Resumos
As estimativas de máxima verossimilhança restrita (REML), das variâncias e das covariâncias genéticas aditivas e residuais, do peso ao nascimento e dos pesos ajustados aos 120, 205, 240, 365, 420 e 550 dias de idade foram empregadas para determinar funções de covariâncias (CFs) do crescimento de 41.415 bovinos da raça Tabapuã, nascidos entre 1975 e 1997 e criados em regime de pastagem. A estimação das CFs mostrou-se bastante útil, pois, além de avaliar covariâncias entre qualquer par de idades, a análise das autofunções, associada aos autovalores das matrizes de coeficientes das CFs, revelou que as curvas de crescimento dos animais podem ser rapidamente alteradas pela seleção. Fatores como o estresse provocado pelo desmame, o ganho compensatório e a seleção de animais, nos períodos finais, provocaram várias mudanças na trajetória das (co)variâncias genéticas, fazendo com que apenas as CFs de ordens de ajuste mais complexas estimassem valores mais próximos das estimativas da REML. Entretanto, nessas funções de alta ordem de ajuste, os polinômios de Legendre tenderam a descrever ondulações nas trajetórias das variâncias, nas extremidades do período, o que parece não ter uma razão biológica coerente.
crescimento de bovinos; funções de covariância; parâmetros genéticos; polinômios de Legendre
Restricted maximum likelihood (REML) estimates of additive and residual variances and covariances for birth weight and adjusted weights at 120, 205, 240, 365, 420 e 550 days of age were used to estimate growth covariance functions (CFs) of Tabapuã beef calves. Data were observed on 41,415 animals born from 1975 to 1997 and raised under pasture conditions. Estimation of CFs is a very useful tool to analyze beef cattle growth. It was possible to estimate covariance between any pair of ages and the analyses of eigenfunctions associated with the eigenvalues of coefficients matrix of CFs showed that the growth curves of Tabapuã calves could be easily changed by selection. Weaning stress, compensatory growth and selection of animals in the final period caused changes on (co)variance trajectories. Therefore only CFs of more complex order were able to estimate values near to REML estimates. However, high order Legendre polynomials drew sharp waves on variances trajectories at the period edges, witch does not have a coherently biological reason.
beef cattle growth; covariance functions; genetic parameters; Legendre polynomial
Avaliação do crescimento de bovinos jovens da raça Tabapuã, por meio de análises de funções de covariâncias1 1 Parte da tese de doutorado apresentada à UFV e financiada pela CAPES.
Growth evaluation of young Tabapuã beef cattle by covariance functions analyses
Eduardo Shiguero SakagutiI; Martinho de Almeida e SilvaII; Richard Louis QuaasIII; Elias Nunes MartinsI; Paulo Sávio LopesIV; Luiz Otávio Campos da SilvaV
IProfessor do Depto. de Zootecnia/UEM Maringá, PR 87020-900. E.mail: essakaguti@uem.br
IIProfessor do Depto. de Zootecnia/UFMG Belo Horizonte, MG
IIIProfessor do Dept. of Animal Sceince/Cornell University Ithaca, NY, EUA
IVProfessor do Depto. de Zootecnia/UFV Viçosa, MG
VPesquisador da Embrapa Gado de Corte Campo Grande, MS
RESUMO
As estimativas de máxima verossimilhança restrita (REML), das variâncias e das covariâncias genéticas aditivas e residuais, do peso ao nascimento e dos pesos ajustados aos 120, 205, 240, 365, 420 e 550 dias de idade foram empregadas para determinar funções de covariâncias (CFs) do crescimento de 41.415 bovinos da raça Tabapuã, nascidos entre 1975 e 1997 e criados em regime de pastagem. A estimação das CFs mostrou-se bastante útil, pois, além de avaliar covariâncias entre qualquer par de idades, a análise das autofunções, associada aos autovalores das matrizes de coeficientes das CFs, revelou que as curvas de crescimento dos animais podem ser rapidamente alteradas pela seleção. Fatores como o estresse provocado pelo desmame, o ganho compensatório e a seleção de animais, nos períodos finais, provocaram várias mudanças na trajetória das (co)variâncias genéticas, fazendo com que apenas as CFs de ordens de ajuste mais complexas estimassem valores mais próximos das estimativas da REML. Entretanto, nessas funções de alta ordem de ajuste, os polinômios de Legendre tenderam a descrever ondulações nas trajetórias das variâncias, nas extremidades do período, o que parece não ter uma razão biológica coerente.
Palavras-chave: crescimento de bovinos, funções de covariância, parâmetros genéticos, polinômios de Legendre
ABSTRACT
Restricted maximum likelihood (REML) estimates of additive and residual variances and covariances for birth weight and adjusted weights at 120, 205, 240, 365, 420 e 550 days of age were used to estimate growth covariance functions (CFs) of Tabapuã beef calves. Data were observed on 41,415 animals born from 1975 to 1997 and raised under pasture conditions. Estimation of CFs is a very useful tool to analyze beef cattle growth. It was possible to estimate covariance between any pair of ages and the analyses of eigenfunctions associated with the eigenvalues of coefficients matrix of CFs showed that the growth curves of Tabapuã calves could be easily changed by selection. Weaning stress, compensatory growth and selection of animals in the final period caused changes on (co)variance trajectories. Therefore only CFs of more complex order were able to estimate values near to REML estimates. However, high order Legendre polynomials drew sharp waves on variances trajectories at the period edges, witch does not have a coherently biological reason.
Key Words: beef cattle growth, covariance functions, genetic parameters, Legendre polynomial
Introdução
As principais fontes de informação das avaliações genéticas de gado de corte são os pesos corporais observados ao longo da vida dos animais. Com vistas em padronizar as análises, os programas de melhoramento estabelecem determinadas idades às quais os pesos são ajustados, antes de serem avaliados. Essas idades recebem a denominação de idades-padrão e são adotadas segundo princípios biológicos ou meramente cronológicos. As análises consideram que os pesos ajustados a determinada idade são observações de característica diferente das formadas pelos pesos em outras idades.
O número de dias que determina as idades-padrão tem variado nos programas de melhoramento. Na literatura, podem ser encontradas idades de 200, 205 e 240 dias, para peso ao desmame; de 365 e 400 dias, para peso no ano; de 420, 550 e 600 dias, para peso ao sobreano ou final (e.g., Meyer, 1993; Lobo, 1996; CNPGC, 2000). Essas variações podem ser justificadas pelas diferenças no clima da região de abrangência de cada programa, pelo sistema de manejo adotado pelas fazendas integrantes, pelos objetivos econômicos e nas características fisiológicas de cada raça ou linhagem.
Independente do critério adotado para definição das idades-padrão, a adoção do conceito de característica distinta, para peso a cada idade-padrão, impõe uma classificação descontínua a um conjunto de dados que têm base contínua, a idade (Kirkpatrick & Lofsvold, 1992).
A utilização de técnicas multivariadas em análises conjuntas dos pesos ajustados às idades-padrão fornece informações sobre as relações lineares existentes entre cada par de idades, porém nenhuma inferência pode ser feita a respeito dos pesos nas idades intermediárias. Por outro lado, melhor detalhamento, por meio da inclusão de várias características (i.e., pesos), pode fazer com que os modelos tenham número excessivo de parâmetros.
Utilizado na definição das características que podem ser descritas por funções contínuas, o conceito de "características de dimensão infinita" (Kirkpatrick & Heckman, 1989) pode ser adequadamente aplicado aos pesos dos animais, quando o objeto de estudo é o crescimento em determinado período. As variâncias e covariâncias desse tipo de características também são representadas por funções contínuas, denominadas de Funções de Covariâncias (CFs). As CFs podem ser empregadas para estimar as variâncias e covariâncias em quaisquer das idades abrangidas pelo período considerado. Assim, as CFs são equivalentes às matrizes de (co)variâncias que poderiam ser estimadas em análise multivariada do conjunto formado por todas as idades do período (Kirkpatrick et al., 1990 e 1994). Entretanto, funções contínuas relativamente simples podem ser capazes de representar as (co)variâncias de determinado intervalo, e os modelos empregados teriam número relativamente pequeno de parâmetros.
Teoricamente, a estimação de CFs envolveria operações com matrizes de dimensões infinitas; contudo, Kirkpatrick et al. (1990) utilizaram a metodologia de quadrados mínimos generalizados (GLS) para apresentar um método de estimação de CFs, a partir de estimativas truncadas de matrizes de (co)variâncias de número finito de idades. Neste método, admite-se que as matrizes de (co)variâncias genéticas sejam previamente determinadas por regressão pai-filho ou por covariâncias entre meios-irmãos ou irmãos completos, nos correspondentes delineamentos experimentais.
Considerando-se que, em geral, as estimativas de parâmetros genéticos de gado de leite, obtidas pela metodologia de máxima verossimilhança restrita (REML), sejam próximas das estimativas obtidas de covariâncias entre meias-irmãs, Van der Werf & Schaeffer (1997) sugeriram a aplicação do método, proposto por Kirkpatrick et al. (1990), em dados de campo, adimitindo-se que estes sejam provenientes de um dos três delineamentos citados.
As análises propostas por Kirkpatrick et al. (1990) envolvem operações algébricas relativamente simples, as quais podem ser prontamente executadas por meio dos vários pacotes matemáticos que, atualmente, se encontram disponíveis. Dessa forma, este trabalho objetivou avaliar a aplicação de análises de funções de covariâncias na avaliação genética do crescimento de bovinos jovens, da raça Tabapuã.
Materiais e Métodos
Parâmetros genéticos dos pesos às idades-padrão
As idades-padrão foram escolhidas com base nos programas de melhoramento genético de raças zebuínas existentes no Brasil (e.g., Lobo, 1996; CNPGC, 2000).
Considerando-se que o peso dos animais ao nascimento (PN) e os pesos ajustados aos 120 (P120), 205 (P205), 240 (P240), 365 (P365), 420 (P420) e 550 (P550) dias de idade eram características diferentes, estimaram-se as variâncias e covariâncias genéticas aditivas e residuais por meio do programa REMLF90 (Misztal, 1999), que utiliza a metodologia de máxima verossimilhança restrita (REML) e o algoritmo de Maximização da Esperança (EM), com processo de aceleração da convergência. Admitiu-se que a convergência era atingida quando o quadrado das diferenças relativas entre estimativas consecutivas era menor que 10-9.
Nas análises, utilizaram-se modelos com efeito genético aditivo e efeitos fixos da idade da vaca ao parto, como covariável linear e quadrática, e de grupo contemporâneo, formado pela combinação das classes de sexo do animal, fazenda, ano e estação de nascimento, para todos os pares de pesos.
Os dados usados foram obtidos da Associação Brasileira de Criadores de Zebu (ABCZ) e foram cedidos pelo Centro Nacional de Pesquisa de Gado de Corte - Embrapa, Campo Grande, MS, sendo provenientes de animais da raça Tabapuã, nascidos entre 1975 e 1997 e criados em sistema de pastagem (Tabela 1).
Equivalência entre multicaracterísticas e características de dimensão infinita
Admitindo-se um período com t idades, de forma que o peso em cada idade correspondesse à observação em uma característica diferente, um modelo misto multivariado, que abrangesse todas as t características, poderia ser representado, para cada animal, por:

em que y é vetor com t pesos; b, vetor de efeitos fixos; u e e, vetores de efeitos aleatórios (e.g., genético aditivo e residual, respectivamente), tal que cada elemento corresponda ao respectivo efeito em uma das idades.
Considerando-se a natureza contínua das idades (t = ¥), os elementos de u poderiam ser descritos por função contínua do tipo:

em que F é a matriz (ou vetor) de covariáveis (constantes), e au, um vetor de t coeficientes, ou seja, caso t = 3, os três elementos de u poderiam ser exatamente representados por função quadrática do peso dos animais em relação às três idades, sendo au formado por an0 (constante da regressão), por au1 (coeficiente de regressão linear) e por au2 (coeficiente de regressão quadrático).
Desmembrado-se e, da equação (1), em dois termos independentes, c (e.g., efeito permanente de ambiente) e e (e.g., efeito temporário de ambiente), e admitindo-se que c também possa ser representado por função contínua, tem-se

em que ac é um vetor de coeficientes, relativos a c.
Assim, empregando-se as igualdades (2) e (3), a equação (1) pode ser reescrita da seguinte forma:

Como existem infinitas idades em qualquer intervalo, a equação (4) pode ter número infinito de covariáveis em F e de coeficientes em au e ac.
Na prática, admite-se que um número finito de coeficientes seja suficiente para descrever os efeitos u e c, em todo o período, e que o modelo (4) seja equivalente aos chamados modelos de regressão aleatória (RRM) (Henderson Jr., 1982). Assim, a partir das estimativas de au e ac podem-se estimar os efeitos aleatórios referentes a u e c, para qualquer idade, por:

em que fi(al*) são covariáveis correspondentes à idade al, de acordo com determinado tipo de função contínua (ver na próxima seção), de ordem de ajuste o. Nota-se que uma função contínua, de ordem de ajuste o = 3, corresponde a um polinômio quadrático (segundo grau).
Polinômios de Legendre
Optou-se pela utilização dos polinômios de Legendre para compor as funções contínuas, pois, além de serem normalizados e ortogonais, são também indicados para analisar padrões de variações genéticas em trajetórias de crescimento (Kirkpatrick et al., 1990).
Cada polinômio k foi definido pela seguinte fórmula:

em que ai* = 2(ai - amin)/ (amax - amin)-1, tal que amin e amax são, respectivamente, a primeira (menor) e a última (maior) idade do intervalo considerado, e os colchetes, [.], sobre o somatório indicam que o número em seu interior é arredondado para baixo, admitindo-se o valor inteiro mais próximo.
Em termos matriciais, as covariáveis resultantes dos polinômios de Legendre podem ser calculadas por F = ML (Van der Werf & Schaeffer, 1997), em que M é matriz com t linhas e o colunas, com elementos mij = (ai*)j-1, e L é matriz de ordem o, composta pelos coeficientes polinomiais produzidos pela equação (6). Dessa forma, para o=3, tem-se

Funções de covariância (CFs)
Sendo G e R as matrizes de variâncias e covariâncias de u e e, respectivamente, e empregando-se as igualdades (2) e (3), tem-se que

em que Ku = Var(au); Kc = Var(ac); I é uma matriz identidade de ordem t; e se2 é a variância do ambiente temporário de cada peso observado; Ku e Kc são as matrizes de coeficientes das CFs, e as estimativas dos seus elementos, kuij e kcij, podem ser empregadas para calcular (co)variâncias entre efeitos de u e de c, para qualquer par de idades (al e am), por meio das seguintes funções de covariâncias:

Para o máximo de ajuste, ou seja, quando a ordem de ajuste da função contínua for igual ao número de idades (i.e., o = t), a matriz F é matriz quadrada e tem inversa comum; assim, as matrizes dos coeficientes das CFs podem ser estimadas por

em que o sobrescrito -t significa a inversa da matriz transposta.
Entretanto, o que se pretende, na maioria dos casos, é estimar as CFs, que, além de serem capazes de descrever os valores das covariâncias ao longo das idades, sejam simples o bastante (i.e., o pequeno) para facilitar as análises e as interpretações, principalmente quando t for relativamente grande. Para se estimarem funções de covariâncias de ordem de ajuste reduzida (i.e., o < t), utilizou-se o método proposto por Kirkpatrick et al. (1990), citado, resumidamente, no próximo item.
Em razão da pequena magnitude das variâncias residuais para PN, quando comparada às demais, e do desconhecimento da proporção entre efeitos temporários e permanentes que compõem o resíduo, optou-se pela estimação das CFs dos resíduos, em vez de se estimarem apenas as CFs dos efeitos permanentes de ambiente. Admitiu-se que apenas a diagonal da matriz de covariância seria inflacionada e que a heterocedasticidade dos resíduos também era decorrente da idade do animal. Assim, neste estudo, o modelo (4) e a igualdade (8) foram substituídos por

Estimação de CFs pelo Método de Kirkpatrick et al. (1990).
Considerando-se que g seja um vetor-coluna, de ordem t2, composto pelas estimativas das matrizes de (co)variâncias (e.g., G ou R), empilhado por colunas sucessivas, tal que g' = [G11, , Gt1, G12, , Gt2, , Gtt] ou g' = [R11, , Rt1, R12, , Rt2, , Rtt], admitiu-se o seguinte modelo:

em que ko é vetor-coluna, de ordem o2, composto pelos elementos da matriz de coeficientes das CFss, dispostos da seguinte forma: ko'= [K11, , Ko1, K12, , Ko2, , Koo]; Xo = FoÄFo, que é o produto de Kronecker (Ä) da matriz de covariáveis (Fo), referentes a uma função de ordem o; e do é o vetor de erros de amostragem.
Para aplicar a metodologia dos quadrados mínimos generalizados (GLS), é necessário estimar , que é a matriz de estimativas de covariâncias, referente aos erros nas estimativas de g. Ao se admitir que g tenha sido obtida de delineamento composto por 800 famílias (s = 800 reprodutores) de 50 meios-irmãos (n = 50 filhos por reprodutor), estimaram-se os elementos de por

em que Ma e Me são, respectivamente, produtos cruzados de reprodutores e residuais, estimados por

em que  e
e  são estimativas das covariâncias genéticas e fenotípicas, respectivamente.
são estimativas das covariâncias genéticas e fenotípicas, respectivamente.
As covariâncias entre os produtos cruzados foram obtidas por

Os denominadores dessas duas equações são os graus de liberdade dos respectivos produtos cruzados, adicionados de dois para se obterem estimativas não-tendenciosas de Vij,kl, uma vez que foram empregadas as estimativas Maij e Meij.
Tendo em vista que as matrizes de covariâncias são simétricas, o vetor g contém redundâncias (e.g., Gij = Gji), o que faz com que  seja singular. Assim, adotaram-se as seguintes modificações:
seja singular. Assim, adotaram-se as seguintes modificações:
1. Para todos os Gi ou Ri, em que i < j, excluir as correspondentes linhas em g , e Xo e as correspondentes colunas em .
2. Para todos os Kij, tal que i < j, excluir as correspondentes linhas em ko, e em Xo, somar cada coluna referente a Kij com a coluna referente a Kji, e excluir as colunas referentes a Kij.
Após essas modificações, as funções de covariâncias foram estimadas de acordo com o sistema de equações abaixo:

em que o asterisco (*) indica que as matrizes tiveram suas dimensões reduzidas.
A partir das estimativas de ko*, estimaram-se (co)variâncias genéticas e residuais por  , para qualquer idade no período analisado, ajustando-se apenas os valores em Xo*.
, para qualquer idade no período analisado, ajustando-se apenas os valores em Xo*.
Testes para falta de ajustamento
Para testar a consistência entre os valores de covariâncias, calculados pelas CFs de ordem reduzida ( ), e as covariâncias estimadas para as idades-padrão, pela metodologia REML (
), e as covariâncias estimadas para as idades-padrão, pela metodologia REML ( ), foi empregado o procedimento proposto por Kirkpatrick et al. (1990), que admitiram que a soma de quadrados dos erros de amostragem, de determinado modelo de ordem reduzida, seguiu distribuição de c2, aproximada com m- p graus de liberdade, ou seja,
), foi empregado o procedimento proposto por Kirkpatrick et al. (1990), que admitiram que a soma de quadrados dos erros de amostragem, de determinado modelo de ordem reduzida, seguiu distribuição de c2, aproximada com m- p graus de liberdade, ou seja,

em que m = t(t + 1)/2 e p = o(o + 1)/2.
Foi também empregada a estatística F, proposta por Van der Werf & Schaeffer (1997), para testar a significância do aumento de precisão com adição de um coeficiente de ordem superior, dado pela fórmula:

Consistência das matrizes de covariâncias
As matrizes de covariâncias genéticas (G) e residuais (R), empregadas na estimação das CFs, foram formadas pelas estimativas da REML de covariâncias dos pares de pesos às idades-padrão (fora da diagonal) e pelas médias das estimativas de variâncias (diagonal) das análises bicaracterísticas. A consistência dessas matrizes foi assegurada pela substituição dos autovalores negativos, contidos na diagonal da matriz Di, por 1x10-8 e pela reestruturação das matrizes, da seguinte forma:

em que Di* é matriz diagonal dos novos autovalores (não-negativos); Ai tem as colunas formadas pelos autovetores; e ri é a razão da soma dos autovalores originais pela soma dos novos autovalores.
Autofunções e autovalores
Autofunções são funções contínuas, cujos coeficientes são formados pelos elementos dos autovetores das matrizes de coeficientes das CFs. Para cada autofunção existe um autovalor, que representa a proporção de variação total que ela explica. As trajetórias descritas pelas autofunções podem ser utilizadas na avaliação das idades em que a seleção poderia ser mais eficiente.
Fazendo-se a decomposição da matriz de coeficientes das CFs genéticas (Ku) em uma matriz diagonal (D) de autovalores e em uma matriz (U) cujas colunas são formadas pelos autovetores, tal que Ku = UDU', as autofunções foram obtidas pela multiplicação da matriz de covariáveis F pela matriz de autovetores U, i.e., FU.
Resultados e Discussão
Pesos às idades-padrão
As estimativas EM-REML de (co)variâncias genéticas aditivas e residuais e herdabilidades, para peso no nascimento e pesos ajustados aos 120, 205, 240, 365, 420 e 550 dias de idade, obtidas em 21 análises bicaracterísticas, são apresentadas na Tabela 2. Encontrou-se variabilidade genética entre os pesos dos animais em todas as idades analisadas, o que indica que a seleção pode promover ganhos consideráveis no peso corporal dos animais da raça Tabapuã, em todas as idades-padrão.
A Figura 1 mostra que as taxas de aumento da magnitude das médias das variâncias genéticas aditivas e residuais são semelhantes até os 205 dias, ou seja, enquanto os animais estão sendo amamentados. Após os 205 dias, a variância residual continua aumentando a taxas semelhantes às do período pré-desmame, enquanto a variância genética aditiva reduz a taxa de aumento, entre 240 e 365 dias. Meyer et al. (1993) observaram diminuição de variabilidade genética entre o peso ao desmame (200 dias de idade) e o peso aos 400 dias de idade, em dados de bovinos da raça Polled Hereford e da raça sintética Wolkalup (¼ zebuína e ¾ taurina), criados em condições de pastagem. O ganho compensatório pode ter contribuído para essa redução na variabilidade genética por volta de um ano de idade, ao fazer com que os animais muito leves, por ocasião do desmame, tivessem ganho pós-desmame superior à média de seus rebanhos, provocando certa uniformização dos pesos, por volta do primeiro ano de idade.
Embora a variância genética tenha tido considerável aumento, dos 420 aos 550 dias de idade, as estimativas de herdabilidades aos 550 dias foram semelhantes aos valores encontrados para as idades de 365 e 420 dias, sendo que a herdabilidade média (Tabela 3), aos 550 dias, foi a menor de todas as idades consideradas. A menor herdabilidade da última idade considerada pode ser atribuída ao menor número de observações nessa idade. É bastante provável que exista tendência de eliminação dos animais que apresentaram baixo desempenho nas idades mais novas.
Quanto mais próximas são as idades, maiores são as correlações genéticas e fenotípicas (Tabela 3), visto que o distanciamento entre as idades provoca maior queda na correlação fenotípica que na genética.
Os valores de correlação genética entre o peso ao nascer e as demais idades são relativamente baixos, sendo a maior correlação (0,44) observada no peso aos 550 dias de idade, e as menores (0,36), nos pesos próximos ao desmame (205 e 240 dias de idade).
Embora se possa esperar que animais mais pesados ao nascer mantenham essa vantagem nas idades posteriores, quando a seleção é praticada, o peso excessivo ao nascer pode comprometer o desenvolvimento pós-natal. Bezerros muito pesados podem sofrer maior estresse em situações de restrição de alimentos, quando comparados com animais mais leves, que apresentam menor exigência nutricional. As vacas zebuínas de corte, em geral, não são boas produtoras de leite e são criadas em regime extensivo de pastagem, estando sujeitas a períodos de escassez de alimento. Além disso, o desgaste provocado pela gestação e pelo aleitamento de bezerros muito pesados pode dificultar a retomada do ciclo reprodutivo, o que não é economicamente interessante para os criadores.
Altas correlações genéticas aditivas (>0,80) foram encontradas entre pesos em idades posteriores ao nascimento, sendo as correlações próximas à unidade quando os pesos foram separados por menos de três meses. Essa expressiva relação linear entre os pesos após o nascimento indica que a seleção, para maior peso nas primeiras idades, tende a aumentar o peso nas idades mais avançadas e vice-versa.
Na pecuária de corte, questionamentos sobre a eficiência econômica do sistema de produção com animais de tamanho adulto excessivo são antigos. Entretanto, até recentemente, na avaliação genética de bovinos da raça Tabapuã, consideravam-se apenas os pesos ajustados às idades-padrão (Josahkian, 2000).
Funções de covariâncias
Com a estimação das funções de covariâncias dos efeitos genéticos aditivos e residuais, há mudança do panorama descontínuo das estimativas de variâncias e covariâncias nas idades-padrão para uma superfície contínua de covariâncias (Figura 2), o que permite que se estimem as covariâncias entre os pesos corporais de qualquer par de idades abrangidas pelo intervalo estudado.
A utilização de polinômios ortogonais é apenas uma das alternativas de estimação das funções de covariâncias a partir de matrizes de (co)variâncias (e.g., G e P). Independentemente do tipo de função adotado, quando se empregam funções com o máximo de ajuste (o = t), os valores estimados pelas CFs, para os pontos da matriz original (i.e., as (co)variâncias das idades que compõem G), são sempre iguais aos valores originais. Entretanto, a escolha do tipo de polinômio influencia os valores das idades intermediárias, calculados por interpolação, e as estimativas obtidas por CFs de ordem de ajuste reduzida (Kirkpatrick et al., 1990).
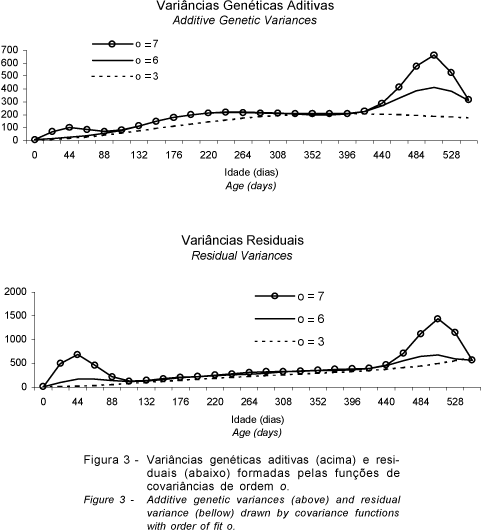
Apesar de reproduzirem as covariâncias entre as idades-padrão, as CFs de máxima ordem de ajuste (o = 7) desenharam várias ondulações na superfície de (co)variâncias, o que dificultou uma interpretação biológica do crescimento dos animais.
Os modelos de ordens de ajuste mais baixos (o < 6) tenderam a subestimar as (co)variâncias genéticas dos pesos dos animais nas idades mais avançadas (P420 e P550). A superfície descrita pelo modelo de ordem de ajuste igual a seis percorre pontos próximos aos valores estimados para as idades-padrão, e suas ondulações parecem ter maior coerência com o comportamento do crescimento.
Na Tabela 4 encontram-se as estimativas de somas de quadrados e graus de liberdade dos erros de amostragem, empregados nos testes estatísticos que avaliaram o ajustamento dos modelos de CFs de ordem de ajuste reduzida. Todas as estimativas de covariâncias genéticas e residuais, obtidas a partir das CFs de ordens reduzidas, foram significativamente diferentes (P<0,01) das respectivas estimativas EM-REML, pelo teste de qui-quadrado.
A necessidade de polinômio de alta ordem de ajuste pode ser atribuída à grande extensão do período avaliado, no qual houve fases de crescimento distintas que fizeram com que as covariâncias, principalmente genéticas, tivessem várias mudanças de trajetórias. Segundo Van der Werf & Schaeffer (1997), ao se subestimarem os valores de V, admite-se que as estimativas de covariâncias, empregadas na estimação das CFs, sejam mais acuradas que a realidade, o que torna mais difícil encontrar um modelo de ordem de ajuste reduzido que seja suficiente.
Pelo teste F, constatou-se que apenas as inclusões do terceiro (o = 3) e do sexto (o = 6) polinômios de Legendre produziram aumento significativo na precisão dos modelos de CFs genéticas, enquanto todas as inclusões de polinômios adicionais foram significativas nos modelos de CFs residuais.
Na Figura 3, estão representadas as variâncias genéticas e residuais estimadas pelas CFs, de ordens iguais a 3, 6 e 7. Observa-se que as CFs, de ordens mais altas, tendem a descrever picos nas extremidades, o que não apresenta justificativa biológica. Foram também encontrados problemas relativos à estimação de parâmetros genéticos nas extremidades do período de lactação, nos trabalhos de Jamrozik & Schaeffer (1997), Kettunen et al. (1998), Van der Werf et al. (1998) e Strabel & Misztal (1999), que empregaram os modelos de regressão aleatória para estimar CFs em características de produção de leite. Nos dois primeiros trabalhos, foi admitido que apenas as (co)variâncias genéticas variavam em decorrência do tempo.
Para Van der Werf et al. (1998), o comportamento anômalo das CFs, no início e no final da lactação, pode ter sido causado pela falta de um modelo correto para todos os componentes de variância e pela utilização de polinômios, que podem não se comportar bem nas periferias. Valores distantes das médias exercem grande influência em uma análise de regressão (Meyer, 1999). O menor número de observações no final da lactação foi outra possível causa de viés, segundo Strabel & Misztal (1999), que obtiveram estimativas mais plausíveis, quando foram realizadas análises conjuntas de duas lactações.
Neste trabalho, o comportamento estranho das CFs, nas idades extremas, pode ser atribuído à não-inclusão do efeito genético materno nos modelos que estimaram as (co)variâncias nas idades-padrão, principalmente nas primeiras idades, enquanto a seleção de animais pode ter influenciado as estimativas referentes às últimas idades. Entretanto, Meyer (1999) eliminou as observações das últimas idades e continuou observando comportamento distorcido das CFs, nas últimas idades. Segundo essa autora, os problemas encontrados eram também atribuídos à ordem de ajuste das CFs.
Nas idades intermediárias (dos 100 a 420 dias de idade), os valores das variâncias genéticas e residuais, estimados pelos modelos de ordem de ajuste igual a seis, são muito semelhantes às estimativas do modelo de máxima ordem de ajuste, sendo as estimativas de todas as idades-padrão muito próximas nos dois modelos. Com exceção da ondulação na fase final, a trajetória descrita pelas variâncias genéticas, estimadas pelo modelo de ordem de ajuste igual a seis, parece ser biologicamente mais coerente que a trajetória descrita pelo modelo de máxima ordem de ajuste.
Os modelos de regressão quadrática (o = 3) subestimaram as variâncias genéticas e residuais em quase todo o período estudado, e, nas idades próximas ao desmame e no período final, as diferenças entre as estimativas de variância genética dos modelos de ordem superior foram mais acentuadas. Entretanto, a ausência de ondulações nas extremidades faz com que o comportamento da trajetória das variâncias residuais, do modelo quadrático, pareça ser biologicamente mais apropriado.
Ao se analisarem os autovalores das matrizes de coeficientes das CFs de ordens de ajuste iguais a 3, 6 e 7 (Tabela 5), observa-se que o primeiro autovalor é sempre dominante sobre os demais, sendo equivalentes a 97, 88 e 78% da variação genética total dos modelos de o = 3, 6 e 7, respectivamente. Uma porção considerável (10 e 17%) da variação genética dos modelos de o = 6 e 7, respectivamente, é atribuída ao segundo autovalor. Entretanto, no modelo quadrático, o segundo autovalor representou menos que 0,3% da variação genética.
A grande dominância dos primeiros autovalores também foi observada por Meyer (1999), ao analisar peso adulto (de 2 a 10 anos de idade) de vacas de corte das raças Polled Hereford e Wokalup, e por Koenen & Veerkamp (1998), ao analisarem o peso corporal, durante o período de lactação (25 semanas), de vacas da raça Holandesa. Entretanto, em ambos os trabalhos, as autofunções, associadas ao primeiro autovalor de cada modelo, descreveram curvas de comportamento bem menos oscilatórias que as encontradas neste trabalho (Figura 4).
A utilização de pesos de animais mais jovens, durante o período em que a seleção foi praticada, pode ter sido a causa da maior oscilação das autofunções deste trabalho. Por estarem associadas às autofunções que explicam grande proporção da variância genética, essas oscilações indicam que a curva de crescimento (até 2 anos de idade) dos animais da raça Tabapuã pode ser rapidamente alterada pela seleção.
Trajetórias planas descritas pelas autofunções associadas aos primeiros autovetores, como as encontradas nos trabalhos de Kirkpatrick et al. (1990) e Koenen & Veerkamp (1998), indicam que as curvas de crescimento, nos períodos estudados, estão pouco sujeitas às alterações pela seleção.
Conclusões
As variabilidades genéticas, encontradas em todas as idades-padrão, indicaram a possibilidade de alcançar respostas pela seleção de animais, em todo o período considerado.
A estimação das CFs mostrou-se útil na avaliação do crescimento de bovinos, pois, além de determinar covariâncias entre qualquer par de idades, a análise das autofunções, associada aos autovalores das matrizes de coeficientes das CFs, revelou que a curva de crescimento dos dois primeiros anos de vida pode ser rapidamente alterada pela seleção.
A segmentação, em períodos mais curtos que o estudado neste trabalho (2 anos), pode ser empregada para se tentar evitar a utilização de polinômios de alta ordem de ajuste, os quais podem descrever trajetórias de difícil interpretação biológica.
Agradecimento
Ao professor Ignacy Misztal e ao pesquisador Shogo Tsuruta, da University of Georgia, EUA, por viabilizarem a utilização do programa REMLF90, ao professor Julius Van der Werf, da University of New England, Austrália, e ao pesquisador Roberto A. A. Torres Jr., da Embrapa gado de Corte, pelas valiosas colaborações prestadas.
Literatura Citada
Recebido em: 13/05/02
Aceito em: 20/11/02
- CENTRO NACIONAL DE PESQUISA EM GADO DE CORTE - CNPGC. Sumário das raças zebuínas de corte 1999 MA/ABCZ/EMBRAPA. http://www.cnpgc.embrapa.br/~locs/sumario Acessado em 12/03/2000.
- HENDERSON JR., C.R. Analysis of covariance in the mixed model: higher-level, nonhomogeneous, and random regressions. Biometrics, v.38, n.9, p.623-640, 1982.
- JAMROZIK, J.; SCHAEFFER, L.R. Estimates of genetic parameters for a test day model with random regressions for yield traits of first lactation Holsteins. Journal of Dairy Science, v.80, n.4, p.762-770, 1997.
- JOSAHKIAN, L.A. Programa de melhoramento genético das raças zebuínas. In: SIMPÓSIO NACIONAL DE MELHORAMENTO ANIMAL, 3, 2000, Belo Horizonte. Anais Belo Horizonte: SBMA, 2000, p.76-93.
- KETTUNEN, A.; MÄNTYSAARI, E.A.; STRANDÉN, I. et al. Estimation of genetic parameters for first lactation test day milk production using random regression models. In: WORLD CONGRESS ON GENETICS APPLIED TO LIVESTOCK PRODUCTION, 6., 1998, Armidale. Proceedings... Armidale: AGBU-UNE, 1998. p.307-310.
- KIRKPATRICK, M.; HECKMAN, N. A quantitative genetic model for growth, shape, reaction norms, and other infinite-dimensional characters. Journal of Mathematical Biology, v.27, n.5, p.429-450, 1989.
- KIRKPATRICK, M.; HILL, W.G.; THOMPSON, R. Estimating the covariance structure of traits during growth and ageing, illustrated with lactation in dairy cattle. Genetic Research, v.64, n.1, p.57-69, 1994.
- KIRKPATRICK, M.; LOFSVOLD, D. Measuring selection and constraint in the evolution of growth. Evolution, v.46, n.4, p.954-971, 1992.
- KIRKPATRICK, M.; LOFSVOLD, D.; BULMER, M. Analysis of the inheritance, selection and evolution of growth trajectories. Genetics, v.124, n.2, p.979-993, 1990.
- KOENEN, E.P.C.; VEERKAMP, R.F. Genetic covariance functions for live weight, condition score, and dry-matter intake measured at different lactation stages of Holstein Friesian heifers. Livestock Production Science, v.57, n.1, p.67-77, 1998.
- LOBO, R.B. Programa de melhoramento genético da raça Nelore PMGRN 3.ed., Ribeirão Preto: Universidade de São Paulo, 1996. 88p.
- MEYER, K. Estimates of covariance components for growth traits of Australian Charolais cattle. Australian Journal of Agricultural Research, v.44, n.6, p.1501-1508, 1993.
- MEYER, K. Estimates of genetic and phenotypic covariance functions for postweaning growth and mature weight of beef cows. Journal of Animal Breeding and Genetics, v.116, n.3, p.181-203, 1999.
- MEYER, K.; CARRICK, M.J.; DONNELLY, B.J.P. Genetic Parameters for growth traits of Australian beef cattle from a multibreed selection experiment. Journal of Animal Science, v.71, n.10, p.2614-2622, 1993.
- MISZTAL, I. REMLF90 Manual. ftp:/nce.ads.uga.edu/pub/ignacy/blupf90/. 1999. (Acessado em janeiro de 1999).
- STRABEL. T.; MISZTAL, I. Genetic parameters for first and second lactation milk yields of Polish Black and White cattle with random regression test-day models. Journal of Dairy Science, v.82, n.12, p.2805-2810, 1999.
- Van der WERF, J.H.J.; GODDARD, M.E.; MEYER, K. The use of covariance functions and random regressions for genetic evaluation of milk production based on test day records. Journal of Dairy Science, v.81, n.12, p.3300-3308, 1998.
- Van der WERF, J.H.J.; SCHAEFFER, L. Random regression in animal breeding. CGIL Guelph, Canada, 1997. (Course Notes).
Datas de Publicação
-
Publicação nesta coleção
10 Dez 2003 -
Data do Fascículo
Ago 2003
Histórico
-
Aceito
20 Nov 2002 -
Recebido
13 Maio 2002