Abstracts
The Lumbrera Formation is the uppermost unit of the Salta Group, which crops out in northwestern Argentina. The paleoenvironment of the Lumbrera Formation is interpreted as a perennial lake deposited under temperate climatic conditions during the early to middle Eocene. Its fossil content is made up of palynomorphs, insects, crocodiles, turtles, lizards, and mammals, besides an ichthyofauna formed by cichlids, poeciliids and dipnoans. †Plesioheros chauliodus is described based on a single individual from this formation, which was fossilized as a lateral view impression (missing anal and caudal fins). It can be distinguished from other cichlids by a moderately deep body, enlarged anterior dentary teeth bearing subapical cusp, a low abdominal vertebral count (10), five canal openings in the dentary, and XI + 12 dorsal-fin rays. A phylogenetic analysis, using the matrix by Kullander (1998), recovered †Plesioheros within Heroini. This species was recovered most closely related to Australoheros and to the deep-bodied South American heroins. The occurrence of an Eocene Heroini, as well as of other cichlid lineages in the same stratigraphical level, is evidence of an ancient diversification in this family. This ancient age supports the hypothesis that the Cichlidae originated on Gondwana.
Argentina; Cichlinae; Fossil cichlid; Lumbrera Formation; Plesioheros chauliodus; Ypresian-Lutetian
A Formação Lumbrera é a unidade do topo do Grupo Salta, aflorante na região noroeste da Argentina. O paleoambiente da Formação Lumbrera tem sido interpretado como um lago perene depositado sob um clima temperado durante o início do Eoceno. Seu conteúdo fóssil é formado por palinomorfos, insetos, crocodilos, tartarugas, lagartos, mamíferos, além de uma ictiofauna que inclui ciclídeos, poeciliídeos e dipnóicos. †Plesioheros chauliodus é descrito com base em um único indivíduo coletado nesta formação, preservado como impressão em vista lateral (faltando as nadadeiras anal e caudal). Ele pode ser distinguido de outros ciclídeos por um corpo moderadamente alto, dentes anteriores do dentário aumentados e portando uma cúspide lingual subapical, uma baixa contagem de vértebras abdominais, cinco aberturas do canal sensorial no dentário, e XI + 12 raios na dorsal. Na análise filogenética, usando a matriz apresentada por Kullander (1998), †Plesioheros resultou dentro da Tribo Heroini mais relacionado a Australoheros e aos heroinis de corpo alto sul-americanos. A ocorrência de um heroini fóssil desta idade, bem como das outras linhagens de ciclídeos no mesmo nível estratigráfico, é uma evidência da diversificação inicial da família, corroborando fortemente a hipótese de que os Cichlidae se originaram no Gondwana ainda durante o Cretáceo.
A new genus and species of Heroini (Perciformes: Cichlidae) from the early Eocene of southern South America
Patricia Alano PerezI; Maria Claudia MalabarbaI; Cecilia del PapaII
IMuseu de Ciências e Tecnologia, Pontifícia Universidade Católica do Rio Grande do Sul. Av. Ipiranga, 6681, 90619-900 Porto Alegre, RS, Brazil. patriza@hotmail.com, mariacm@pucrs.br
IICONICET, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba. Av. Velez Sarsfield 1611, Córdoba, Argentina. delpapacecilia@efn.uncor.edu
ABSTRACT
The Lumbrera Formation is the uppermost unit of the Salta Group, which crops out in northwestern Argentina. The paleoenvironment of the Lumbrera Formation is interpreted as a perennial lake deposited under temperate climatic conditions during the early to middle Eocene. Its fossil content is made up of palynomorphs, insects, crocodiles, turtles, lizards, and mammals, besides an ichthyofauna formed by cichlids, poeciliids and dipnoans. †Plesioheros chauliodus is described based on a single individual from this formation, which was fossilized as a lateral view impression (missing anal and caudal fins). It can be distinguished from other cichlids by a moderately deep body, enlarged anterior dentary teeth bearing subapical cusp, a low abdominal vertebral count (10), five canal openings in the dentary, and XI + 12 dorsal-fin rays. A phylogenetic analysis, using the matrix by Kullander (1998), recovered †Plesioheros within Heroini. This species was recovered most closely related to Australoheros and to the deep-bodied South American heroins. The occurrence of an Eocene Heroini, as well as of other cichlid lineages in the same stratigraphical level, is evidence of an ancient diversification in this family. This ancient age supports the hypothesis that the Cichlidae originated on Gondwana.
Key words: Argentina, Cichlinae, Fossil cichlid, Lumbrera Formation, †Plesioheros chauliodus, Ypresian-Lutetian.
RESUMO
A Formação Lumbrera é a unidade do topo do Grupo Salta, aflorante na região noroeste da Argentina. O paleoambiente da Formação Lumbrera tem sido interpretado como um lago perene depositado sob um clima temperado durante o início do Eoceno. Seu conteúdo fóssil é formado por palinomorfos, insetos, crocodilos, tartarugas, lagartos, mamíferos, além de uma ictiofauna que inclui ciclídeos, poeciliídeos e dipnóicos. †Plesioheros chauliodus é descrito com base em um único indivíduo coletado nesta formação, preservado como impressão em vista lateral (faltando as nadadeiras anal e caudal). Ele pode ser distinguido de outros ciclídeos por um corpo moderadamente alto, dentes anteriores do dentário aumentados e portando uma cúspide lingual subapical, uma baixa contagem de vértebras abdominais, cinco aberturas do canal sensorial no dentário, e XI + 12 raios na dorsal. Na análise filogenética, usando a matriz apresentada por Kullander (1998), †Plesioheros resultou dentro da Tribo Heroini mais relacionado a Australoheros e aos heroinis de corpo alto sul-americanos. A ocorrência de um heroini fóssil desta idade, bem como das outras linhagens de ciclídeos no mesmo nível estratigráfico, é uma evidência da diversificação inicial da família, corroborando fortemente a hipótese de que os Cichlidae se originaram no Gondwana ainda durante o Cretáceo.
Introduction
The cichlids comprise the most diverse non-ostariophysian freshwater fish group in the world. They constitute one of the most speciose vertebrate families including more than 1500 valid species (Eschmeyer & Fong, 2010) with estimates exceeding 1900 species (Kullander, 2003). Due to this richness, and their ecological and evolutionary diversity, cichlids are among the most widely studied fishes.
Nevertheless, the cichlid fossil record is poor. Cichlid fossils are limited to few species in Africa, Europe, the Neotropics, and the Near East. In South America, cichlids are recorded for the Eocene-Oligocene [Tremembichthys pauloensis (Schaeffer, 1947); T. garciae Malabarba & Malabarba, 2008] and Pliocene (Macracara prisca Woodward, 1939) from Brazil; and for the Miocene [Aequidens saltensis Bardack, 1961; Paleocichla longirostrum (Bardack, 1961)] and Eocene [Piroterocara argentina Malabarba et al., 2006; Gymnogeophagus eocenicus Malabarba et al., 2010] from Argentina. The first cichlid record for the American continent is the heroin Nandopsis woodringi (Cockerell, 1924; Chakrabarty, 2006) from the Miocene of Haiti (Hispaniola).
The Lumbrera Formation is the top unit of the Salta Group, cropping out in northwestern Argentina (Fig. 1). It constitutes a complex continental Eocene lithostratigraphic unit divided in two informal members: the lower and upper Lumbrera (Gómez Omil et al., 1989; del Papa, 2006). The specimen described here comes from the uppermost section of the Lower Lumbrera known as "Faja Verde" because of its lacustrine origin. The age constraint of the Faja Verde Lake is not precisely dated, and mostly estimated on the basis of regional considerations, the mammalian fossil record, and an absolute U/Pb zircon date (Salfity & Marquillas, 1994; del Papa et al., 2010); which in conjunction point to a Lower Eocene age. The strongest dating evidence is provided by an U/Pb zircon dating of 39.9 Ma from a tuff bed stratigraphically located 240 m above the fossiliferous level (del Papa et al., 2010). Other indirect evidence comes from the paleoclimatic inferences based on the study of aluvial paleosol horizons in the Lumbrera Formation, where a stratigraphic fluvial section correlative to the Faja Verde Lake, has been preliminarily assigned to the Early Eocene Climatic Optimum (EECO; White et al., 2009). Finally, the mammalian fossil association suggests a Casamayoran-Vacan SALMA age, for the deposits studied here (see discussion in del Papa et al., 2010). The overall geological evidence points to a Lower Eocene, more precisely upper Ypresian-Lower Lutetian (~ 48.6 Ma) age for the Faja Verde of the Lumbrera Formation.
The debate about the origin and biogeography of cichlids has been recently stimulated by phylogenetic studies, mainly based on molecular data. In this context, each cichlid fossil record contributes information which can eventually be crucial to understanding the origin and evolution of the group. This paper deals with a new fossil cichlid represented by the impression of a single articulated individual. Despite the limitations of this information that can be obtained from the specimen, there are sufficient features observable to diagnose it as a new cichlid related to the Heroini.
Material and Methods
Material. The fossil material was collected in the Faja Verde of the Lumbrera Formation, at Alemanía in northwestern Argentina (Fig. 1). It is made up of a single specimen, articulated and almost complete, preserved as impression in lateral view. Poor preservation quality prevents the observation of most anatomical details. The fossil is deposited in the Universidad Nacional de Salta, in Salta, Argentina (CNS-V10026).
Methods: Given that the fossil is preserved as an impression, bones, scales and teeth are represented in a negative relief. To optimize the observation and interpretation of anatomical structures, a silicone peel was made. The peel was sprinkled with ammonium chloride, allowing better visualization and photography.
Measurements were made with a digital caliper with data recorded to tenths of millimeters. Measurements follow Reis & Malabarba (1988) and Casciotta & Arratia (1993a). Standard length was estimated from the tip of the snout to a projected vertical line at the most posterior point of the dorsal margin of the caudal peduncle. Counts of fin spines are indicated by upper case Roman numerals and soft rays by Arabic numerals. General osteological terminology follows Kullander (1986) and Reis & Malabarba (1988). The terminology, measurements and angles adopted for jaw descriptions follow Casciotta & Arratia (1993a). The term coulter area is that presented by Casciotta & Arratia (1993a) to indicate the portion of the suspensorium formed by the base of the anguloarticular and the retroarticular. The classification of cichlids follows Smith et al. (2008), with the Neotropical cichlids grouped into the subfamily Cichlinae, which includes seven tribes: Cichlini, Retroculini, Astronotini, Chaetobranchini, Geophagini, Heroini and Cichlasomatini. We also follow Smith et al. (2008), in using for informal suprageneric names with the suffix -ine(s) for subfamilies (e.g. "cichlines" for Cichlinae) and -in(s) for tribe (e.g. "heroins" for Heroini).
The numerical ages (in Ma) provided for the geological time periods are based on the Standard Global Chronostratigraphic Scale (Ogg et al., 2008).
Cladistic methodology. The phylogenetic position of †Plesioheros (CNS-V10024) is discussed based on the hypothesis of Kullander (1998), which includes 50 Recent cichlid taxa and 91 characters. †Plesioheros was added to the matrix of Kullander (1998) in the Winclada software. Only 32 characters could be coded for †Plesioheros (states in parentheses): 15(2), 16(1), 17(0), 25(0), 34(1), 35(0), 36(0), 38(1), 39(1), 40(0), 41(1), 42(1), 43(0),44(2), 45(0), 46(0), 47(1), 48(0), 51(0), 63(1), 64(2), 65(1), 66(0), 67(1), 69(0), 70(2), 73(0), 77(1),79(0), 84(1), 85(1), 87(0); the other 59 characters are informed as missing data ("?"). The phylogenetic analysis was performed using TNT 1.1, Traditional Search TRB, and not ordered characters, resulting in three equally parsimonious trees: 735 steps, CI 20, RI 50. The differences among the trees were restrict to the arrangement of some Geophagini genera.
Institutional abbreviations. CNS-V, collection of paleontology, Facultad de Ciencias Naturales, Universidad Nacional de Salta, Salta; MCP, Ichthyology, Museu de Ciências e Tecnologia, Porto Alegre; UFRGS, Ichthyology, Universidade Federal do Rio Grande do Sul, Porto Alegre; UNG-T, Universidade de Guarulhos, Guarulhos.
Results
CICHLIDAE Bonaparte, 1840
CICHLINAE Bonaparte, 1840 (sensu Smith et al., 2008)
HEROINI Kullander, 1998 (sensu Smith et al., 2008)
Plesioheros, new genus
Type species.Plesioheros chauliodus, new species
Diagnosis. Heroin cichlid distinguished from other cichlid genera by the following combination of characters: premaxilla with equally sized conical teeth over the entire length of the dentigerous arm; lower jaw with anterior teeth enlarged and bearing a small subapical cusp; presence of spatulate teeth in the lower jaw; mandibular canal with five foramina; two supraneurals; 10 abdominal vertebrae with equal or higher number of caudal vertebrae; dorsal fin rays XI + 12.
Etymology. From the Greek words plesios, meaning near, and heros, after the nominotypical genus of Heroini.
Plesioheros chauliodus, new species
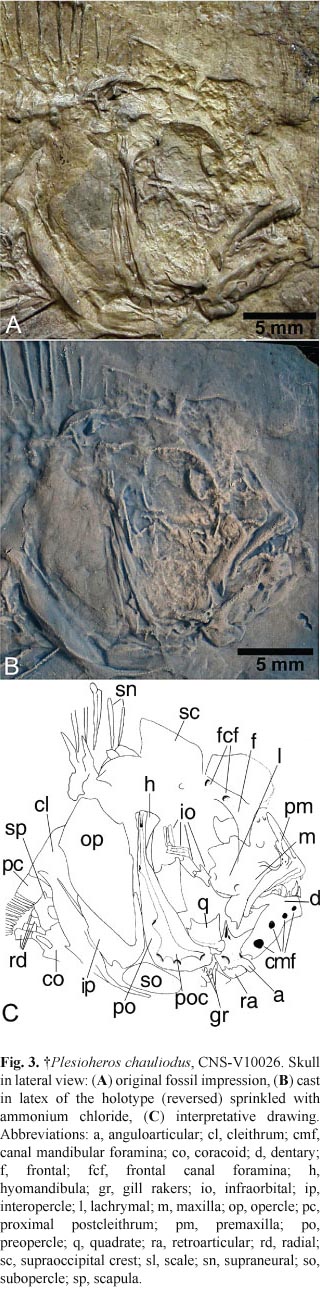
Holotype. CNS-V10026, an articulated individual preserved as an impression in lateral view, with the posteroventral region of the body missing (Fig. 2).
Locality. Alemanía, northwestern Argentina, level Faja Verde II, Lumbrera Formation, Lower Eoceno (~ 48 Ma).
Diagnosis. As for the genus.
Etymology. From the Greek word chauliodous (χαυλιοδουσ), meaning having large teeth, referring to the enlarged dentary teeth of the specimen. Treated as noun in apposition.
Description
Shape. Morphometric data for †Plesioheros chauliodus is presented in Table 1. †Plesioheros is a deep-bodied cichlid with the greatest body depth is at about the seventh spine of the dorsal fin. The dorsal contour of head is steep, slightly arched from the snout to the supraoccipital crest. The dorsal body contour is convex from the dorsal-fin origin to the caudal peduncle. As judged from the prepelvic region the ventral body contour is curved.
Skull. The skull is roughly round in lateral view, a little deeper (21.53 mm) than long (17.08 mm), with a rounded snout and a terminal, slightly inferior mouth (Fig. 3). The anteroventral portion of the vomer is preserved and presents a rounded outline. The frontal follows the contours of the orbit dorsally and contacts the lateral ethmoid anteroventrally and the supraoccipital posteriorly, but most of its surface is concealed by the scales. The supraoccipital crest is moderately high with the posterior margin slightly curved. Part of the sensory canal system and three foramina can be observed in the frontal. The median frontal lateralis openings are coalesced forming a single coronalis foramen (NLF0 of Stiassny, 1991) not covered by the supraoocipital. The parietal crest is low, starting in the posterior region of the frontal, near frontal lateralis foramen 3 and extending through the parietal and epiotic. The mesethmoid and lateral ethmoid are represented by impressions inside the orbit.
Over the anterior region of the skull roof, dorsal to frontal, there is a weak impression of a roundish protuberance, apparently covered by small scales. Although the bad preservation does not allow a more secure identification, it could be a "nuchal hump", very common in males of some cichlid species.
Circumorbital series. The orbital region show signs of distortion and disarticulation. The orbit is nearly oval, with the vertical diameter (6.01 mm) longer than the horizontal (5.06 mm). Only the posterior half of the lachrymal is visible, it appears nearly square with the posteriordorsal corner projected and extending over infraorbital 2. Only two pores of the sensory canal are observed on the lachrymal: a posteroventral and a posterodorsaly which is directed to infraorbital 1. Posteriorly to the lachrymal, there are four infraorbitals. They are short, nearly tubular and slightly dislocated from the original positions.
Jaws. The mouth is terminal and short, with the upper jaw slightly longer than the lower (Fig. 4). The premaxilla is robust with a round symphysial region. The ascending arm is barely longer (5.72 mm) than the dentigerous arm (5.69 mm); however, as its spine is not completely preserved dorsally, we are assuming the ascending arm is a little longer. The confluence of the ascending and dentigerous arms forms a slightly obtuse angle (97°). The ascending arm exceeds half of the orbit diameter; it is wide (width 27% of height) with an articular process moderately distinct. The dentigerous arm is slightly curved ventrally with no symphysial extension or processes.
The dentary is short and deep with the posterodorsaly portion (coronoid process) concealed by the maxilla. There are five openings in the dentary for the lateral canal. The anguloarticular is deeper (4.73 mm) than long (4.16 mm), with the primordial process wide, low and slightly forward projected. The coulter area is nearly squared, almost so deep (2.16 mm) as wide (2.18 mm). The section of the mandibular canal in the anguloarticular is short and slightly oblique. The posterior border of the coulter area is slightly curve, almost straight with the retroarticular forming the posteroventral region.
The hyomandibular has a clear vertical crest and is dorsally expanded; it is ventrally sutured to mesopterygoid. The quadrate is typically triangular with the ventral border following the horizontal arm of the preopercle. The robust condyle articulates with the mandible at the level of the middle of the orbit.
Teeth. In the premaxilla, impressions of alveoli and teeth are preserved in the most anterior and posterior regions of the dentigerous arm, indicating that the teeth were implanted along its entire length and arranged in three rows symphysially (Figs. 3, 4). Four conical teeth are preserved: two in the anterior region and two at the caudal end (Fig. 4). The most anterior teeth measure 0.45 mm; the posterior ones are smaller, measuring 0.2 mm.
There are seven tooth impressions in the oral border of the dentary. The teeth increase size toward the symphysis. The two most anterior teeth are conical with their apex slightly recurved and a very small lingual cusp; they measure 0.5 and 1.13 mm and probably belong to the outer row. The next posterior tooth is much smaller (0.41 mm), conical and anteriorly directed; it appears to be broken and to belong to an inner row; it is not possible determine the presence of a lingual cusp. The fourth tooth in the sequence is large (0.7 mm), conical, with the apex slightly recurved rearward. The next tooth is small (0.43 mm) and badly preserved; it appears to be pedunculate, with a triangular tip (spatulate tooth?). Posteriorly, it seems to have four more teeth in the sequence, but it is not possible to define their shapes. Based on this observation, the dentition of the lower jaw of this species appears to be made up of at least two rows: the outer row includes enlarged anterior teeth, conical, slightly curved and bearing a smaller lingual cusp; and an inner row would include smaller conical (with or without lingual cusp) and doubtfully spatulate teeth. The presence of the enlarged canines bearing an accessory lingual cusp in the outer jaw rows is exclusive of heroins among Neotropical cichlids (Casciotta & Arratia, 1993b; Kullander, 1996, 1998; Chakrabarty, 2007).
Opercular apparatus. The preopercle is narrow with the horizontal arm much shorter than the vertical arm, which form an approximately rectangle angle. It is crossed longitudinally by the preopercular sensory canal presenting two terminal and four medial pores. The opercle is trapezium-shaped with a round posteroventral angle. The subopercle is narrow with a long process projecting between the opercle and preopercle. The interopercle is ovoidal tapering rostrally. All opercular elements have smooth surfaces and borders and they have impressions of cycloid scales all over their surface (except for the preopercle).
Gill rakers. Just below the most anteroventral part of the preopercle there are impressions of three gill rakers (Fig. 3). They are short, fingerlike and aligned, close together, on its caudal border. Based on the position they fossilized, it is assumed they should belong to first gill arch.
Vertebrae and supraneurals. The vertebral column is almost completely preserved, only the most posterior region is missing including the caudal peduncle and the fin. There are 10 abdominal and 10 caudal vertebrae and a short hypapophysis is preserved in the fourth abdominal vertebra.
There are two narrow and parallel supraneurals with no neural spines associated with them (Fig. 6). Ten abdominal vertebrae is the basal percoid number and is too low for South American cichlids, which have 12-14 abdominal vertebrae (Kullander, 1998). In †Plesioheros the first preserved neural spine is projected between the second and third dorsal pterygiophores, whereas American cichlids with two supraneurals have the first neural spine inserted between the supraneurals. Therefore, it is assumed here that there was a vertebra with the neural spine inserted between the supraneurals, resulting in 11 abdominal vertebrae for †Plesioheros.
The last preserved caudal vertebra (10th vertebra) lies just before the caudal peduncle (Fig. 5). Posteriorly, there is part of the dorsal margin of the caudal peduncle where it is possible to observe 4 procurrent rays (Fig. 5). Considering the distance between the last preserved vertebra and the most anterior procurrent ray (5 mm) and the distance between the neural spines (1.2 mm), we estimate that 2-3 caudal vertebrae are missing from our vertebral count. Then, including these 2-3 missing vertebrae, †Plesioheros has 12-13 caudal vertebrae, totalizing 23-24 vertebrae.
Girdle and pectoral fin. Most of the pectoral girdle and fin are preserved. Dorsally in the girdle, there are parts of the posttemporal and the proximal (medial) extrascapular adjacent to the neurocranium. The supracleithrum is not visible. The cleithrum contours the opercle and subopercle ending in a point anteroventrally. Just above the pectoral insertion, the cleithrum has a notch with a lamina projecting for the proximal postcleithrum articulation. The fin rays conceal most of the postcleithra leaving only part of the proximal and the ventral pointed end of the distal postcleithrum observable. The scapula and coracoid are partially preserved articulating with each other and with the four rectangular radials. The pectoral fin has 12 soft rays and appears rounded.
Girdle and pelvic fin. Both pelvic basipterygia are preserved. They are robust and triangular bones, measuring 5.9 mm. Their most proximal end is covered by the cleithrum and it is not visible. Distally the pelvic bone is expanded to articulate with the rays; the processus medialis is short and stout. The articulation between basipterygium and the pelvic fin spine is strong and evident. There are I + 5 rays in the pelvic fin.
Dorsal fin. The dorsal fin is completely preserved with XI + 12 rays. The base of the spiny portion is longer (14.9 mm) than the soft portion (12.1 mm), and they are continuous with no gap between them. The first four spines increase gradually in length; the length does not vary from the 5th to 10th spine; the last one (eleventh) conspicuously longer. The soft dorsal fin is represented mainly by the proximal (unsegmented) portion of the soft rays; the last four soft rays are more completely preserved including their distal segmentation. There are 22 pterygiophores; the first three are about the same length; they decrease in length from the fourth to the last.
Anal and caudal fins. As stated above, the anal and caudal fins were not preserved. However, it is possible to observe the dorsal profile of the caudal peduncle, where 4 procurrent caudal-fin rays are present (Fig. 5). This allows us to obtain a probable length for the caudal peduncle, which was measured from the end of the dorsal-fin base to the last point preserved (7.7 mm). We estimated that 2-3 caudal vertebrae referring to the caudal peduncle were lost.
Only the presumed first anal-fin pterygiophore (it is not possible determine how many pterygiophores), whi ch is contacting the first caudal vertebra (11th vertebra), is preserved.
Scales. The squamation was preserved as impressions all over the body (Fig. 2). In the head, differently sized scales are present in the cheek and postorbital region. Cheek scales are ctenoid and irregularly distributed, with a few covering the rostral border of the preopercle. Differently sized cycloid scales fully cover the opercle, subopercle and interopercle, but not the preopercle. The scales covering the body are mainly cycloid, large sized (1.9-2.8 mm) with the chest scales slightly smaller than those on the flanks. In the occipital region, the scales are ctenoid. There is a predorsal series with about six scales (Fig. 6), decreasing in size caudad, from the supraoccipital crest to dorsal-fin origin; however, it is not possible to determine accurately whether they constitute a uniserial or triserial arrangment. There are scale impressions on the bases of spines and rays and over the soft portion of the dorsal fin.
Lateral line. As typical in cichlids, the lateral line in †Plesioheros is divided into two segments: upper (anterior) and lower (posterior). The upper segment begins next to supracleithrum region and it extends posteriorly, ascending gradually and finishing at the vertical through the 4th caudal vertebra (5th soft ray). The trajectory of the lower section of the lateral line initiates at the vertical through the 6th caudal vertebra (7th soft dorsal-fin ray), following close to the vertebral column and entering the caudal peduncle. It is counted 17 scales (impressions) in the upper and 6 in the lower lateral line; there is no overlap of the upper and lower sections. We estimate that 3 lateral line scales referring to the caudal peduncle (see Vertebrae and supraneurals) were lost, totalizing 26 scales in the lateral line. There are 3 scales between the upper lateral line and dorsal fin anteriorly, and 2 posteriorly; 2 scales between the lateral lines.
Discussion
Phylogenetic position of Plesioheros
Casciotta & Arratia (1993b) proposed the first phylogeny for the Cichlinae focusing the American cichlids based on morphological data including two fossil taxa from the Tertiary of Argentina. Later, Kullander (1998) presented the broadest morphological study emphasizing South American cichlids, based on a comprehensive data set of 51 taxa and 91 characters. Character conflict and undersampling influenced the resolution and support of the analysis (Kullander, 1998:487), which did not recover the Neotropical cichlids as monophyletic.
Later, Smith et al. (2008) combined nucleotide characters to Kullander's (1998) morphological data, and increased the number of terminals to 90. They also include the Eocene cichlid †Proterocara (also coming from the Lumbrera Formation). Although, most relationships and tribal structures were congruent with Kullander (1998), Smith et al. (2008) managed to recover a monophyletic Cichlinae. They partitioned Cichlinae in seven tribes: Retroclini, Cichlini, Astronotini, Chaetobranchini, Geophagini, Cichlasomatini and Heroini. †Proterocara was recovered not as originally hypothesized sister-group to Geophagini + Heroini + Cichlasomatini as suggested by Malabarba et al. (2006), but nested within Geophagini.
Currently, cichlids can be recognized by nine synapomorphies (Kullander, 2003), five of which refer to soft anatomy and the other four that correspond to very fragile structures, which hardly fossilize. Among them, the short hypapophysis on the fourth vertebra is the only one that was observed in †Plesioheros. In addition, the disjunct lateral line presented by †Plesioheros is typical of cichlids, being a relatively rare pattern among fishes (Webb, 1990).
Although molecular and combined analyses (e.g. Farias et al., 1999, 2000; Sparks & Smith, 2004; Chakrabarty, 2006; Smith et al., 2008) have recovered Neotropical cichlids as monophyletic, morphological evidence is slim. The strong interdigitating suture between the vomerine shaft and parasphenoid bar suggested by Stiassny (1991) seems to be the most constant synapomorphy for Cichlinae (Kullander, 1998; Smith et al., 2008). Unfortunately, the fossil preservation does not allow the observation of the vomerine-parasphenoid suture in †Plesioheros.
The Heroini synapomorphies listed by Kullander (1998) and Smith et al. (2008) were not preserved in †Plesioheros. However, the presence in †Plesioheros of anterior enlarged canines with a great size difference to the other teeth, is not found in any other Neotropical cichlid and was pointed by Kullander (1996) as a synapomorphy of Heroini. In addition, the presence of a lingual subapical cusp on outer anterior teeth in the oral jaws, as found in the fossil, was considered as a derived character of Heroini or of a subset within this group (Casciotta & Arratia, 1993b; Kullander, 1996; Chakrabarty, 2007). The occurrence of these two characters in †Plesioheros justifies its assignment to the Heroini cichlines.
In order to test this assignment and to try to establish fossil relationships, †Plesioheros was added to the Kullander's (1998) matrix and a phylogenetic analysis was performed. The analysis resulted in three MPT (L: 733, CI 0.20, RI 0.50), only differing in the arrangement of some geophagin genera. The strict consensus tree (Fig. 7) is largely similar to that proposed by Kullander (1998; using a weighted analysis), with the same major groupings, and even the same position for Heterochromis among the cichlines, but there are differences. The most evident involves part of the Crenicaratina (Biotoecus, Crenicara and Dicrossus) and Apistogramma nested within Cichlasomatini. These aspects of the cichlid tribal structures and generic arrangements not involving †Plesioheros are beyond the scope of this paper and will not be discussed here.
In our tree, the Heroini tribe has the same generic composition as the consensus tree of Kullander (1998: fig. 9), but the arrangement changes. In both trees, Hoplarchus is sister-group to all other heroins; however, in our tree Hypselecara turn out to be the second most basal heroin and Australoheros has a more derived position near other deep-bodied Heroini. †Plesioheros was positioned within Heroini as the sister-group to a clade formed by Australoheros + (Heroina + (Pterophyllum+ Symphysodon) + (Mesonauta + (Uaru + Heros))). The only support for this clade is the presence of lingual cusps on the anterior teeth (65:1).
The only autapomorphy found for †Plesioheros chauliodus in the analysis here performed was the presence of five lateral canal openings in the dentary (45:0). Five openings is the most common number in cichlids, occurring in geophagins and Old World cichlids. The reduction to four openings is a synapomorphy for the Cichlasomatini + Heroini (Kullander, 1998; our analysis). Among the heroins, only Thorichthys presents five openings in the dentary (Miller & Nelson, 1961; Kullander, 1983, 1996; Chakrabarty, 2007).
Plesioheros and the heroins
Although our analysis results corroborate the assignment of †Plesioheros to the Heroini, its position within the tribe and relationships to deep-bodied heroins (Fig. 7) must be considered with caution. The Heroini form a very diverse and large tribe distributed from middle South America to Texas and including the Greater Antilles. Although the tribe monophyly has been largely recognized (Cichocki, 1976; Kullander, 1998; Farias et al., 1998, 1999, 2000; Smith et al., 2008; López-Fernández et al., 2010; and others), the analyses have recovered South American heroin lineages nested within the Central American clade, demonstrating that these two assemblages are not monophyletic (Chakrabarty, 2007; ían & Kullander, 2006, 2008; ían et al., 2008; López-Fernández et al., 2010). The Kullander (1998) matrix does not contain any Middle American representatives, and three (Australoheros, "Cichlasoma" atromaculatum, Petenia) of the 11 South American genera have been found to be more related to Middle American taxa in recent analyses (ían & Kullander, 2006, 2008; Smith et al., 2008; López-Fernández et al., 2010). Additionally, the heroin taxonomy is deficient with some genera having never been defined and other many species are still waiting a generic assignment after they have been removed from the catch-all genus 'Cichlasoma' (Kullander, 1983). Therefore, the †Plesioheros allocation within the clade could possibly change if Middle American heroin lineages were included in the analysis.
Facing the large diversity and taxonomic intricacy of the modern forms, there is the limited and fragmentary information that is inherent to the fossil record. In the case of †Plesioheros, a mixture of primitive and derived characters is associated to the impossibility of verifying important characters referring to the caudal and anal fins. This fossil specimen here described is the only known representative of its taxon and the available characters are just those observable in an impression of the lateral view. Even with this limited information, it is possible to recognize this taxon as a new heroin genus. Hopefully, future fieldwork may provide more complete specimens displaying additional characters and synapomorphies, which will allow improve knowledge of †Plesioheros anatomy and relationships.
Plesioheros and the fossil cichlids
Cichlids have a notably poor fossil record considering their modern diversity. Among the few records of fossil cichlids, eight have been described for the Neotropics. Besides those characters used in the phylogenetic analysis, †Plesioheros can be distinguished from other American fossil cichlids by meristics and metric characters.
Cockerell (1924) described †Nandopsis woodringi in the genus Cichlasoma, based on a specimen from Las Cahobas (Haiti). Examining the fossil, Chakrabarty (2006) recognized lingual cusps on the oral teeth and 4 spines in the anal fin, whose combination is unique to Nandopsis species. Although the Haitian fossil is poorly preserved, it can be distinguished from †Plesioheros by the head profile (straight in †N. woodringi and rounded in †Plesioheros), the total number of the vertebrae (28-29 versus 23-24 in †Plesioheros), and the number of in the dorsal-fin spines (14 vs. 11 in †Plesioheros).
†Tremembichthys is from the Eocene-Oligocene (~ 34 Ma) of southeastern Brazil, and includes two species: †T. pauloensis and †T. garciae. †Tremembichthys can be distinguished it from †Plesioheros by the number of dorsal-fin spines (14-15 vs. 11 in †Plesioheros) and the total number of vertebrae (29-31 in †Tremembichthys vs. 23-24 in †Plesioheros).
†Aequidens saltensis and †Palaeocichla longirostrum are from the Miocene (~ 13 Ma) sediments of northwestern Argentina. †Aequidens saltensis can be differentiated from †Plesioheros by the total number of vertebrae (27-28 vs. 23-24 in †Plesioheros) and the number of dorsal-fin rays (XIII+13 vs. XI+12 in †Plesioheros). †Palaeocichla longirostrum can be distinguished from †Plesioheros by the number of pectoral-fin rays (16 vs. 12 in †Plesioheros), the total number of vertebrae (31-32 vs. 23-24 in †Plesioheros), the shape of the dorsal fin with the posterior spines decreasing in size, and by the length of the body and caudal peduncle (more elongated than †Plesioheros).
†Macracara prisca is from Brazilian northeastern sediments, which would have been deposited in an abandoned meander of the Parnaíba River during the Pliocene (~3.6 Ma; Dino et al., 2006). Macracara can be differentiated from †Plesioheros by the number of vertebrae (36 vs. 23-24 in †Plesioheros) and the number of dorsal-fin spines (16 vs. 11 in †Plesioheros).
The other two cichlids from the Lumbrera Formation, †Proterocara argentina and †Gymnogeophagus eocenicus, are from the same Faja Verde II level than †Plesioheros. Together they comprise the oldest American cichlid record and attest an ancient diversification in this family. In a recent analysis (Smith et al., 2008), †Proterocara was recovered in an apical position within Geophagini. Although the counting of dorsal-fin rays in †Proterocara (XI+12-13) coincides with that of †Plesioheros, these species can be differentiated by the body depth (41.3% SL vs. 47.8% SL in †Plesioheros), number of vertebrae (28 vs. 23-24 in †Plesioheros), and number of pectoral-fin rays (14 vs. 12 in †Plesioheros). The Lumbrera geophagin, †G. eocenicus, is distinguished by the number of dorsal-fin spines (14 vs. 11 in †Plesioheros), and by the Gymnogeophagus synapomorphies: lack of supraneurals associated with a forward-directed spine at the distal tip of the first dorsal pterygiophore.
Comparative material. Fossil specimens: Gymnogeophagus eocenicus, CNS-V10024; Proterocara argentina, CNS-V10020; Tremembichthys garciae, UNG 2T-148. Extant specimens: (All cleared and stained) Aequidens epae, MCP 15184; Australoheros facetus, MCP 23791; Crenicichla punctata, MCP 13187; Geophagus sp., MCP 10318; Gymnogeophagus meridionalis, MCP 14767.
Acknowledgements
This research was financially supported by CNPq grant (MCM) and scholarship (PAP); and PIP 781, PICT2006-381 grants (CdP). Prosanta Chakrabarty and Sven O. Kullander gave valuable comments for revising this manuscript. TNT was provided free by the Willi Hennig Society.
Literature Cited
Accepted June 18, 2010
Published September 24, 2010
- Bardack, D. 1961. New Tertiary teleosts from Argentina. American Museum Novitates, 2041: 1-27.
- Casciotta, J. & G. Arratia. 1993a. Jaws and teeth of American cichlids (Pisces: Labroidei). Journal of Morphology, 217: 1-36.
- Casciotta, J. & G. Arratia. 1993b. Tertiary cichlid fishes from Argentina and reassessment of the phylogeny of New World Cichlids (Perciformes: Labroidei). Kaupia, 2: 195-240.
- Chakrabarty, P. 2006. Taxonomic status of the Hispaniolan Cichlidae. Occasional papers of the Museum of Zoology, 737: 1-17.
- Chakrabarty, P. 2007. A morphological phylogenetic analysis of middle american cichlids with special emphasis on the section 'Nandopsis' sensu Regan. Miscellaneous Publications Museum of Zoology University of Michigan, 198: 1-31.
- Cichocki, F. P. 1976. Cladistic history of cichlid fishes and reproductive strategies of the american genera Acarichthys, Biotodoma and Geophagus Unpublished Ph.D. Dissertation, University of Michigan, Ann Arbor, 356p.
- Cockerell, T. D. A. 1924. A fossil cichlid fish from the Republic of Haiti. Proceedings of the United States National Museum, 63: 1-2.
- Concheiro Pérez, G. A., O. ían, G. Ortí, E. Bermingham, I. Doadrio & R. Zardoya. 2007. Phylogeny and biogeography of 91 species of heroine cichlids (Teleostei: Cichlidae) based on sequences of the cytochrome b gene. Molecular phylogenetics and Evolution, 43: 91-110.
- del Papa, C. E. 2006. Estratigrafia y paleoambientes de la Formación Lumbrera, Grupo Salta, noroeste argentino. Revista de la Asociación Geológica Argentina, 61: 313-327.
- del Papa, C., A. Kirschbaum, J. Powell, A. Brod, F. Hongn & M. Pimentel. 2010. Sedimentological, geochemical and paleontological insights applied to continental omission surfaces: a new approach for reconstructing Eocene foreland basin in NW Argentina. Journal of South American Earth Sciences, 29: 327-345.
- Dino, R., M. J. Garcia, L. Antonioli & M. R. Lima. 2006. Palinoflora das Camadas Nova Iorque, registro sedimentar do Plioceno na Bacia do Parnaíba (Maranhão). Pp. 42. In: Pearinotto, J. A., I. C. Lino, A. R. Saad, M. L. Etchebehere & N. Morales (Eds.). Boletim do VII Simpósio do Cretáceo do Brasil, UNESP.
- Eschmeyer, W. N. & J. D. Fong. 2010. Species of Fishes by family/subfamily. On-line version 22/ mar/ 2010. http://research.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp
- Farias, I. P., H. Schneider & I. Sampaio. 1998. Molecular phylogeny of Neotropical cichlids: the relatioships of cichlasomines and heroines. Pp. 499-508. In: Malabarba, L. R., R. E. Reis, R. P. Vari, Z. M. S. Lucena & C. A. S. Lucena (Eds.). Phylogeny and Classification of Neotropical Fishes. Porto Alegre, Edipucrs, 603p.
- Farias, I. P., G. Ortí & A. Meyer. 2000. Total evidence: Molecules, morphology, and the phylogenetics of cichlid fishes. Journal of Experimental Zoology, 288: 76-92.
- Farias, I. P., G. Ortí, I. Sampaio, H. Schneider & A. Meyer. 1999. Mitochondrial DNA phylogeny of the family Cichlidae: Monophyly and fast molecular evolution of the Neotropical Assemblege. Journal of Molecular Evolution, 48: 703-711.
- Gómez Omil, R. J., A. Boll & R. M. Hernández. 1989. Cuenca cretácico-terciaria del Noroeste argentino (Grupo Salta). Pp. 43-64. In: Chebli, G. A. & L. A. Spalletti (Eds.). Cuencas Sedimentarias Argentinas. Universidad Nacional de Tucumán, San Miguel de Tucumán, 512p.
- Kullander, S. O. 1983. Taxonomic studies on the percoid freshwater fish Family Cichlidae in South America. Stockolm, Swedish Museum of Natural History, 440p.
- Kullander, S. O. 1986. Cichlid fishes of the Amazon River drainage of Peru. Stockolm, Swedish Museum of Natural History, 431p.
- Kullander, S. O. 1996. Heroina isonycterina, a new genus and species of cichlid fish from Western Amazonia, with comments on cichlasomine systematics. Ichthyological Exploration of Freshwaters, 7(2): 149-172.
- Kullander, S. O. 1998. A phylogeny and classification of the South American Cichlids (Teleostei: Perciformes). Pp. 461-498. In: Malabarba, L. R., R. E. Reis, R. P. Vari, Z. M. S. Lucena & C. A. S. Lucena (Eds.). Phylogeny and Classification of Neotropical Fishes. Porto Alegre, Edipucrs, 603p.
- Kullander, S. O. 2003. Family Cichlidae (Cichlids). Pp. 605-654. In: Reis, R. E., S. O. Kullander & C. J. Ferraris, Jr. (Eds.). Check List of the Freshwater Fishes of South and Central America, Porto Alegre, Edipucrs, 729p.
- López-Fernández, H., K. O. Winemiller & R. L. Honeycutt. 2010. Multilocus phylogeny and rapid radiations in Neotropical cichlid fishes (Perciformes: Cichlidae: Cichlinae). Molecular Phylogenetics and Evolution, 55: 1070-1086.
- Malabarba, M. C. & L. R. Malabarba. 2008. A new cichlid Tremembichthys garciae (Actinopterygii, Perciformes) from the Eocene-Oligocene of eastern Brazil. Revista Brasileira de Paleontologia, 11(1): 59-68.
- Malabarba, M. C., L. R. Malabarba & C. del Papa. 2010. Gymnogeophagus eocenicus (Perciformes: Cichlidae), an Eocene cichlid from the Lumbrera Formation in Argentina. Journal of Vertebrate Paleontology, 30(2): 341-350.
- Malabarba, M. C., O. D. Zuleta & C. del Papa. 2006. Proterocara argentina, a new fossil cichlid from the Lumbrera Formation, Eocene of Argentina. Journal of Vertebrate Paleontology, 26: 267-275.
- Miller, R. R. & J. S. Nelson. 1961. Variation, life colors, and ecology of Cichlasoma callolepis, a cichlid fish from southern Mexico, with a discussion of the Thorichthys species group. Occasional papers of the Museum of Zoology, 622: 1-9.
- Ogg, J. G., G. Ogg & F. M. Gradstein. 2008. The concise Geologic Time Scale. New York, Cambridge University Press, 184p.
- Reis, R. E. & L. R. Malabarba. 1988. Revision of the Neotropical genus Gymnogeophagus Ribeiro, 1918, with descriptions of two new species (Pisces, Perciformes). Revista Brasileira de Zoologia, 4: 259-305.
- ían, O. & S. O. Kullander. 2006. Character- and tree-based delimitation of species in the 'Cichlasoma' facetum group (Teleostei, Cichlidae) with the description of a new genus. Journal of Zoological Systematics and Evolutionary Research, 44: 136-152.
- ían, O. & S. O. Kullander. 2008. The Australoheros (Teleostei: Cichlidae) species of the Uruguay and Paraná River drainages. Zootaxa, 1724: 1-51.
- ían, O., R. Zardoya & I. Doadrio. 2008. Phylogenetic relationships of Middle American cichlids (Cichlidae, Heroini) based on combined evidence from nuclear genes, mtDNA, and morphology. Molecular Phylogenetics and Evolution, 49: 941-957.
- Salfity, J. A. & R. A. Marquillas. 1994. Tectonic and Sedimentary Evolution of the Cretaceous-Eocene Salta Group Basin, Argentina. Pp. 266-315. In: Salfity, J. A. (Ed.). Cretaceous Tectonics of the Andes. Berlin, Friedrich Vieweg and Sohn, 345p.
- Schaeffer, B. 1947. Cretaceous and Tertiary actinopterygian fishes from Brazil. Bulletin of the American Museum of Natural History, 89(1): 1-38.
- Silva Santos, R. & H. S. Santos. 1993. Tremembichthys pauloensis (Schaeffer, 1947) (Pisces, Cichlidae) da Formação Tremembé, Estado de São Paulo, Brasil. Anais da Academia Brasileira de Ciências, 65: 41-55.
- Smith, W. L., P. Chakrabarty & J. S. Sparks. 2008. Phylogeny, taxonomy, and evolution of Neotropical cichlids (Teleostei: Cichlidae: Cichlinae). Cladistics, 24: 625-641.
- Sparks, J. S. & W. L. Smith. 2004. Phylogeny and biogeography of cichlid fishes (Teleostei: Perciformes: Cichlidae). Cladistics, 20: 501-517.
- Sparks J. S. & W. L. Smith. 2005. Freshwater fishes, dispersal ability, and nonevidence: "Gondwana Life Rafts" to the rescue. Systematic Biology, 54: 158-165.
- Stiassny, M. L. 1991. Phylogenetic intrarelationships of the family Cichlidae: an overview. Pp. 1-35. In: Keenleyside, M. H. A. (Ed.). Cichlid fishes. Behaviour, ecology and evolution. Chapman & Hall, 378p.
- Webb, J. F. 1990. Ontogeny and phylogeny of the trunk lateral line system in cichlid fishes. Journal of Zoology, 221: 405-418.
- White, T., C. del Papa & R. Brizuela. 2009. Paleosol-based paleoclimate reconstruction of Late Paleocene through Middle Eocene Argentina. Geological Society of America, Abstracts with Programs, 41(7): 567.
- Woodward, A. S. 1939. Tertiary fossil fishes from Maranhão, Brasil. Annals and magazine of Natural History, 11: 450-453.
Publication Dates
-
Publication in this collection
04 Nov 2010 -
Date of issue
2010
History
-
Received
18 June 2010 -
Accepted
14 Sept 2010