413...
; Bastos et al., 2021Bastos DA, Zuanon JAS, Py-Daniel LR, de Santana CD. Social predation in electric eels. Ecol Evol. 2021; 11(3):1088–92. https://doi.org/10.1002/ece3.7121
https://doi.org/10.1002/ece3.7121...
). However, the feeding behavior of early ontogenetic phases have been neglected and there is a lack of information on how the electric eels search and locate their prey. Due to the highly reduced visual systems of most gymnotiforms (Albert, Crampton, 2006Albert JS, Crampton WGR. Diversity and phylogeny of neotropical electric fishes (Gymnotiformes). In: Bullock TH, Hopkins CD, Popper AN, Fay RR, editors. Electroreception. New York: Springer; 2006. p.360–409.), the mechanosensory lateral line system may supplement the electrosensory system in facilitating prey localization (Westby, 1988Westby GWM. The ecology, discharge diversity and predatory behaviour of gymnotiform electric fish in the coastal streams of French Guiana. Behav Ecol Sociobiol. 1988; 22:341–54. https://doi.org/10.1007/BF00295103
https://doi.org/10.1007/BF00295103...
). Here we describe the early development and organization of cephalic lateral-line canals in Electrophorus varii and explore correlations between lateral line ontogeny and larval and juvenile feeding behavior.

Sampling areas (Adolpho Ducke Reserve and Purupuru Lake, Amazonas, Brazil) where individuals of Electrophorus varii were captured. The white stars represent the exact sampling spots.
MATERIAL AND METHODS
Sampling sites. Specimens of E. varii were collected in two different sites of the Brazilian Amazon Basin: a terra-firme stream at Adolpho Ducke Forest Reserve (Rio Negro drainage, 2°59’02.4”S 59°55’55.2”W), located at the northeastern outskirts of the city of Manaus, and a whitewater lake, Purupuru Lake (Rio Solimões drainage, 3°24’03.6”S 59°51’03.6”W), in the Careiro da Várzea municipality (Fig. 1). Procedures for sampling followed INPA’s ethics committee rules (protocol number 044/2016).
Adolpho Ducke Forest Reserve. Nursery sites were monthly monitored from July 2017 to September 2018 at Igarapé Bolivia, a terra-firme stream in the reserve, using electric fish finders (Crampton et al., 2007Crampton WGR, Jonathan KW, Smyth C, Walz SA. Design and construction of an Electric Fish Finder. Neotrop Ichthyol. 2007; 5(3):425–28. https://doi.org/10.1590/S1679-62252007000300022
https://doi.org/10.1590/S1679-6225200700...
). Five nests were located under stream banks, where the development of the nestlings was monitored. The nests were visited every two-three days for behavioral record-taking and to sample up to five specimens per nest and event; these were used to build an ontogenetic series. To collect specimens in the first weeks after hatching, we dug small openings on the stream bank above the nest cavities. After the second month of life, the eels were collected at night as they foraged outside the nest. All specimens were preserved in 4% buffered formalin and conserved in 75% ethanol solution.
Purupuru Lake. Sampling in Purupuru Lake focused on adult individuals and was conducted in November 2018, using hook-and-line. The specimens were preserved in 10% formalin solution and conserved in 75% ethanol solution.
Ontogenetic series and staging. The number and distribution of lateral-line pores and canals were examined in 22 larvae and 10 juvenile specimens, ranging from 1.5 to 38.0 cm TL, and two adults of 148.0 and 111.5 cm TL. The distinction between larvae and juveniles was made following a staging protocol based on anatomic and behavioral characteristics (Verçoza, 2020Verçoza G. Desenvolvimento ontogenético do poraquê Electrophorus varii (Gymnotiformes: Gymnotidae). [Master dissertation] Manaus: Universidade Federal do Amazonas; 2020. Available from: https://tede.ufam.edu.br/bitstream/tede/7916/9/Disserta%c3%a7%c3%a3o_GabrielVer%c3%a7oza_PPGZOO.pdf
https://tede.ufam.edu.br/bitstream/tede/...
), where larvae are recognized as the individuals measuring between 1.0 to 15.0 cm TL under parental care, juveniles as the specimens without parental care ranging from 15.0 to 85.0 cm, and adults as the mature specimens above 85.0 cm TL (Mendes-Júnior et al., 2016Mendes-Junior RNG, Oliveira JCS, Ferrari SS. Biology of the electric eel, Electrophorus electricus, Linnaeus, 1766 (Gymnotiformes: Gymnotidae) on the floodplain of the Curiaú River, eastern Amazonia. Rev Fish Biol Fisheries. 2016; 26:83–91. https://doi.org/10.1007/s11160-015-9407-9
https://doi.org/10.1007/s11160-015-9407-...
) (shown in the Fig. S1).
Larval and juvenile feeding behavior. The larval and juvenile observations were carried out only at the Ducke Reserve due to environmental conditions such as transparent water and nest location. Field observations were made from both outside water (from the banks) and underwater (using a GoPro Hero 3+ and a Panasonic HX-A1 camera, as well as a portable borescope) at the same days of specimens collection for anatomical analyses. To obtain images from the interior of the nest cavity, one of the image-capturing devices was inserted next to the foam structure that composes the upper layer of the nest. The feeding behavior was classified in two phases following Bastos (2020)Bastos DA. História Natural de poraquês (Electrophorus spp.), Gymnotiformes: Gymnotidae. [PhD Thesis] Manaus: Instituto Nacional de Pesquisas da Amazônia; 2020.: I – larval feeding inside the nest by eggs provided by the adult eel(s), and II – juvenile foraging outside the nest, consuming small invertebrates.
Anatomical analysis. Larvae and juvenile specimens were examined under a stereomicroscope (16x). Adult specimens were examined both by naked eye and using a stereomicroscope, after the removal of the epidermis of the head and the anterior surface of the trunk to expose the cephalic (or cranial) lateral-line canal’s structure (S2). Three-dimensional visualization of the cranium as well as the analysis of the reconstructed data was performed using VGStudio MAX 2.2.3 64 bit (Volume Graphics GmbH, Heidelberg, Germany).
Pores were quantified by counting on both left and right sides of the head and for each canal. Organization and connections of the canals were observed by injecting a dye (0.5% toluidine blue solution) into the pores. The terminology of cephalic lateral line canals followed Northcutt (1989)Northcutt RG. The phylogenetic distribution and innervation of craniate mechanoreceptive lateral lines. In: Coombs S, Görner P, Müns H, editors. The mechanosensory lateral line: neurobiology and evolution. New York: Springer; 1989. p.17–78.: infraorbital (IO), mandibular (MD), otic (O), pre-opercular (PO), supraorbital (SO) and supratemporal (ST). Some other terms used in the current study followed Webb (2013). “Groove” refers to the canal structure before its enclosure, when the canal neuromasts are totally exposed. “Segment of canals” refers to a short section of canals that after the enclosure process becomes fused to the adjacent segment, resulting in a lateral-line pore. “Pore” refers to the canal opening on the skin surface that serves to maintain canal neuromasts in contact with the external environment.
Voucher specimens of E. varii were catalogued at Fish Collection of the Instituto Nacional de Pesquisas da Amazônia (INPA), Manaus, Brazil (larvae and juveniles: INPA-ICT 059502 – 059510; adults: INPA-ICT 059511 – 059513).

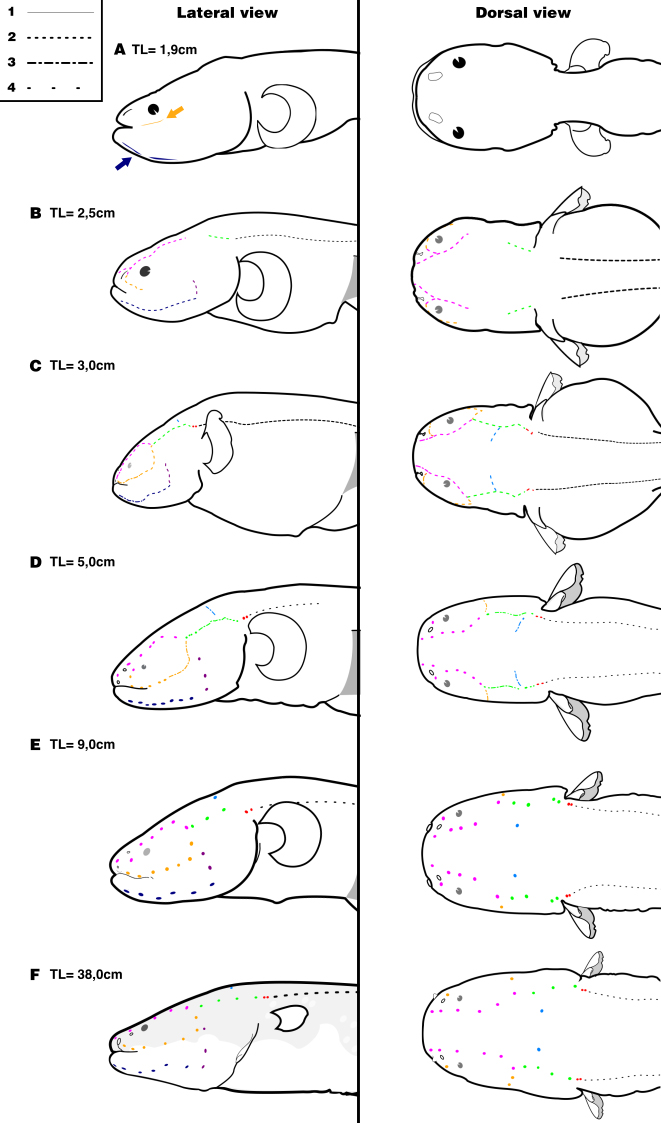
Ontogenetic development of the lateral-line canals in larvae and juveniles of Electrophorus varii, in lateral and dorsal views. A. 1.9 cm TL; B. 2.5 cm TL; C. 3.0 cm TL; D. 5.0 cm TL; E. 9.0 cm TL; F. 38 cm TL. Types of dashes indicate the (1) shallow and short grooves on the skin; (2) deep grooves; (3) segments of grooves start enclosure; (4) pores of canals on the skin. Yellow: infraorbital canal; navy blue: mandibular canal; green: otic canal; purple: pre-opercular canal; pink: supraorbital canal; light blue: supratemporal canal; grey: posterior lateral-line canal of the trunk and; red: otic pit between cephalic and trunk canals. Arrows indicate short grooves of the presumptive infraorbital and mandibular canals.
RESULTS
Lateral-line canals in larval and juvenile specimens. Although the hatchlings range from 0.8 to 1.1 cm TL, the occurrence of externally visible lateral-line canals was firstly detected in individuals of 1.9 cm TL. At this size, short grooves start to appear on the head surface, marking the beginning of canal morphogenesis and originating the presumptive infraorbital and mandibular canals (Fig. 2A). This event occurs after consumption of the yolk and just before the individuals initiate the exogenous feeding on provided eggs.
At 2.5 cm TL, the grooves become deeper into the skin, clearly marking the supraorbital, infraorbital, mandibular, otic and pre-opercular canals (Fig. 2B). At this size, the individuals start exhibiting more directional, oriented swimming and are able to actively avoid being caught by a hand net.
After reaching nearly 3.0 cm TL, the anteriormost grooves of the infraorbital, mandibular and supraorbital canals start their closure, forming a series of short canal segments. Lastly, the fusion between two canal segments is accompanied by the appearance of spots that represent where the lateral-line pores will be located (Fig. 2C). At this stage the otic and pre-opercular canals are still open and the supratemporal canal becomes visible. The otic canal is located posterior to the confluence of the infraorbital and supraorbital canals on the cephalic surface. A small pit is formed at the end of the otic canal, posteriorly in the skull, marking the limit to the trunk lateral-line canal. Laterally positioned on the head, the pre-opercular canal forms a continuum connected to the posterior section of the mandibular canal. Ventrally, the anteriormost pores of the mandibular canal are located at the tip of the jaws, just below the dentaries symphysis.
The formation of canal segments continues until fish reach approximately 5.0 cm TL. At this length, the posterior portion of the supraorbital and mandibular canals, as well the dorsal portion of the pre-opercular, are closed. The infraorbital, otic and supratemporal canals already have the grooves edges fused, but most parts of those canals remain open. The surrounding epithelium covers the otic pit, forming a double-pored structure dorsally to the base of the pectoral fins (Fig. 2D).
The complete set of pores and fused canal segments is present at nearly 9.0 cm TL, when individuals show a cephalic lateral-line morphology similar to that of the adults, with large, well-separated pores over the skin surface (Figs. 2E–F). Nevertheless, pores may be commonly found doubled in all canal series. Individuals of this size start to forage outside the nest, feeding on small invertebrates associated with the substrate.
The number of pores also varies on the mandibular canal, specifically for the symphyseal pores (located on the symphysis of the mandible) that connect both left and right branches of the canal. In small specimens, there are two pores on this region that seem to fuse, forming an arch-like pattern (Fig. 3C). The number of pores or short segments since the first appearance to the end of morphogenesis in each canal is shown in Tab. 1.
Number of pores of cephalic lateral-line canals during the ontogeny of Electrophorus varii. Mean number of segments or pores are presented for each canal in larval/juvenile and adult specimens.
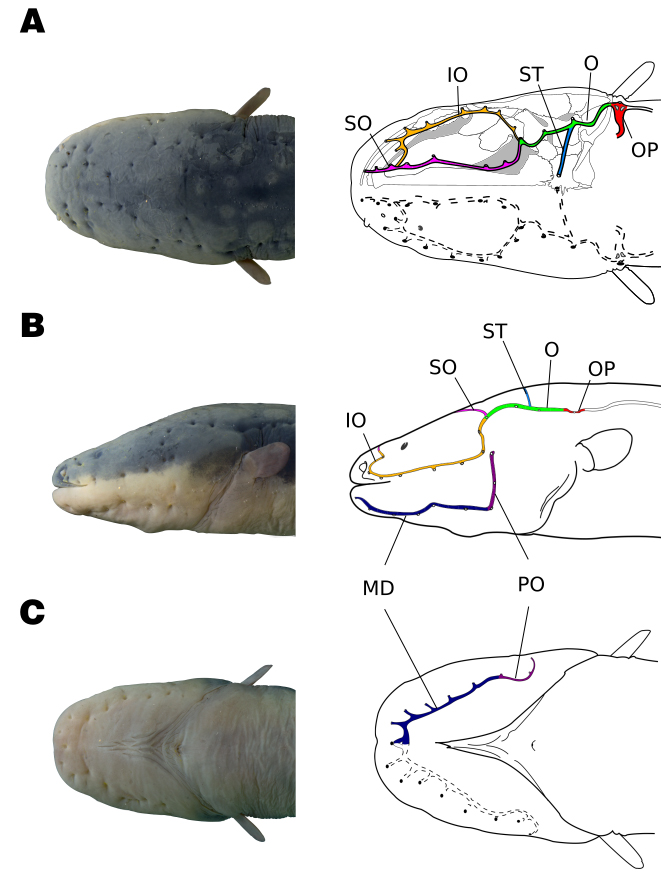
Cephalic lateral-line canals in adults. The cephalic lateral-line canals in adults of Electrophorus varii are positioned beneath the thick epidermis and pores are visible on the head surface. All canals present tubules terminating in large pores with stiffened edges (Fig. 3). The canal walls are relatively rigid and become partially ossified in adult stages (Fig. 4).
The supraorbital canal stretches along the frontals and extends towards the tip of the snout. At midway between the posterior nare and the orbit, the supraorbital receives a branch of the infraorbital canal. The infraorbital is a long canal, running along a large stretch from the back of the skull, surrounding the eye, until deviating dorsally towards the supraorbital canal. Thus, infraorbital and supraorbital canals are connected anteriorly and posteriorly, making a closed circuit. Both canals branch off small tubules that open in the skin pores. The posterior confluence of the supraorbital and infraorbital is connected to the otic canal, forming a triple joint.

Organization of the cephalic lateral-line canals in adult Electrophorus varii. Arrangement of all canals in A. dorsal, B. lateral and C. ventral views. Yellow: infraorbital canal; navy blue: mandibular canal; green: otic canal; purple: pre-opercular canal; pink: supraorbital canal; light blue: supratemporal canal; grey: posterior lateral-line canal of the trunk; red: otic pit (OP) between cephalic and trunk canals. The photographs are from a juvenile that completed the development and is morphologically similar to an adult. The drawings are from an individual of 148 cm TL.

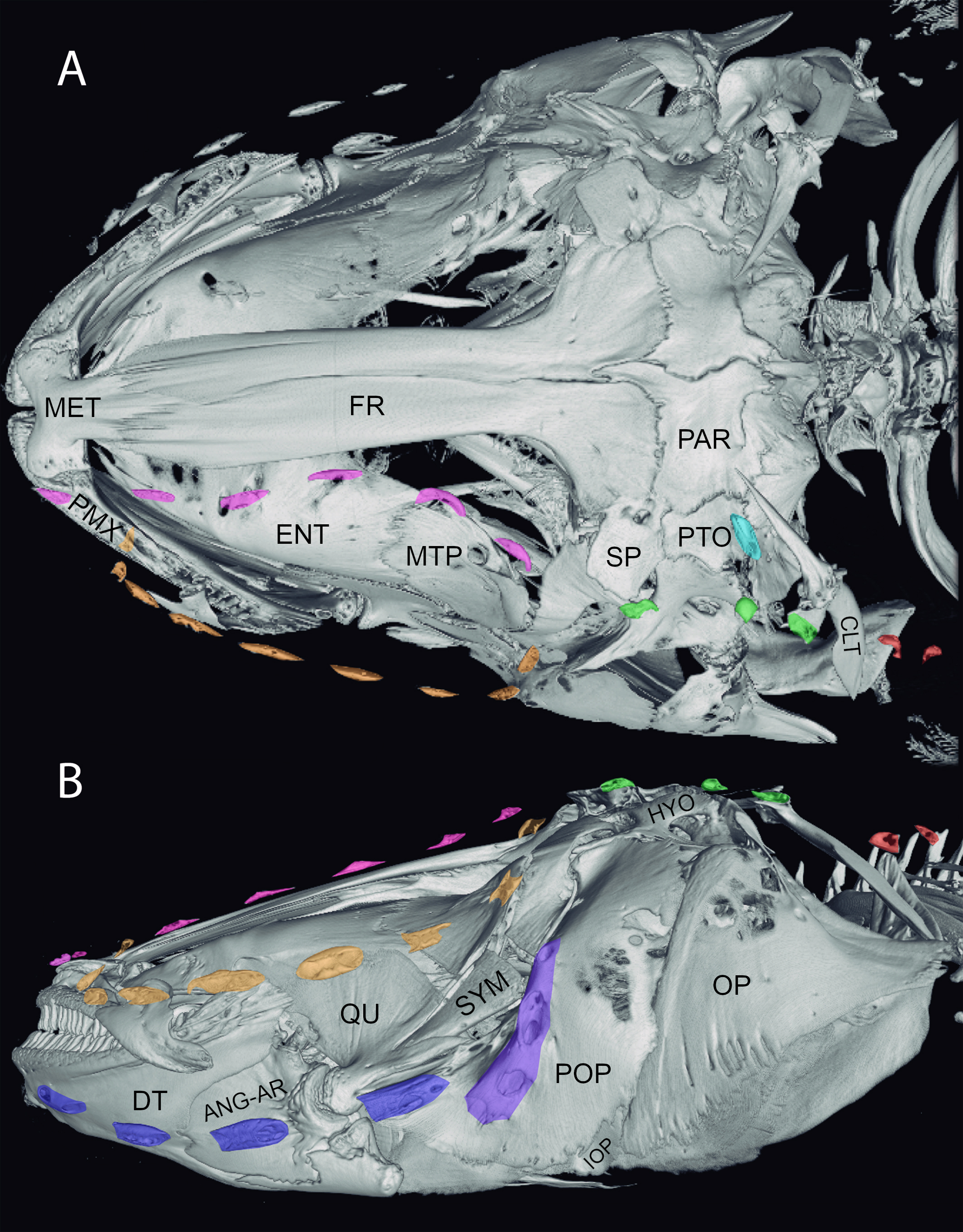
Images of CT scan of the head of Electrophorus varii. A. dorsal and B. left lateral views of the cranium show the ossification of the lateral-line canals. The ossified segments of canals are marked by colors. Yellow: infraorbital canal; navy blue: mandibular canal; green: otic canal; purple: pre-opercular canal; pink: supraorbital canal; light blue: supratemporal canal; red: otic pit ossicles. ANG-AR: Angulo-articular; CLT: cleithrum; DT: dentary; ENT: entopterygoid; FR: frontal bone; HYO: Hyomandibular; IOP: interopercle; MET: mesethmoid; MTP: metapterygoid; OP: opercle; PAR: parietal; PMX: premaxila; POP: preopercle; PTO: pterotic; QU: quadrate; SP: sphenotic; SYM: sympletic. Specimen size = 82 cm TL.
The otic canal is relatively long, extending from its joint to the supraorbital and infraorbital confluence until the otic pit, bearing three to four pores. The supratemporal canal is represented by a single long tubule, derived from the otic canal mid area, oriented mesially on the parietal portion, and terminating in a single pore. The posterior edge of the otic canal ends on the otic pit, which is connected to the lateral-line canal on the trunk and extends towards the end of the body. The otic pit is a depression on the skin that is also observed in the larval phase (Fig. 2C, the early stage of its appearance is represented by two dots in red), but much more conspicuous in adults. The otic pit, underneath the skin, shows three exits: more anteriorly, it is the exit of the end of the otic canal, more posteriorly, it is the surfacing of the body lateral line, and another in the middle. In fact, the otic pit represents a deep penetration of the otic canal into the body before surfacing as the body lateral line: opposite to the deep external pore, the thin and flexible canal wall is connected to an internal tubule that penetrates behind the head between the epaxial and hypaxial (obliquus superioris) muscles and expands as a chamber. This expanded chamber-like tubule presents a conspicuous sinuosity and is apparently connected to the anterior portion of the swim bladder by a short chamber with a flexural wall (Fig. 3A).
The mandibular canal is positioned latero-ventrally to the mouth and follows underneath and along the jaw and branches off in six to seven tubules that terminate in pores. Both sides of the mandibular canal are connected by a junction of two tubules that end in a pore in the symphysis of the jaw. Posteriorly, the pre-opercular canal forms a continuum with the mandibular canal, makes a turn on the opercular elements and turns dorsally, towards the cranium. In contrast to all other canals, the pre-opercular canal is partially embedded in the underlying dermal bone, with pores located at the point of junction between two segments of canals. Pores are present in all canals and their number may vary between individuals due to cases of ‘pore doubling’, i.e., where one tubule opens in two pores (Tab. 1).
DISCUSSION
The morphogenesis of cephalic sensory canals in larvae and juveniles of E. varii begins just before the onset of the exogenous feeding. This timing is also observed in other sensory structures, such as the electric organ (Schwassmann et al., 2014Schwassmann HO, Assunção MIS, Kirschbaum F. Ontogeny of the electric organs in the electric eel, Electrophorus electricus: Physiological, histological and fine structural investigations. Brain Behav Evol. 2014; 84(4):288–302. https://doi.org/10.1159/000367884
https://doi.org/10.1159/000367884...
), which in some fishes occurs synchronically to the yolk depletion and a rapid skull development. These synchronous changes are commonly associated with the start of exogenous food sources consumption (e.g., Mathias, LI, 1982Mathias JA, LI S. Feeding habits of walleye larvae and juveniles: Comparative laboratory and field studies. T Am Fish Soc. 1982; 111(6):722–35. https://doi.org/10.1577/1548-8659(1982)111<722:FHOWLA>2.0.CO;2
https://doi.org/10.1577/1548-8659(1982)1...
).
Unlike most fishes, where the morphogenesis of the cephalic lateral-line canals is closely related to dermatocranial bones (Webb, Shirey, 2003Webb JF, Shirey JE. Postembryonic development of the cranial lateral line canals and neuromasts in zebrafish. Dev Dyn. 2003; 228(3):370–85. https://doi.org/10.1002/dvdy.10385
https://doi.org/10.1002/dvdy.10385...
), E. varii presents ossified lateral-line canals separately formed from the skull and overlying cranial and mandibular bones and muscles. The pre-opercular canal, however, is an exception, as it is embedded in the underlying bone. Similar morphology of cephalic lateral-line canals has also been observed in some apteronotid species (Albert, 2001Albert JS. Species diversity and phylogenetic systematics of American knifefishes (Gymnotiformes: Teleostei). Misc Publ Mus Zool Univ Mich. 2001; 190:1–127.) and Ophidiiformes (Pastana et al., 2020Pastana MNL, Bockmann FA, Datovo A. The cephalic lateral-line system of Characiformes (Teleostei: Ostariophysi): anatomy and phylogenetic implications. Zool J Linn Soc. 2020; 189(1):1–46. https://doi.org/10.1093/zoolinnean/zlz105
https://doi.org/10.1093/zoolinnean/zlz10...
). The tubules of E. varii are arranged in the sub-epidermal layer and terminated in pores on the skin surface. Nevertheless, ontogenetic shifts of the formation of lateral-line canals are similar to those observed for other teleost fish (Webb, Shirey, 2003Webb JF, Shirey JE. Postembryonic development of the cranial lateral line canals and neuromasts in zebrafish. Dev Dyn. 2003; 228(3):370–85. https://doi.org/10.1002/dvdy.10385
https://doi.org/10.1002/dvdy.10385...
; Mukai et al., 2008Mukai Y, Tuzan AD, Lim LS, Wahid N, Sitti Raehanah MS, Senoo S. Development of sensory organs in larvae of African catfish Clarias gariepinus. J Fish Biol. 2008; 73(7):1648–61. https://doi.org/10.1111/j.1095-8649.2008.02038.x
https://doi.org/10.1111/j.1095-8649.2008...
; Pastana et al., 2020Pastana MNL, Bockmann FA, Datovo A. The cephalic lateral-line system of Characiformes (Teleostei: Ostariophysi): anatomy and phylogenetic implications. Zool J Linn Soc. 2020; 189(1):1–46. https://doi.org/10.1093/zoolinnean/zlz105
https://doi.org/10.1093/zoolinnean/zlz10...
). Early larvae of E. varii present canal segments turning into deeper depressions and arranged mostly as conspicuous longitudinal grooves. As already described in some teleosts (e.g., Tarby, Webb, 2003Tarby ML, Webb JF. Development of the supraorbital and mandibular lateral line canals in the cichlid, Archocentrus nigrofasciatus. J Morphol. 2003; 255(1):44–57. https://doi.org/10.1002/jmor.10045
https://doi.org/10.1002/jmor.10045...
), asynchronous development of the cephalic lateral line canals is also observed in E. varii. While infraorbital, mandibular and supraorbital canals are completed from rostral to caudal direction, the opposite orientation (caudal to rostral) was observed for the otic canal. No evidence of development orientation could be detected for the pre-opercular and supratemporal canals. However, this should be verified with histological analysis.
The overall development and ossification patterns of the cephalic lateral-line canals of larval phase of E. varii are similar to those reported in the literature for most bony fishes (Tarby, Webb, 2003Tarby ML, Webb JF. Development of the supraorbital and mandibular lateral line canals in the cichlid, Archocentrus nigrofasciatus. J Morphol. 2003; 255(1):44–57. https://doi.org/10.1002/jmor.10045
https://doi.org/10.1002/jmor.10045...
; Webb, Shirey, 2003Webb JF, Shirey JE. Postembryonic development of the cranial lateral line canals and neuromasts in zebrafish. Dev Dyn. 2003; 228(3):370–85. https://doi.org/10.1002/dvdy.10385
https://doi.org/10.1002/dvdy.10385...
; Pastana et al., 2020Pastana MNL, Bockmann FA, Datovo A. The cephalic lateral-line system of Characiformes (Teleostei: Ostariophysi): anatomy and phylogenetic implications. Zool J Linn Soc. 2020; 189(1):1–46. https://doi.org/10.1093/zoolinnean/zlz105
https://doi.org/10.1093/zoolinnean/zlz10...
; Rizzato et al., 2020Rizzato PP, Pospisilova A, Hilton EJ, Bockmann FA. Ontogeny and homology of cranial bones associated with lateral-line canals of the Senegal Bichir, Polypterus senegalus (Actinopterygii: Cladistii: Polypteriformes), with a discussion on the formation of lateral-line canal bones in fishes. J Anat. 2020; 237(3):439–67. https://doi.org/10.1111/joa.13202
https://doi.org/10.1111/joa.13202...
). However, in E. varii all canals lie in the skin, completely independent of, and located over cranial and mandibular bones and muscles. This total lack of bony connection of the canals with the cranium is an unusual feature among teleosts (Webb, 2013Webb J. Morphological diversity, development and evolution of the mechanosensory lateral line system. In: Coombs S, Bleckmann H, Fay R, Popper A, editors. The Lateral Line System. Springer Handbook of Auditory Research, vol. 48. New York: Springer; 2013. p.17–72.). The integration of cranial lateral-line canals into the underlying bones has been thoroughly examined in fishes and classified as ‘two-component’ (when there is a fusion of two minor ossifications) or ‘one-component’ (where the underlying bones remain independent throughout the fish’s life) (Lekander, 1949Lekander B. The sensory line system and the canal bones in the head of some Ostariophysi. Acta Zool. 1949; 30(1-2):1–131. https://doi.org/10.1111/j.1463-6395.1949.tb00503.x
https://doi.org/10.1111/j.1463-6395.1949...
). However, a distinct and unusual pattern was observed in E. varii. The cephalic lateral-line canals are immersed into skin, without a direct association with the cranial bony elements during the canals formation. This situation precludes classifying E. varii as possessing either a two-component or one-component process of lateral line bones development (Lekander, 1949Lekander B. The sensory line system and the canal bones in the head of some Ostariophysi. Acta Zool. 1949; 30(1-2):1–131. https://doi.org/10.1111/j.1463-6395.1949.tb00503.x
https://doi.org/10.1111/j.1463-6395.1949...
; Tarby, Webb, 2003Tarby ML, Webb JF. Development of the supraorbital and mandibular lateral line canals in the cichlid, Archocentrus nigrofasciatus. J Morphol. 2003; 255(1):44–57. https://doi.org/10.1002/jmor.10045
https://doi.org/10.1002/jmor.10045...
; Webb, Shirey, 2003Webb JF, Shirey JE. Postembryonic development of the cranial lateral line canals and neuromasts in zebrafish. Dev Dyn. 2003; 228(3):370–85. https://doi.org/10.1002/dvdy.10385
https://doi.org/10.1002/dvdy.10385...
). Instead, canal formation in E. varii occurs independently of the underlying bones. As mentioned above, an exception was observed in the pre-opercular canal, which has the lateral line dependent on the underlying bone; this being a more typical case of developmental integration between lateral line canals and dermatocranial bones.
Electrophorus varii begins voracious predatory feeding in areas close to the nest at around 60 days after hatching. The larvae were seen foraging close to the substrate searching for prey (Bastos, pers. obs.), such as invertebrates and fishes (Assunção, Schwasmann, 1995Assunção MIS, Schwassmann HO. Reproduction and larval development of Electrophorus electricus on Marajó Island (Pará, Brazil). Ichthyol Explor Fres. 1995; 6(2):175–84. ). The robust and thick skin of the head of electric eels possibly help support injuries caused by objects in the water such as those caused by pieces of palm spines that were commonly found inserted in their epidermis. Along the ontogenetic development of E. varii, the lateral-line canals become progressively ossified in segments; this segmented ossification is maintained throughout the adults. Tomographic images show a restricted laminar dermal ossification on the infraorbital and mandibular canals, a condition similar to some Apteronotidae (Bernt et al., 2018Bernt MJ, Crampton WGR, Orfinger AB, Albert JS. Melanosternarchus amaru, a new genus and species of electric ghost knifefish (Gymnotiformes: Apteronotidae) from the Amazon Basin. Zootaxa. 2018; 4378(4):451–79. https://doi.org/10.11646/zootaxa.4378.4.1
https://doi.org/10.11646/zootaxa.4378.4....
). The pores present stiffened edges on the skin surface but structural composition analysis is required to explore whether this stiffening is the result of keratinization or mineralization.
Even though the lateral-line canal of the trunk has not been studied in detail in this paper, we noted that its wall is more flexible than in the cephalic branches (although this remains to be confirmed with histological analysis). In general, the evolutionary significance of the partial ossification of cephalic lateral-line canal’ walls and its disconnection of the cranial bones in E. varii is not clear among Gymnotiformes or other Ostariophysi, and deserves further study (see e.g., Arratia, Huaquín, 1995Arratia G, Huaquín L. Morphology of the lateral line system and of the skin of diplomystid and certain primitive loricarioid catfishes and systematic and ecological considerations. Bonn Zool Monogr. 1995; 36:1–110.; Britz, Moritz, 2007Britz R, Moritz T. Reinvestigation of the osteology of the miniature African freshwater fishes Cromeria and Grasseichthys (Teleostei, Gonorynchiformes, Kneriidae), with comments on kneriid relationships. Zoosyst Evol. 2007; 83(1):3–42. https://doi.org/10.1002/mmnz.200600016
https://doi.org/10.1002/mmnz.200600016...
).
Complete formation of the canal pores was detected only in late larvae (~9.0 cm TL) when foraging behavior had already begun. Foraging behavior in (adult) Gymnotiformes is usually associated only (or mostly) to the electrosensory system. However, considering that feeding is a multisensory task (Liang et al., 1998Liang XF, Liu JK, Huang BY. The role of sense organs in the feeding behaviour of Chinese perch. J Fish Biol. 1998; 52(5):1058–67. https://doi.org/10.1111/j.1095-8649.1998.tb00603.x
https://doi.org/10.1111/j.1095-8649.1998...
) and given the known role of the lateral-line canals in both prey detection (Coombs et al., 2001Coombs S, Braun CB, Donovan B. Orienting response of Lake Michigan mottled sculpin is mediated by canal neuromasts. J Exp Biol. 2001; 204(2):337–48. Available from: https://jeb.biologists.org/content/204/2/337
https://jeb.biologists.org/content/204/2...
) and schooling (Blaxter, Fuimann, 1989Blaxter JHS, Fuiman LA. Function of the free neuromasts of marine teleost larvae. In: Coombs S, Görner P, Münz H, editors. The mechanosensory lateral line: Neurobiology and evolution. New York: Springer-Verlag; 1989. p.481–99.), the ontogenetic timing of the finalization of the canal pores highlights the likelihood of a strong role for mechanoreception in the foraging behavior of E. varii in individuals of ca. 9+ cm size range. Because parental guarding still occurs at this size, we conjecture that the lateral-line system may also act as an additional source of parent-offspring communication through intraspecific mechanical signals, which occurs during mating and spawning behaviors in some other fish groups (e.g., Satou et al., 1994Satou M, Takeuchi HA, Nishii J, Tanabe M, Kitamura S, Okumoto N, Iwata M. Behavioral and electrophysiological evidences that the lateral line is involved in the inter-sexual vibrational communication of the himé salmon (landlocked red salmon, Oncorhynchus nerka). J Comp Physiol A. 1994; 174:539–49. https://doi.org/10.1007/BF00217373
https://doi.org/10.1007/BF00217373...
; Coombs, Braun, 2003Coombs S, Braun CB. Information processing by the lateral line system. In: Collin SP, Marshall NJ, editors. Sensory processing in aquatic environments. New York: Springer; 2003. p.122–38.).
Pores of the lateral-line canals system open to the skin surface by short tubules that extend from the main canal through the thick epidermis. All pores in E. varii adults are large and clearly visible on the skin surface, as is common in other fishes occupying habitats with limited visibility and low levels of acoustic noise (Branson, Moore, 1962Branson BA, Moore GA. The lateralis components of the acoustico-lateralis system in the sunfish family Centrarchidae. Copeia. 1962; 1962(1):1–108. https://doi.org/10.2307/1439483
https://doi.org/10.2307/1439483...
; McAllister, 1968McAllister DE. Mandibular pore pattern in the sculpin family Cottidae. Bull Nat Mus Can Contr Zool. 1968; 223:58–69. ; Hassan, 1989Hassan ES. Hydrodynamic imaging of the surroundings by the lateral line of the blind cave fish Anoptichtys jordani. In: Coombs S, Görner P, Münz H, editors. The Mechanosensory Lateral Line. New York: Springer Verlag; 1989. p.217–27.).
Despite being well developed, the cephalic lateral-line canals of E. varii are relatively simple in comparison to those of other Otophysi (e.g., Blaxter, 1987Blaxter JHS. Structure and development of the lateral line. Biol Rev. 1987; 62(4):471–514. https://doi.org/10.1111/j.1469-185X.1987.tb01638.x
https://doi.org/10.1111/j.1469-185X.1987...
). All the dorsal cephalic canals (SO, IO, ST, and O) are connected to each other and are linked to the ventral canals (MD and PO) by a connection between PO and IO. Connection of canals of both left and right sides of the head was observed only for the mandibular canal. Although the supratemporal commissure is present in other fishes (Arratia, Huaquín, 1995Arratia G, Huaquín L. Morphology of the lateral line system and of the skin of diplomystid and certain primitive loricarioid catfishes and systematic and ecological considerations. Bonn Zool Monogr. 1995; 36:1–110.; Webb, 2013Webb J. Morphological diversity, development and evolution of the mechanosensory lateral line system. In: Coombs S, Bleckmann H, Fay R, Popper A, editors. The Lateral Line System. Springer Handbook of Auditory Research, vol. 48. New York: Springer; 2013. p.17–72.), the lack of connection between the left and right sides of the head probably provides more accuracy for the localization of mechanical disturbances in the water. The anterior and posterior connections between supra- and infraorbital have been also observed in some species of Characiformes (e.g., Pastana, 2020), and Siluriformes (i.e., Rhamdella Eigenmann & Eigenmann, 1888, in Bockmann, Miquelarena, 2008Bockmann FA, Miquelarena AA. Anatomy and phylogenetic relationships of a new catfish species from northeastern Argentina with comments on the phylogenetic relationships of the genus Rhamdella Eigenmann and Eigenmann 1888 (Siluriformes, Heptapteridae). Zootaxa. 2008; 1780(1):1–54. and Pimelodella Eigenmann & Eigenmann, 1888 by Slobodian, Pastana, 2018Slobodian V, Pastana MNL. Description of a new Pimelodella (Siluriformes: Heptapteridae) species with a discussion on the upper pectoral girdle homology of Siluriformes. J Fish Biol. 2018; 93(5):901–16. https://doi.org/10.1111/jfb.13795
https://doi.org/10.1111/jfb.13795...
). For Gymnotiformes, anterior and/or posterior connections were described for Orthosternarchus species (Albert, 2001Albert JS. Species diversity and phylogenetic systematics of American knifefishes (Gymnotiformes: Teleostei). Misc Publ Mus Zool Univ Mich. 2001; 190:1–127.) and for some other apteronotids (Bernt et al., 2018Bernt MJ, Crampton WGR, Orfinger AB, Albert JS. Melanosternarchus amaru, a new genus and species of electric ghost knifefish (Gymnotiformes: Apteronotidae) from the Amazon Basin. Zootaxa. 2018; 4378(4):451–79. https://doi.org/10.11646/zootaxa.4378.4.1
https://doi.org/10.11646/zootaxa.4378.4....
) and appear to be an uncommon but phylogenetically widely-distributed feature in Otophysi.
A single pore dorsally located on the basis of the pectoral fin, the otic pit, sunk in the surrounded skin is of particular interest since it delimits an internal cavity that comprises the interface of the cephalic lateral-line canals and the trunk (posterior lateral-line) canal. In a deeper dissection, we noted that the internal wall, just opposite the pore of the cavity, has a thin membrane that covers a large lateral chamber filled with a lymph-like fluid, similar to those observed in many other fishes, such as the catfish Ancistrus sp. and cobitids (Bleckmann et al., 1991Bleckmann HU, Niemann U, Firtzsch B. Peripheral and central aspects of the acoustic and lateral line system of a bottom dwelling catfish, Ancistrus sp. J Comp Neurol. 1991; 314(3):452–66. https://doi.org/10.1002/cne.903140304
https://doi.org/10.1002/cne.903140304...
; Kratochvil, Ladich, 2000Kratochvil H, Ladich F. Auditory role of lateral trunk channels in cobitid fishes. J Comp Physiol A. 2000; 186:279–285. https://doi.org/10.1007/s003590050428
https://doi.org/10.1007/s003590050428...
).
The otic pit cavity is located in the same position of the structure described as a pseudotympanum by Dutra et al., (2015)Dutra GM, Jerep FC, Vari RP, de Santana CD. The pseudotympanum in the Gymnotiformes (Teleostei, Ostariophysi, Otophysi): homology and evolution of a previously unexplored system in Neotropical electric fishes. Zool J Linn Soc. 2015; 174(1):114–29. https://doi.org/10.1111/zoj.12221
https://doi.org/10.1111/zoj.12221...
for a 28.2 cm TL specimen of E. varii (listed therein as Electrophorus electricus (Linnaeus, 1766)). A pseudotympanum has been described as a hiatus devoid of musculature that is thought to enhance the reception of external body vibration (Birindelli, Shibatta, 2011Birindelli JLO, Shibatta OA. Morphology of the gas bladder in bumblebee catfishes (Siluriformes, Pseudopimelodidae). J Morphol. 2011; 272(7):890–96. https://doi.org/10.1002/jmor.10961
https://doi.org/10.1002/jmor.10961...
; Dutra et al., 2015Dutra GM, Jerep FC, Vari RP, de Santana CD. The pseudotympanum in the Gymnotiformes (Teleostei, Ostariophysi, Otophysi): homology and evolution of a previously unexplored system in Neotropical electric fishes. Zool J Linn Soc. 2015; 174(1):114–29. https://doi.org/10.1111/zoj.12221
https://doi.org/10.1111/zoj.12221...
). In our analyses we have not found any evidence of an empty area in adult specimens, but instead observed the expansion of the sinking canal of the otic pit in a walled chamber at this location (S2C–D).
Moreover, the connection of cranial canals (SO and IO via O, and OP) to the trunk lateral-line canal and to a lateral chamber, supposedly related to the auditory region, resembles the recessus lateralis of the lateral-line system described for clupeomorph fishes (Blaxter, 1987Blaxter JHS. Structure and development of the lateral line. Biol Rev. 1987; 62(4):471–514. https://doi.org/10.1111/j.1469-185X.1987.tb01638.x
https://doi.org/10.1111/j.1469-185X.1987...
; Di Dario, 2004Di Dario F. Homology between the recessus lateralis and cephalic sensory canals, with the proposition of additional synapomorphies for the Clupeiformes and the Clupeoidei. Zool J Linn Soc. 2004;141(2):257–70. https://doi.org/10.1111/j.1096-3642.2004.00122.x
https://doi.org/10.1111/j.1096-3642.2004...
). By unraveling novelties about cephalic lateral line structure and organization on E. varii, this work marks the starting point of a detailed investigation to understand the functional relevance of the cephalic lateral line system in those organisms and its connections with the other sensorial systems in Electrophorus varii.
ACKNOWLEDGEMENTS
The authors acknowledge the contributions of two anonymous reviewers as well as the editor William Crampton to the manuscript. The authors thanks the FAPESP/Smithsonian project # 2016/19075–9 “Diversity and Evolution of Gymnotiformes”, the MZUSP Laboratório de Processamento de Imagens de Microtomografia Computadorizada de Alta Resolução for generating images of μCT scan, Alberto Carvalho and Luiz Peixoto for editing the images. GV thanks his ancestors for opening his paths. GV received a grant from the Fundação de Amparo à Pesquisa do Estado do Amazonas-FAPEAM. AS receives fellowship from FAPEAM (FIXAM #062.01520/2018). DAB was funded by the Conselho Nacional de Desenvolvimento Científico e Tecnológico-CNPq (# 140145/2016–8). LRP and JZ received productivity grants from CNPq (#311236/2015–4 and #313183/2014–7, respectively).
REFERENCES
- Albert JS Species diversity and phylogenetic systematics of American knifefishes (Gymnotiformes: Teleostei). Misc Publ Mus Zool Univ Mich. 2001; 190:1–127.
- Albert JS, Crampton WGR Diversity and phylogeny of neotropical electric fishes (Gymnotiformes). In: Bullock TH, Hopkins CD, Popper AN, Fay RR, editors. Electroreception. New York: Springer; 2006. p.360–409.
- Arratia G, Huaquín L Morphology of the lateral line system and of the skin of diplomystid and certain primitive loricarioid catfishes and systematic and ecological considerations. Bonn Zool Monogr. 1995; 36:1–110.
- Assunção MIS, Schwassmann HO Reproduction and larval development of Electrophorus electricus on Marajó Island (Pará, Brazil). Ichthyol Explor Fres. 1995; 6(2):175–84.
- Bastos DA História Natural de poraquês (Electrophorus spp.), Gymnotiformes: Gymnotidae. [PhD Thesis] Manaus: Instituto Nacional de Pesquisas da Amazônia; 2020.
- Bastos DA, Zuanon JAS, Py-Daniel LR, de Santana CD Social predation in electric eels. Ecol Evol. 2021; 11(3):1088–92. https://doi.org/10.1002/ece3.7121
» https://doi.org/10.1002/ece3.7121 - Bernt MJ, Crampton WGR, Orfinger AB, Albert JS Melanosternarchus amaru, a new genus and species of electric ghost knifefish (Gymnotiformes: Apteronotidae) from the Amazon Basin. Zootaxa. 2018; 4378(4):451–79. https://doi.org/10.11646/zootaxa.4378.4.1
» https://doi.org/10.11646/zootaxa.4378.4.1 - Birindelli JLO, Shibatta OA Morphology of the gas bladder in bumblebee catfishes (Siluriformes, Pseudopimelodidae). J Morphol. 2011; 272(7):890–96. https://doi.org/10.1002/jmor.10961
» https://doi.org/10.1002/jmor.10961 - Blaxter JHS Structure and development of the lateral line. Biol Rev. 1987; 62(4):471–514. https://doi.org/10.1111/j.1469-185X.1987.tb01638.x
» https://doi.org/10.1111/j.1469-185X.1987.tb01638.x - Blaxter JHS, Fuiman LA Function of the free neuromasts of marine teleost larvae. In: Coombs S, Görner P, Münz H, editors. The mechanosensory lateral line: Neurobiology and evolution. New York: Springer-Verlag; 1989. p.481–99.
- Bleckmann HU, Niemann U, Firtzsch B Peripheral and central aspects of the acoustic and lateral line system of a bottom dwelling catfish, Ancistrus sp. J Comp Neurol. 1991; 314(3):452–66. https://doi.org/10.1002/cne.903140304
» https://doi.org/10.1002/cne.903140304 - Bockmann FA, Miquelarena AA Anatomy and phylogenetic relationships of a new catfish species from northeastern Argentina with comments on the phylogenetic relationships of the genus Rhamdella Eigenmann and Eigenmann 1888 (Siluriformes, Heptapteridae). Zootaxa. 2008; 1780(1):1–54.
- Branson BA, Moore GA The lateralis components of the acoustico-lateralis system in the sunfish family Centrarchidae. Copeia. 1962; 1962(1):1–108. https://doi.org/10.2307/1439483
» https://doi.org/10.2307/1439483 - Britz R, Moritz T Reinvestigation of the osteology of the miniature African freshwater fishes Cromeria and Grasseichthys (Teleostei, Gonorynchiformes, Kneriidae), with comments on kneriid relationships. Zoosyst Evol. 2007; 83(1):3–42. https://doi.org/10.1002/mmnz.200600016
» https://doi.org/10.1002/mmnz.200600016 - Catania K The shocking predatory strike of the electric eel. Science. 2014; 346(6214):1231–34. https://doi.org/10.1126/science.1260807
» https://doi.org/10.1126/science.1260807 - Catania KC The astonishing behavior of electric eels. Front Integr Neurosci. 2019; 13(1):23. https://doi.org/10.3389/fnint.2019.00023
» https://doi.org/10.3389/fnint.2019.00023 - Coombs S, Braun CB Information processing by the lateral line system. In: Collin SP, Marshall NJ, editors. Sensory processing in aquatic environments. New York: Springer; 2003. p.122–38.
- Coombs S, Braun CB, Donovan B Orienting response of Lake Michigan mottled sculpin is mediated by canal neuromasts. J Exp Biol. 2001; 204(2):337–48. Available from: https://jeb.biologists.org/content/204/2/337
» https://jeb.biologists.org/content/204/2/337 - Crampton WGR Electroreception, electrogenesis and electrical signal evolution. J Fish Biol. 2019; 95(1):92–134. https://doi.org/10.1111/jfb.13922
» https://doi.org/10.1111/jfb.13922 - Crampton WGR, Albert JS Evolution of electric signal diversity in gymnotiform fishes. In: Ladich F, Collin SP, Moller P, Kapoor BG, editors. Communication in fishes. Gwalior: Science Publishers, 2006. p.647–731.
- Crampton WGR, Jonathan KW, Smyth C, Walz SA Design and construction of an Electric Fish Finder. Neotrop Ichthyol. 2007; 5(3):425–28. https://doi.org/10.1590/S1679-62252007000300022
» https://doi.org/10.1590/S1679-62252007000300022 - Crampton WGR, de Santana CD, Wadell JC, Lovejoy NR A taxonomic revision of theNeotropical electric fish genus Brachyhypopomus (Ostariophysi: Gymnotiformes: Hypopomidae), with descriptions of 15 new species. Neotrop Ichthyol. 2016, 14(4):e150146. http://dx.doi.org/10.1590/1982-0224-20150146
» http://dx.doi.org/10.1590/1982-0224-20150146 - Di Dario F Homology between the recessus lateralis and cephalic sensory canals, with the proposition of additional synapomorphies for the Clupeiformes and the Clupeoidei. Zool J Linn Soc. 2004;141(2):257–70. https://doi.org/10.1111/j.1096-3642.2004.00122.x
» https://doi.org/10.1111/j.1096-3642.2004.00122.x - Di Dario F, de Pinna MC The supratemporal system and the pattern of ramification of cephalic sensory canals in Denticeps clupeoides (Denticipitoidei, Teleostei): additional evidence for monophyly of Clupeiformes and Clupeoidei. Pap Avulsos Zool. 2006; 46(10):107–23. https://doi.org/10.1590/S0031-10492006001000001
» https://doi.org/10.1590/S0031-10492006001000001 - Dutra GM, Jerep FC, Vari RP, de Santana CD The pseudotympanum in the Gymnotiformes (Teleostei, Ostariophysi, Otophysi): homology and evolution of a previously unexplored system in Neotropical electric fishes. Zool J Linn Soc. 2015; 174(1):114–29. https://doi.org/10.1111/zoj.12221
» https://doi.org/10.1111/zoj.12221 - Gotter AL, Kaetzel MA, Dedman JR Electrophorus electricus as a model system for the study of membrane excitability. Comp Biochem Physiol A Mol Integr Physiol. 1998; 119(1):225–41. https://doi.org/10.1016/S1095-6433(97)00414-5
» https://doi.org/10.1016/S1095-6433(97)00414-5 - Hassan ES Hydrodynamic imaging of the surroundings by the lateral line of the blind cave fish Anoptichtys jordani In: Coombs S, Görner P, Münz H, editors. The Mechanosensory Lateral Line. New York: Springer Verlag; 1989. p.217–27.
- Hagiwara S, Szabo T, Enger PS Physiological properties of electroreceptors in the electric eel, Electrophorus electricus J Neurophysiol. 1965; 28(5):775–83. https://doi.org/10.1152/jn.1965.28.5.775
» https://doi.org/10.1152/jn.1965.28.5.775 - Kratochvil H, Ladich F Auditory role of lateral trunk channels in cobitid fishes. J Comp Physiol A. 2000; 186:279–285. https://doi.org/10.1007/s003590050428
» https://doi.org/10.1007/s003590050428 - Lekander B The sensory line system and the canal bones in the head of some Ostariophysi. Acta Zool. 1949; 30(1-2):1–131. https://doi.org/10.1111/j.1463-6395.1949.tb00503.x
» https://doi.org/10.1111/j.1463-6395.1949.tb00503.x - Liang XF, Liu JK, Huang BY The role of sense organs in the feeding behaviour of Chinese perch. J Fish Biol. 1998; 52(5):1058–67. https://doi.org/10.1111/j.1095-8649.1998.tb00603.x
» https://doi.org/10.1111/j.1095-8649.1998.tb00603.x - Mathias JA, LI S Feeding habits of walleye larvae and juveniles: Comparative laboratory and field studies. T Am Fish Soc. 1982; 111(6):722–35. https://doi.org/10.1577/1548-8659(1982)111<722:FHOWLA>2.0.CO;2
» https://doi.org/10.1577/1548-8659(1982)111<722:FHOWLA>2.0.CO;2 - McAllister DE Mandibular pore pattern in the sculpin family Cottidae. Bull Nat Mus Can Contr Zool. 1968; 223:58–69.
- Mendes-Junior RNG, Oliveira JCS, Ferrari SS Biology of the electric eel, Electrophorus electricus, Linnaeus, 1766 (Gymnotiformes: Gymnotidae) on the floodplain of the Curiaú River, eastern Amazonia. Rev Fish Biol Fisheries. 2016; 26:83–91. https://doi.org/10.1007/s11160-015-9407-9
» https://doi.org/10.1007/s11160-015-9407-9 - Moller P Electric fishes: history and behavior. Springer; 1995.
- Mukai Y, Tuzan AD, Lim LS, Wahid N, Sitti Raehanah MS, Senoo S Development of sensory organs in larvae of African catfish Clarias gariepinus. J Fish Biol. 2008; 73(7):1648–61. https://doi.org/10.1111/j.1095-8649.2008.02038.x
» https://doi.org/10.1111/j.1095-8649.2008.02038.x - Northcutt RG The phylogenetic distribution and innervation of craniate mechanoreceptive lateral lines. In: Coombs S, Görner P, Müns H, editors. The mechanosensory lateral line: neurobiology and evolution. New York: Springer; 1989. p.17–78.
- Oliveira MSB, Mendes-Júnior RNG, Tavares-Dias M Diet composition of the electric eel Electrophorus voltai (Pisces: Gymnotidae) in the Brazilian Amazon region. J Fish Biol. 2020. https://doi.org/10.1111/jfb.14413
» https://doi.org/10.1111/jfb.14413 - Pastana MNL Canais e poros do sistema látero-sensorial cefálico de Characiformes (Ostariophysi): anatomia e seu significado filogenético. [Master Dissertation] Ribeirão Preto: Universidade de São Paulo; 2014. Available from: https://www.teses.usp.br/teses/disponiveis/59/59139/tde-06012015-123252/pt-br.php
» https://www.teses.usp.br/teses/disponiveis/59/59139/tde-06012015-123252/pt-br.php - Pastana MNL, Bockmann FA, Datovo A The cephalic lateral-line system of Characiformes (Teleostei: Ostariophysi): anatomy and phylogenetic implications. Zool J Linn Soc. 2020; 189(1):1–46. https://doi.org/10.1093/zoolinnean/zlz105
» https://doi.org/10.1093/zoolinnean/zlz105 - Rizzato PP, Pospisilova A, Hilton EJ, Bockmann FA Ontogeny and homology of cranial bones associated with lateral-line canals of the Senegal Bichir, Polypterus senegalus (Actinopterygii: Cladistii: Polypteriformes), with a discussion on the formation of lateral-line canal bones in fishes. J Anat. 2020; 237(3):439–67. https://doi.org/10.1111/joa.13202
» https://doi.org/10.1111/joa.13202 - de Santana CD, Crampton WGR A review of the South American electric fish genus Porotergus (Gymnotiformes: Apteronotidae) with the description of a new species. Copeia. 2010; 2010(1):165–75. https://doi.org/10.1643/CI-05-136
» https://doi.org/10.1643/CI-05-136 - de Santana CD, Crampton WGR, Dillman CB, Frederico RG, Sabaj MH, Covain R et al Unexpected species diversity in electric eels with a description of the strongest living bioelectricity generator. Nat Commun. 2019; 10:4000. https://doi.org/10.1038/s41467-019-11690-z
» https://doi.org/10.1038/s41467-019-11690-z - Satou M, Takeuchi HA, Nishii J, Tanabe M, Kitamura S, Okumoto N, Iwata M Behavioral and electrophysiological evidences that the lateral line is involved in the inter-sexual vibrational communication of the himé salmon (landlocked red salmon, Oncorhynchus nerka). J Comp Physiol A. 1994; 174:539–49. https://doi.org/10.1007/BF00217373
» https://doi.org/10.1007/BF00217373 - Schwassmann HO, Assunção MIS, Kirschbaum F Ontogeny of the electric organs in the electric eel, Electrophorus electricus: Physiological, histological and fine structural investigations. Brain Behav Evol. 2014; 84(4):288–302. https://doi.org/10.1159/000367884
» https://doi.org/10.1159/000367884 - Slobodian V, Pastana MNL Description of a new Pimelodella (Siluriformes: Heptapteridae) species with a discussion on the upper pectoral girdle homology of Siluriformes. J Fish Biol. 2018; 93(5):901–16. https://doi.org/10.1111/jfb.13795
» https://doi.org/10.1111/jfb.13795 - Stephens RR A description of the cephalic lateralis system of Anchoa mitchilli (Valenciennes) (Clupeomorpha: Engraulidae) with identification of synapomorphies for the Engraulidae. P Biol Soc Wash. 2010; 123(1):8–16. https://doi.org/10.2988/09-13.1
» https://doi.org/10.2988/09-13.1 - Szabo T Sense organs of the lateral line system in some electric fish of the Gymnotidae, Mormyridae and Gymnarchidae. J Morphol. 1965; 117(2):229–49. https://doi.org/10.1002/jmor.1051170208
» https://doi.org/10.1002/jmor.1051170208 - Szabo T The origin of electric organs of Electrophorus electricus Anat Rec. 1966; 155(1):103–10. https://doi.org/10.1002/ar.1091550112
» https://doi.org/10.1002/ar.1091550112 - Szabo T Anatomy of the specialized lateral line organs of electroreception. In: Fessard A, editor. Electroreceptors and other specialized receptors in lower vertebrates. Handbook of Sensory Physiology, vol. 3. Berlin: Springer, Heidelberg; 1974. p.13–58.
- Tarby ML, Webb JF Development of the supraorbital and mandibular lateral line canals in the cichlid, Archocentrus nigrofasciatus J Morphol. 2003; 255(1):44–57. https://doi.org/10.1002/jmor.10045
» https://doi.org/10.1002/jmor.10045 - Verçoza G Desenvolvimento ontogenético do poraquê Electrophorus varii (Gymnotiformes: Gymnotidae). [Master dissertation] Manaus: Universidade Federal do Amazonas; 2020. Available from: https://tede.ufam.edu.br/bitstream/tede/7916/9/Disserta%c3%a7%c3%a3o_GabrielVer%c3%a7oza_PPGZOO.pdf
» https://tede.ufam.edu.br/bitstream/tede/7916/9/Disserta%c3%a7%c3%a3o_GabrielVer%c3%a7oza_PPGZOO.pdf - Webb J Morphological diversity, development and evolution of the mechanosensory lateral line system. In: Coombs S, Bleckmann H, Fay R, Popper A, editors. The Lateral Line System. Springer Handbook of Auditory Research, vol. 48. New York: Springer; 2013. p.17–72.
- Webb JF, Shirey JE Postembryonic development of the cranial lateral line canals and neuromasts in zebrafish. Dev Dyn. 2003; 228(3):370–85. https://doi.org/10.1002/dvdy.10385
» https://doi.org/10.1002/dvdy.10385 - Westby GWM The ecology, discharge diversity and predatory behaviour of gymnotiform electric fish in the coastal streams of French Guiana. Behav Ecol Sociobiol. 1988; 22:341–54. https://doi.org/10.1007/BF00295103
» https://doi.org/10.1007/BF00295103
ADDITIONAL NOTES
-
HOW TO CITE THIS ARTICLE
Verçoza G, Shibuya A, Bastos DA, Zuanon J, Rapp Py-Daniel LH. Organization of the cephalic lateral-line canals in Electrophorus varii de Santana, Wosiacki, Crampton, Sabaj, Dillman, Mendes-Júnior & Castro e Castro, 2019 (Gymnotiformes: Gymnotidae). Neotrop Ichthyol. 2021; 19(1):e200075. https://doi.org/10.1590/1982-0224-2020-0075
Edited-by
Publication Dates
-
Publication in this collection
11 June 2021 -
Date of issue
2021
History
-
Received
6 Aug 2020 -
Accepted
5 Mar 2021