Resumos
Este trabalho teve como objetivo estudar a mobilização de reservas de sementes de Caesalpinia peltophoroides Benth. durante a germinação e crescimento inicial das plântulas. As variações nas reservas de carbiodratos, lipídios e proteínas foram analisadas desde o período pré-germinativo (0 a 5 dias após a semeadura - DAS) até a total senescência e abscisão dos cotilédones, aos 35 DAS, por meio de testes bioquímicos nos cotilédones das sementes. Os resultados indicaram que os lipídios constituem o principal composto de reserva nos cotilédones, contribuindo com cerca de 50% de massa seca. Carboidratos solúveis representaram 32%, as proteínas solúveis 7,7% e o amido 6,8% de massa seca dos cotilédones. Os lipídios sofreram marcante decréscimo entre 5 e 10 dias após a semeadura, período em que se observou elevada taxa de crescimento das plântulas. Carboidratos e proteínas solúveis exibiram tendência gradativa de queda, enquanto no amido, isso quase não foi detectado. A redução do peso de massa seca dos cotilédones foi bem correlacionada com o aumento da biomassa da plântula.
Carboidratos; lipídio e proteínas
This work aimed at studying the mobilization seed reserves during germination and initial growth of seedlings of Caesalpinia peltophoroides Benth. The variations in carbohydrate, lipid and protein reserves were analyzed from the pre-sprouting period (0 to 5 days after sowing -DAS) to the total senescence and abscission of seeds. The results showed that lipids constitute the main reserve compound in the cotyledon, contributing with almost 50 % of its dry mass weight. Soluble carbohydrates represent 32 %, the soluble proteins 7.7 % and starch 6.8 % of the dry mass weight of cotyledons. Lipids showed a marked decrease between 5 and 10 days after sowing, period of a high seedling growth rate. Carbohydrates and soluble proteins showed a gradual tendency to decrease, while starch was almost non-detectable. The reduction in cotyledon dry mass of weight was well correlated with the increase in the seedling biomass.
Carbohydrates; lipids and proteins
Mobilização de reservas durante a germinação das sementes e crescimento das plântulas de Caesalpinia peltophoroides Benth. (Leguminosae-Caesalpinoideae)
Mobilization of the reserves during germination of seeds and growth of seedlings of Caesalpinia peltophoroides Benth. (Leguminosae-Caesalpinoideae)
Viviana Borges CorteI; Eduardo Euclydes de Lima e BorgesII; Cláudia Aparecida PontesI; Idalina Tereza de Almeida LeiteIII; Marília Contin VentrellaIV; Alexsandro de Almeida MathiasV
IPrograma de Pós-Graduação em Ciência Florestal da Universidade Federal de Viçosa. E-mail:<viviborgescorte@yahoo.com.br>
IIDepartamento de Engenharia Florestal da Universidade Federal de Viçosa. E-mail: <elborges@ufv.bv>
IIIDepartamento de Ciências Biológicas da Universidade Federal do Espírito Santo
IVDepartamento de Biologia da Universidade Federal de Viçosa
VBiólogo
RESUMO
Este trabalho teve como objetivo estudar a mobilização de reservas de sementes de Caesalpinia peltophoroides Benth. durante a germinação e crescimento inicial das plântulas. As variações nas reservas de carbiodratos, lipídios e proteínas foram analisadas desde o período pré-germinativo (0 a 5 dias após a semeadura DAS) até a total senescência e abscisão dos cotilédones, aos 35 DAS, por meio de testes bioquímicos nos cotilédones das sementes. Os resultados indicaram que os lipídios constituem o principal composto de reserva nos cotilédones, contribuindo com cerca de 50% de massa seca. Carboidratos solúveis representaram 32%, as proteínas solúveis 7,7% e o amido 6,8% de massa seca dos cotilédones. Os lipídios sofreram marcante decréscimo entre 5 e 10 dias após a semeadura, período em que se observou elevada taxa de crescimento das plântulas. Carboidratos e proteínas solúveis exibiram tendência gradativa de queda, enquanto no amido, isso quase não foi detectado. A redução do peso de massa seca dos cotilédones foi bem correlacionada com o aumento da biomassa da plântula.
Palavras-chave: Carboidratos, lipídio e proteínas.
ABSTRACT
This work aimed at studying the mobilization seed reserves during germination and initial growth of seedlings of Caesalpinia peltophoroides Benth. The variations in carbohydrate, lipid and protein reserves were analyzed from the pre-sprouting period (0 to 5 days after sowing -DAS) to the total senescence and abscission of seeds. The results showed that lipids constitute the main reserve compound in the cotyledon, contributing with almost 50 % of its dry mass weight. Soluble carbohydrates represent 32 %, the soluble proteins 7.7 % and starch 6.8 % of the dry mass weight of cotyledons. Lipids showed a marked decrease between 5 and 10 days after sowing, period of a high seedling growth rate. Carbohydrates and soluble proteins showed a gradual tendency to decrease, while starch was almost non-detectable. The reduction in cotyledon dry mass of weight was well correlated with the increase in the seedling biomass.
Keywords: Carbohydrates, lipids and proteins.
1. INTRODUÇÃO
O estabelecimento de dada espécie está ligado à capacidade de suas sementes germinarem rápida e uniformemente, a fim de vencer a concorrência com outras espécies presentes no local, ou pela capacidade de se manterem viáveis por períodos mais longos até que condições ambientais sejam propícias ao desenvolvimento das plântulas (BORGES, 2003). Para tanto, cada espécie lança mão de adaptações, como a mobilização de determinadas reservas que serão úteis para a formação de estruturas físicas, como parede celular, ou para respiração, sintetizando intermediários metabólicos com finalidades diversas (BORGES e RENA, 1993; BORGES, 2003; BUCKERIDGE et al., 2004).
As principais substâncias de reserva nas sementes são carboidratos, proteínas e lipídios. A proporção dessa composição pode variar de espécie para espécie e até entre espécies de uma mesma família (BORGES e RENA, 1993; BEWLEY e BLACK, 1994). Essas substâncias são mobilizadas durante a germinação, e no decorrer do desenvolvimento das plântulas seus produtos de degradação são usados para diferentes propósitos, como a geração de energia e a produção de matéria-prima para a construção de novas células e tecidos (MAYER e POLJAKOFF-MAYBER, 1975).
As sementes têm sido estudadas quanto à composição química de suas reservas, e tal interesse não se dá apenas por seu teor nutritivo, mas por serem úteis na confecção de produtos industrializados (BUCKERIDGE et al., 2004), entre diversos fins. Além disso, o estudo da composição química é do interesse prático da tecnologia de sementes, porque tanto o vigor quanto o potencial de armazenamento de sementes são influenciados pelo teor dos compostos presentes (CARVALHO e NAKAGAWA, 2000).
Diante da possibilidade de ampliar o conhecimento sobre aspectos germinativos e do crescimento inicial de uma espécie nativa tropical, o presente estudo se fez com Caesalpinia peltophoroides Benth. Essa espécie, conhecida popularmente como sibipiruna ou falso pau-brasil, tem grande potencial ornamental e madeireiro, além de possuir ampla distribuição no Brasil, principalmente na região de Mata Atlântica (LORENZI, 2002).
Recentemente, estudos acerca da fisiologia da germinação de sementes de C. peltophoroides têm sido desenvolvidos BORGES, 2003; BORGES et al., 2005). Entretanto, aspectos bioquímicos e fisiológicos da composição das reservas e sua mobilização nas sementes durante a germinação e crescimento inicial das plântulas ainda são pouco conhecidos.
Este trabalho teve como objetivo avaliar a composição química das sementes, bem como as alterações no conteúdo de carboidratos, lipídios e proteínas durante a germinação e crescimento inicial das plântulas de Caesalpinia peltophoroides.
2. MATERIAL E MÉTODOS
Foram utilizadas sementes de Caesalpinia peltophoroides Benth. Recém-colhidas (provenientes de 15 árvores) em Viçosa, Minas Gerais. Durante o beneficiamento foram eliminadas sementes imaturas, deterioradas ou danificadas.
Os cotilédones utilizados nos ensaios foram obtidos a partir de sementes previamente tratadas com solução de captan 0,5% por 60 seg, as quais foram germinadas sobre areia previamente esterilizada com brometo de metila (20 cc. m-2).
Os cotilédones das sementes ou plântulas foram coletados e amostrados para a determinação dos teores de lipídios, proteínas e carboidratos a cada cinco dias até o estádio de senescência e abscisão dos cotilédones. Assim, os tempos de coleta dos cotilédones compreenderam: sementes quiescentes (T0), sementes aos cinco dias após a semeadura (DAS), período marcado pela protrusão da radícula (T5), plântulas aos 10 DAS (T10), 15 DAS (T15), 20 DAS (T20), 25 DAS (T25), período marcado pela senescência dos cotilédones, 30 e 35 DAS (T30 e T35, respectivamente) e período marcado pela abscisão dos cotilédones.
As amostras (cotilédones intactos) foram secadas em estufa a 45 ºC, por 72 h; após a determinação da matéria seca, foram armazenadas em vidros hermeticamente fechados e mantidos no "freezer" até a extração e quantificação das reservas.
Determinação dos lipídios: A extração e quantificação do teor de lipídios foram realizadas em aparelho tipo Soxhlet, pelo método de extração a frio com éter de petróleo por 24 h, conforme Silva (1990).
Determinação de proteínas solúveis: A extração de proteínas nos cotilédones foi feita conforme Alfenas et al. (1991), utilizando-se o tampão TRIS-HCl, 0,1 M, pH 7,5 como solução de extração. A quantificação dos teores de proteínas foi feita pelo método de Bradford (1976), utilizando-se albumina sérica bovina (BSA) como padrão.
Determinação de carboidratos solúveis e amido: A extração de açúcares solúveis foi feita em banho-maria por 30 min. a 75 ºC (BUCKERIDGE e DIETRICH, 1990) e repetida cinco vezes. Após cada extração, a mistura foi centrifugada, retirando-se o sobrenadante. O precipitado foi secado em estufa a 45 ºC, por 48 h, e submetido à digestão do amido com 1 mL de ácido perclórico 35% (PASSOS, 1996). Os açúcares solúveis e o amido foram quantificados pelo método fenol-sulfúrico (DUBOIS et al., 1956), usando-se glicose (Sigma) como padrão.
O delineamento experimental usado foi inteiramente casualizado. As coletas constaram de cinco repetições de 20 cotilédones com três repetições cada. Amostras da parte aérea e da raiz das plântulas foram coletadas e utilizadas para obtenção das medidas de incremento do peso da matéria seca ao longo do tempo. As equações de regressão foram ajustadas pelo método stepwise (DRAPER e SMITH, 1981), nas concentrações de lipídios, carboidratos e proteínas presentes nos cotilédones das sementes e plântulas ao longo do período de germinação e crescimento, com o objetivo de analisar o comportamento das referidas concentrações em relação ao tempo. Para verificar o grau de associação entre as concentrações dos compostos, assim como a produção de massa seca em cotilédones e plântulas, usou-se a análise do coeficiente de correlação.
3. RESULTADOS E DISCUSSÃO
Sementes de Caesalpinia peltophoroides apresentaram cerca de 50% de lipídios, 32% de carboidratos solúveis, 7,7% de amido e 6,8% de proteínas solúveis, em relação ao peso de matéria seca dos cotilédones (Figura 1). Outros compostos, como carboidratos constituintes de parede, proteínas insolúveis e compostos do metabolismo secundário, estariam compondo os 4% restantes.
Os principais materiais de reserva em sementes de girassol também são estocados nos cotilédones, tendo entre 22 e 36% do seu peso de matéria seca constituído por lipídios (SOROKIN, 1967). Dalbergia miscolobium Benth. apresenta 32% de lipídio e pequena proporção de amido (2,4 a 4,7%) nos cotilédones (SILVA et al., 1998), já em sementes de Euphorbia heterophylla L. o amido não é detectado (SUDA e GIORGINI, 2000).
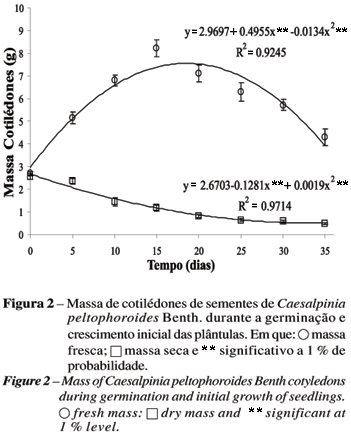
No período pré-germinativo (entre T0 e T5), observou-se aumento exponencial na massa fresca dos cotilédones até T15, em função do incremento da turgescência de seus tecidos, seguido de queda, à medida que as reservas foram exauridas. A massa seca declinou significativamente após T5 (Figura 2).
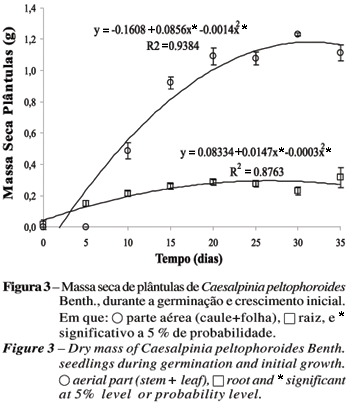
Nota-se, na Figura 3, maior crescimento da raiz em relação à parte aérea no período pré-germinativo. Nesse período, os cotilédones apresentaram pequena diminuição na massa seca (Figura 2), podendo indicar que a hidrólise das reservas ainda estava por se iniciar. Após T5, foi verificada uma aceleração no desenvolvimento da parte aérea da plântula (Figura 3), com o uso do suprimento de nutrientes a partir do tecido de reserva. De T5 a T35, observou-se aumento crescente na produção de biomassa tanto da parte aérea quanto da raiz das plântulas. Segundo Laboriau (1970), a germinação de sementes consiste na retomada do crescimento do eixo embrionário, que é, inicialmente, determinado pela embebição, seguida pela mobilização de reservas para formação de novas estruturas celulares. Entretanto, a produção de massa seca da plântula não pode ser atribuída exclusivamente às reservas energéticas originais dos cotilédones, uma vez que no T15 seu primeiro par de folhas já está presente, havendo provavelmente contribuição de material proveniente da fotossíntese. Da mesma forma, Sassaki e Felippe (1992) verificaram em Dalbergia miscolobium Benth. grande dependência do eixo embrionário em relação às reservas dos cotilédones até os 20 dias, quando começa a maior expansão foliar.
Os carboidratos de reserva armazenados em sementes de leguminosas têm sido aplicados como ferramenta na taxonomia. Muitas Papilionoideae armazenam grandes quantidades de amido em suas sementes; em outras, sua ocorrência é pequena ou ausente. Em Caesalpinioideae e Mimosoideae, essa reserva constitui uma exceção (HEGNAUER e GPAYER-BARKMEIJER, 1993).
Embora haja grandes variações quantitativas entre os constituintes das sementes, qualitativamente eles se apresentam mais estáveis, pelo menos considerando-os em seus grupos e funções nas sementes. Interessante é que parece haver relação inversa em sementes oleaginosas entre o conteúdo de óleo e de proteína (GUIMARÃES, 1999), e tal relação é claramente observada em sementes de Caesalpinia peltophoroides, em que lipídios e proteínas ocuparam proporções máxima e mínima, respectivamente.
A quantidade total de reservas da semente disponível para uma plântula não é determinada somente pela sua massa, mas é também influenciada pela composição química (KITAJIMA, 1996). Considerando a alta proporção de lipídios nas sementes de Caesalpinia peltophoroides, pode-se sugerir essa característica como uma possível vantagem adaptativa à espécie, possibilitando o estabelecimento de plântulas em ambientes menos iluminados, devido ao maior conteúdo energético armazenado. Além disso, Kitajima (1996) relatou que os altos conteúdos de lipídios nas sementes de algumas espécies deve indicar uma seleção compensatória (maior energia/volume), à medida que as sementes mais leves são fortemente selecionadas, por exemplo, para melhor dispersão.
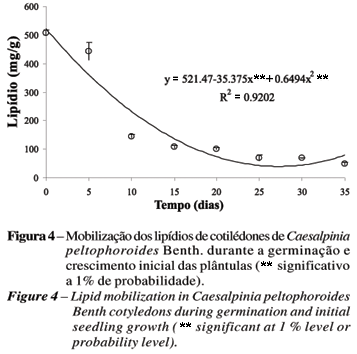
Esses lipídios apresentaram forte queda entre T0 e T10, sendo esta mais suave a partir do T10 (Figura 4). Em espécies do gênero Impatiens, há acentuado declínio de lipídio após o 4º dia após a embebição (NOZZOLILLO e THIE, 1984).
Consumo ainda mais rápido foi encontrado por Suda e Giorgini (2000), em sementes de Euphorbia heterophylla L., cuja degradação dos lipídios se iniciou logo após a embebição, atingindo cerca de 70% entre três e quatro dias após a embebição. Em Cucumis sativus L., a degradação dos lipídios iniciou no 2º dia após a germinação, só restando 3% do total inicial aos seis dias. Essa rápida degradação se inicia com o aparecimento da radícula e termina com a completa expansão dos cotilédones (MATSUI et al., 1999). Entretanto, segundo Suda e Giorgini (2000), esse padrão não é usual, quando comparado com outras sementes oleaginosas, nas quais o conteúdo de lipídio permanece inalterado durante o período inicial da germinação, diminuindo posteriormente. Observou-se em Arabidopsis thaliana (L.) Heynh. que a mobilização das reservas de lipídio é inibida na presença de açúcares solúveis, como glicose e sacarose, em geral provindos do metabolismo do amido (TO et al., 2002).
Teores de carboidratos solúveis reduziram-se continuamente durante todo o período observado (Figura 5). O conteúdo de carboidratos apresentou queda mais acentuada após o 10º dia, concomitantemente com a diminuição na degradação das reservas de lipídios (Figura 4). Resultados semelhantes foram observados por Buckeridge et al. (1992), em sementes de Copaifera langsdorfii Desf., cujo conteúdo de açúcares solúveis reduziu durante e após a germinação. Ao contrário, sementes de Brassica oleracea L. e Euphorbia heterophylla L., tiveram o conteúdo de carboidratos solúveis inalterados até 48 e 72 h, respectivamente (QOUTA et al., 1991; SUDA e GIORGINI, 2000).
Pode-se sugerir que a grande quantidade de carboidratos solúveis observada em sementes de Caesalpinia peltophoroides se deve não só ao fornecimento de energia para a germinação, mas também confee maior armazenabilidade a essas sementes.
As reservas de amido apresentaram pequenas concentrações em sementes de Caesalpinia peltophoroides, semelhante ao que ocorreu em outras leguminosas (IRVING, 1984; HEGNAUER e GPAYERBARKMEIJER, 1993; SILVA et al., 1998; SUDA e GIORGINI, 2000), tendo apresentado pequeno consumo durante o período germinativo e de crescimento das plântulas (Figura 5).
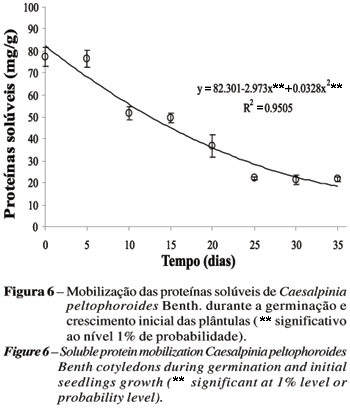
O conteúdo protéico apresentou-se inalterado no período pré-germinativo, com redução gradativa após a protrusão da radícula (T5). Borges et al. (2005) relataram que o teor de proteína mantém-se estável nos cotilédones dessa espécie até 24 h de embebição. Schlereth et al. (2000) observaram que em sementes de Vicia sativa L. a mobilização das proteínas nos cotilédones só começa após 48 h de embebição.
O conteúdo de proteínas solúveis decresce durante o crescimento das plântulas (Figura 6), o que concorda com a afirmação de Bewley e Black (1994), de que as proteínas são mobilizadas durante a germinação e subseqüente crescimento das plântulas. Esses resultados também concordam com os obtidos por Müntz et al. (2001) de que a mobilização das proteínas armazenadas nos cotilédones só é detectável após a protrusão da radícula (pós-germinação). Em sementes de Erythrina velutina Willd., o conteúdo de proteína cotiledonar solúvel decresceu rapidamente durante a germinação e crescimento da plântula (OLIVEIRA et al., 1998), e em sementes de Euphorbia heterophylla as proteínas foram continuamente degradadas após o inicio da embebição (SUDA e GIORGINI, 2000).
Isso ocorre porque a germinação da semente é iniciada graças às reservas próprias do embrião e depois mantida com o consumo dos componentes dos tecidos de reserva, pela atividade enzimática e pelo fluxo dos componentes solúveis às regiões de crescimento onde há rápido consumo (CARVALHO e NAKAGAWA, 2000).
Yamaguchi et al. (1996) e Matsui et al. (1999) observaram que em sementes de Cucumis sativus a degradação das reservas de proteínas inicia-se aos três dias após a germinação e que a diminuição desta está claramente correlacionada com a transição do metabolismo dos cotilédones, dependente da fonte da energia preservada nas sementes para o crescimento autotrófico. Nesse contexto, Buckeridge et al. (2004) também evidenciaram a mobilização das reservas de proteínas para a estruturação dos processos que conferem capacidade de absorver nutrientes e realizar fotossíntese.
A Tabela 1 ilustra o grau de associação entre as concentrações dos compostos químicos analisados nos cotilédones, bem como a produção ou consumo de massa seca em plântulas e cotilédones, através do coeficiente de correlação. Para análise do coeficiente de correlação, Shimakura (2002) descreveu: correlação muito fraca (0,00 a 0,19), correlação fraca (0,20 a 0,39), correlação moderada (0,40 a 0,69), correlação forte (0,70 a 0,89), e correlação muito forte (0,90 a 1,00).
O peso da massa seca dos cotilédones apresenta correlação positiva muito forte em relação aos teores de lipídio, amido e proteína (0,9691; 0,9311 e 0,9772, respectivamente) e correlação forte (0,8107) com o teor de carboidrato. Ao contrário, o peso da massa seca dos cotilédones apresenta correlação negativa muito forte (- 0,9872) com o peso de massa seca das plântulas. Do mesmo modo, o peso de massa seca das plântulas apresenta correlação negativa muito forte com os teores de lipídio e proteínas (- 0,9480 e - 0,9513, respectivamente) e correlação negativa forte com os teores de carboidratos solúveis e amido (- 0,8017 e - 0,8947, respectivamente).
Pode-se afirmar, então, que a diminuição no peso da massa seca dos cotilédones está diretamente associada à redução nos compostos de reserva neles estocados e associada ao aumento do peso de massa seca da plântula. Essas relações evidenciam a mobilização dos compostos de reserva dos cotilédones (fonte) e sua translocação para outros órgãos (dreno), à medida que a redução na massa seca cotiledonar reflete o aumento na produção de biomassa de parte aérea e raiz da nova plântula. Resultados semelhantes foram obtidos durante a germinação e o crescimento inicial de Brassica oleracea e espécies do gênero Impatiens (NOZZOLILLO e THIE, 1984; QOUTA et al., 1991).
Em relação aos teores de lipídio, carboidratos solúveis, amido e proteínas solúveis, bservam-se correlações sempre positivas. As proteínas apresentam correlação positiva muito forte com lipídio e amido (0,9122 e 0,9334, respectivamente) e forte com carboidratos (0,8859). O amido apresentou correlação forte com lipídio e carboidratos (0,8674 e 0,8840, respectivamente). Já os carboidratos exibiram correlação moderada com o teor de lipídio (0,6541). Essas correlações positivas demonstram semelhante comportamento de degradação entre esses compostos.
As correlações positivas de degradação observadas entre os compostos não significam que o consumo de um causa, necessariamente, o consumo do outro ou vice-versa, de forma que nem sempre os dados observados estabelecem relações causais. Para afirmações dessa natureza, seria necessário conduzir experimentos específicos para a obtenção de mais evidências de uma relação causal. Isso indica a possível existência de processos de conecção que atuariam inseridos num amplo e complexo sistema metabólico, no sentido de mobilizar as reservas energéticas estocadas nos cotilédones das sementes com o objetivo final de estabelecer uma nova planta.
4. CONCLUSÃO
Sementes de Caesalpinia peltophoroides possuem alto conteúdo de lipídios, os quais são predominantemente consumidos durante a germinação.
Lipídios, proteínas e carboidratos representam as reservas consumidas durante a germinação e crescimento inicial das plântulas.
O consumo das reservas energéticas dos cotilédones foi fortemente correlacionado com a redução no peso de massa seca destes e, inversamente correlacionado com o aumento na produção de biomassa nas plântulas.
5. REFERÊNCIAS BIBLIOGRÁFICAS
Recebido em 12.06.06 e aceito para publicação em 19.11.2006.
- ALFENAS, A.C. et al. Eletroforese de proteínas e isoenzimas de fungos e essências florestais. Viçosa, MG: SIF, 1991. 242 p.
- BEWLEY, J.D.; BLACK, M. Seeds: Physiology of development and germination. 2. ed. New York: Plenum Press, 1994. 445 p.
- BORGES, E.E.L. Comportamento bioquímico e fisiológico de sementes florestais nativas durante a embebição 2003. 100 f. Tese (Doutorado em Ecologia e Recursos Naturais) Universidade Federal de São Carlos, São Carlos, 2003.
- BORGES, E.E.L.; RENA, A.B. Germinação de sementes. In: AGUIAR, I.B.; PIÑA-RODRIGUES, F.C.M.; FIGLIOLIA, M.B. Sementes florestais tropicais Brasília: ABRATES, 1993. 350 p.
- BORGES, E.E.L. et al.. Caracterização de Alfagalactosidase e sua relação com a germinação das sementes de Caesalpinia peltophoroides (Leguminosae Caesalpinoideae). Revista Árvore, v. 29, n. 4, p. 525-533, 2005
- BRADFORD, M. M. A rapid and sensitive method for the quantification of microgram quantities of proteins utilizing the principle of protein-dye binding. Analytical Biochemistry, v. 72, p. 248-254, 1976.
- BUCKERIDGE, M.S. et al. Xiloglucan structure and post-germinative metabolism in seeds of Copaifera langsdorfi from savanna and forest populations. Physiologia Plantarum, v. 86, p. 145-151, 1992.
- BUCKERIDGE, M.S. et al. Acúmulo de Reservas. In: FERREIRA, A.G.; BORGHETTI, F. Germinação: do básico ao aplicado. Porto Alegre: Artmed, 2004. 324 p.
- BUCKERIDGE, M.S.; DIETRICH, S.M.C. Galactomanans from Brazilian legume seeds. Revista Brasileira de Botânica, v. 13, p. 109-112, 1990.
- CARVALHO, J.E.U.; NAKAGAWA, J. Sementes: ciência, tecnologia e produção. Jaboticabal: Funep, 2000. 588 p.
- DRAPER, W.R.; SMITH, H. Applied regression analysis New York: Jonh Willey & Sons, 1981. 709 p.
- DUBOIS, M. et al. Colorimetric method for determination of sugars and related substances. Annalical Chemestry, v. 28, p. 350-356, 1956.
- GUIMARÃES, R.M. Fisiologia de sementes Lavras:UFLA/ FAEPE, 1999. 79 p.
- HEGNAUER, R.; GPAYER-BARKMEIJER, R.J. Relevance of seed polysaccharides and flavonoids for the classification of the leguminosae: A chemotaxonomic approach. Phytochemistry, v. 34, n. 1, p. 3-16, 1993.
- IRVING, D.W. Seed structure and histochemistry of Prosopis velutina (Leguminosae). Botanical Gazete, v. 145, n. 3, p. 340-345, 1984.
- KITAJIMA. K. Ecophysiology of tropical tree seedlings. In: MULKEY, S.S.; CHAZDON, R.L.; SMITH, A.P. Tropical forest plant ecophysiology New York: Chapman & Hall, 1996. p. 559-596.
- LABORIAU, L.G. On the physiology of seed germination in Vicia graminea Sm. I. Anais da Academia Brasileira de Ciências, v. 42, n. 2, p .235-262, 1970.
- LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. Nova Odessa: Platarum, 2002. 352 p.
- MATSUI, K. et al. Cucumber Cotyledon Lipoxygenase during Postgerminative Growth. Its Expression and Action on Lipid Bodies. Plant Physiology, v. 119, p. 1279-1287, 1999.
- MAYER A.M.; POLJAKOFF-MAYBER A. The germination of seeds New York: Pergamon Press, McMillan, 1975. 236 p.
- MÜNTZ, K. et al. Stored proteinases and the initiation of storage protein mobilization in seeds during germination and seedling growth. Journal of Experimental Botany, v. 52, n. 362, p. 1741-1752, 2001.
- NOZZOLILLO, C.; THIE, I. A comparative study of mobilization of lipid and carbohydrate reserves during germination of seeds of three species of Impatiens (Balsaminaceae): I. balsamina L., I. capensis Meerb. and I. ballida Nutt. Torrey Botanical Club, v. 111, n. 2, p. 200-208, 1984.
- OLIVEIRA, J.T.A. et al. Protein and lecitin mobilization during Erythrina velutina forma Aurantiaca seed germination and seedling growth in the dark. Revista Brasileira de Fisiologia Vegetal, v. 10, n. 1, p. 25-30, 1998.
- PASSOS, L. P. Métodos analíticos e laboratoriais em fisiologia vegetal Coronel Pacheco: EMBRAPA-CNPGL, 1996. 223 p.
- PONTES, A. P. et al. Mobilização de reservas em sementes de Apuleia leiocarpa (Vogel) J. F. Macbr. (garapa) durante a embebição. Revista Árvore, v. 26, n. 5, p. 593-601, 2002.
- QOUTA, L.A. et al. Changes in seed reserves and cell wall composition of component organs during germination of cabbage (Brassica oleracea) seeds. Journal of Plant Physiology, v. 138, p. 700-707, 1991.
- SASSAKI, R.M.; FELIPPE, G.M. Remoção dos cotilédones e desenvolvimento inicial de Dalbergia miscolobium Revista Brasileira de Botânica, n. 15, p. 5-16, 1992.
- SCHLERETH, A. et al. Comparison of globulin mobilization and cysteine proteinases in embryonic axes and cotyledons during germination and seedlings growing of vetch (Vicia sativa L.). Journal of Experimental Botany, v. 51, n. 349, p. 1423-1433, 2000.
- SHIMAKURA, S.E. Interpretação do coeficiente de correlação [Disponível em: www.est.ufpr.br acessado em 07 de junho de 2005].
- SILVA, D.J. Análise de alimentos - métodos químicos e biológicos. Viçosa, MG: Universidade Federal de Viçosa, 1990. 165 p.
- SILVA, T.R.G.; CORTELAZZO, A.L.; DIETRICH, S.M.C. Variations in storage compounds during germination and early plantlet growth of Dalbergia miscolobium Revista Brasileira de Fisiologia Vegetal, v. 10, n. 2, p. 119-124, 1998.
- SOROKIN, H.P. The spherosomes and the reserve fat in plant cells. American Journal Botany, v. 54, n. 8, p. 1008-1016, 1967.
- SUDA, C.N.K.; GIORGINI, J.F. Seed reserve composition and mobilization during germination and initial seedling development of Euphorbia heterophylla Revista Brasileira Fisiologia Vegetal, v. 12, n. 3, p. 226-245, 2000.
- TO, J.P.C.; REITER, W.; GIBSON, S.I. Mobilization of seed storage lipid by Arabidopsis seedlings is retarded in the presence of exogenous sugars. BMC Plant Biology, v. 2, p. 4-15, 2002.
- YAMAGUHCI, Y. et al. Emergence of proteases in germinating cucumber cotyledons and their roles in the two-step degradation of storage protein. Plant Cell Physiology, v. 37, p. 279284, 1996.
Datas de Publicação
-
Publicação nesta coleção
27 Fev 2007 -
Data do Fascículo
Dez 2006
Histórico
-
Aceito
19 Nov 2006 -
Recebido
12 Jun 2006