Abstracts
Otocinclus arnoldi from the La Plata basin is resurrected from the synonymy of O. flexilis described from the rio Jacuí drainage, based on three distinguishing features: the possession of five branched pectoral-fin rays, the larger number of enlarged odontodes on the tip of the parieto-supraoccipital posterior process, and having the prootic involved in the contact with the hyomandibular articular condyle. These species are also compared to O. mimulus, a third species described from the Paraná River basin, and the three species are rediagnosed. A reassessment of the phylogenetic relationships of all species of Otocinclus shows a well-supported clade composed of (O. xakriaba ((O. mimulus, O. arnoldi) (O. affinis, O. flexilis))) from the eastern-draining river basins of the Brazilian Shield as sister-group to a clade including all remaining Otocinclus species which are distributed on a wide lowland area of the Amazonas, Paraguay, and Orinoco basins.
La Plata; Synonymy; Hypoptopomatinae; Taxonomy; Catfish; Cascudinho
Otocinclus arnoldi da bacia do rio da Prata é revalidada da sinonímia de O. flexilis, descrito da bacia do rio Jacuí, baseado em três características distintivas: a presença de cinco raios ramificados na nadadeira peitoral, o maior número de odontódeos hipertrofiados na ponta do processo posterior do parieto-supraoccipital, e por ter o proótico envolvido no contato com o côndilo articular do hiomandibular. Essas espécies são também comparadas com O. mimulus, outra espécie descrita da bacia do rio Paraná, e as três espécies são re-diagnosticadas. Uma nova análise filogenética de todas as espécies de Otocinclus revelou um clado bem suportado composto por (O. xakriaba ((O. mimulus, O. arnoldi)(O. affinis, O. flexilis))) dos rios do escudo Brasileiro que drenam para leste, como grupo-irmão de um clado que inclui todas as demais espécies de Otocinclus que são distribuídas em uma grande área baixa das bacias do Amazonas, Paraguai e Orinoco.
Re-validation of Otocinclus arnoldi Regan and reappraisal of Otocinclus phylogeny (Siluriformes: Loricariidae)
Pablo Lehmann A.I; Fernanda MayerII; Roberto E. ReisII
ILaboratório de Ictiologia, Universidade do Vale do Rio dos Sinos. Av. Unisinos, 950; 93022-000 São Leopoldo, RS, Brazil. pablole@unisinos.br
IILaboratório de Sistemática de Vertebrados, Pontifícia Universidade Católica do Rio Grande do Sul. P. O. Box 1429, 90619-900 Porto Alegre, RS, Brazil. reis@pucrs.br
ABSTRACT
Otocinclus arnoldi from the La Plata basin is resurrected from the synonymy of O. flexilis described from the rio Jacuí drainage, based on three distinguishing features: the possession of five branched pectoral-fin rays, the larger number of enlarged odontodes on the tip of the parieto-supraoccipital posterior process, and having the prootic involved in the contact with the hyomandibular articular condyle. These species are also compared to O. mimulus, a third species described from the Paraná River basin, and the three species are rediagnosed. A reassessment of the phylogenetic relationships of all species of Otocinclus shows a well-supported clade composed of (O. xakriaba ((O. mimulus, O. arnoldi) (O. affinis, O. flexilis))) from the eastern-draining river basins of the Brazilian Shield as sister-group to a clade including all remaining Otocinclus species which are distributed on a wide lowland area of the Amazonas, Paraguay, and Orinoco basins.
Key words: La Plata, Synonymy, Hypoptopomatinae, Taxonomy, Catfish, Cascudinho.
RESUMO
Otocinclus arnoldi da bacia do rio da Prata é revalidada da sinonímia de O. flexilis, descrito da bacia do rio Jacuí, baseado em três características distintivas: a presença de cinco raios ramificados na nadadeira peitoral, o maior número de odontódeos hipertrofiados na ponta do processo posterior do parieto-supraoccipital, e por ter o proótico envolvido no contato com o côndilo articular do hiomandibular. Essas espécies são também comparadas com O. mimulus, outra espécie descrita da bacia do rio Paraná, e as três espécies são re-diagnosticadas. Uma nova análise filogenética de todas as espécies de Otocinclus revelou um clado bem suportado composto por (O. xakriaba ((O. mimulus, O. arnoldi)(O. affinis, O. flexilis))) dos rios do escudo Brasileiro que drenam para leste, como grupo-irmão de um clado que inclui todas as demais espécies de Otocinclus que são distribuídas em uma grande área baixa das bacias do Amazonas, Paraguai e Orinoco.
Introduction
Otocinclus flexilis Cope, 1894 was described from the rio Jacuí, Rio Grande do Sul State, Brazil, based on a syntype series with two lots and 17 specimens, collected by Herbert H. Smith in 1882 and deposited in the Academy of Natural Sciences of Philadelphia under catalog numbers ANSP 2162221626 and ANSP 21756-21767. Cope (1894: 97) diagnosed O. flexilis from O. affinis Steindachner, 1877, mentioning the presence of six branched pectoral-fin rays and describing the color pattern as ''light yellowish brown, with a row of about six oblong dusky spots along the lateral line, which become obscure anteriorly. A series of corresponding spots along the dorsal region. Dorsal and caudal fins light colored with numerous dusky spots. A black spot at the base of caudal fin in some specimens''.
In the same publication, Otocinclus fimbriatus Cope, 1894 was described from the same type locality of O. flexilis, based on differences ''... in the tubercular and fringed lip ... more brightly colored and with less numerous lateral spots. The ventral fins are relatively longer, and the dorsal fin originates above their base, and not behind it, as is the case in O. flexilis''. Schaefer (1997) examined the syntype series of O. fimbriatus (ANSP 21585-21597, 21752-21755, 17 specimens) and concluded that it does not present consistent differences from O. flexilis and, therefore, O. fimbriatus was synonymized with Otocinclus flexilis, as already suggested by Regan (1904) and Aquino (1996), and maintained by Schaefer (2003).
Otocinclus arnoldi Regan, 1909 was described from ''La Plata'' based on a single aquarium specimen donated by J. P. Arnold. The rather short original description does not state the number of pectoral-fin rays. Aquino (1996), based on a morphometric and meristic comparison, and also on the color pattern, synonymized O. arnoldi with O. flexilis. More recently, Otocinclus mimulus Axenrot & Kullander, 2003 was described from the río Paraná drainage in Paraguay, being diagnosed from O. flexilis by possessing elevated, enlarged odontodes at the posterior parieto-supraoccipital tip and distinct modal number of premaxillary and dentary teeth and caudal vertebrae.
Otocinclus is the only genus among the Hypoptopomatinae that received strong phylogenetic attention by previous authors. Schaefer (1997) produced the first phylogenetic analysis including all species of Otocinclus known to date. After that, Britto & Moreira (2002) described O. tapirape and reassessed the phylogenetic relationships among Otocinclus species. Axenrot & Kullander (2003) described O. mimulus and again reassessed the phylogeny of Otocinclus species, adding one character to the matrix of Schaefer (1997). Reis (2004) and Lehmann (2006) described O. cocama and O. batmani, but did not provide updated phylogenetic analyses of the genus.
In this paper we present the results of a reassessment of the validity of Otocinclus arnoldi, and reassess the phylogenetic relationships among all Otocinclus species.
Material and Methods
The specimens examined belong to the following institutions: Natural History Museum, London (BMNH); Instituto de Ciencias Naturales, Museo de Historia Natural, Universidad Nacional de Colombia, Bogotá (ICNMHN); Museo de Ciencias Naturales, Guanare (MCNG); Museu de Ciências e Tecnologia, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre (MCP); Museo Nacional de Historia Natural del Paraguay, Asunción (MNHNP); Museu de Zoologia da Universidade de São Paulo, São Paulo (MZUSP); Swedish Museum of Natural History, Stockholm (NRM); Universidade Federal do Rio Grande do Sul, Porto Alegre (UFRGS); Universidade Federal do Rio de Janeiro, Rio de Janeiro (UFRJ); and National Museum of Natural History, Smithsonian Institution, Washington (USNM). Measurements were calculated as interlandmark distances based on homologous landmarks acquired with a video digitizer, using the same set of landmarks as Schaefer (1997). An additional landmark was digitized at the end of the hypural plate and standard length was calculated as the interlandmark distance from this point to the snout tip. The software LMDis (by R. E. Reis, 1996) was used to extract interlandmark distances. Counts and anatomical terminology follow Schaefer (1997), Arratia (2003) and Axenrot & Kullander (2003). Specimens studied were cleared and stained (c&s) using the methods of Taylor & van Dyke (1985). Osteological terminology follows Schaefer (1997) and Arratia (2003).
Principal component analysis (PCA) was used to assess morphometric variation among studied Otocinclus populations. A total of 18 morphometric variables (Table 1) was taken from 70 specimens representing comparable size ranges: 30 specimens from the rio Jacuí basin, type-locality of Otocinclus flexilis (29.4-40.3 mm SL), 27 specimens from the rio Uruguai and lower río Paraná drainage, type-locality of O. arnoldi (26.2-44.4 mm SL), and 13 paratypes of O. mimulus from the middle río Paraná basin in Paraguay (30.9-36.3 mm SL). The analysis was performed on the covariance matrix of the 18 log10-transformed measurements.
For the phylogenetic analysis we used the original data matrix of Schaefer (1997) with the addition of Otocinclus arnoldi, O. tapirape, O. mimulus, O. cocama, O. batmani, and another undescribed species from the rio Madeira basin provisionally called Otocinclus sp. ''madeira''. We also included one character from Axenrot & Kullander (2003) on a mimetic association of Otocinclus with the callichthyid Corydoras, and six additional new characters. We submitted the data matrix to 10,000 replications of Random Addition Sequence (RAS) followed by TBR branch swapping using the software NONA (by P. Goloboff, 1993) and WinClada (by Nixon, 2002). Trees were rooted on Microlepidogaster perforatus. All multistate characters were set as unordered. Bremer branch support was calculated with NONA.
Results
A direct comparison of proportional measurements between specimens of Otocinclus flexilis from the laguna dos Patos basin and specimens from the rio Paraná and rio Uruguai drainages (Table 1) reveals no differences, as already demonstrated by Schaefer (1997). The principal component analysis also failed to reveal unambiguous differences among the three populations analyzed. The first principal component included a large proportion of the total variance (78.3%) and all variable loadings were negative and varied little in magnitude, indicating that it represents a general size factor. Plots of factor scores of principal components 2 versus 3 and 2 versus 4 both grouped specimens into three broadly overlapping clusters (Fig. 1). PC 2, 3, and 4 included 7.5, 3.3, and 2.4% of the total variance, respectively.
Lateral trunk coloration in these populations is also similar, as the three groups have either a row of 3-6 distinct dark blotches or a distinct dark stripe, extending from the compound pterotic to the caudal-fin base, or a diffuse mixture of those two color patterns (Figs. 2, 3 and 4). On the other hand, however, there is an important difference in the pectoral-fin ray counts among these fishes, as already demonstrated by Schaefer (1997). All examined specimens from the La Plata basin (285 specimens, including the holotype of O. arnoldi and the 13 paratypes of O. mimulus) have five branched rays in the pectoral fin (except for one specimen each in lots MCP 25254, UFRGS 7180, and USNM 176023 which have six branched rays in one side), and all 226 specimens examined from the laguna dos Patos basin (in addition to the entire syntype series of O. flexilis and O. fimbriatus examined by Schaefer, 1997) have six branched rays in the pectoral fin, except for one specimen in UFRGS 4963 and nine juveniles from MCP 15068, with five rays in one or both sides.
Schaefer (1997: 107) described a raised tuft of odontodes forming a raised crest on the parieto-supraoccipital tip as a juvenile character of hypoptopomatines, and regarded the presence of enlarged parieto-supraoccipital crest odontodes in mature adult specimens within Otocinclus as a derived condition representing paedomorphosis, and therefore synapomorphic for O. affinis and O. xakriaba. The same tuft of enlarged, raised odontodes is present in O. mimulus, which was coded as having the derived state in the analysis of Axenrot & Kullander (2003).
Contrary to the three species above, however, mature adult specimens of Otocinclus from the Jacuí and La Plata drainages do not have a raised crest of odontodes on the parieto-supraoccipital tip. However, most of the examined specimens present a patch of hypertrophied odontodes on the parieto-supraoccipital tip which are not raised to form a crest, but instead are laid on the bone surface and cannot be visualized laterally. These odontodes are 2-5 times larger than the surrounding odontodes and are present in both juveniles and adults. The population in the rio Jacuí basin has 0-10 (mode = 0, mean = 3.0) enlarged odontodes, while the fishes from the Uruguai and lower Paraná basins possess 2-23 (mode = 7, mean = 8.3) enlarged odontodes (Table 2).
The incorporation of the prootic in the contact with the hyomandibular articular condyle was described by Schaefer (1997: 104) as an autapomorphic trait of O. xakriaba. This feature, however, is also present in the Otocinclus populations from the La Plata basin, including O. mimulus, but not in O. flexilis from the rio Jacuí basin, in which only the compound pterotic contacts the hyomandibular articular condyle.
Based on the differences in pectoral-fin count, number of hypertrophied odontodes on the parieto-supraoccipital, and the involvement of the prootic in the contact with the hyomandibular articular condyle, O. arnoldi is resurrected from the synonymy of O. flexilis.
In a similar manner, a direct comparison of proportional measurements and meristics between Otocinclus arnoldi and a series of 13 paratypes of O. mimulus (Table 1) shows no differences, as already mentioned by Axenrot & Kullander (2003: 255). In the original description, O. mimulus was distinguished from O. arnoldi (O. flexilis of Axenrot & Kullander, 2003, who only examined specimens from the La Plata basin), based on the elevated tuft of enlarged odontodes at tip of parieto-supraoccipital. The tuft is distinctly conspicuous in O. mimulus because the predorsal plates immediately posterior to the parieto-supraoccipital are slightly sunk below the level of the parieto-supraoccipital border, making the tuft clearly apparent. Other meristic characters originally used to diagnose O. mimulus include modal number of premaxillary and dentary teeth, which are variable and largely overlapping, and caudal vertebrae, which suggest a distinction between the two species (Table 2). The PCA also failed to separate the two species, but showed a trend towards separation, especially on PC 4, and the two species are maintained and recognized as valid in this study.
Otocinclus flexilis Cope, 1894 Fig. 2
Otocinclus flexilis Cope, 1894: 97, pl. 8, fig. 13. Type locality: rio Jacuhy, Rio Grande do Sul [rio Jacuí, Brazil]. Lectotype: ANSP 21756, 40.0 mm SL, designated by Schaefer (1997: 53).
Otocinclus fimbriatus Cope, 1894: 98, pl. 9, fig. 16. Type locality: rio Jacuhy, Rio Grande do Sul [rio Jacuí, Brazil]. Syntypes: ANSP 212752 (4) and ANSP 21585-97 (14).
Diagnosis. Otocinclus flexilis is distinguished from all other Otocinclus (except for O. xakriaba, O. affinis, O. hasemani, O. arnoldi, and O. mimulus) by possessing an iris operculum and (except for O. xakriaba, O. arnoldi, and O. mimulus) by having a lateral trunk coloration consisting of a series of 3-6 diffuse pigment blotches (Fig. 2). An inconspicuous and not elevated patch of 0-10 (mode = 0, mean = 3.0) enlarged odontodes on the postero-dorsal parieto-supraoccipital tip, the pectoral skeleton with closed arrector fossae, and the presence of a triangular pigment mark at the dorsal-fin base distinguishes O. flexilis from O. xakriaba, which possesses an elevated patch of enlarged odontodes on the parietosupraoccipital, has an open arrector fossae, and lacks the dorsal-fin mark. From O. arnoldi and O. mimulus it is easily distinguished by having six branched pectoral-fin rays (vs. five branched rays) and few (0-10, mode = 0, mean = 3.0) not elevated enlarged odontodes on the parieto-supraoccipital tip (vs. 2-23, mode = 7, mean = 8.3) not elevated enlarged odontodes in O. arnoldi and 5-14, mode = 7, mean = 8.4 conspicuously elevated odontodes in O. mimulus).
Distribution. Otocinclus flexilis is restricted to the laguna dos Patos drainage basin (Fig. 5).
Otocinclus arnoldi Regan, 1909 Fig. 3
Otocinclus arnoldi Regan, 1909: 234. Type locality: La Plata, South America. Holotype: BMNH 1908.12.5:13, 43 mm SL.
Diagnosis. Otocinclus arnoldi is distinguished from all other Otocinclus species except O. mimulus, by having five branched rays in the pectoral-fin (vs. six branched pectoralfin rays), and except from O. mimulus and O. xakriaba by having the prootic involved in the contact with the hyomandibular articular condyle (vs. only compound pterotic contacting the hyomandibular articular condyle). It is distinguished from O. mimulus by having a not elevated patch of enlarged odontodes on the postero-dorsal parietosupraoccipital tip and 15 caudal vertebrae (vs. having an elevated patch of enlarged odontodes on the parietosupraoccipital and typically having 16-17 caudal vertebrae see Table 2). It is further distinguished (except for O. affinis, O. hasemani, O. xakriaba, O. mimulus and O. flexilis) by possessing an iris operculum, and from O. affinis, O. mimulus, and O. xakriaba by having a not elevated patch of enlarged odontodes on the postero-dorsal parieto-supraoccipital tip. It is also distinguished from all remaining Otocinclus species (except for O. xakriaba, O. flexilis, and O. mimulus) by having a lateral trunk coloration composed of either a row of 3-6 distinct dark blotches or a distinct dark stripe extending from the compound pterotic to the base of the caudal fin, or a diffuse mixture of those two color patterns (Fig. 3).
Distribution. Otocinclus arnoldi occurs in the tributaries of the lower río Paraná drainage, lower and middle rio Uruguai, and the río de La Plata (Fig. 5).
Otocinclus mimulus Axenrot & Kullander, 2003 Fig. 4
Otocinclus mimulus Axenrot & Kullander, 2003: 251. Type locality: Paraguay: Departamento Caaguazú: río Paraná drainage, small stream (arroyo) at Estancia María Belén, 8 km from Colonel Patricio Colman; 25º40'13''S 55º5'52''W. Holotype: MNHNP uncatalogued (ex NRM 43480), 34.1 mm SL [Holotype originally cited as MNHNP uncat.].
Diagnosis. Otocinclus mimulus is distinguished from all other Otocinclus species except O. arnoldi, by having five branched rays in the pectoral-fin (vs. six branched pectoral-fin rays), and except from O. arnoldi and O. xakriaba by having the prootic involved in the contact with the hyomandibular articular condyle (vs. only compound pterotic contacting the hyomandibular articular condyle). It is distinguished from O. arnoldi by having an elevated patch of enlarged odontodes on the postero-dorsal parieto-supraoccipital tip and typically having 16-17 caudal vertebrae (see Table 2; vs. having a not elevated patch of enlarged odontodes on the parietosupraoccipital and having 15 caudal vertebrae). It is further distinguished (except for O. affinis, O. hasemani, O. xakriaba, O. arnoldi, and O. flexilis) by possessing an iris operculum, and (except for O. affinis and O. xakriaba) by having an elevated patch of enlarged odontodes on the postero-dorsal parietosupraoccipital tip. It is also distinguished from all remaining Otocinclus species (except for O. xakriaba, O. arnoldi, and O. flexilis) by having a lateral trunk coloration composed of eitherfin, or a diffuse mixture of those two color patterns (Fig. 4).
Distribution. Otocinclus mimulus is only known from the río a row of 3-6 distinct dark blotches or a distinct dark stripe Mondai in Paraguay, a left-bank tributary of the lower rio extending from the compound pterotic to the base of the caudal Paraná (Fig. 5).

Phylogenetic relationships
The phylogenetic position of Otocinclus arnoldi, O. batmani, O. cocama and Otocinclus sp. ''madeira'' were never investigated. In order to test their phylogenetic position we added to the 27-character data matrix of Schaefer (1997) the character proposed by Axenrot & Kullander (2003) on a mimetic association with Corydoras, and six additional characters (Table 3), which are described below.
Character 28: Mimetic association with a species of Corydoras. According to Axenrot & Kullander (2003) ''Otocinclus mimulus, O. flexilis, O. affinis, and O. xakriaba are considered to be mimics of particular sympatric Corydoras species (C. diphyes, C. paleatus, C. nattereri, and C. garbei, respectively)''. This interpretation of mimetic association is based on the syntopically co-occurrence and on the shared color pattern of the Otocinclus-Corydoras species pair. We add to this list O. arnoldi, which has the same color pattern of O. flexilis and C. paleatus and also shares the syntopic co-occurrence with C. paleatus (nine of the 23 MCP lots of O. arnoldi were collected syntopically with C. paleatus).
Character 29: Position of pleural ribs. In loricariids the pleural ribs posterior to the well-developed rib of the sixth vertebral centrum are thin and delicate, and variably occur in the first centra posterior to the sixth centrum. In basal loricariids, most neoplecostomines, and most hypoptopomatines the first pleural rib posterior to the sixth centrum is associated with the seventh or eighth centrum (state 0). In Otocinclus bororo, O. mariae, O. affinis, O. caxarari, O. flexilis, O. arnoldi, O. xakriaba, O. mimulus, and Otocinclus sp. ''madeira'' the first delicate rib is associated to the ninth vertebral centrum (state 1). Contrastingly, in Hisonotus notatus, Microlepidogaster perforatus, O. hoppei, O. huaorani, O. macrospilus, O. vestitus, O. vittatus, O. tapirape, O. cocama, and O. batmani the first delicate rib is associated with the tenth vertebral centrum (state 2). Otocinclus mura and O. hasemani have no pleural ribs, and this character is thus not applicable. The character is highly variable among the genera of the Hypoptopoma group, but the tribe was coded as having state 0.
Character 30: Shape of the ventral process of the complex centrum (VPCC). In basal loricariids and most hypoptopomatines the VPCC has the shape of a rectangular arch attached to the ventral surface of the complex centrum and contacting the swimbladder capsule by means of a thin, dorsolaterally directed splint. Among species of Otocinclus this state is only shared by O. xakriaba (state 0). In all remaining species of Otocinclus the VPCC lost the dorsolaterally splint, being straight, curved or shaped as a golf-stick, but never forming a complete arch (state 1).
Character 31: Shape of the dorsal-fin spinelet. In basal loricariids the first dorsal-fin spine is transformed in a Vshaped spinelet, which acts with the nuchal plate as a locking mechanism for the second dorsal-fin spine (state 0). In the neoplecostomines and hypoptopomatines, however, the spinelet lost its function as a locking mechanism and is either oval or rectangular and plate-like in shape (state 1), a state shared with Microlepidogaster perforatus and Hisonotus notatus. In all species of Otocinclus, in contrast, the dorsalfin spinelet is V-shaped and the dorsal-fin spine locking mechanism is functional, which represents a reversion in Otocinclus. All species on the Hypoptopoma group completely lost the dorsal-fin spinelet (state 2).
Character 32: Number of branched pectoral-fin rays. Most hypoptopomatines, including the Hypoptopoma group, Microlepidogaster perforatus, Hisonotus notatus, and most species of Otocinclus share the possession of six branched rays in the pectoral fin (state 0). Contrastingly, O. mimulus and O. arnoldi posses five branched rays in the pectoral fin (state 1).
Character 33: Number of predorsal plates. The number of predorsal median plates between the parieto-supraoccipital tip and the nuchal plate is variable among the hypotopomatines. Microlepidogaster perforatus, Hisonotus notatus, Otocinclus xakriaba, O. flexilis, O. mimulus, and O. arnoldi have three or four predorsal plates (state 0). This condition is shared by the Hypoptopoma group, which can have three or more predorsal plates. In contrast, all the
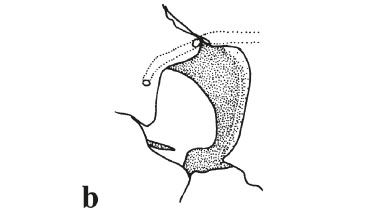
Fig. 3. Color variation of Otocinclus arnoldi. (a) and (b): MCP 25234, SL = 43.8 and 39.0 mm; (c): MCP 32565, SL = 31.9 mm; and (d): UFRGS 6838, SL = 30.4 mm. remaining species of Otocinclus share the presence of two Character 34: Teeth on pharyngeal jaws. predorsal plates between the parieto-supraoccipital and the Hypoptopomatines generally have well developed teeth nuchal plate (state 1). arranged in multiple series on both the upper pharyngeal tooth plate and the fifth ceratobranchial (state 0), a condition shared tooth on both the upper pharyngeal tooth plate and the fifth by Hisonotus notatus, Microlepidogaster perforatus and the ceratobranchial (state 1). Hypoptopoma group. All species of Otocinclus share a Besides adding the above characters, we provide modified reduction in the pharyngeal teeth, only having one series of interpretations of some of the characters as originally described by Schaefer (1997). His character 2 is an expansion of the subnasal lamina of the lateral ethmoid. Microlepidogaster perforatus and Hisonotus notatus share with O. affinis, O. flexilis, O. xakriaba, O. mimulus, and O. arnoldi the lateral ethmoid only slightly expanded medially, covering less than 20% of the nasal capsule when viewed through the nares (Fig. 6a, state 0). Contrastingly, in O. bororo, O. caxarari, O. hasemani, O. hoppei, O. huaorani, O. macrospilus, O. mariae, O. tapirape, O. vestitus, O. vittatus, O. cocama, O. batmani, and Otocinclus sp. ''madeira'' the subnasal lamina of the lateral ethmoid is moderately exposed and expanded medially in a concave shelf, covering 30-70% of the nasal capsule as viewed through the nares (Fig. 6b, state 1). In the Hypoptopoma group and in O. mura the lateral ethmoid is greatly expanded medially, covering 80-100% of the nasal capsule (Fig. 6c; state 2).

Schaefer's (1997) character 21 is the midlateral dark stripe that can be solid and continuous from the compound pterotic to the caudal-fin base (state 0) or broken in a series of three or more large, diffuse blotches of irregular size and shape (state 1). To this character we added a second state where the midlateral dark stripe is not confluent with the spot at caudalfin base, usually being interrupted one or two plates before the spot (state 2). This later condition is shared by Otocinclus mariae, O. hoppei, O. macrospilus, and Otocinclus sp. ''madeira''. Otocinclus cocama has a very distinct lateral color pattern, but was coded as having state 1.
Some other characters that deserve comments are: Character 15 is the possession of 23 or fewer lateral plates. Despite that O. cocama and the undescribed species from the rio Madeira possess 21-24 lateral plates, we coded these species as presenting state 1, because the count of 24 plates is rare. Character 18 refers to the number of canal-bearing plates in the anterior field of perforated lateral line plates. Because O. cocama has the lateral line continuous, without a mid-body gap, we coded this character as inapplicable. Finally, the character 22 describes the paired W-shaped marks of the caudal-fin pigmentation. As both O. cocama and O. batmani have one single, wide W-shaped mark on the caudal fin, we added the state 2 for this character to code the condition shared by these two species.
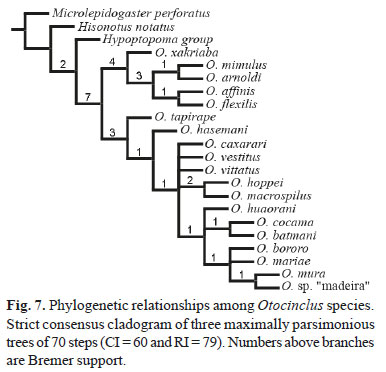
The phylogenetic analysis of this expanded data matrix yielded three maximally parsimonious trees with 70 steps (CI = 60 and RI = 79), the strict consensus of which is presented in Fig. 7.
Discussion
Aquino (1996) proposed that Otocinclus arnoldi is a junior synonym of Otocinclus flexilis based on overlapping results in morphometric and meristic characters, and the color pattern. However, she did not include specimens of O. flexilis from the rio Jacuí basin in the morphometric analysis, only comparing the holotype of O. arnoldi with 34 specimens (table 1, lots ILPLA 204 and 207) from the La Plata basin. Furthermore, in her table 1, the holotype of O. arnoldi is erroneously reported as having six pectoral-fin branched rays, what is contrary to our own and Schaefer's (1997: 53) count of five branched rays. The results presented by Aquino (1996), therefore, do not provide compelling justification for considering O. arnoldi as junior synonymy of O. flexilis.
Following the synonymy of Aquino (1996), Schaefer (1997: 53) reported on the number of pectoral-fin rays present in O. flexilis, indicating that this feature is variable in this species. The reason for such variability is that specimens from both the rio Jacuí and the La Plata basin were considered to be conspecific. As shown above, all 285 specimens examined of O. arnoldi have five branched rays in the pectoral fin, contrary to all other species of Otocinclus (except O. mimulus), which retain the plesiomorphic state of having six branched rays.
In his revision of Otocinclus Schaefer (1997) reported three lots of O. affinis from the La Plata basin in Argentina (USNM 176023 and USNM 177900) and rio Uruguai in southern Brazil (MCP 9388), significantly extending the geographical range of that species from the coastal rivers of Rio de Janeiro and São Paulo to the La Plata basin. We re-examined these three lots and all 60 specimens have five branched rays in the pectoral fins (except one specimen in USNM 176023 which has six rays in one side). The color pattern is rather faded, but the broad longitudinal stripe or series of blotches can be still seen. For this reason, these specimens are re-identified as O. arnoldi and O. affinis is consequently restricted to the coastal rivers of the Brazilian States of São Paulo and Rio de Janeiro.
The recognition of Otocinclus mimulus as a valid species separated from O. arnoldi is based on slight differences. We preferred this course of action based on the absence of dense sampling of Otocinclus in the area between Santa Fé in Argentina and the río Monday and other tributaries of the rio Paraná in Paraguay. Also, the restricted distribution of O. mimulus is not typical of Otocinclus species but that particular region was already detected as the area of endemism of the cichlid Gymnogeophagus setequedas. It is possible that the distribution of O. mimulus is wider than the río Mondaí alone, but like G. setequedas, it can be restricted to the tributaries of the lower rio Paraná in southeastern Paraguay and the Brazilian state of Paraná, between the río Mondaí and the Brazilian city of Guaíra.
Despite the addition of six morphological characters, the phylogenetic relationships uncovered by the present analysis did not gain much resolution when compared to the previous analyses (Schaefer, 1991; Britto & Moreira, 2002; andAxenrot & Kullander, 2003). Our results show a well-supported clade composed of (O. xakriaba ((O. mimulus, O. arnoldi)(O. affinis, O. flexilis))) as sister-group to all remaining Otocinclus species. Inside this clade, the most basal species Otocinclus xakriaba is endemic to the rio São Francisco basin. Otocinclus mimulus and O. arnoldi are endemic to the upper rio Paraná and lower rio Paraná/rio Uruguai basins, respectively. Finally, O. affinis and O. flexilis are endemic to coastal streams of Rio de Janeiro and São Paulo in southeastern Brazil and the laguna dos Patos coastal basin in southern Brazil. This clade evolved and differentiated on the watersheds draining the Brazilian Shield, and is the sister group to the clade comprising the species inhabiting the Paraguay, Amazonas and Orinoco basins.
The other Otocinclus clade in the present hypothesis is not well resolved, showing a large amount of polytomy and low internal branch support values. The two most basal species in that clade, however, O. tapirape and O. hasemani, are both endemic to the rio Tocantins, a river emptying near the mouth of the rio Amazonas but still running on rocks of the Brazilian Shield. The remaining of the species are distributed on a wide lowland area of the Amazonas, Paraguay, and Orinoco basins, suggesting that the genus Otocinclus began to differentiate on the watersheds draining the Brazilian Shield uplands and invaded lowlands of the continent only once.
Material examined. Acestridium discus: Brazil: Amazonas: MZUSP 85320, 7, 2 c&s, 26.8-56.6 mm SL, igarape Barroso, tributary to rio Preto da Eva on Francisco Mendes road, 2º44'N, 59º38'W. Hisonotus notatus: Brazil: Espírito Santo: MCP 18098, 208, 3 c&s, 31.1-41.0 mm SL, rio São José dos Torres on road BR-101, between São José dos Torres and Travessão, 21º4'43''S 41º14'2''W. Hypoptopoma guentheri: Brazil: Mato Grosso: MCP 15744, 2 c&s, 49.1-52.7 mm SL, flooded area of rio Paraguai, channel in the middle course, Cáceres, 16º3'S 57º42'W. Microlepidogaster perforatus: Brazil: Minas Gerais: MCP 17717, 1, 1c&s, 31.9-33.7 mm SL, rio Carandaí, tributary of rio das Velhas at Carandaí, upper rio Paraná basin, 20º57'17''S, 43º46'41''W. Nannoptopoma spectabilis: Venezuela: MCNG 26579, 2c&s, 22.2-23.7 mm SL, Orinoco basin. Niobichthys ferrarisi: Venezuela: Amazonas State: MCP 34810, 2, 1 c&s, 39.1 mm SL, Municipio Autónomo Río Negro, río Baria, approximately 200 m above Neblina base camp. 0º55'N, 66º10'W. Otocinclus affinis: Brazil: Rio de Janeiro: UFRJ 4660, 2 c&s, 28.8 30.7 mm SL, rio Preto, tributary to rio Piabanha, São José do Rio Preto, 22º10'S 42º55'W. Otocinclus arnoldi (285 specimens): Argentina: Río de La Plata: BMNH 1908.12.5:13, holotype, 43 mm SL, río de La Plata. Lower Paraná drainage: MCP 32565, 5, 1 c&s, 27.3-34.3 mm SL, arroyo Nogoya, Vitória/Gualeguay, 32º50'S 59º50'W. MCP 11591, 10, 27.3-37.1 mm SL, río Zapata on Ruta 11, Puerto Magdalena, 35º05'S 57º30'W. MZUSP 51092, 18, 24.5-34.9 mm SL, río Cayasta, tributary to río San Javier, Helvecia, 31º10'S 60º10'W. USNM 175023, 6, 25.1-35.2 mm SL, Buenos Aires. USNM 177900, 4, 27.0-33.6 mm SL, Santa Fé. Río Uruguay drainage: Entre Rios: MCP 43131, 7, 28.4-32.2 mm SL, canal on isla Pelada on río Uruguay, Colonia Hugues, 32º21'S 58º09'W. Brazil: Rio Grande do Sul: rio Uruguai drainage: MCP 9271, 1, 30.6 mm SL, creek on road from Santa Maria to Mata, 29º33'S 54º27'W. MCP 9388, 50, 14.7 31.1 mm SL, rio Ibicui on road from São Rafael to Cacequi, 29º49'S 54º41'W. MCP 9644, 21, 26.7-34.9 mm SL, rio Santa Maria on km 246 of road BR 293 between Santana do Livramento and Dom Pedrito, 30º59'S 54º42'W. MCP 10590, 1, 30.31 mm SL, creek at Faculdade de Zootecnia, Uruguaiana, 29º45'S 57º05'W. MCP 11563, 1, 32.2 mm SL, arroio Touro Passo, Uruguaiana, 29º38'S 56º56'W. MCP 14165, 1, 26.1 mm SL, arroio SantoAntonio on road from Rosário do Sul to Santana do Livramento, 30º18'S 54º59'W. MCP 14196, 1, 34.3 mm SL, Rosário do Sul, 30º15'S 54º55'W. MCP 16189, 7, 24.5 27.6 mm SL, marginal lagoon on praia Formosa, São Marcos, Uruguaiana, 29º30'15''S 56º50'37''W. MCP 21626, 1, 33.4 mm SL, rio Uruguai and lateral pools at praia da Formosa, São Marcos, Uruguaiana, 29º29'54''S 56º49'12''W. MCP 23086, 1, 31.7 mm SL, rio Jaguari-Mirim, ca 5 km SE of São Francisco de Assis, 29º36'03''S 55º05'08''W. MCP 23110, 2, 33.5-33.7 mm SL, rio Inhacundá near São Francisco de Assis, 29º32'27''S 55º07'45''W. MCP 23153, 8, 30.2-39.3 mm SL, and MCP 25210, 5, 30.5-39.6 mm SL, rio Inhacundá near São Francisco do Assis, 29º32'51''S 55º08'11''W. MCP 23881, 1, 39.4 mm SL, arroio Ibirocai at Passo do Vai, Alegrete, 29º24'S 56º37'W. MCP 25234, 8, 2 c&s, 32.7-44.3 mm SL, rio Inhacundá near São Francisco de Assis, 29º32'27''S 55º07'45''W. MCP 25245, 46, 1 c&s, 30.1-42.7 mm SL, stream tributary to rio Inhacundá, São Francisco de Assis, 29º32'39''S 55º07'50''W. MCP 26766, 1, 28.1 mm SL, arroio Ibicuí da Faxina on road BR 158, Santana do Livramento, 30º47'31''S 55º12'35''W. MCP 26814, 2, 22.1-22.9 mm SL, rio Ibicuí between São Vicente do Sul and Cacequi, 29º50'22''S 54º47'53''W. MCP 26833, 1, 26.3 mm SL, creek tributary to rio Santa Maria, Rosário do Sul, 30º10'44''S 54º51'22''W. MCP 26864, 8, 2 c&s, 20.7-31.9 mm SL, arroio do Salso, Rosário do Sul, 30º22'27''S 55º02'07''W. MCP 26970, 2, 24.5-37.1 mm SL, rio Caxambu, Panambi, 28º35'50''S 53º27'31''W. MCP 27658, 1, 29.0 mm SL, arroio Caraí-Passo on road from São Francisco de Assis to Manoel Viana, 29º31'03''S 55º10'49''W. MCP 34613, 20, 19.9-24.5 mm SL, rio Ibicuí, São Vicente do Sul, 29º48'S 54º58'W. MCP 43818, 1, 29.2 mm SL, arroio Quarai-Chico at the Parque Estadual do Espinilho, Quarai, 30º47'S 57º28'W. UFRGS 5297, 1, 29.2 mm SL, arroio do Salso at Rosário do Sul, 30º22'27''S 55º21'00''W. UFRGS 5336, 4, 21.6-31.6 mm SL, arroio do Salso, Rosário do Sul, 30º22'27''S 55º21'00''W. UFRGS 5366, 1, 33.0 mm SL, rio Ibicuí-Mirim on road from Cacequi to São Vicente, Cacequi, 29º50'14''S 54º47'53''W. UFRGS 6838, 4, 31.7-38.2 mm SL, stream on road BR-290 ca 10 km from Rosário do Sul, 30º12'42.8''S 55º03'17.5''W. UFRGS 8339, 1, 43.5 mm SL, arroio do Salso, tributary to rio Santa Maria on road BR 293 between Bagé and Dom Pedrito. Uruguay: Río Uruguay drainage: MCP 10003, 5, 29.9-36.5 mm SL, río Negro, Arreria, Cerro Largo, 31º50'S 54º28'W. UFRGS 7180, 17, 31.5-47.7 mm SL, tributary to río Yi, Durazno, 33º15'S 56º00'W. UFRGS 7181, 1, 47.8 mm SL, arroyo Corrales on route 27, Tacuarembó, 31º23'S 55º12'W. UFRGS 7182, 2, 40.4-45.5 mm SL, arroyo Cuñapiru on route 27, Tacuarembó, 31º05'S 55º25'W. UFRGS 9244, 1, 19.8 mm SL, arroyo Batovi at km 24 of route 27, río Tacuarembó basin, Rivera, 31º06'58''S 55º24'56''W. Otocinclus bororo: Brazil: Mato Grosso: MCP 15721, 5 paratypes, 2 c&s, 19.1-26.6 mm SL, creek on road from Barra do Bugres to Cáceres, 15º45'S 57º20'W. Otocinclus caxarari: Brazil: Mato Grosso: MCP 19286, 7 paratypes, 2 c&s, 21.4-25.7 mm SL, tributary to rio Guaporé, Guajará-Mirim, 10º48'S 65º23'W. Otocinclus cocama: Peru: Loreto: MCP 34842, 8 paratypes, 2 c&s, 29.7-40.7 mm SL, Quebrada Yanayacu, Jenaro Herrera, 04º53'55''S 73º39'00''W. Otocinclus flexilis (226 specimens): Brazil: Rio Grande do Sul, laguna dos Patos drainage: MCP 9628, 3, 22.1-23.1 mm SL, arroio dos Ratos, Arroio dos Ratos, 30º07'S 51º43'W. MCP 15068, 145, 9.7-46.1 mm SL, swamp of rio Gravatai near road RS 118, ca. 500m from highway BR 290, Gravataí, 29º58'S 50º56'W. MCP 17414, 13, 2 c&s, 23.5-45.1 mm SL, arroio Itaetá, Passo das Pedras, 31º50'S 52º43'W. MCP 18307, 2, 37.6-37.8 mm SL, arroio Sapucaia, Esteio, 29º52'S 51º09'W. MCP 21426, 2, 28.4-33.9 mm SL, arroio Arambaré, Pedro Osório, 31º54'3 1''S 53º01'44''W. MCP 25131, 1, 37.5 mm SL, arroio Arambaré, Pedro Osório, 31º54'35''S 53º01'40''W. MCP 37681, 11, 38.5-44.4 mm SL, arroio Arambaré, Pedro Osório, 31º51'51''S 52º49'24''W. UFRGS 2011, 2, 29.4-31.7 mm SL, creek between highway BR 290 and Santo Antônio, Santo Antônio, 29º52'S 50º35'W. UFRGS 2013, 3, 32.4-34.1 mm SL, arroio Ramos between highway BR 290 and Santo Antônio, Santo Antônio 29º52'S 50º30'W. UFRGS 4963, 22, 23.5-31.0 mm SL, flood of rio Gravataí, Porto Alegre, 29º57'30''S 50º59'42''W. UFRGS 6325, 11 of 171, 33.3 41.5 mm SL, creek near Gravataí, 29º57'26''S 51º00'23''W. UFRGS 7144, 9, 33.5-43.4 mm SL, rio Gravataí at the Passo das Canoas, Gravataí, 29º57'46''S 51º0'7''W. Otocinclus hasemani: Brazil: Piauí: MCP 22547, 94, 21.0-27.9 mm SL, riacho Palo, Formosa, 05º14'27''S 42º40'19''W. Otocinclus hoppei: Brazil: Pará: MCP 22545, 26, 23.8 32.1 mm SL, igararé Apeú, tributary to rio Guamá, Castanhal, 01º18'06''S 47º59'11''W. Otocinclus huaorani: Peru: Loreto: NRM 17994, 13, 21.8-29.0 mm SL, and NRM 37375, 3 c&s, 18.0-29.3 mm SL, quebrada at km 53, San Jacinto, 02º31'S 75º43'W .Otocinclus macrospilus: Colombia: Amazonas, Río Amazonas drainage: ICNMHN 4155, 5, 27.3-37.3 mm SL, and ICNMHN 5213, 4, 23.6 31.2 mm SL, Quebrada Yahuaracaca, km 8, Letícia, 04º08'05''S 69º56'32''W. ICNMHN 5030, 1, 30.3 mm SL, río Puré, Leticia, 02º07'05''S 69º37'50''W. Peru: Loreto, Iquitos, Río Amazonas drainage: MCP 28205, 1, 23.3 mm SL, quebrada Pintuyacu, 43 km SW of Iquitos on road to Nauta, 04º5'58''S 73º27'18''W. Otocinclus mimulus: Paraguay: NRM 42332, 13 paratypes, 30.1-36.9 mm SL, and NRM 43479, 1 paratype c&s, 36.2 mm SL, small stream at Estancia María Belén 8 km from Coronel Patricio Colman, Caaguazu, 25º40'13''S 55º05'52''W. Otocinclus mura: Brazil: Pará: MCP 22550, 19, 21.2-32.7 mm SL, igarapé Urucuré, Tomé-Acú, 02º29'13''S 48º31'31''W.Otocinclus tapirape: Brazil: Goiás: UFRJ 5421, 2 c&s, 20.1-20.3 mm SL, córrego Água Parada, 11 km W of Novo Planalto, 13º17'S 49º62'W. Otocinclus vestitus: Colombia: ICNMHN 4981, 1, 23.6 mm SL, caño La Arenosa, 10 km S on road Leticia to Tarapacá, 04º8'S 69º56'W. Otocinclus vittatus: Colombia: ICNMHN 1316, 1, 20.6 mm SL, laguna de Menegua, Puerto Lopez, 04º06'S 72º54'W. Venezuela: MCNG 15667, 16, 9.1-25.8 mm SL, caño Maraca, on road Guanare to Guanarito, 08º50'N 69º21'W. Bolivia: CBF 3069, 4, 21.9-27.5 mm SL, 1 c&s, 26.8 mm SL, arroyo Aguas Negras, río Curiraba, río Apere, río Mamoré, drenaje río Madeira, 10º22'52''S 65º22'47''W.Otocinclus xakriaba (30 specimens): Brazil: Minas Gerais: rio São Francisco drainage: MCP 16879, 25, 4 c&s, 22.5-28.7 mm SL, rio Peru-Açu, Januária, 15º11'S 44º12'30''W. MCP 23506, 1 c&s, 30.2 mm SL, rio Paraopeba, Jatuaba, 19º57'S 44º18'W. Otocinclus sp. ''madeira'' (49 specimens): Brazil: Amazonas: rio Madeira drainage: MCP 35594, 34, 4 c&s, 19.8-27.6 mm SL, igarapé do Doze on Transamazônica Road ca. 12 km W of Humaitá, 07º34'25''S 63º06'39''W. MCP 35595, 15, 2 c&s, 22.7-28.2 mm SL, igarapé do Vinte e Dois at Recanto do Sanari, ca. 22 km W of Humaitá, 07º35'36''S 63º10'27''W.
Acknowledgements
We thank Sven Kullander (NRM) for the loan of paratypes of Otocinclus mimulus, James Maclaine (BMNH) for the photos of the holotype of O. arnoldi and assistance during a visit to the NHM. We are grateful to Luiz R. Malabarba for support in fieldwork in Uruguay and loan of material from UFRGS. Thanks to Fernando Jerep and Vivianne Sant'Anna for help with specimens at USNM and photographs of specimens, to Aloisio Braun for the photographs in Figs. 2-4, and to Tiago Carvalho for assistance. Sven Kullander read different versions of the manuscript and presented valuable suggestions. Research associated with this paper was partially financed by the All Catfish Species Inventory (NSF-DEB #0315963). PLA received a doctoral fellowship from CNPq/ CAPES (process # 190033/02-9), FM had an IC fellowship from CNPq (process # 502643/2007-2), and RER is partially financed by CNPq (process # 303362/2007-3).
Literature Cited
Accepted November 30, 2009
Published March 31, 2010
- Aquino, A. E. 1996. Redescripción de Otocinclus flexilis Cope, 1894 (Siluriformes, Loricariidae, Hypoptopomatinae) con un nuevo sinónimo. Iheringia, Série Zoologia, 81: 13-22.
- Arratia, G. 2003. Catfish head skeleton - an overview. Pp. 3-46. In. Arratia, G., B. G. Kapoor, M. Chardon & R. Diogo (Eds.). Catfishes, v1. Enfield, Science Publishers.
- Axenrot, T. E. & S. O. Kullander. 2003. Corydoras diphyes (Siluriformes: Callichthyidae) and Otocinclus mimulus (Siluriformes: Loricariidae), two new species of catfishes from Paraguay, a case of mimetic association. Ichthyological Exploration of Freshwaters, 14(3): 249-272.
- Britto, M. R & C. R. Moreira. 2002. Otocinclus tapirape: A new hypoptopomatine catfish from central Brazil (Siluriformes: Loricariidae). Copeia, 2002: 1063-1069.
- Lehmann A, P. 2006. Otocinclus batmani: A new species of hypoptopomatine catfish (Siluriformes: Loricariidae) from Colombia and Peru. Neotropical Ichthyology, 4: 379-383.
- Regan, C. T. 1904. A monograph of the fishes of the family Loricariidae. Transactions of the Zoological Society of London, 18: 191-350.
- Regan, C. T. 1909. Descriptions of three new freshwater fishes from South America presented to the British Museum by Herr J. Paul Arnold. Annals and Magazine of Natural History, 3: 234-235.
- Reis, R. E. 2004. Otocinclus cocama, a new uniquely colored loricariid catfish from Peru (Teleostei: Siluriformes), with comments on the impact of taxonomic revisions to the discovery of new taxa. Neotropical Ichthyology, 2: 109-115.
- Schaefer, S. A. 1997. The neotropical cascudinhos: Systematics and biogeography of the Otocinclus catfishes (Siluriformes: Loricariidae). Proceedings of the Academy of Natural Sciences of Philadelphia, 148: 1-120.
- Schaefer, S. A. 2003. Subfamily Hypoptopomatinae. Pp. 321-229. In: Reis, R. E., S. O. Kullander & C. J. Ferraris, Jr. (Eds). Check list of the freshwater fishes of South and Central America. Porto Alegre, Edipucrs, 729p.
- Taylor, W. R. & G. G. van Dyke. 1985. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium, 9: 107-119.
Publication Dates
-
Publication in this collection
30 Apr 2010 -
Date of issue
Mar 2010
History
-
Received
30 Nov 2009 -
Accepted
31 Mar 2010