Abstract
The present study aimed at assessing the effects of methionine and arginine on the immune response of broiler chickens submitted to immunological stimuli. Three methionine concentrations (0.31, 0.51, and 0.66% from 1 to 21 days of age; 0.29, 0.49, and 0.64% from 22 to 42 days of age) and 2 arginine concentrations (1.33 and 1.83%; 1.14 and 1.64% for the same life periods) were tested. Birds were divided into two groups for immunological stimuli (3x2x2 arrangement). Vaccines against Marek's disease, fowl pox, infectious bronchitis, Freund's Complete Adjuvant, Sheep red blood cells (SRBC), and avian tuberculin were administered to one group as immunological stimuli; the other group did not receive any stimulus. The experiment was carried out with 432 one-day-old male Ross broilers, distributed into 12 treatments with 6 replicates of 6 birds each. Performance data were weekly collected. Anti-SRBC antibodies were collected by hemagglutination test and cell immune response (CIR) was measured by tubercularization reaction in one wattle 24 hours after administration of the second tuberculin injection at 42 days of age. The weight difference between the two wattles of each bird (one injected with tuberculin and the other not) was the measure of CIR. Arginine levels did not influence either bird performance or immune response. Methionine concentrations higher or lower than usually adopted in broiler production (0.51 and 0.49%) equally failed to influence the birds' immune humoral response, but the best CIR was observed at the intermediate methionine level. Vaccines administered on the first day of age impaired bird performance up to the 21st day of age.
Amino acids; birds; immune system; vaccines
Effects of methionine and arginine dietary levels on the immunity of broiler chickens submitted to immunological stimuli
Rubin LLI; Canal CWI; Ribeiro ALMII, III; Kessler AII; Silva III; Trevizan LII; Viola TII; Raber MII; Gonçalves TAII; Krás RII
ILaboratory of Virology, Departamento de Zootecnia, Departamento de Medicina Animal, Universidade Federal do Rio Grande do Sul
IILaboratory of "Ensino Zootécnico", Departamento de Medicina Animal, Universidade Federal do Rio Grande do Sul
IIIAssociated Professor, Departamento de Medicina Animal, Universidade Federal do Rio Grande do Sul
Mail Address Mail Address: Andréa Machado Leal Ribeiro UFRGS Av. Bento Gonçalves, 7712 91.540-000. Porto Alegre, RS, Brasil E-mail: aribeiro@ufrgs.br
ABSTRACT
The present study aimed at assessing the effects of methionine and arginine on the immune response of broiler chickens submitted to immunological stimuli. Three methionine concentrations (0.31, 0.51, and 0.66% from 1 to 21 days of age; 0.29, 0.49, and 0.64% from 22 to 42 days of age) and 2 arginine concentrations (1.33 and 1.83%; 1.14 and 1.64% for the same life periods) were tested. Birds were divided into two groups for immunological stimuli (3x2x2 arrangement). Vaccines against Marek's disease, fowl pox, infectious bronchitis, Freund's Complete Adjuvant, Sheep red blood cells (SRBC), and avian tuberculin were administered to one group as immunological stimuli; the other group did not receive any stimulus. The experiment was carried out with 432 one-day-old male Ross broilers, distributed into 12 treatments with 6 replicates of 6 birds each. Performance data were weekly collected. Anti-SRBC antibodies were collected by hemagglutination test and cell immune response (CIR) was measured by tubercularization reaction in one wattle 24 hours after administration of the second tuberculin injection at 42 days of age. The weight difference between the two wattles of each bird (one injected with tuberculin and the other not) was the measure of CIR. Arginine levels did not influence either bird performance or immune response. Methionine concentrations higher or lower than usually adopted in broiler production (0.51 and 0.49%) equally failed to influence the birds' immune humoral response, but the best CIR was observed at the intermediate methionine level. Vaccines administered on the first day of age impaired bird performance up to the 21st day of age.
Keywords: Amino acids, birds, immune system, vaccines.
INTRODUCTION
Several kinds of animal feed supplements of different origins have been recently developed. The general aim is to enhance, either through an exogenous pathway or by stimulating the animal's immune response, a health status that in turn may allow a gradual decrease in antibiotics use. Among such compounds, there are some additives widely acknowledged and extensively used in animal nutrition, such as fatty acids, vitamins and minerals, as well as acidifiers, enzymes, flavoring and antioxidant agents, coccidiostats, and probiotics, which are usually employed at higher levels than strictly nutritional compounds. In addition, a more recent class of substances has been used to activate animal immune system, including oral immunoglobulins, intestinal flora stimulators, and oligosaccharides that block the adhesion of pathogenic bacteria.
Methionine (Met) (Swain & Johri, 2000; Rama Rao et al., 2003; Shini & Brydeen, 2005) and arginine (Tayade et al., 2006ab; Lee et al., 2002) are two amino acids that have proven immune regulatory action. Methionine is an essential amino acid with at least four main roles. First, Met participates in protein synthesis. Second, Met is a glutathione precursor, a tripeptide that reduces reactive oxygen species (ROS) and thus protects cells from oxidative stress. Third, methionine is required for the synthesis of polyamines (spermine and spermidine), which take part in nucleus and cell division events. Fourth, methionine is the most important methyl group donor for methylation reactions of DNA and other molecules.
Several studies demonstrated that methionine constructively affects the immune system, improving both cellular and humoral immune response. It was reported that methionine requirements for optimal immunity are higher than for optimal growth (Tsiagbe et al., 1987; Swain & Johri, 2000; Shini & Bryden, 2005), and that restriction of sulfur amino acids (SAA) results in severe lymphocyte depletion in intestinal tissues (Peyer's patches) and lamina propria (Swain & Johri, 2000). One of the mechanisms proposed to explain methionine interference in the immune system is the proliferation of T cells, which are sensitive to intracellular glutathione and cysteine levels, compounds also participate in methionine metabolism (Kinscherf et al., 1994).
Arginine, a dibasic amino acid, (Boorman & Lewis, 1971) is essential to birds. In addition of being required to optimal growth, arginine is also important for a series of biological and physiological functions, including protein biosynthesis, nitrogen transport and excretion, production of polyamides and nitric oxide, and stimulation of several endocrine glands (Efron & Barbul, 1998; Efron & Barbul, 2000). Due to the lack of some enzymes in the urea cycle, broiler chickens are unable to biosynthesize arginine from ornithine; thus, this amino acid must be supplied in diets (Wu et al., 1995). It was recently demonstrated that L-arginine increases specific immune response against Infectious Bursal Disease (IBD) in chickens (Tayade et al., 2006a). In humans, arginine supplements increase the proliferation of lymphocytes in the blood, and boost suppressor T-cell counts (Barbul et al., 1981). In addition, it was observed that arginine increases phagocytic activity of alveolar macrophages in rats (Tachibana et al., 1985). Arginine is a precursor of nitric oxide, an important microbiocidal molecule involved in the role played by macrophages as "exterminators" (Koshland, 1992). Moreover, arginine participates in the regulation of the interactions between macrophages and lymphocytes, as well as in lymphocyte activation and adhesion.
This study was designed to verify if dietary methionine levels and arginine:methionine ratio modulate the immune response of broiler chickens submitted to an immunological stimulus series.
MATERIALS AND METHODS
This experiment used 432 1-day-old Ross 308 male broiler chickens. Chickens were housed in metal battery brooders placed in a controlled temperature environment, and were allowed free access to water and feed throughout the growth period (1 to 42 days). The baseline diet (Table 1) was supplemented with two methionine concentrations (0.20 and 0.35 percent), which ultimately resulted in 3 digestible methionine levels (0.31, 0.51, and 0.66% in the starter phase, and 0.29, 0.49, and 0.64% in the grower phase). Similarly, the addition of a single arginine concentration (0.50 percent) led to 2 digestible levels in diets (1.33 and 1.83% in the starter phase, and 1.14 and 1.64% in the grower phase). Methionine was supplemented as methionine hydroxyl analog (HMTBA liquid-phase methionine), considering 88% bioavailability, whereas arginine was supplemented as L-arginine, with a 99% bioavailability.
Birds were divided into two groups. The first group was submitted to immunological stimulus (IS) with vaccines commonly used in industrial poultry production, Freund's Complete Adjuvant, avian tuberculin, and Sheep Red Blood Cells (SRBC). The second group was not challenged (positive control).
On the first day of life, vaccines against Marek's disease and fowl pox were administered via intradermal injection, and infectious bronchitis as ocular vaccine. Freund's Complete Adjuvant (inactivated Mycobacterium avium + mineral oil) was administered at 0.5mL (4 mg organisms/mL) via intramuscular injection in the chest muscle on the 15th day of age. On days 29 and 41, birds received one intradermal avian tuberculin injection at 0.1mL in one of the wattles to trigger tubercularization reaction. Twenty four hours after the administration of the second tuberculin injection (42 days of age), birds were sacrificed by cervical dislocation. The two wattles of the same bird (one wattle submitted to the tubercularization reaction) were collected and weighed. The analytic criterion was the weight difference between the two wattles of each bird.
SRBC diluted at 10% in sterile phosphate buffered saline solution (PBS), pH 7.2 (v/v), was administered at 0.5mL dose per bird by intramuscular injection on the 21st and the 35th day of age. Sera were collected on the 42nd day of age, and specific antibodies against SRBC were enumerated by Bartlett & Smith (2003) hemagglutination technique and expressed in log2. Antibody titers were analyzed for normality of variance (Shapiro & Wilk, 1965), and, when variance was determined as normal, titer data were submitted to analysis of variance.
Challenged and non-challenged bird groups were raised in two separate rooms with equal humidity and temperature. In each room, treatments were randomly distributed. Non-vaccinated birds were kept in the experiment until birds from both groups reached the same body weight. Performance was weekly measured as weight gain (WG), feed intake (FI), and feed conversion ratio (FCR).
The 12 treatment series (3 methionine concentrations x 2 arginine concentrations x 2 IS arrangements) were carried out as 6 replicates of 6 birds each until 21 days, when the two IS groups reached the same body weight. The 6 remaining treatments were analyzed (3 methionine concentrations x 2 arginine concentrations) for the period of 22 to 42 days of age, as well for the entire experimental period (1 to 42 days). Experimental data were evaluated using ANOVA procedure of the SAS software package (2001). When the F test produced significant results, means were submitted to LS means test.
RESULTS AND DISCUSSION
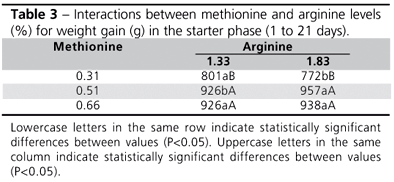
In the starter phase, a significant interaction between methionine and arginine levels was observed (P<0.05) for WG, and will be discussed later. As to the main effects (Table 2), the group of non-vaccinated birds presented better WG, FI, and FCR results. This effect was not observed after 21 days of age (data not shown), when birds from both groups reached the same weight. It was also noticed that the lowest methionine level led to lower WG and FI, and also to higher FCR as compared to the other levels. Arginine levels, regardless the nature of the correlation with methionine levels, did not produce any significant difference in bird performance.
Routine vaccines, as adopted in large-scale poultry production impaired bird performance during the starter rearing phase (Table 2), which is consistent with previous studies (Rubin et al., 2007; Chamblee et al., 1992). One hypothesis to explain these results is that the vaccines administered on the first day of age lead to FI decrease, and to inefficient nutrient absorption and metabolism, caused by the triggered immune response (Klasing et al., 1987), which eventually negatively affected FI in vaccinated chickens. A series of metabolic events may lie underneath this finding, such as redistribution of bivalent cations, higher energy expenditure, higher breakdown degradation of skeletal muscle, more intense amino acid oxidation, and also higher glycogenesis mediated by amino acids (Klasing & Barnes, 1988).
The interaction showed that, at the lowest methionine level (0.31%), the lowest arginine level (1.83%) produced the best WG figure as compared to the highest arginine concentration. At the intermediate methionine level, the highest arginine level produced better WG. At the highest methionine concentration in broiler chicken feed, arginine concentration did not influence WG (Table 3).
The highest arginine level used along with the lowest methionine level in broiler chicken feeds led to lower WG value. This was also previously observed by Keshavaraz & Fuller (1971) in 1- to 21-day-old chickens using similar concentrations as those adopted in the present study (1.68% arginine and 0.36% methionine). Fuller et al. (1967) also reported lower WG in young chickens when higher arginine concentrations were supplemented to corn- and soybean meal-based feed. In chickens, creatine biosynthesis is an important factor in arginine metabolism during the first 4 weeks of life (Fisher et al., 1956), since arginine and glycine are biological precursors of creatine (they form glycocyamine, which, together with the methionine methyl group, is converted into creatine) (Du Vigneaud et al., 1941). The methionine-arginine interaction is a typical example of "amino acid imbalance", according to Harper's classification (1964). High dietary arginine concentrations may cause heavy consumption of methionine methyl groups to generate creatine, which lowers the amount of methionine effectively available for growth.
As to performance parameters, Ruiz-Feria et al. (2001) observed that the increase in arginine levels from 1.25 to 1.47% failed to significantly improve WG of 1- to 21-day-old broiler chickens. Similarly, Cuca & Jensen (1990) reported that, at this phase, arginine requirements for optimal WG and FCR were 1.24 and 1.28%, respectively, in a 21% crude protein diet. Nevertheless, Kidd et al. (2001) observed that increasing dietary arginine concentrations from 1.25 to 1.50% resulted in higher WG (P<0.05) in 1- to 18-day-old broilers at a dietary methionine concentration of 1.05%. In another experiment, the same author did in fact observe that increasing arginine concentration from 1.12 to 1.50% did not affect WG or FCR (Kidd et al., 2001). Deng et al. (2005) reported that no statistical difference was observed in FCR and WG in male Leghorn 1- to 28-day-old chickens when arginine levels varying from 100 to 130% over the values recommended by the NRC (1994) were fed. In a similar way, the present study used arginine concentrations between 106 and 146% of the suggested NRC values (for 1- to 21-day-old broilers), and no statistically significant differences were observed in FCR or WG.
Vieira et al. (2004) observed that 0.35% methionine resulted in lower WG and higher FCR values as compared to 0.48 and 0.58% methionine levels recommended for commercial broilers breeds (Ross and Cobb) between the 14th and the 35th day of age, which is consistent with the results of the present study. That study reported a trend (P<0.07) of FI reduction, under the same experimental conditions. Conversely, Fatufe & Rodehutscord (2005) stated that the best methionine concentration for optimal WG and FC of Ross broilers, between 8 to 21 days of age, was 0.33%, when tested levels ranged between 0.18-0.70%.
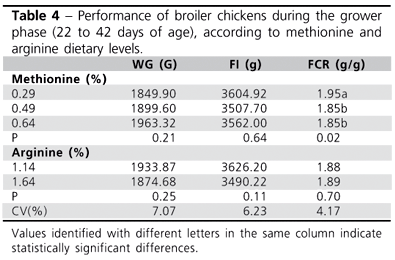
In the grower phase (Table 4), when only the vaccinated birds were examined, no significant difference in WG and FC was observed for the methionine levels used, but an impaired FC was already observed at the lowest methionine concentration. During this phase, there were no treatment interactions. Arginine content did not produce significant differences in performance, although the highest arginine level numerically decreased FI (P<0.11), and this deserves attention in future studies. Such finding may also be interpreted as a result of amino acid imbalance, but broilers at this age are less sensitive to nutritional changes.
Gonzales-Esquerra and Leeson (2006) did not observe differences in FI or in WG in 26- to 33-day-old broilers fed arginine levels of 0.94 or 1.54%. Also, Tayade et al. (2006a) reported that the supplementation of 2% L-arginine in feeds was safe, and did not produce any detrimental effect when used in 21- to 42-day-old broiler chickens. Likewise, Corzo et al. (2006) did not find any statistically significant difference (P>0.05) in WG when Met content in diet varied between 0.31-0.45%. Yet, in a previous study carried out by our research group (Rubin et al., 2007), it was shown that methionine levels between 0.37 and 0.47% reduced WG (P<0.07), as compared to a 0.57% level. As for FCR and FI, our previous results agree with the findings of the present study: methionine did not influence FI, whereas methionine levels under 0.49% negatively affected FCR.
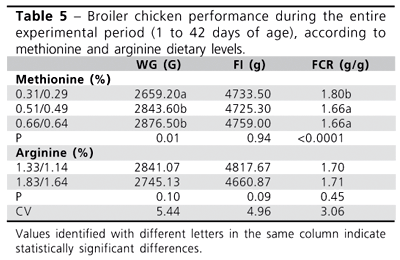
For the entire experimental period (Table 5), the hypothesis that the lowest methionine level would lead to worse WG and FCR was confirmed, as compared to the other concentrations tested. As for arginine, the different concentrations did not produce any statistically significant difference in performance parameters.
As to immune humoral response (Table 6), the tested dietary methionine and arginine levels did not influence the production of antibodies against SRBC. In addition, there was no significant action between treatments.
Takahashi et al. (1993), Takahashi et al. (1994), and Swain & Johri (2000) demonstrated that neither the excess nor the deficiency of methionine in diets influenced the production of primary antibodies in chickens. On the other hand, Rama Rao et al. (2003) showed that methionine supplementation increased anti-SRBC antibody titers, and that the best methionine concentration for antibody production was the highest (0.55%). The SRBC administration pathway, genetic traits, as well as bird sex may have influenced antibody production. Yet, in that study, the authors used females of four different genetic strains, and SRBC was intravenously injected. The present study used male Ross chickens as the animal model, and SRBC was injected in the muscle.
As for arginine, Kidd et al. (2001) demonstrated that arginine levels between 1.48 and 1.68% failed to enhance the immune response against SRBC and Newcastle vaccine, which is consistent with the results obtained in the present study. However, Tayade et al. (2006a) proved that the supplementation of 2% arginine in diets increased antibody counts and the protection against infectious bursal disease virus.
In terms of immune cell response (Table 6), the lowest Met level resulted in a diminished tubercularization reaction, as compared to the intermediate concentration. Such finding suggests a decreased proliferation of Th1 lymphocytes in inflammation sites. The present study is in agreement with the results of Swain & Johri (2000) and Rama Rao et al. (2003), showing that methionine levels lower than 0.50% generate a poorer immune cell response as compared to higher concentrations.
No significant effect of arginine on immune cell response was observed, which is again consistent with the results of Kidd et al. (2001) and Deng et al. (2005), whose studies that did not reveal any significant difference either in anti-SRBC antibody production or in immune cell response against phytohemagglutinine. However, Tayade et al. (2006b) demonstrated that the supplementation of 2% L-arginine stimulated both intestinal and systemic immune responses against infectious bronchitis virus (IBV). Lee et al. (2002) observed that CD8+ cells percentage in the lung lavage fluid of IBV-challenged chickens was lower than in chickens fed with arginine-deficient diets (0.50%), as compared with regular arginine-adequate diets and with feeds containing excess arginine (3.00%). It is important to emphasize the study mentioned was conducted with specific-pathogen-free birds of a different strain (Leghorn), using a different immunogen and different arginine levels (0.5, 1.0 and 3.0%) as compared to the present study (1.33 and 1.83%). Interactions between nutrients and the immune system are largely dependent on the variables mentioned above. Therefore, results obtained using different experimental methods have to be compared with caution.
CONCLUSIONS
In general, methionine levels below or above the concentrations usually applied in commercial poultry production did not influence the humoral immune response of birds, though the likelihood of a dose-response correlation between methionine and cell-mediated immune response cannot be neglected, and the best response is achieved when usual methionine concentrations are used. The present study also establishes that the vaccines administered on the first day of age impaired broiler chicken performance up to 21 days of age. Therefore, it is recommended to administer vaccines carefully, taking into account the risk of losses caused by disease as compared to losses caused by vaccines. It is also important to emphasize that further studies are needed to establish alternative optimal nutrition characteristics in order to minimize such "inevitable" losses. The arginine concentrations tested in the present study did not influenced either broiler chicken performance, or humoral and cell immune responses.
Acknowledgements
We thank Professor Jaime Cobuci that helped the statistical analysis.
Arrived: December / 2006
Approved: November / 2007
- Barbul A, Sisto DA, Wasserkrug HL, Efron G. Arginine stimulates lymphocyte immune response in healthy human beings. Surgery 1981; 90(2):244-51.
- Bartlett JR, Smith MO. Effects of different levels of zinc on the performance and immunocompetence of broilers under heat stress. Poultry Science 2003; 82(10):1580-8.
- Boorman KN, Lewis D. Protein metabolism. In: Bell DJ, Freeman BM, editors. Physiology and Biochemistry of the Domestic Fowl. New York: Academic Press; 1971. p. 339-372.
- Chamblee TN, Brake JD, Schultz CD, Thaxton JP. Yolk sac absorption and initiation of growth in broilers. Poultry Science 1992; 71:1811-1816.
- Corzo A, Kidd MT, Dozier III WA, Shack LA, Burgess SC. Protein expression of pectoralis major muscle in chickens in response to dietary methionine status. British Journal of Nutrition 2006; 95: 703-708.
- Cuca M, Jensen LS. Arginine requirement of starting broiler chicks. Poultry Science 1990; 69(8):1377-82.
- Deng K, Wong CW, Nolan JV. Long-term effects of early life L-arginine supplementation on growth performance, lymphoid organs and immune responses in Leghorn-type chickens. British Poultry Science 2005; 46(3):318-324.
- Du Vigneaud V, Chon M, Chandler JP, Schenck JR, Simmonds S. The utilization of the methyl group of methionine in the biological synthesis of choline and creatine. The Journal of Biological Chemistry 1941; 140:625.
- Efron DT, Barbul A. Modulation of inflammation and immunity by arginine supplements. Current Opinion in Clinical Nutrition and Metabolic Care 1998; 1:531-538.
- Efron DT, Barbul A. Role of arginine in immunonutrition. Journal of Gastroenterology 2000; 35(Suppl. 12):20-23.
- Fatufe AA, Rodehutscord M. Growth, body composition, and marginal efficiency of methionine utilization are affected by nonessential amino acid nitrogen supplementation in male broiler chicken. Poultry Science 2005; 84(10):1584-92.
- Fisher H, Salander RC, Taylor MW. The influence of creatine biosynthesis on the arginine requirements of chick. The Journal of Nutrition 1956; 59:491.
- Fuller HL, Chang SI, Potter DK. Detoxication of dietary tannic acid by chicks. The Journal of Nutrition 1967; 91:477.
- Gonzalez-Esquerra R, Leeson S. Concentrations of putrescine, spermidine, and spermine in duodenum and pancreas as affected by the ratio of arginine to lysine and source of methionine in broilers under heat stress. Poultry Science 2006; 85:1398-1408.
- Harper AE. Amino acid toxicities and imbalances. In: Munro HN, Allison JB, editors. Mammalian Protein Metabolism. New York: Academic Press; 1964. p. 87.
- Keshavarz K, Fuller HL. Relationship of arginine and methionine in the nutrition of the chick and the significance of creatine biosynthesis in their interaction. The Journal of Nutrition 1971; 101(2):217-222.
- Kidd MT, Peedles ED, Whitmarsh SK, Yeatman JB, Wideman RF Jr. Growth and immunity of broiler chicks as affected by dietary arginine. Poultry Science 2001; 80:1535-1542.
- Kinscherf R, Fischbach T, Mihm S, Roth S, Hohenhaus-Sievert E, Weiss C, Edler L, Bärtsch P, Dröge W. Effect of glutathione depletion and oral N-acetyl-cysteine treatment on CD4+ and CD8+ cells. The FASEB Journal 1994; 8:448-451.
- Kirk SJ, Regan MC, Wasserkrug HL, Sodeyama M, Barbul A. Arginina enhances T-cell responses in athymic nude mice. Journal of Parenteral and Enteral Nutrition 1992; 16:429-432.
- Klasing KC, Barnes DM. Decreased amino acid requirements of growing chicks due to immunological stress. The Journal of Nutrition 1988; 118:1158-1164.
- Klasing KC, Laurin DE, Peng RK, Fry DM. Immunologically mediated growth depression in chicks: influence of feed intake, corticosterone, and interleukin-1. The Journal of Nutrition 1987; 117:1629-1639.
- Koshland Jr DE. The microbial wars. Science 1992; 257(5073):1021.
- Lee JE, Austic RE, Naqi SA, Golemboski KA, Dietert RR. Dietary arginine intake alters avian leukocyte population distribution during infectious bronchitis challenge. Poultry Science 2002; 81:793-798.
- National Research Council (NRC). Nutrient requirements of poultry. 9th ed. Washington: National Academy Press; 1994.
- Rama Rao SV, Praharaj NK, Reddy MR, Panda AK. Interaction between genotype and dietary concentrations of methionine for immune function in commercial broilers. British Poultry Science 2003; 44: 104-112.
- Rubin LL, Ribeiro ALM, Canal CW, Silva IC, Trevizan L, Vogt LK, Pereira RA, Lacerda L. Influence of sulfur amino acid levels in diets of broiler chickens submitted to immune stress. Brazilian Journal of Poultry Science 2007; 9(1):305-311.
- Ruiz-Feria CA, Kidd MT, Wideman Jr RF. Plasma levels of arginine, ornithine, and urea and growth performance of broilers fed supplemental L-arginine during cool temperature exposure. Poultry Science 2001; 80:358-369.
- SAS Institute. SAS user's guide: statistics. Cary, NC; 2001.
- Shapiro SS, Wilk MB. An analysis of variance test for normality (complete sample). Biometrika Great Britain 1965; 52(3):591-611.
- Shini S, Li X, Bryden WL. Methionine requirement and cell-mediated immunity in chicks. British Journal of Nutrition 2005; 94:746-752.
- Swain BK, Johri TS. Effect of supplemental methionine, choline and their combinations on the performance and immune response of broilers. British Poultry Science 2000; 41:83-88.
- Tachibana K, Mukai K, Hiraoka I, Moriguchi S, Takama S, Kishino Y. Evaluation of the effect of arginine-enriched amino acid solution on tumor growth. Journal of Parenteral and Enteral Nutrition 1985; 9(4):428-434.
- Takahashi K, Konashi S, Akiba Y, Horiguchi M. Effects of dietary threonine level on antibody production in growing broilers. Animal Science and Technology 1994; 65:956-960.
- Takahashi K, Konashi S, Akiba Y, Horiguchi M. Effects of marginal excess or deficiency of dietary methionine on antibody production in growing broilers. Animal Science and Technology 1993; 64:13-19.
- Takahashi K, Orta N, Akiba Y. Influence of dietary methionine and cystine on metabolic responses to immunological stress by Escherichia coli lipopolysaccharide injection and mitogenic response in broiler chickens. British Journal of Nutrition 1997; 78: 815-821.
- Tayade C, Jaiswal TN, Mishra SC, Koti M. L-Arginine stimulates immune response in chickens immunized with intermediate plus strain of infectious bursal disease virus. Vaccine 2006a; 24(5):552560.
- Tayade C, Koti M, Mishra SC. L-Arginine stimulates intestinal intraepithelial lymphocyte functions and immune response in chickens orally immunized with live intermediate plus strain of infectious bursal disease vaccine. Vaccine 2006b; 24:5473-5480.
- Tizard IR. Imunologia veterinária. 6th ed. São Paulo: Roca; 2002.
- Tsiagbe VK, Cook ME, Harper AE, Sunde ML. Enhanced immune responses in broiler chick fed methionine supplemented diets. Poultry Science 1987; 66:1147-1154.
- Vieira SL, Lemme A, Goldenberg DB, Brugalli I. Responses of growing broilers to diets with increased sulfur amino acids to lysine ratios at two dietary protein levels. Poultry Science 2004; 83:1307-1313.
- Wu G, Flynn NE, Yan W, Barstow GG Jr. Glutamine metabolism in chick enterocytes: Absence of pyrroline-5-carboxylase synthase and citrulline synthesis. The Biochemical Journal 1995; 306:717-721.
Mail Address:
Publication Dates
-
Publication in this collection
12 Mar 2008 -
Date of issue
Dec 2007
History
-
Accepted
Nov 2007 -
Received
Dec 2006