RESUMO
Os atributos da microbiota do solo são importantes indicadores de alterações causadas por práticas agrícolas. Foi conduzido um experimento em condição de campo, em um delineamento de blocos casualizados com quatro repetições, com o objetivo de avaliar os efeitos da forma de manejo (sistema de semeadura direta - SSD e sistema de plantio convencional - SPC) e da fase de condução da cultura do arroz de terras altas (pré-plantio - PP, floração - FA e pós-colheita - PC) sobre atributos da biomassa microbiana do solo (Cmic, Nmic, RBS, qCO2), atividade enzimática (AET, Aβ-G e AFA) e população de microrganismos (bactérias, fungos, actinobactérias e diazotróficos) em amostras de solo coletadas na profundidade de 0-20 cm, tendo como referência uma área de mata, próxima ao experimento. Os sistemas de cultivo provocaram alterações nos atributos microbiológicos do solo quando comparados com a mata, usada como referência. Foi observado diferença entre os sistemas de manejo para Cmic, RBS e qCO2, sendo que sob SSD os valores de Cmic e qCO2 foram próximos aos observados para a mata. A atividade enzimática do solo não detectou diferença entre SSD e SPD; contudo, foi observado maior AFA no PP. No geral, o número de microrganismos foi maior sob SSD e SPC, comparados à mata, exceto para fungos. As maiores contagens de microrganismos foram observadas no PP, exceto para os actinomicetos cujo maior valor foi observado na PC.
Palavras-chave:
Oryza sativa; Manejo do solo; Qualidade do solo; Contagem de microrganismos
ABSTRACT
The attributes of soil microbiota are important indicators of the changes caused by agricultural practices. An experiment was carried out under field conditions in a randomized block design with four replications, with the aim of evaluating the effects of the type of management (a system no tillage - SSD and of conventional tillage - SPC) and the growth stage of upland rice (pre-plant - PP, flowering - FA and postharvest - PC) on attributes of the soil microbial biomass (Cmic, Nmic, BSR, qCO2), enzyme activity (TEA, Aß-G and APA) and population of microorganisms (bacteria, fungi, actinobacteria and diazotrophs) in soil samples collected at a depth of 0-20 cm. An area of forest near the experiment was taken as reference. The management systems caused changes in the microbiological attributes of the soil when compared to the reference forest. A difference was found between the management systems for Cmic, BSR and qCO2; under SSD, the values for Cmic and qCO2 were close to those observed for the forest. No difference was detected between SSD and SPD in the enzyme activity of the soil; however, greater values were found for APA at the PP stage. Overall, the number of microorganisms was higher under SSD and SPC compared to the forest, except for fungi. The greatest counts of microorganisms were found at the PP stage, except for actinobacteria, for which the highest value was found at the PC stage.
Key words:
Oryza sativa; Soil management; Soil quality; Microorganism counts
INTRODUÇÃO
A adoção de sistemas sustentáveis de produção, como os sistemas agroecológicos, vem aumentando consideravelmente nos últimos anos, impulsionada principalmente pela demanda da sociedade por alimentos que apresentem maior qualidade e que, em seu processo produtivo, resultem em menores impactos ambientais. Como reflexo, o mercado de produtos orgânicos cresce de 20 a 30% ao ano, tendo movimentado em 2014 R$ 2 bilhões, com projeção de R$ 2,5 bilhões para o ano de 2016 (BRASIL, 2016BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Mercado brasileiro de orgânicos deve movimentar R$ 2,5 bi em 2016. Disponível em: < http://www.agricultura.gov.br/comunicacao/noticias/2015/09/mercado-brasileiro-de-organicos-deve-movimentar-rs-2-bi-em-2016>. Acesso em: 08 Mar. 2016.
http://www.agricultura.gov.br/comunicaca...
). Além do aumento do número de produtores e área cultivada, a agregação de valor a esse produto, decorrente de um mercado com comercialização diferenciada e da sua forma de produção, contribuem para o bom desempenho desse mercado.
Nesses sistemas de produção adota-se um conjunto de práticas, entre elas a semeadura direta, procurando preparar e cultivar o solo de maneira a minimizar a destruição dos agregados e a inversão das camadas de solo, diferente do que ocorre, em geral, no preparo convencional (VEZZANI; MIELNICZUK, 2009VEZZANI, F. M.; MIELNICZUK, J. Uma visão sobre qualidade do solo. Revista Brasileira de Ciência do Solo, v. 33, n. 4 p.743-755, 2009.). A principal contribuição da semeadura direta está associada à adição de matéria orgânica, a qual tem sido relacionada à melhoria das propriedades edáficas do solo (CARDOZO et al., 2008CARDOZO, S. V. et al. Caracterização de propriedades edáficas em áreas sob manejo orgânico e natural na região serrana do Estado do Rio de Janeiro. Semina: Ciências Agrárias, v. 29, n. 3, p. 517-530, 2008.), às alterações na composição da comunidade microbiana e na atividade enzimática do solo, resultando em impactos benéficos nos atributos microbianos do solo (ELFSTRAND; HEDLUND; MARTENSSON, 2007ELFSTRAND, S.; HEDLUND, K.; MÅRTENSSON, A. Soil enzyme activities, microbial community composition and function after 47 years of continuous green manuring. Applied Soil Ecology, v. 35, n. 3, p. 610-621, 2007.; ELFSTRAND; BATH; MARTENSSON, 2007ELFSTRAND, S.; BÅTH, B.; MÅRTENSSON, A. Influence of various forms of green manure amendment on soil microbial community composition, enzyme activity and nutrient levels in leek. Applied Soil Ecology, v. 36, n. 1, p. 610-621, 2007.).
Os atributos microbianos do solo, tais como a diversidade de microrganismos, atividade enzimática, taxa de respiração e biomassa microbiana, são indicadores sensíveis que podem ser utilizados no monitoramento de alterações ambientais decorrentes do uso agrícola (EPELDE et al., 2014EPELDE, L. et al. Microbial properties and attributes of ecological relevance for soil quality monitoring during a chemical stabilization field study. Applied Soil Ecology, v. 75, p. 1-12, 2014.; FERREIRA et al., 2010FERREIRA, E. P. B. et al. Microbial soil quality indicators under different crop rotations and tillage management. Revista Ciência Agronômica, v. 41, n. 2, p. 177-183. 2010.; FERREIRA; WENDLAND; DIDONET, 2011FERREIRA, E. P. B.; WENDLAND, A.; DIDONET, A. D. Microbial biomass and enzyme activity of a Cerrado Oxisol under agroecological production system. Bragantia, v. 70, n. 4, p. 1-9, 2011.). Esses indicadores, além de serem úteis para o monitoramento, também podem servir para orientar o planejamento e a avaliação das práticas de manejo utilizadas.
A microbiota do solo é a principal responsável pela decomposição dos resíduos orgânicos, pela ciclagem de nutrientes e pelo fluxo de energia no solo, podendo ter influência tanto na transformação da matéria orgânica, quanto na estocagem do carbono e nutrientes minerais (JENKINSON; LADD, 1981JENKINSON, D. S.; LADD, J. N. Microbial biomass in soil: measurement and turnover. In: PAUL, E. A.; LADD, J. N. (Ed.). Soil Biology and Biochemistry, v. 5, p. 415-471, 1981.). Entretanto, somente a quantificação da biomassa não fornece indicações sobre os níveis de atividade das populações microbianas do solo, sendo importante também avaliar parâmetros que estimem a atividade microbiana, como a respiração microbiana e atividade enzimática, para verificar o estado metabólico das comunidades de microrganismos do solo (BOWLES et al., 2014BOWLES, T. et al. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biology and Biochemistry, v. 68, p. 252-262, 2014.), bem como quantificar grupos de microrganismos que desempenham papel chave na dinâmica da matéria orgânica do solo.
A literatura apresenta informações sobre o impacto de sistemas de produção e de plantas de cobertura sobre indicadores microbianos da qualidade do solo (SANTOS et al., 2007SANTOS, T. E. B. et al. Variáveis microbiológicas e produtividade do arroz sob diferentes manejos do solo e água. Acta Scientiarum. Agrononomy, v. 29, n. 03, p. 355-366, 2007.; SOUZA et al., 2006SOUZA, L. M. et al. Influência da aplicação de diferentes vermicompostos na biomassa microbiana do solo após cultivo de alface. Revista Brasileira de Agrociência, v. 12, n. 4, p. 429-434, 2006.). Nesses estudos, áreas cultivadas têm sido referendadas a áreas de mata/floresta localizadas próximas, que são consideradas áreas onde os indicadores de qualidade do solo estão submetidos a condições de baixo estresse (FERREIRA et al., 2010FERREIRA, E. P. B. et al. Microbial soil quality indicators under different crop rotations and tillage management. Revista Ciência Agronômica, v. 41, n. 2, p. 177-183. 2010.; FERREIRA; WENDLAND; DIDONET, 2011FERREIRA, E. P. B.; WENDLAND, A.; DIDONET, A. D. Microbial biomass and enzyme activity of a Cerrado Oxisol under agroecological production system. Bragantia, v. 70, n. 4, p. 1-9, 2011.). Embora bastante explorados nos mais diversos sistemas de produção, dados sobre esses indicadores ainda são escassos em sistemas de produção agroecológicos, principalmente no Cerrado brasileiro. Dessa forma, o objetivo desse trabalho foi avaliar o efeito de sistemas de manejo do solo e época de amostragem nos atributos microbianos do solo cultivado com arroz em sistema agroecológico de produção.
MATERIAL E MÉTODOS
O experimento foi conduzido na Estação Experimental em Agroecologia, localizada na Embrapa Arroz e Feijão, em Santo Antônio de Goiás, GO, cujas coordenadas geográficas são: latitude 16º29'30,18" S, longitude 46º17'33,62" W e altitude de 786 m. O clima, conforme classificação de Köppen, é Aw, tropical de savana, megatérmico. O regime pluvial é bem definido, com precipitação pluviométrica média anual de 1.498 mm, ocorrendo cerca de 90% da precipitação no período de outubro a abril e o período seco ocorre de maio a setembro (SILVA et al., 2014SILVA, S. C. da et al. Informações meteorológicas para pesquisa e planejamento agrícola, referentes ao município de Santo Antônio de Goiás, GO, 2012. Santo Antônio de Goiás: Embrapa Arroz e Feijão, 2014. 29 p. (Embrapa Arroz e Feijão. Documentos, 298).).
O solo do local é um Latossolo Vermelho distrófico (EMBRAPA, 2013EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Sistema brasileiro de classificação de solos. 3. ed. Brasília, 2013. 353 p.), de textura franco argilosa, com 410 g kg-1 de areia, 270 g kg-1 de silte e 320 g kg-1 de argila. O pH foi determinado em água. O fósforo e o potássio foram extraídos com a solução de Mehlich 1 (HCl a 0,5 N + H2SO4 a 0,025N) e determinados, respectivamente, em colorímetro e fotômetro de chama. O Ca2+ e o Mg2+ foram extraídos em KCl a 1N e determinados por titulação de EDTA. A matéria orgânica foi determinada pelo método de Walkley e Black (1934)WALKLEY, A.; BLACK, I. A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Science, v. 37, n. 01, p. 29-38, 1934.. As análises laboratoriais foram realizadas de acordo com EMBRAPA (1997)EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Centro Nacional de Pesquisa de Solos. Manual de métodos de análise de solos. 2. ed. Rio de Janeiro: Embrapa CNPS, 1997. 212 p. (Embrapa CNPS. Documentos, 1).. Os atributos químicos do solo nas áreas avaliadas estão apresentados na Tabela 1.
Atributos químicos do solo sob mata e sistemas de semeadura direta (SSD) e sistema de plantio convencional (SPC)
A vegetação original da área experimental era do tipo Cerradão e, após retirada da vegetação natural, a área foi cultivada no sistema convencional de preparo do solo (gradagens aradora e niveladora) com a sucessão milho e soja até a implantação do sistema agroecológico de produção, em 2007. O sistema de produção agroecológico consistiu do uso de leguminosas como plantas de cobertura, principalmente para o fornecimento de nitrogênio (N), e uso de insumos para o controle de pragas e doenças e que foram regulamentados pela Lei 10.831 de 23 de dezembro de 2003 e pelo Decreto nº 6.323 de 27 de dezembro de 2007.
O experimento foi conduzido de 2007 a 2012 com as plantas de cobertura cultivadas de outubro a dezembro de cada ano e a cultura do arroz de terras altas, cultivar Aimoré, conduzida no sistema de semeadura direta (SSD) e em sistema de plantio convencional (SPC) de outubro a dezembro de cada ano. A cultura do arroz foi semeada com espaçamento de 0,45 m entre linhas e com 60 sementes por metro. O experimento foi instalado em delineamento de blocos casualizados com arranjo fatorial (2 x 3) dos tratamentos (dois sistemas de manejo do solo: SSD e SPC e três épocas de coleta, com 4 repetições). As parcelas apresentavam 75 m2 de área útil, deixando como bordadura as duas linhas laterais e um metro em cada cabeceira das parcelas.
No último ano de condução do experimento (2012), amostras de solo foram coletadas com trado na profundidade de 0 a 20 cm nas épocas de pré-plantio, floração e pós-colheita do arroz. Além da coleta, nas parcelas experimentais, foi coletado solo em uma área de mata nativa próxima a área experimental, localizada há cerca de 700 m da área experimental (16º29'52,89" S; 49º17'52,56" W).
Os valores diários de precipitação pluviométrica e temperatura média durante a condução do ensaio são apresentados na Figura 1 (SILVA et al., 2014SILVA, S. C. da et al. Informações meteorológicas para pesquisa e planejamento agrícola, referentes ao município de Santo Antônio de Goiás, GO, 2012. Santo Antônio de Goiás: Embrapa Arroz e Feijão, 2014. 29 p. (Embrapa Arroz e Feijão. Documentos, 298).).
Precipitação, temperatura média diária e épocas de coleta de amostras de solo durante o período de avaliação dos atributos microbianos (janeiro a março de 2012)
As amostras de solo, compostas por três subamostras, foram passadas em peneira com malha de 2 mm de abertura sendo removidos os restos vegetais e os fragmentos de raízes. Foram pesados 20 g de solo de cada amostra, em triplicata, para fumigação e não-fumigação. Os teores de umidade dessas amostras foram ajustados para 80% da capacidade de campo, obtendo-se equilíbrio com a tensão de 30 kPa.
Para a extração do carbono e nitrogênio da biomassa (Cmic e Nmic) foram adicionados 60 mL de uma solução 0,5 M de K2SO4 às amostras que foram submetidas à agitação (40 min; 150 rpm), seguida de decantação por 30 minutos. Em seguida, as amostras foram filtradas em papel filtro para a separação do extrato que foi utilizado na determinação do Cmic e Nmic.
A determinação do Cmic foi realizada pelo método da fumigação-extração, conforme Vance, Brookes e Jenkinson (1987)VANCE, E. D.; BROOKES, P. C.; JENKINSON, D. S. An extraction method for measuring soil microbial biomass C. Soil Biology and Biochemistry, v. 19, n. 6, p. 703-707, 1987., utilizando-se o fator de correção (kC) de 0,33 preconizado por Sparling e West (1988)SPARLING, G. P.; WEST, A. W. A direct extraction method to estimate soil microbial C: calibration in situ using microbial respiration and 14C labelled cells. Soil Biology and Biochemistry, v. 20, n. 3, p. 337-343, 1988., a fim de expressar a fração do Cmic recuperado após o processo de fumigação-extração. A determinação do Nmic foi feita por meio da digestão sulfúrica seguida de destilação Kjeldhal, conforme descrito por Tedesco et al. (1995)TEDESCO, M. J. et al. Análises de solo, plantas e outros materiais. 2. ed. Porto Alegre: Universidade Federal do Rio Grande do Sul, 1995. 174 p., utilizando-se o fator de correção (kN) de 0,54 sugerido por Brookes et al. (1985)BROOKES, P. C. et al. Chloroform fumigation and the release of soil nitrogen: a rapid direct extration method to measure soil microbial biomass nitrogen in soil. Soil Biology and Biochemistry, v. 17, n. 6, p. 837-842, 1985..
A respiração basal do solo (RBS) foi determinada segundo Jenkinson e Powlson (1976)JENKINSON, D. S.; POWLSON, D. S. The effects of biocidal treatments on metabolism in soil: a method for measuring soil biomass. Soil Biology and Biochemistry, v. 8, n. 3, p. 209-213, 1976., em que 20 g de cada amostra de solo foram incubadas (168 h; 120 ºC) à temperatura ambiente em frascos de 0,5 litros hermeticamente fechados e o CO2 foi capturado em frascos de 100 mL contendo 10 mL de uma solução de KOH (0,3 M). Após a incubação, foram adicionados 3 ml de BaCl2 (20%) e o excesso de KOH foi determinado por titulação com solução de HCl (0,1 mol L-1). A avaliação do quociente metabólico (qCO2) foi realizada seguindo o procedimento descrito por Anderson e Domsch (1990)ANDERSON, T. H.; DOMSCH, K. H. Application of eco-physiological quotients (qCO2 and qD) on microbial biomasses from soils of different cropping histories. Soil Biology and Biochemistry, v. 22, n. 2, p. 251-255, 1990., obtido da razão entre o C respirado por unidade de C microbiano por tempo.
A avaliação da atividade enzimática total (AET) do solo foi realizada pelo método da hidrólise do diacetato de fluoresceína, proposto por Ghini, Mendes e Bettiol (1998)GHINI, R.; MENDES, M. D. L.; BETTIOL, W. Métodos de hidrólise de diacetato de fluoresceína (FDA) como indicador da atividade microbiana no solo e supressividade a Rhizoctonia solaniSumma Phytopathologica, v. 24, n. 3, p. 239-242, 1998.. As atividades de β-glicosidase (Aβ-G) e da fosfatase ácida (AFA) foram determinadas de acordo com Tabatabai (1994)TABATABAI, M. A. Soil enzymes. In: WEAVER, R. W.; SCOTT, A.; BOTTOMELEY, P. J. (Ed. Methods of soil analysis: microbiological and biochemical properties. Madison: Soil Science Society of America, 1994. p. 778-835. (Special Publication, 5)., usando a determinação colorométrica do p-nitrofenol liberado pelas enzimas quando as amostras de solo foram incubadas na presença de seus substratos específicos, p-nitrofenol-β-D-glicopironosideo e p-nitrofenol-fosfato, respectivamente.
A quantificação dos microrganismos cultiváveis do solo foi feita pelo método de diluição seriada e espalhamento em placa com meio de cultura sólido ou inoculação em frascos de penicilina com meio semisólido. Na contagem de fungos utilizou-se o meio BDA (batata, dextrose, ágar) com antibiótico (150 µg mL-1 de ampicilina e 100 µg mL-1 de tetraciclina) e para actinobactérias foi utilizado meio ISP2 com antifúngico (150 µg mL-1 de cetoconazol). Para a contagem de bactérias foi utilizado o meio NA (nutriente ágar) também com antifúngico (150 µg mL-1 de cetoconazol) e para bactérias diazotróficas associativas o meio Nfb semi-sólido com cantifúngico (150 µg mL-1 de cetoconazol). As placas foram incubadas a 28 ºC por três dias para as bactérias e sete dias para actinobactérias e fungos. A partir da visualização do crescimento microbiano, as colônias de cada grupo foram contadas. Para as bactérias dizotróficas associativas foi utilizada a técnica do número mais provável. Os resultados foram expressos em unidades formadoras de colônias por grama de solo (UFC g-1 solo).
Os dados foram submetidos à análise de variância e as médias comparadas pelo teste de Tukey a 5% de significância. Adicionalmente, efetuou-se a análise de agrupamento pelo método de Ward, utilizando como medida de dissimilaridade a distância euclidiana média. As análises foram feitas com o auxílio do Programa R (R DEVELOPMENT CORE TEAM, 2011R DEVELOPMENT CORE TEAM. R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing, 2011. Disponível em <http://www.R-project.org>. Acesso em: 25 jan. 2013.
http://www.R-project.org...
).
RESULTADOS E DISCUSSÃO
Valores mais elevados do carbono da biomassa (Cmic) foram observados em amostra de solo de mata, em todas as épocas de amostragem, exceto na floração do arroz, quando não foi observado diferença significativa para o valor observado sob SSD. Em média, o valor de Cmic sob mata foi cerca de 2,6 e 2 vezes, respectivamente, superior aos valores encontrados sob plantio convencional (SPC) e semeadura direta (SSD). Entre esses sistemas de manejo, o maior valor de Cmic foi observado sob SSD, comparado ao SPC, na média geral e, dentro das épocas de coleta, para o SSD somente na pós-colheita. Em relação ao efeito da época de coleta, o valor mais elevado foi observado no pré-plantio, seguido da pós-colheita e floração, para as amostras de solo de área de mata. Para o SSD o maior valor foi observado na pós-colheita e os menores no pré-plantio e na floração do arroz. Já sob SPC não foi observado diferença significativa para Cmic entre as épocas avaliadas (Tabela 2).
Atributos da biomassa microbiana afetados pelo uso do solo e épocas de coleta de amostras. Valores médios de quatro repetições
Resultados semelhantes aos do Cmic foram observados para o nitrogênio da biomassa (Nmic), quando da comparação da mata com os sistemas de manejo. Apesar de não ter sido observado diferença significativa entre os sistemas de manejo do solo para a média geral, diferenças foram observadas no pré-plantio e na pós-colheita, sendo nessas épocas maior o valor desse atributo no SPC e no SSD, respectivamente. Em relação ao efeito da época de coleta, o valor mais elevado foi observado no pré-plantio, seguido da pós-colheita e floração para todos os usos do solo (Tabela 2).
Os atributos da biomassa Cmic e Nmic servem como indicadores da qualidade do solo (FERREIRA; WENDLAND; DIDONET, 2011FERREIRA, E. P. B.; WENDLAND, A.; DIDONET, A. D. Microbial biomass and enzyme activity of a Cerrado Oxisol under agroecological production system. Bragantia, v. 70, n. 4, p. 1-9, 2011.); contudo, o uso desses parâmetros isoladamente não é o mais apropriado para determinar o estado metabólico das comunidades microbianas do solo (BOWLES et al., 2014BOWLES, T. et al. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biology and Biochemistry, v. 68, p. 252-262, 2014.), necessitando levar em consideração outros atributos microbiológicos da qualidade do solo.
Nesse sentido, a quantificação da respiração basal do solo (RBS) e do quociente metabólico (qCO2) fornecem informações importantes sobre a atividade da microbiota do solo (ANDERSON; DOMSCH, 1990ANDERSON, T. H.; DOMSCH, K. H. Application of eco-physiological quotients (qCO2 and qD) on microbial biomasses from soils of different cropping histories. Soil Biology and Biochemistry, v. 22, n. 2, p. 251-255, 1990.).
Em relação à RBS, foi observado diferença significativa entre as formas de uso do solo em cada época de coleta, sendo o menor valor observado sob SSD, exceto na floração do arroz, em que não foi observado diferença significativa entre SPC, SSD e mata. Considerando a época de amostragem, no geral, os maiores valores de RBS foram observados na pós-colheita; contudo, elevados valores também foram observados na floração do arroz sob SSD e no pré-plantio sob mata (Tabela 2). A maior liberação de CO2, expressa pela RBS, em solo de mata pode ser em função da constante incorporação de resíduos, promovendo aumento da biomassa microbiana e da atividade biológica sobre esse material, resultando na liberação de CO2 (KUZYAKOV, 2010KUZYAKOV, Y. Priming effects: Interactions between living and dead organic matter. Soil Biology and Biochemistry, v. 42, n. 9, p. 1363-1371, 2010.). Portanto, o maior teor de Cmic encontra-se positivamente relacionado com a liberação de CO2 (ADACHI et al., 2006ADACHI, M. et al. Differences in soil respiration between different tropical ecosystems. Applied Soil Ecology, v. 34, n. 2, p. 258-265, 2006.).
Como consequência, considerando a média, o maior valor de qCO2 foi observado sob SPC em comparação ao SSD e à mata, comportamento que se repetiu na pós-colheita e no pré-plantio, apesar de não diferir da mata nessa última época. Na floração, os sistemas de manejo do solo não diferiram e apresentaram maiores valores de qCO2 do que a mata (Tabela 2). Esses resultados indicam que a taxa de liberação de CO2 por unidade de Cmic é menor sob SSD e mata, comparados ao SPC. Isso pode ser explicado pelo fato de que quando a biomassa microbiana se torna mais eficiente na utilização de recursos do ecossistema, menos CO2 é perdido pela respiração e maior proporção de C é incorporada aos tecidos microbianos, o que resulta em diminuição do qCO2 (SILVA et al., 2007SILVA, M. B. et al. Atributos biológicos do solo sob influência da cobertura vegetal e do sistema de manejo. Pesquisa Agropecuária Brasileira, v. 42, n. 12, p. 1755-1761, 2007.).
Dessa forma, sistemas de manejo em que a comunidade microbiana do solo é pouco perturbada apresentam grande estabilidade, contribuindo para maior sustentabilidade (FERREIRA; WENDLAND; DIDONET, 2011FERREIRA, E. P. B.; WENDLAND, A.; DIDONET, A. D. Microbial biomass and enzyme activity of a Cerrado Oxisol under agroecological production system. Bragantia, v. 70, n. 4, p. 1-9, 2011.). Tomando como bases os resultados observados para os atributos da biomassa microbiana, pode-se inferir que, comparado ao SPC, o SSD promove menor perturbação da biomassa microbiana do solo, proporcionando maiores valores de Cmic e menores valores de RBS e qCO2, o que possivelmente resulta em maior sustentabilidade da atividade agrícola sob SSD.
Os resultados de atividade enzimática do solo (AET, Aβ-G e AFA) mostraram que os maiores valores de AET foram observados em solo de mata, especialmente no pré-plantio. Embora não tenha sido observado diferença significativa entre SSD e SPC para a AET, o SSD proporcionou um acréscimo de cerca de 3,5% em relação ao SPC (Tabela 3). Segundo Green et al. (2007)GREEN, V. S. et al. Tillage impacts on soil biological activity and aggregation in a Brazilian Cerrado Oxissol. Soil and Tillage Research, v. 92, n. 1, p. 114-121, 2007. o SSD pode aumentar os valores de AET de 18 a 186% na camada de 0-0,05 cm; contudo, em camadas mais profundas diferenças entre SSD e SPC podem não ser encontradas, como nesse trabalho.
Atividade de enzimas afetada pelo uso do solo e épocas de coleta. Valores médios de quatro repetições
Os elevados valores de Aβ-G observados em solo sob SSD, comparado com a mata (Tabela 3), podem estar relacionados a um efeito positivo das plantas de cobertura do solo na atividade da enzima, provavelmente devido à alta biomassa microbiana produzida em resposta (TEJADA et al., 2006TEJADA, M. et al. Organic amendment based on fresh and composted beet vinasse: influence on physical, chemical and biological properties and wheat yield. Soil Science Society of American Journal, v. 70, n. 3, p. 900-908, 2006.), uma vez que a Aβ-G reflete o status da matéria orgânica e os processos de decomposição ocorrendo no solo.
Já para a AET os maiores valores foram observados em solo da mata; contudo, foi observado diferença significativa entre os sistemas de manejo do solo, sendo o maior valor observado para o SSD (Tabela 3), corroborando os resultados de Wei et al. (2014)WEI, K. et al. Application of 31P NMR sprectroscopy in determining phosphatase activities and P consumption in soil aggregates influenced by tillage residue management practices. Soil and Tillage Research, v. 138, p. 35-43, 2014..
Considerando a média, a contagem de bactérias do solo não diferiu entre os sistemas de manejo do solo e foi muito superior nesses sistemas em comparação com a mata. A época de coleta influenciou a comunidade bacteriana do solo somente sob SSD, no qual maiores valores foram observados no pré-plantio do arroz. Esse comportamento da comunidade bacteriana pode ter sido influenciado pela disponibilidade hídrica no momento das coletas, uma vez que na semana antecedente a cada coleta a ocorrência de chuvas aconteceu de forma diferenciada, com maior pluviometria no pré-plantio (Figura 1). Considerando que a variação sazonal da comunidade de bactérias pode ser afetada pela umidade do solo (BUTENSCHOEN; SCHEU; EISENHAUER, 2011BUTENSCHOEN, O.; SCHEU, S.; EISENHAUER, N. Interactive effects of warming, soil humidity and plant diversity on litter decomposition and microbial activity. Soil Biology and Biochemistry, v. 43, p. 1902-1907, 2011.), a maior disponibilidade de água no pré-plantio pode ter estimulado a comunidade bacteriana, refletindo nos altos valores observados nessa época. Além disso, um somatório dos efeitos dos sistemas de manejo (SPC e SSD), das características químicas e bioquímicas do solo, assim como da influência dos fatores abióticos (temperatura e umidade) pode ter contribuído para tal resultado (Tabela 4).
População de microrganismos do solo em função do sistema de manejo e da época de coleta. Médias de quatro repetições expressas em ufc x104 g-1 de solo seco
Em cada época de coleta foi observado grande variação da população bacteriana do solo entre os sistemas de manejo, sendo que no pré-plantio foi observado maior número no SSD, na floração do arroz no SPC e não foi observado diferença significativa na pós-colheita (Tabela 4), indicando que a forma de uso do solo não afeta significativamente essa comunidade microbiana.
A comunidade de fungos do solo foi a que apresentou menores variações sob as condições avaliadas. De modo geral, ela não foi afetada pelo uso do solo uma vez que não foi observado diferenças entre os sistemas de manejo e a mata. Contudo, no pré-plantio foi observado elevado valor para a contagem de fungos sob SSD, ocasionando diferença significativa desse sistema de manejo em relação à mata e entre as épocas de coleta dentro desse sistema de manejo do solo (Tabela 4). Esses resultados corroboram parcialmente os resultados apresentados por Wang et al. (2010)WANG, Y. et al. Tillage, residue burning and crop rotation alter soil fungal community and water-stable aggregation in arable fields. Soil and Tillage Research, v. 107, n. 2, p. 71-79, 2010., os quais afirmaram que o SSD favorece a população de fungos do solo, apresentando maiores índices de riqueza e diversidade e número total de espécies do que em SPC.
A população de actinobactérias apresentou, na média, valores sob SSD e SPD estatisticamente superiores ao observado para a mata, porém sem diferença significativa entre os dois sistemas de manejo do solo. Foi observado também efeito significativo da época de coleta dentro dos sistemas de manejo do solo, com maior valor para a contagem de actinobactérias na pós-colheita; contudo, não houve efeito da época de coleta sobre a comunidade de actinobactérias em solo de mata (Tabela 4). Resultados semelhantes são relatados na literatura e indicam que diferentes práticas de manejo do solo apresentam efeito sobre a comunidade de actinobactérias. Ainda, em solo de mata os efeitos mais marcantes sobre a comunidade desses microrganismos estão relacionados a variações climáticas (GHORBANI-NASRABADI et al., 2013GHORBANI-NASRABADI, R. et al. Distribution of actinomycetes in different soil ecosystems and effect of media composition on extracellular phosphatase activity. Journal of Soil Science and Plant Nutrition, v. 13, n. 1, p. 223-236, 2013.).
A contagem de diazotróficos sob SSD e SPC apresentou valores superiores ao observado em solo de mata, tanto na média geral, quanto no pré-plantio, embora não tenha sido observado diferença significativa entre os sistemas de manejo do solo. Ainda, na pós-colheita, o valor encontrado sob SPC foi superior ao encontrado para solo de mata. Quanto ao efeito da época de coleta, foi observado diferença significativa somente sob SSD, sendo observado maior contagem de diazotróficos no pré-plantio, comparado à floração do arroz e pós-colheita (Tabela 4).
O número de microrganismos diazotróficos presente em amostras de solo cultivado (SSD e SPD) apresentou tendência de ser maior em comparação ao número observado em solo de mata, possivelmente em função do histórico de cultivo de leguminosas nessa área, como o feijoeiro comum, crotalária, feijão de porco e mucuna, as quais estimulam o aumento da comunidade desses microrganismos. Em trabalho usando solo não cultivado e cultivado com feijão, Junier et al. (2009)JUNIER, P. et al. Composition of diazotrophic bacterial assemblages in bean-planted soil compared to unplanted soil. European Journal of Soil Biology, v. 45, n. 2, p. 153-162, 2009. atribuíram a presença preferencial de bactérias diazotróficas em solo cultivado a um estímulo específico do feijão à proliferação das bactérias diazotróficas ou à formação de microhabitats mais favoráveis ao crescimento saprofítico dessas bactérias. Outros fatores, como a espécie de planta, uso de fertilizantes, N-total, C orgânico total e pH do solo podem também causar variação na estrutura da comunidade de diazotróficos do solo (REARDON; GOLLANY; WUEST, 2014REARDON, C. L.; GOLLANY, H. T.; WUEST. S. B. Diazotrophic community structure abundance in wheat-fallow and wheat-pea crop rotations. Soil Biology and Biochemistry, v. 69, p. 406-412, 2014.).
De forma geral, os atributos da biomassa microbiana do solo, atividade enzimática e contagem de comunidades microbianas foram afetados pelo sistema de manejo do solo (Tabelas 2; 3 e 4), sendo que a mata apresentou valores de Cmic, Nmic, RBS, AET e AFA superiores aos observados sob SSD e SPC e valores de qCO2, Aβ-G, contagem de bactérias, actinomicetos e diazotróficos inferiores aos verificados nos sistemas de manejo do solo. Por outro lado, o SSD apresentou valores de Cmic e AFA superiores aos observados sob SPC e valores de RBS e qCO2 menores que aqueles observados sob SPC, além de haver apresentado valores de qCO2 semelhantes aos observados na mata, corroborando com dados da literatura, os quais indicam que o SSD oferece maior sustentabilidade ao sistema de produção (FERREIRA et al., 2010FERREIRA, E. P. B. et al. Microbial soil quality indicators under different crop rotations and tillage management. Revista Ciência Agronômica, v. 41, n. 2, p. 177-183. 2010.; FERREIRA; WENDLAND; DIDONET, 2011).
Os elevados valores observados para os parâmetros avaliados em solo de mata podem estar relacionados ao maior conteúdo de matéria orgânica, em comparação ao solo sob SSD e SPC (Tabela 1), uma vez que a matéria orgânica promove resultados benéficos às propriedades edáficas (CARDOZO et al., 2008CARDOZO, S. V. et al. Caracterização de propriedades edáficas em áreas sob manejo orgânico e natural na região serrana do Estado do Rio de Janeiro. Semina: Ciências Agrárias, v. 29, n. 3, p. 517-530, 2008.) e microbiológicas do solo (ELFSTRAND; BÅTH; MÅRTENSSON, 2007ELFSTRAND, S.; BÅTH, B.; MÅRTENSSON, A. Influence of various forms of green manure amendment on soil microbial community composition, enzyme activity and nutrient levels in leek. Applied Soil Ecology, v. 36, n. 1, p. 610-621, 2007.; ELFSTRAND; HEDLUND; MÅRTENSSON, 2007ELFSTRAND, S.; HEDLUND, K.; MÅRTENSSON, A. Soil enzyme activities, microbial community composition and function after 47 years of continuous green manuring. Applied Soil Ecology, v. 35, n. 3, p. 610-621, 2007.).
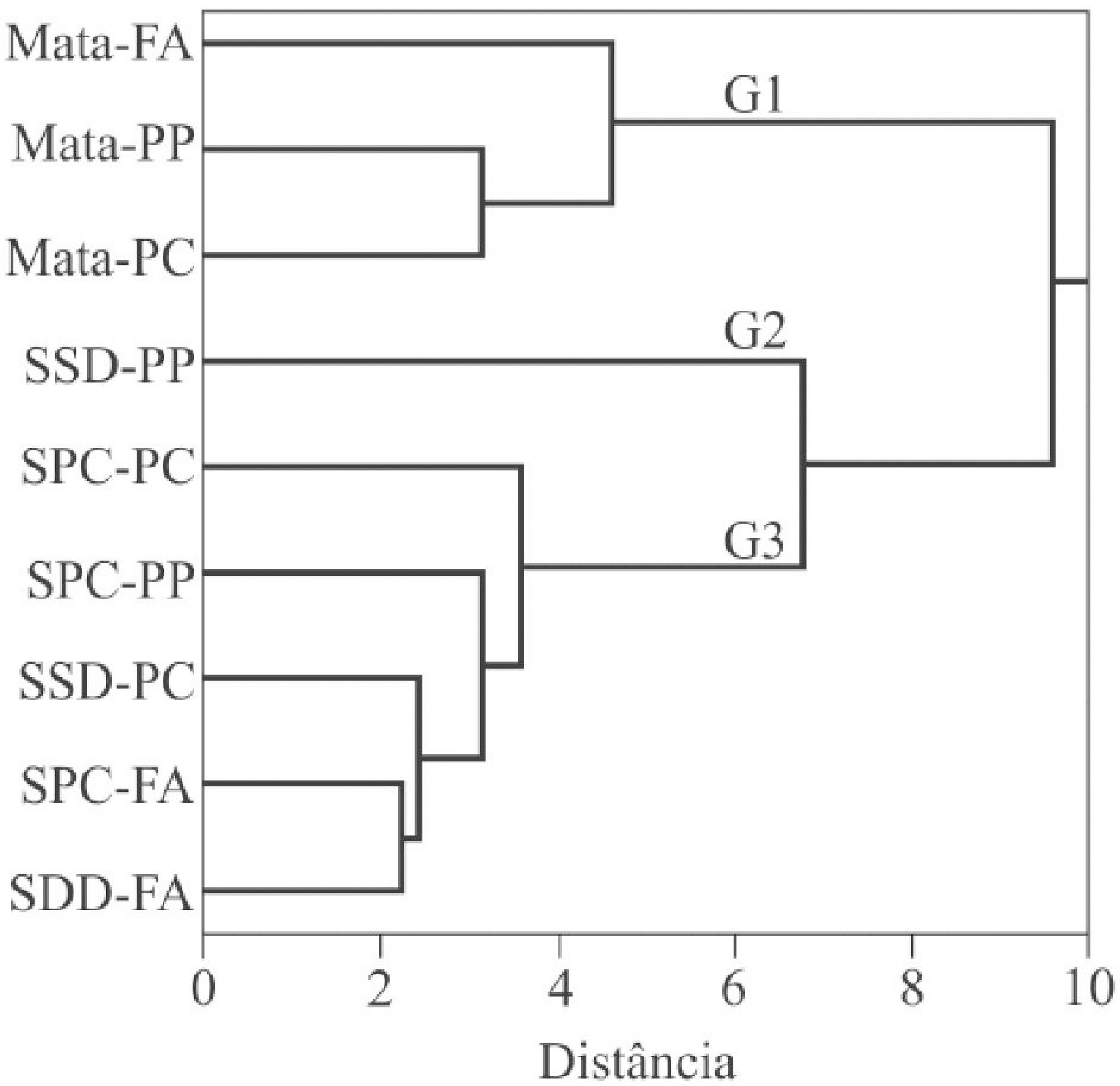
Com base nos dados dos atributos da biomassa microbiana (Tabela 2), da atividade de enzimas do solo (Tabela 3) e da contagem de microrganismos do solo (Tabela 4) foi realizada uma análise multivariada de agrupamento baseado na dissimilaridade entre os tratamentos. Essa análise confirmou a separação dos tratamentos, sendo formados três grupos a uma distância euclidiana de 9,5 (Figura 2).
Dendrograma de agrupamento dos dados microbianos no pré-plantio (PP), floração do arroz (FA) e pós-colheita (PC), sob sistema de plantio convencional (SPC) e sistema de semeadura direta (SSD) e mata
O grupo G1 é formado pelas diferentes épocas de amostragem do solo de mata, enquanto o grupo G2 é formado pelo SSD no pré-plantio, sendo o tratamento que apresentou a menor distância em relação à mata. O grupo G3 é formado pelos outros cinco tratamentos, sendo que, de um lado, o SPC no pré-plantio e pós-colheita ficaram mais próximos, enquanto o SSD e SPC na floração do arroz formaram outro subgrupo.
Apesar da análise de agrupamento não apresentar uma clara separação entre os tratamentos em função dos sistemas de manejo do solo e das épocas de amostragem, um dos tratamentos sob SSD (SSD-PP) foi o que mais se aproximou da área de referência (mata), sugerindo que a atividade agrícola conduzida sob esse tipo de manejo é menos impactante sob o ponto de vista dos indicadores microbiológicos. Resultados semelhantes foram relatados na literatura (CUNHA et al., 2012CUNHA, E. Q. et al. Atributos físicos, químicos e biológicos de solo sob produção orgânica impactados por sistemas de cultivo. Revista Brasileira de Engenharia Agrícola e Ambiental, v. 16, n. 1, p. 56-63, 2012.; FERREIRA et al., 2010FERREIRA, E. P. B. et al. Microbial soil quality indicators under different crop rotations and tillage management. Revista Ciência Agronômica, v. 41, n. 2, p. 177-183. 2010.), cujos autores atribuíram esse comportamento a um possível efeito do SSD na melhoria das qualidades física, química e biológica do solo.
Por outro lado, as diferenças nos agrupamentos observadas nesse trabalho entre as épocas de coleta podem ter sido decorrentes de fatores edafoclimáticos ou da fase de desenvolvimento da cultura do arroz. Contudo, a literatura aponta que o sistema de manejo do solo empregado desempenha papel preponderante na formação dos agrupamentos (FERREIRA; WENDLAND; DIDONET, 2011FERREIRA, E. P. B.; WENDLAND, A.; DIDONET, A. D. Microbial biomass and enzyme activity of a Cerrado Oxisol under agroecological production system. Bragantia, v. 70, n. 4, p. 1-9, 2011.).
CONCLUSÕES
-
Os sistemas de cultivo provocam alterações nos atributos microbiológicos do solo quando comparados com a condição preservada do ambiente de mata;
-
Comparado ao SPC, o SSD proporciona valores de Cmic e qCO2 mais próximos aos observados no solo de mata, denotando menor impacto desse sistema sobre os atributos da biomassa microbiana;
-
A atividade enzimática do solo é pouco influenciada pelos sistemas de manejo do solo; contudo, maior atividade da fosfatase ácida é observada no pré-plantio do arroz sob SSD;
-
De forma semelhante à atividade enzimática, o número de microrganismos cultiváveis avaliados é pouco impactado pelos sistemas de manejo do solo; contudo, menor número de microrganismos foi observado em solo de mata.
-
1
Resultado de projeto financiado pela Empresa Brasileira de Pesquisa Agropecuária/EMBRAPA
REFERÊNCIAS
- ADACHI, M. et al Differences in soil respiration between different tropical ecosystems. Applied Soil Ecology, v. 34, n. 2, p. 258-265, 2006.
- ANDERSON, T. H.; DOMSCH, K. H. Application of eco-physiological quotients (qCO2 and qD) on microbial biomasses from soils of different cropping histories. Soil Biology and Biochemistry, v. 22, n. 2, p. 251-255, 1990.
- BOWLES, T. et al Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biology and Biochemistry, v. 68, p. 252-262, 2014.
- BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Mercado brasileiro de orgânicos deve movimentar R$ 2,5 bi em 2016 Disponível em: < http://www.agricultura.gov.br/comunicacao/noticias/2015/09/mercado-brasileiro-de-organicos-deve-movimentar-rs-2-bi-em-2016>. Acesso em: 08 Mar. 2016.
» http://www.agricultura.gov.br/comunicacao/noticias/2015/09/mercado-brasileiro-de-organicos-deve-movimentar-rs-2-bi-em-2016 - BROOKES, P. C. et al Chloroform fumigation and the release of soil nitrogen: a rapid direct extration method to measure soil microbial biomass nitrogen in soil. Soil Biology and Biochemistry, v. 17, n. 6, p. 837-842, 1985.
- BUTENSCHOEN, O.; SCHEU, S.; EISENHAUER, N. Interactive effects of warming, soil humidity and plant diversity on litter decomposition and microbial activity. Soil Biology and Biochemistry, v. 43, p. 1902-1907, 2011.
- CARDOZO, S. V. et al Caracterização de propriedades edáficas em áreas sob manejo orgânico e natural na região serrana do Estado do Rio de Janeiro. Semina: Ciências Agrárias, v. 29, n. 3, p. 517-530, 2008.
- CUNHA, E. Q. et al Atributos físicos, químicos e biológicos de solo sob produção orgânica impactados por sistemas de cultivo. Revista Brasileira de Engenharia Agrícola e Ambiental, v. 16, n. 1, p. 56-63, 2012.
- ELFSTRAND, S.; BÅTH, B.; MÅRTENSSON, A. Influence of various forms of green manure amendment on soil microbial community composition, enzyme activity and nutrient levels in leek. Applied Soil Ecology, v. 36, n. 1, p. 610-621, 2007.
- ELFSTRAND, S.; HEDLUND, K.; MÅRTENSSON, A. Soil enzyme activities, microbial community composition and function after 47 years of continuous green manuring. Applied Soil Ecology, v. 35, n. 3, p. 610-621, 2007.
- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Sistema brasileiro de classificação de solos 3. ed. Brasília, 2013. 353 p.
- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Centro Nacional de Pesquisa de Solos. Manual de métodos de análise de solos 2. ed. Rio de Janeiro: Embrapa CNPS, 1997. 212 p. (Embrapa CNPS. Documentos, 1).
- EPELDE, L. et al Microbial properties and attributes of ecological relevance for soil quality monitoring during a chemical stabilization field study. Applied Soil Ecology, v. 75, p. 1-12, 2014.
- FERREIRA, E. P. B. et al Microbial soil quality indicators under different crop rotations and tillage management. Revista Ciência Agronômica, v. 41, n. 2, p. 177-183. 2010.
- FERREIRA, E. P. B.; WENDLAND, A.; DIDONET, A. D. Microbial biomass and enzyme activity of a Cerrado Oxisol under agroecological production system. Bragantia, v. 70, n. 4, p. 1-9, 2011.
- GHINI, R.; MENDES, M. D. L.; BETTIOL, W. Métodos de hidrólise de diacetato de fluoresceína (FDA) como indicador da atividade microbiana no solo e supressividade a Rhizoctonia solaniSumma Phytopathologica, v. 24, n. 3, p. 239-242, 1998.
- GHORBANI-NASRABADI, R. et al. Distribution of actinomycetes in different soil ecosystems and effect of media composition on extracellular phosphatase activity. Journal of Soil Science and Plant Nutrition, v. 13, n. 1, p. 223-236, 2013.
- GREEN, V. S. et al Tillage impacts on soil biological activity and aggregation in a Brazilian Cerrado Oxissol. Soil and Tillage Research, v. 92, n. 1, p. 114-121, 2007.
- JENKINSON, D. S.; LADD, J. N. Microbial biomass in soil: measurement and turnover. In: PAUL, E. A.; LADD, J. N. (Ed.). Soil Biology and Biochemistry, v. 5, p. 415-471, 1981.
- JENKINSON, D. S.; POWLSON, D. S. The effects of biocidal treatments on metabolism in soil: a method for measuring soil biomass. Soil Biology and Biochemistry, v. 8, n. 3, p. 209-213, 1976.
- JUNIER, P. et al Composition of diazotrophic bacterial assemblages in bean-planted soil compared to unplanted soil. European Journal of Soil Biology, v. 45, n. 2, p. 153-162, 2009.
- KUZYAKOV, Y. Priming effects: Interactions between living and dead organic matter. Soil Biology and Biochemistry, v. 42, n. 9, p. 1363-1371, 2010.
- R DEVELOPMENT CORE TEAM. R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing, 2011. Disponível em <http://www.R-project.org>. Acesso em: 25 jan. 2013.
» http://www.R-project.org - REARDON, C. L.; GOLLANY, H. T.; WUEST. S. B. Diazotrophic community structure abundance in wheat-fallow and wheat-pea crop rotations. Soil Biology and Biochemistry, v. 69, p. 406-412, 2014.
- SANTOS, T. E. B. et al Variáveis microbiológicas e produtividade do arroz sob diferentes manejos do solo e água. Acta Scientiarum. Agrononomy, v. 29, n. 03, p. 355-366, 2007.
- SILVA, M. B. et al Atributos biológicos do solo sob influência da cobertura vegetal e do sistema de manejo. Pesquisa Agropecuária Brasileira, v. 42, n. 12, p. 1755-1761, 2007.
- SILVA, S. C. da et al Informações meteorológicas para pesquisa e planejamento agrícola, referentes ao município de Santo Antônio de Goiás, GO, 2012 Santo Antônio de Goiás: Embrapa Arroz e Feijão, 2014. 29 p. (Embrapa Arroz e Feijão. Documentos, 298).
- SOUZA, L. M. et al Influência da aplicação de diferentes vermicompostos na biomassa microbiana do solo após cultivo de alface. Revista Brasileira de Agrociência, v. 12, n. 4, p. 429-434, 2006.
- SPARLING, G. P.; WEST, A. W. A direct extraction method to estimate soil microbial C: calibration in situ using microbial respiration and 14C labelled cells. Soil Biology and Biochemistry, v. 20, n. 3, p. 337-343, 1988.
- TABATABAI, M. A. Soil enzymes. In: WEAVER, R. W.; SCOTT, A.; BOTTOMELEY, P. J. (Ed. Methods of soil analysis: microbiological and biochemical properties. Madison: Soil Science Society of America, 1994. p. 778-835. (Special Publication, 5).
- TEDESCO, M. J. et al Análises de solo, plantas e outros materiais 2. ed. Porto Alegre: Universidade Federal do Rio Grande do Sul, 1995. 174 p.
- TEJADA, M. et al Organic amendment based on fresh and composted beet vinasse: influence on physical, chemical and biological properties and wheat yield. Soil Science Society of American Journal, v. 70, n. 3, p. 900-908, 2006.
- VANCE, E. D.; BROOKES, P. C.; JENKINSON, D. S. An extraction method for measuring soil microbial biomass C. Soil Biology and Biochemistry, v. 19, n. 6, p. 703-707, 1987.
- VEZZANI, F. M.; MIELNICZUK, J. Uma visão sobre qualidade do solo. Revista Brasileira de Ciência do Solo, v. 33, n. 4 p.743-755, 2009.
- WALKLEY, A.; BLACK, I. A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Science, v. 37, n. 01, p. 29-38, 1934.
- WANG, Y. et al Tillage, residue burning and crop rotation alter soil fungal community and water-stable aggregation in arable fields. Soil and Tillage Research, v. 107, n. 2, p. 71-79, 2010.
- WEI, K. et al Application of 31P NMR sprectroscopy in determining phosphatase activities and P consumption in soil aggregates influenced by tillage residue management practices. Soil and Tillage Research, v. 138, p. 35-43, 2014.
Datas de Publicação
-
Publicação nesta coleção
Jan-Mar 2017
Histórico
-
Recebido
03 Fev 2015 -
Aceito
10 Abr 2016

 Fonte:
Fonte: