Abstracts
Atrioventricular valve complex of 30 Jafarabadi water buffaloes, adult males were studied in this research with no heart diseases. The animals were obtained from a slaughterhouse in Brazilian State of Parana. The hearts were opened at the third portion affording access to the valve complex. The complexes had its area, number and type of tendinous cords submitted to analysis. The results showed that the complex is composed by two cusps and four accessory cusps, two or three papillary muscles in which 10-25 tendinous cords fix on the cusps that face the ventricle wall. The total area of the complex was on average 38.56cm², with a minimum of 24.96cm² and a maximum of 55.54cm². Statistically, no relation between the number of cords and the cusps' area where they are inserted or with the number of papillary muscle where they originated from was observed.
Heart; Jaffarabadi water buffaloes; cardiac ventricle; mitral valve
Foram estudados os complexos valvares atrioventricular esquerdo de 30 búfalos da raça Jafarabadi, machos e adultos, sem alterações cardíacas, provenientes de abatedouros do Estado do Paraná. Os corações foram examinados em seu terço médio para acesso ao complexo valvar, que foi submetido a estudos de área, número e tipificação de cordas tendíneas. Os resultados demonstram que este complexo é formado por duas cúspides principais e quatro cúspides acessórias, apresentam em sua formação de 2-3 músculos papilares, nos quais se inserem de 10-25 cordas tendíneas, que se fixam em cúspides voltadas para a parede do ventrículo. A área total deste complexo apresenta uma média de 38,56cm² com um mínimo de 24,96cm² e um máximo de 55,54cm². Estatisticamente não há relação entre número de cordas e a área da cúspide onde estas estão inseridas, nem com o número de músculos papilares dos quais elas provem.
Coração; búfalo Jafarabadi; ventrículo cardíaco; valva mitral
Study of the cardiac left atrioventricular valvar complex in water buffaloes (Bubalus bubalis) of the Jafarabadi breed
Estudo do complexo valvar atrioventricular cardíaco esquerdo em búfalos (Bubalus bubalis) da raça Jafarabadi
Patrícia R. Esteves; Karla P.C. Araújo; Carlos E. Ambrósio* * Autor para correspondência: ceambrosio@usp.b r ; Dulcinéa G. Teixeira; Daniele S. Martins; Adriana C. Morini; João C. Morini Junior; Caio N. Duarte; Pedro P. Bombonato; Maria A. Miglino; Arani N.B. Mariana
Departamento de Cirurgia, Setor de Anatomia dos Animais Domésticos e Silvestres, Faculdade de Medicina Veterinária e Zootecnia (FMVZ), Universidade de São Paulo (USP), Av. Prof. Dr. Orlando Marques de Paiva 87, Cidade Universitária, São Paulo, SP 05508-900, Brazil
ABSTRACT
Atrioventricular valve complex of 30 Jafarabadi water buffaloes, adult males were studied in this research with no heart diseases. The animals were obtained from a slaughterhouse in Brazilian State of Parana. The hearts were opened at the third portion affording access to the valve complex. The complexes had its area, number and type of tendinous cords submitted to analysis. The results showed that the complex is composed by two cusps and four accessory cusps, two or three papillary muscles in which 10-25 tendinous cords fix on the cusps that face the ventricle wall. The total area of the complex was on average 38.56cm2, with a minimum of 24.96cm2 and a maximum of 55.54cm2. Statistically, no relation between the number of cords and the cusps' area where they are inserted or with the number of papillary muscle where they originated from was observed.
Index terms: Heart, Jaffarabadi water buffaloes, cardiac ventricle, mitral valve.
RESUMO
Foram estudados os complexos valvares atrioventricular esquerdo de 30 búfalos da raça Jafarabadi, machos e adultos, sem alterações cardíacas, provenientes de abatedouros do Estado do Paraná. Os corações foram examinados em seu terço médio para acesso ao complexo valvar, que foi submetido a estudos de área, número e tipificação de cordas tendíneas. Os resultados demonstram que este complexo é formado por duas cúspides principais e quatro cúspides acessórias, apresentam em sua formação de 2-3 músculos papilares, nos quais se inserem de 10-25 cordas tendíneas, que se fixam em cúspides voltadas para a parede do ventrículo. A área total deste complexo apresenta uma média de 38,56cm2 com um mínimo de 24,96cm2 e um máximo de 55,54cm2. Estatisticamente não há relação entre número de cordas e a área da cúspide onde estas estão inseridas, nem com o número de músculos papilares dos quais elas provem.
Termos de indexação: Coração, búfalo Jafarabadi, ventrículo cardíaco, valva mitral.
INTRODUCTION
This species breed is growing so fast in the World and mainly in Brazil, who looks for an important protein source in this resistant ruminant specie. Few articles on the literature are showing morphological aspect of Buffaloes, like placenta morphology and physiology (Pereira et al. 2001, 2009, Morini et al. 2008), mammary gland morphology (Santos et al. 2007), or even congenital malformation of the body (Schild et al. 2002). Also some pathology directed link to this specie in Brazil fields, as described by Silva et al. (1998) about botulism as a common disease in buffaloes rose in low lands of Maranhão State. However, this research is focusing on cardiac apparatus of this species, showing basic morphology of all heart.
Scientific literature consulted showing the disposition of the cardiac valve complex in mammals were explored although those researches did not specifically refer to the water buffaloes. Supply morphological bases and interpre-tations for experimental researches contributed in a significant manner to the development of the Cardiology. The subject has merited ever more detailed studies, whether for their morphological significance or for their importance to Comparative Anatomy as well as for Experimental Surgery.
The complexity of the various components of the atrioventricular valves has brought out scientific interest in detailing these structures and presenting a suitable understanding of the cardiac and circulation functions in water buffaloes.
Zimmerl (1949) states that the heart is made of two halves: the right heart and the left heart. Each portion has two superimposed cavities, atrium and ventricle, that communicate though broad openings, the atrioventricular ostium, provided with the mitral valve that regulates and guides the bloodstream from the atrium to the ventricle.
Mammals' left atrioventricular ostium (Ostium atrioventricularis sinister) establishes a communication between the two compartments of the arterial heart presents a circular or oval form and is formed by a valve made of two elements called bicuspids (Bossi et al.1909). The cusps of this valve on the left side are bigger and thicker than those on the right side of the heart (Bourdelle & Bressou 1938, Sisson & Grossman 1959, Dobberstein & Hoffnann 1964).
The bicuspid valve or mitral valve is similar to an Episcopal miter, presenting one auricular face, one ventricular face, one base and one free margin. Due to its location, the mitral cusps are designated as cranial and caudal. The first is smaller and is in the transition of the two orifices (atrioventricular and aortic), limiting a kind of infundibulum where the primitive aorta arise. The caudal cusp is larger and results of the union of three cusps which become one at its base. Between the two valves there is an accessory valve seeming a third cusp (Bossi et al. 1909, Gonzalez y Garcia & Gonzalez Alvarez 1949, Schwarze & Schroder 1972).
The tendinous cords (chordae tendineae) may be classified according to their insertion into the valve's cusps in three orders: first order are those inserted into the free margin of the cusps, except in the central portion of the aortic cusps and walls; are more numerous than the second order cords, these being more tapered (Bossi et. al. 1909, Davison 1947, Brock 1952, Chierchi et. al. 1956, Lam et. al. 1970). The third order cords, according to Ellenberger & Baum (1977), are named accessories and originate from the fleshy column or directly from the ventricle wall, are inserted into the base of the cuspid and fuse with the atrioventricular fibrous ring. Montagna (1964) states that the cardiac muscle, when contracts, tenses the papillary muscles and the tendinous cords.
As said by Getty (1981) and Schummer et al. (1981) the septal cusp separates the atrioventricular ostium from the aortic vestibule; the parietal, which in turn is situated in the papillary muscles of the left ventricle, called subauricularis and their tendinous cords are directed to the bicuspid valve.
In this study we chose water buffaloes from the Jafarabadi breed because they are a large sized species and of undisputable zootechnics/veterinary interest. We propose to contribute with the study of the water buffaloes ' Anatomy and Comparative Veterinary, studying in a detailed manner, the area of the septal, parietal and accessorial cuspids, the number of tendinous cords that reach each one of these in particular, the classifications as to the type of tendinous cords and the left atrioventricular valve as a whole.
MATERIALS AND METHODS
Thirty hearts of water buffaloes (Bubalus bubalis Linnaeus, 1758) of the Jafarabadi breed, adult males, obtained at slaughter-houses of the Brazilian Paraná State were used.
After removing the lungs and heart and checking the integrity of the organ, the heart with the vases of the base was isolated and submitted to running water to remove sanguine clots from their cameras. Soon afterwards the pieces were identified with labels, fastened in formol 10% and maintained submerged in this solution, in appropriate containers, for a period of 72 hours. The dissection began with the total removal of the atrial walls and partially of the wall of the left ventricle, to allow a first analysis in situ of the anatomical elements of the atrioventricular valve complex, the papillary muscles, the tendinous strings and cuspids of that ventricle. Subsequently, a discerning withdrawal of the valve group was promoted, maintaining the relationships among the above described components that make possible a detailed analysis of its anatomy. To facilitate the visualization, analysis and measurement of different anatomical components of the valve complex, after the removal of the set a cut in the medium third of the septal cusp was chosen, perpendicularly to the margins. The set was divided into two parts to allow the opening of the fibrous ring that sustains it. This procedure allows the prolongation and a rectified exhibition of all the cusps. Precise measurements through Images Analyzing System (MINIMOP) to evaluate the area of the different cusps and the total area of the atrioventricular valve were performed. The description of the tendinous cord types was made according to Machado; Peduti Neto (1991) who typify them for swine of the Landbreed breed.
Statistical analyses
For descriptive statistics the UNIVARIATE procedure of the Statistical Analysis System, version 6.12 (SAS 1995) program, was used. Entirely casualized experimental delineation to compare the different accessory areas and the areas of the parietal and septal cusps was used. Each comparative group (CG) represented one treatment. The ANOVA procedure (SAS 1991) was used in agreement with the following statistical model:
Yij = m + gci + eij
Where:
Yij = observed value of the j-th measured heart of i-th comparative group;
m = overall average;
gci = effect of i-th comparative group;
eij = random error inherent to the observation yij.
To investigate the relationship amongst the measures of the different areas Pearson analysis, product-moment correlation, was used by means of the PROC CORR.
RESULTS
After the examination of the left atrioventricular valve complex of 30 water buffaloes of the Jafarabadi breed, we could observe the presence of four accessory cusps in 100% of the cases; being denominated as subauricular or accessory cusps 1 and 2 those that turn back to the left wall of the ventricle, and subatrial or accessory cusps 3 and 4 those that turn back to the right wall of the left ventricle.
For better understanding of our discoveries they are presented separately: first regarding the data of the areas of the septal and parietal cusps, the accessory cusps and the total area of the left atrioventricular valve, all in square centimeters; second concerning the data of the total number of cords, number of cords per papillary muscle and the types of tendinous strings.
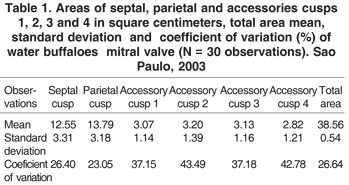
Values of the mean, average and Standard deviation as well the coefficient of variation of the septal cusps, parietal cusps, accessory cusps 1, 2, 3 and 4 in water buffaloes hearts can be seen in Table 1 and Figure 1 (A,B).
The variance analysis of the areas between the parietal and the septal cusps did not show significant results (P>0.05) that can suggest possible comparative differences among the areas of the groups.
Pearson correlation analyses (hereafter called correlation/s), were accomplished of product-moment, seeking to identify possible associations among specific blocks of variables. The correlation among the areas of the septal cusps, parietal and accessory cusps 1, 2, 3 and 4 was estimated in order to verify possible associations among those structures. We found "positive and low intensity correlations" among the areas of the septal cusps, parietal and accessory cusps 1, 2, 3 and 4.
The correlations of the subatrial muscle with the septal, parietal and accessory cusps 1, 2, 3 and 4 resulted in "positive and low intensity correlations" among the papillae subatrial muscle with the septal cusps, papillae subatrial muscle with the accessory cusp 1 and the subatrial muscle with the parietal cuspid.
"Positive and average intensity correlations" among the papillary subatrial muscle and the accessory cusp 4 and "negative correlations of low intensity" among the papillary subatrial muscle and the accessory cusps 2 and 3 has been observed.
The analysis of the correlations among the sub-auricular muscle and the septal, parietal and accessory cusps 1, 2, 3 and 4 exhibited "positive and low intensity correl-ations" among the papillary sub-auricular muscle and the accessory cusp 4. "Negative and low intensity correlations" among the papillary subauricular muscle and the septal, parietal and accessory cusps 1, 2 and 3 has been observed.
We found "positive and low intensity correlations" among the total number of tendinous cords and the area of the septal cusps, as well as with the area of the accessory cusp 1. "Positive correlations of average intensity" were observed among the total number of cords and the accessory cusp 4 as well "negative correlations of low intensity" among the total number of cords with the parietal and accessory cusps 2 and 3. Regarding the analysis of the correlations among the total number of cords with the papillary subatrial, subauricular and accessory muscles "positive and high intensity correlation" has been observed among the total number of cords and the subatrial and subauricular muscles. "Negative and low intensity correlation" was seen among the total number of cords and the papillary accessory muscle.
About the components of the left atrioventricular valve, the number of tendinous cords which origin is in the subauricular papillary muscle and that reach the septal, parietal and accessory cusps 1 and 2; and the number of tendinous cords that arise from the subatrial papillary muscle and reach the septal, parietal and accessory cusps 3 and 4 are expressed in Table 2. The total number of tendinous cords and number of cords by papillae muscle of the water buffaloes left atrioventricular valve are expressed in Table 3.
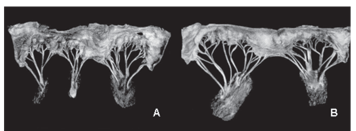
The type of tendinous cords that arise from the subauricular papillary muscle and reach the septal, parietal and accessory cusps 1 and 2 are presented in Table 4; those that arise from the subatrial papillary muscle and reach the septal, parietal and accessory cusps 3 and 4 are presented in Table 5. The types of tendinous cords can be seen in Figure 2 (A,B,C,D,E). In Figure 2 (F,G) the exemples of representative scheme of the water buffaloes's left atrioventricular valves of studied hearts are shown.
DISCUSSION
Regarding the description of the valve complex of domestic animals, the authors do not disagree much as to their form.
Referring the mammals Bossi et al. (1909), Martin (1915), Lesbrè (1923), Davison (1947), Dobberstein & Hoffmann (1964), Montagna (1964) state that the left atrioventricular orifice is provided with a valve made of two elements (bicuspid or mitral), whose free margins are cranially and caudally found (Ellenberger & Baum 1977, Getty 1981) being the one smaller and in the transition of two orifices (auriculoventricular and aortic) and the other larger caudal that results from the union of three elements (Gonzalez y Garcia & Gonzalez Alvarez 1949) which fuse in its base, agreeing with the description made by Montané &Bourdelle (1913) for equines and Bourdelle & Bressou (1938) for human beings.
Montané & Bourdelle (1913) confirm that the internal conformation of the heart in ruminants is similar to the solipeds both in the walls and in the orifices.
According to Zimmerl (1930), Zimmerl et al. (1930) and Bradley (1922) the bicuspid valve is to be found around the atrioventricular ostium, with an anterior cuspid and another posterior, both large, quadrilateral, deeper and more consistent than on the right side. However Sisson & Grossman (1959), Koch (1965) and Schwarze & Schroder (1972) limit themselves to commenting that these valves have larger and deeper cuspids than the right side of the heart. Dyce et al. (1990) stated that in ruminants the left atrioventricular valve is compatible with the right side valve but has only two larger cuspids.
Testut & Latarjet (1979) and Schummer et al. (1981) describe that, due to the two cusps location they are named septal and parietal, the septal cuspid is extensive and separates the aorta's ostium from the atrioventricular ostium, therefore the data found in the present research are that four accessory cusps have been identified in all samples, besides two main cusps, and their location and behavior are similar to that described by these authors. Regarding specific studies in equines Bourdelle & Bressou (1938) and Miller & Holmes (1984) works are evidenced since the authors stated that the left atrioventricular valve is formed by two larger cusps, anterior and posterior, in the entrance of the atrium to the ventricle. Between these cusps are the lesser or commissural cusps (left and right), frequently described as part of the posterior cusp. In the same way as the authors cited, we also found the valve complex to be constituted by two main cusps and four accessory cusps. However no author mentions the sizes of the different valves, what impede us to compare their data with our findings. We observed a septal cuspid with an average size of 12.55cm2, the parietal with 13.59cm2; the accessory cusp 1 with 1.14cm2; the accessory cusp 2 with 3.20cm2; the accessory cusp 3 with 3.13cm2; and the accessory cusp 4 with 2.82cm2. In relation to the whole area we could observe that the valve complex achieves an average value of 38.56cm2.
However, no author makes mention to the areas of the different valves, what in he disables them to compare its data with our findings
Regarding the number of accessory cusps, Bossi et al. (1909) indicated the presence of an accessory valve that appears like a third cuspid between the two main valves. Montané & Bourdelle (1913), Lesbré (1923) and Bourdelle & Bressou (1938) stated that there are 2 to 3 secondary leaflets interleaved between the main ones. £ukaszewska-Otto (1967) observed that the number of cusps varied from 2 to 7 and that the additional cusps were situated close to the commisures.
After a comparative study between the equine and bovine species Riella (1996) states that the number of accessory valves varied for each of the studied species. Lesnau (2001) verified a minimum of two accessory cusps for each heart. On the other hand Martin (1915), Montané & Bourdelle (1917), Bradley (1922), Zimmerl (1930), Bruni & Zimmerl (1951), Sisson & Grossman (1959), Montagna (1964), Getty (1981), Soares & Ferreira (1984), Dyce et al. (1990), Machado & Peduti Neto (1991) and Costa (1999) did not mention the number of accessory valves in their works. In our findings, regarding the number of accessory cusps, in all cases we found 4 accessory cusps that we call accessory cusps 1 and 2 turned towards the left wall of the ventricle, and those turned towards the right wall of the ventricle, accessory cusps 3 and 4, different from the consulted literature.
In respect to the number of tendinous cords, Dobber-stein & Hoffmann (1964), Koch (1965) and Ellenberger & Baunm (1977) mention the presence of 6-8 tendinous cords in both the papillary muscles (subauricular and subatrial). In specific works, £ukaszewska-Otto (1966) studying the Rhesus and Cynomolgus monkeys, describes that the average number of tendinous cords found is 18.70. In 1967 this same author, states that in swine the number of tendinous cords that are oriented towards the septal cusp is 55.75 on average, and 99.73 to the parietal cusp on average. Riella (1996) while studying 30 equine hearts and 30 bovine hearts observed that the number of tendinous cords varied from 7 to 15 for equines and from 8 to 15 for bovines. Other consulted authors as Bossi et al. (1909), Zimmerl (1930, 1949), Sisson & Grosmann (1959), Getty (1981) and Dyce (1990) did not refer to the number of tendinous cords. Since none of the authors worked specifically with water buffaloes, in regard to tendinous cords, we cannot establish comparisons.
Our study sought to establish the numbers of these structures, that originating in the subauricular and subatrial papillary muscles were destined to the septal, parietal cusps and the accessories cups 1, 2, 3 and 4. Therefore, it was verified that from the subauricular papillary muscle to the septal cusp there were a minimum of 1 and a maximum of 2 tendinous cords, and from the same muscle to the parietal cusp we found from 0 to 5 tendinous cords. Still from the subauricular muscle to the accessory cusp 1, tendinous cords varied from 1 to 3. We observed that there was a minimum of 0 to a maximum of 3 tendinous cords for the accessory cusp 2. Referent to the sub-atrial papillary muscle, the following number of tendinous cords originating were: to the septal cusp, a minimum of 1 and maximum of 2 cords; directed to the parietal cusp, the minimum of 0 and maximum of 6 cords; in accessory cusp 3, the cords varied from 1 to 4 and accessory cusp 4, they presented a minimum of one and a maximum of two cords. As to the total number of tendinous cords present in the left atrioventricular valve we found a minimum of 10 and a maximum of 25 cords with an average of 13 cords.
Rusted et al. (1951) and Frater & Ellis (1961) affirm that all the species of mammals examined by them have similar mitral valves and that the standard of the tendinous cords of the dogs' heart corresponds closely to that of the human heart.
Regarding the classification of the tendinous cords, Brock (1952), Chierchi et al. (1956), Lam et al. (1970) and Costa et al. (1999) state that the first order cords, those inserted into the free margin of the cusps, in all their parts, except in the central portions of the aortic and mural cusps, are more numerous than the second order cords and described as those that flow all over the ventricular face of the cusp by its sticky margin. In part of their trajectory in relation to the cusp, some remain free from adherence and others are to be found intimately united, in whole or in part to this membrane.
Machado & Peduti Neto (1991) classified four types of tendinous cords, which we adopt to classify the water buffaloes of Jafarabadi breed cords, thus: type I cords are those generally thicker arising from the papillary muscle and that reach the cusps through branches, starting from the cuspid margin, passing by the wrinkled area, up to the limit between this area and the smooth area of the ventricular face; type II cords are those terminating in slender branches, that project to the wrinkled area and/or margin of the cuspid to which they are destined; type III cords are very slender, represented by a single filament and exclusively destined to the cuspid margin; and, type IV cords are that which like the others arises from the papillary muscle, however is distributed only in the smooth area of the ventricular face of the cuspid, but the distance between its insertion point and the atrioventricular fibrous ring may vary. Although the types of the tendinous cords have been the same as those found in this research. The frequency in water buffaloes is different from that cited by the author for swine of the Landbreed breed, suggesting that number variation being of this animal species. That is why a comparative anatomic study is necessary.
Lesnau (2001) is the only author who makes reference to the existence of an accessory papillary muscle that was also found by us in one of the observations.
In agreement with the results obtained in our investigations about the left atrioventricular valve complex, we concluded that possesses a total area of 38.56cm2, on average a minimum of 24.96cm2 and a maximum of 55.54cm2, presenting in its formation a bicuspid valve, fastened by two or three papillae muscles in which on average insert 13.1 tendinous cords, with a minimum of 10 and a maximum of 25, that are fastened to the cusps turned to the ventricular wall.
We still concluded that after several statistical analyses none relationship among the number of cords and the area of the cusps at the insertion place, as well with the number of papillary muscles from which the cusps arise has been observed.
Reveived on February 15, 2009
Accepted for publication on July 3, 2009.
- Bossi V., Caradonna G.B., Spampani G., Varaldi L. & Zimmerl U. 1909. Tratado de Anatomia Veterinária. Vol.2. Francisco Vallardi, Milano, p.21-24.
- Bourdelle E. & Bressou C. 1938. Anatomie Regionale dês Animaux Domestiques. 9ième ed. J.B. Balliére, Paris. 538p.
- Bradley O.C. 1922. Tórax e Abdômen de Eqüino. W. Green, Edinburgh. 35p.
- Brock R.C. 1952. Pathologic and cirurgical anatomy of mitral valve. Brit. Heart J. 14:489-498.
- Bruni A.C. & Zimmerl U. 1951. Anatomia degli Animali Domestici. Francesco Vallardi, Milano, p.293-294.
- Chierchi M.A., Lees W.M. & Thompson R. 1956. Functional anatomy of the normal mitral valve. J. Thoracic Surg. 32:378.
- Costa F.S., Appolinário A.V.M., Morais-Pinto L. & Oliveira A. 1999. Um tipo especial de Chordae tendinae no ventrículo direito de cães sem raça definida (Canis familiaris Linnaeus, 1758). Braz. J. Morphol. Sci. 16:217-218.
- Davison A. 1947. Mammalian Anatomy. Blakiston, Philadelphia, p.169-170.
- Dobberstein J. & Hoffman G. 1964. Lehrbuch der vergleichenden Anatomie der Haustiere. Vol.3. S. Hirzel, Leipzig, p.13.
- Dyce K.M., Sack W.O. & Wensing C.J.G. 1990. Tratado de Anatomia Veterinária. Guanabara Koogan, Rio de Janeiro, p.343-437.
- Ellenberger W. & Baum H. 1977. Handbuch der vergleichenden Anatomie der Haustiere. Springer-Verlag, Berlin. 624 p.
- Frater R.W.M. & Ellis Jr. F.H. 1961. The anatomy of mitral valve canine. J. Surg. Res. 1:171.
- Getty R. 1981. Anatomia dos Animais Domésticos. Vol.1. 5Ş ed. Interamericana, Rio de Janeiro, p.171.
- Gonzalez y Garcia J. & Gonzalez Alvarez R. 1949. Anatomia Comparada de los Animales Domesticos. 6Ş ed. Canales, Madrid. 708p.
- Koch T. 1965. Lehrbuch der Veterinär-Anatomie. Vol.3. Gustav Fisher, Jena, p.27-28.
- Lam J.H.C., Ranganathan N., Wigle E.D. & Silver M.D. 1970. Morphology of the human mitral valve. I. Chordae tendinae: A new classification. Circulation 41:449-458.
- Lesbré F.X. 1923. Prècis d'Anatomie Comparée dês Animaux Domestiques.Tome 2. Baillière, Paris, p.264.
- Lesnau G.G. 2001. Anatomia do complexo valvar atrioventricular cardíaco esquerdo de baleias Minke (Balaenoptera acutorostrata Lacépéde, 1804). 168p. Dissertação de Mestrado em Anatomia, Faculdade de Medicina Veterinária e Zootecnia, USP, São Paulo.
- Machado G.V. & Peduti Neto J. 1991. Sobre o comportamento anatômico das chordae e tendinae, em relação à valva atrioventricular esquerda, em suínos da raça Landbreed. Braz. J. Vet. Res. Anim. Sci. 28:31-37.
- Martin P. 1915. Lehrbuch der Anatomie der Haustiere. Vol.2. Schickhardt & Ebner, Stuttgart, p.130-131.
- Miller P.J. & Holmes J.R. 1984. Observations on structure and function of the equine mitral valve. Equine Vet. J. 16:457-460.
- Montagna W. 1964. Anatomia Comparada. Omega, Barcelona. 204p.
- Montané L. & Bourdelle E. 1913. Anatomie Régionale dês Animaux Domestiques. Bailliére, Paris. 770 p.
- Montané L. & Bourdelle E. 1917. Anatomie Régionale dês Animaux Domestiques. Bailliére, Paris, p.242-243.
- Morini A.C., Barbosa P.L.G., Melnic R.V., Martins D.S., Morini-Junior J.C., Pereira F.T.V., Franciolli A.L.R., Favaron P.O., Ambrósio C.E. & Miglino M.A. 2008. Caracterização das membranas fetais em búfalas no terço inicial da gestação. Pesq. Vet. Bras. 28:437-445.
- Pereira F.T., Braga F.C., Burioli K.C., Kfoury Jr J.R., Oliveira L., Papa P., Carvalho A.F., Ambrósio C.E., Bazer F.W. & Miglino M.A. 2009. transplacental transfer of iron in the water buffalo (Bubalus bubalis): Uteroferrin and erythrophagocytosis. Reprod. Domest. Anim. 2. (Ahead of print)
- Pereira F.T.V., Miglino M.A., Bevilacqua E. & Carvalho A.F. 2001. Aspectos morfológicos dos hematomas placentários da placenta do búfalo (Bubalus bubalis bubalis Linnaeus, 1758). Braz. J. Vet. Res. Anim. Sci. 38:151-154.
- Riella A.C.M. 1996. Aspectos anatômicos das válvulas componentes da valva atrioventricular esquerda em eqüinos e bovinos sem raça definida. Dissertação de Mestrado em Anatomia, Faculdade de Medicina Veterinária e Zootecnia, USP, São Paulo. 111p.
- Rusted I.E., Scheifley C.H., Edwards J.E. & Kirklin J.W. 1951. Guides to the coissures in operations upon the mitral valve. Proc. Staff Meetings of the Mayo Clinic 26:297-305.
- Santos D.A.S., Ricci R.E.G., Prado I.M.M., Carvalho A.F., Bombonato P.P., Ambrósio C.E., Teixeira D.G., Santos T.C., H.-Brazquez F.J., Martins D.S., Morini A.C., Kfoury Jr J.R. & Miglino M.A. 2007. Morphology and morphometry of buffalo mammary papillae. Pesq.Vet. Bras. 27:95-102.
- Schild A.L., Soares M.P., Damé M.C., Portianski E.L. & Riet-Correa F. 2002. Arthrogryposis in Murrah buffaloes in southern Brazil. Pesq. Vet. Bras. 23:13-16
- Schummer A., Wilkens H., Vollmerhaus B. & Habermehl K.H. 1981. The circulatory system, the skin, on the cutaneous organ of the domestic mammals, p.32-33. In: Nickel L., Schummer A. & Seifferle E. (Eds), Anatomy of the Domestic Animals. Vol.3. Paul Parey, Berlin.
- Schwarze E. & Schroder L. 1972. Compêndio de Anatomia Veterinária. Vol.3. Acribia, Zaragoza, p.21-22.
- Silva T.M.D., Dutra I.S., Castro R.N. & Döbereiner J. 1998. Occurrence and distribution of Clostridium botulinum type C and D spores in buffalo breeding areas of the Baixada Maranhense, Maranhão, Brazil. Pesq. Vet. Bras. 18:127-131.
- Sisson S. & Grossman J.D. 1959. Anatomia de los Animals Domesticos. 4Ş ed. Salvat, Barcelona, p.505-506.
- Soares J.C. & Ferreira A.L. 1984. A valva atrioventricular esquerda: correlação entre área das cúspides e número de cordas tendíneas. Braz. J. Morphol. Sci. 1:28-31.
- Testut L. & Latarjet. A. 1979. Tratado de Anatomia Humana. Vol.2. 9Ş ed. Salvat, Barcelona, p.28-33.
- Zimmerl U. 1930. Trattato di Anatomia Veterinária. Vol.2. Francesco Vallardi, Milano, p.11.
- Zimmerl U. 1949. Trattato di Anatomia Veterinária. Vol.1. Francesco Vallardi, Milano, p.279.
- Zimmerl U., Bruni A.C., Caradonna G.B., Mannu A. & Preziuzo L. 1930. Trattato di Anatomia Veterinária. Vol.2. Francesco Vallardi, Milano, p.8-12.
Publication Dates
-
Publication in this collection
20 Jan 2010 -
Date of issue
Oct 2009
History
-
Accepted
03 July 2009 -
Received
15 Feb 2009