Resumos
A Mata Atlântica do médio rio Doce é considerada uma das formações vegetais mais ameaçadas de Minas Gerais, devido à intensa destruição ocorrida no passado. As florestas encontradas representam a única fonte de informação sobre a vegetação da região. O objetivo deste estudo foi caracterizar algumas comunidades arbóreas do Parque Estadual do Rio Doce e de fragmentos do entorno quanto à estrutura, composição, similaridade florística e distribuição das espécies. Foram estudadas seis áreas usando o método de ponto quadrante. Em cada área foram amostrados 70 pontos tomando-se as medidas de circunferência (> 15 cm) e altura das árvores. No total foram identificadas 61 famílias, 195 gêneros e 373 espécies. O número de famílias variou de 30 a 36 por área de estudo e o de espécies de 80 a 101. A similaridade florística (Jaccard) entre as áreas variou de 6,3 a 24,1 %, mostrando serem as florestas bastante distintas. Houve correlação significativa (CCA) entre a distribuição de espécies e as variáveis de solo. Os índices de diversidade de Shannon (H') variaram de 3,66 a 4,10. O valor mediano dos diâmetros variou de 8,0 a 10,9 cm e o das alturas de 7,0 a 10,0 m. A heterogeneidade florística encontrada no Parque Estadual do Rio Doce e nos fragmentos do entorno refletiu o histórico de perturbações e a influência dos fatores edáficos na distribuição das espécies arbóreas no médio rio Doce.
comunidade arbórea; fitossociologia; heterogeneidade
The Atlantic Forest of the middle Rio Doce is considered one of the most endangered vegetation of Minas Gerais, because of intensive destruction occurred in the past. The forests are the only sources of information of the original vegetation in the region. The aim of this study was to characterize the floristic composition, structure, floristic similarity and the species distribution of the forest community from Rio Doce state park and neighboring areas. We studied six sites, using the point-centered quarter method. In each area, we sampled 70 points, and measured circumference (> 15 cm) and height of trees. We found 61 families, 195 genera and 373 species. The number of families varied from 30 to 36 for study area and, the number of species from 80 to 101. Floristic similarity indexes (Jaccard) among areas ranged from 6.3 to 24.1 %, showing that the areas were very different. There was significant relationship between species distribution and soil variables. Shannon Diversity Indexes varied from 3.66 to 4.10. The median diameter varied from 8.0 to 10.9 cm and the height from 7.0 to 10.0 m. The floristic heterogeneity seems to reflect the disturbance history and the influence of soil factors in the distribution of tree species in the middle Rio Doce.
tree community; phytosociology; heterogeneity
ARTIGOS ORIGINAIS
Florística e estrutura do componente arbóreo de remanescentes de Mata Atlântica do médio rio Doce, Minas Gerais, Brasil
Floristic and structure analysis of a tree component in remnants of Atlantic forest in the Middle Rio Doce, Minas Gerais, Brazil
Glauco Santos FrançaI,1 1 Autor para correspondência: glauco.franca@ifsudestemg.edu.br ; João Renato StehmannII
IParte da tese de Doutorado - Pós-Graduação em Biologia Vegetal, ICB, UFMG
IIUniversidade Federal de Minas Gerais, Inst. Ciências Biológicas, Depto. Botânica, Av. Antônio Carlos 6627, 31270-110, Belo Horizonte, MG, Brasil
RESUMO
A Mata Atlântica do médio rio Doce é considerada uma das formações vegetais mais ameaçadas de Minas Gerais, devido à intensa destruição ocorrida no passado. As florestas encontradas representam a única fonte de informação sobre a vegetação da região. O objetivo deste estudo foi caracterizar algumas comunidades arbóreas do Parque Estadual do Rio Doce e de fragmentos do entorno quanto à estrutura, composição, similaridade florística e distribuição das espécies. Foram estudadas seis áreas usando o método de ponto quadrante. Em cada área foram amostrados 70 pontos tomando-se as medidas de circunferência (> 15 cm) e altura das árvores. No total foram identificadas 61 famílias, 195 gêneros e 373 espécies. O número de famílias variou de 30 a 36 por área de estudo e o de espécies de 80 a 101. A similaridade florística (Jaccard) entre as áreas variou de 6,3 a 24,1 %, mostrando serem as florestas bastante distintas. Houve correlação significativa (CCA) entre a distribuição de espécies e as variáveis de solo. Os índices de diversidade de Shannon (H') variaram de 3,66 a 4,10. O valor mediano dos diâmetros variou de 8,0 a 10,9 cm e o das alturas de 7,0 a 10,0 m. A heterogeneidade florística encontrada no Parque Estadual do Rio Doce e nos fragmentos do entorno refletiu o histórico de perturbações e a influência dos fatores edáficos na distribuição das espécies arbóreas no médio rio Doce.
Palavras-chave: comunidade arbórea, fitossociologia, heterogeneidade.
ABSTRACT
The Atlantic Forest of the middle Rio Doce is considered one of the most endangered vegetation of Minas Gerais, because of intensive destruction occurred in the past. The forests are the only sources of information of the original vegetation in the region. The aim of this study was to characterize the floristic composition, structure, floristic similarity and the species distribution of the forest community from Rio Doce state park and neighboring areas. We studied six sites, using the point-centered quarter method. In each area, we sampled 70 points, and measured circumference (> 15 cm) and height of trees. We found 61 families, 195 genera and 373 species. The number of families varied from 30 to 36 for study area and, the number of species from 80 to 101. Floristic similarity indexes (Jaccard) among areas ranged from 6.3 to 24.1 %, showing that the areas were very different. There was significant relationship between species distribution and soil variables. Shannon Diversity Indexes varied from 3.66 to 4.10. The median diameter varied from 8.0 to 10.9 cm and the height from 7.0 to 10.0 m. The floristic heterogeneity seems to reflect the disturbance history and the influence of soil factors in the distribution of tree species in the middle Rio Doce.
Key words: tree community, phytosociology, heterogeneity.
Introdução
A Mata Atlântica e seus ecossistemas associados cobriam originalmente uma área de 1.110.000 km2, que correspondia a aproximadamente 15% do território brasileiro (Fundação SOS Mata Atlântica/INPE/ISA 1998). O processo de ocupação do Brasil levou a drástica redução de sua cobertura vegetal e calcula-se que 88,27% da Floresta Atlântica original tenha sido perdida, restando hoje 11,73% (16.377.472 ha) de vegetação remanescente, disposta de maneira esparsa ao longo da costa brasileira e no interior das Regiões Sul e Sudeste, além de importantes fragmentos no sul dos estados de Goiás e Mato Grosso do Sul e no interior dos estados do Nordeste (Ribeiro et al. 2009). Em Minas Gerais, a porcentagem de cobertura florestal natural em relação à área do estado foi reduzida de 46%, em 1500, para apenas 9,64% (2.624.626 ha) em 2010 (Fundação SOS Mata Atlântica/INPE/ISA 2008-2010).
No Sudeste do Brasil, as bacias do leste, localizadas no sul da Bahia, Espírito Santo, Minas Gerais e norte do Rio de Janeiro, compreendiam uma grande extensão contínua de florestas estacionais e ombrófilas, que foram destruídas ou severamente fragmentadas nos últimos quarenta anos. Essa região compreende as formações florestais associadas às bacias dos rios Jequitinhonha, Mucuri e Doce (Oliveira-Filho et al. 2005).
A bacia do rio Doce, originalmente quase totalmente recoberta com vegetação característica de florestas semideciduais pertencentes ao domínio da Mata Atlântica, possui altíssima riqueza e diversidade biológica, além de abrigar grande número de espécies da fauna e da flora com distribuição restrita a esse ecossistema (Fonseca 1997). A região do médio rio Doce apresenta remanescentes de florestas que sofreram diferentes graus de perturbação, seja pela ação de desmatamentos, corte seletivo de madeira e/ou fogo. Alguns remanescentes encontram-se preservados, principalmente aqueles localizados em Unidades de Conservação como Parques Estaduais e Reservas Particulares do Patrimônio Natural (RPPNs).
Existem poucos levantamentos sobre a flora arbórea do médio rio Doce, destacando-se os estudos fitossociológicos realizados no Parque Estadual do Rio Doce (PERD) (CETEC 1981; Lopes 2002; Camargos et al. 2008), o estudo das tipologias florestais do referido Parque realizado por Andrade et al. (1997), os trabalhos sobre a composição florística do PERD e da RPPN Feliciano Miguel Abdala (Caratinga), apresentados por Lombardi & Gonçalves (2000) e Gonçalves & Lombardi (2004) e os estudos sobre alguns grupos de famílias arbóreas (Pedralli et al. 1986; Bortoluzzi et al. 2004; Nunes et al. 2007).
Padrões florísticos de distribuição das espécies arbóreas, associados às variáveis geográficas e climáticas, têm sido analisados para o sul da Bahia, estados de São Paulo e Rio de Janeiro (Oliveira-Filho & Fontes 2000; Oliveira-Filho et al. 2005). Dentre eles podemos citar o padrão relacionado a sazonalidade da precipitação que interfere na distinção entre florestas ombrófilas e semidecíduas. O segundo padrão refere-se à variação latitudinal associada ao regime de chuvas, que afetaria a distribuição florística, principalmente em florestas ombrófilas. O terceiro padrão florístico trata das variações altitudinais tanto nas florestas ombrófilas quanto semideciduais. Contudo, estes estudos evidenciaram também uma escassez de informações para grande parte da região das bacias do Leste, em especial o leste de Minas Gerais e sul do Espírito Santo.
Portanto, os objetivos deste trabalho compreenderam a caracterização da composição florística e da estrutura de algumas comunidades florestais arbóreas do médio rio Doce, além da análise das variáveis do solo e das relações florísticas entre as formações florestais.
Material e Métodos
O presente estudo foi realizado em áreas de Floresta Estacional Semidecidual Submontana e de Terras Baixas (Oliveira-Filho & Fontes 2000; Oliveira-Filho et al. 2006), também classificadas como Floresta Estacional Semideciduifólia Submontana Crassissólica Pauperinútrica e de Baixada (Oliveira-Filho 2009a, b), pertencentes ao domínio Mata Atlântica, na região do médio rio Doce, no Leste do Estado de Minas Gerais (Fig. 1). Três remanescentes localizaram-se no interior do PERD (Campolina - CAM, Macuco - MAC e Mumbaça - MUMB), nos municípios de Dionísio e Timóteo, e três no seu entorno (RPPN Fazenda Macedônia - MACE, RPPN Fazenda Sacramento - SACR e RPPN Morro do Gavião - GAV), nos municípios de Ipaba, Pingo d'Água e Dionísio, respectivamente.
O PERD possui 35.974 ha com altitudes entre 230 a 515 m. Seus limites naturais são o rio Doce à Leste e o rio Piracicaba ao Norte (Andrade et al. 1997). Os fragmentos estudados no entorno do Parque apresentaram áreas que variaram de 282 a 560 ha e altitudes entre 220 a 586 m. Os solos predominantes na região são os Latossolos Húmicos, Podzólico Vermelho-Amarelo Eutrófico e Aluviais Eutróficos (IGA 1982; EMBRAPA 2007). O clima da região é do tipo Aw de Köppen (Tropical Úmido de Savana, megatérmico, com inverno seco e verão chuvoso). A temperatura média do mês mais frio é superior a 18ºC. A precipitação do mês mais seco é inferior a 60 mm (Antunes 1986). Os dados fornecidos pela Estação Meteorológica da CENIBRA, em Belo Oriente, MG (19º20' S - 42º24' W, 216 m.), no período de janeiro de 1985 a dezembro de 2004, mostraram valores médios de temperatura de 24,8ºC e precipitação anual média de 1.204,3 mm. Observou-se um longo período de déficit hídrico, principalmente nos meses de abril a setembro. Tal fato afeta significativamente a fisionomia das florestas da região, com uma caducifolia típica das florestas semideciduais submontanas.
Cinco áreas são constituídas por uma vegetação classificada como Floresta Estacional Semidecidual de Terras Baixas (Oliveira-Filho & Fontes 2000) e ocorrem em áreas de "baixada", próximas a lagoas ou rios. Um dos fragmentos, GAV, formado por Floresta Estacional Semidecidual Submontana, localiza-se em área de encosta, sem coleções de água e em topografia mais inclinada (Tab 1).
Para a realização do levantamento florístico foram coletados espécimes férteis próximos às áreas de amostragem. Eles foram herborizados (Fidalgo & Bononi 1984) e incorporados ao Herbário BHCB, do Departamento de Botânica do Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais (UFMG). As identificações foram feitas por comparação com espécimes depositados na coleção do herbário BHCB, além de consultas à literatura taxonômica especializada e envio de duplicatas para especialistas em instituições nacionais e estrangeiras. As espécies foram classificadas nas famílias seguindo o sistema da APG II (2003).
O estudo fitossociológico foi realizado através da marcação de 420 pontos quadrantes, sendo 70 pontos em cada uma das seis áreas estudadas. Os pontos foram distribuídos em sete linhas amostrais de 135 m, distanciadas em 15 m e orientadas pela bússola na direção Norte-Sul, para duas áreas (SACR e GAV) e Noroeste-Sudeste para as demais áreas. Em cada linha foram marcados, através de estacas, 10 pontos quadrantes (equidistantes em 15 m), sendo amostrada uma árvore em cada quadrante, totalizando quatro por ponto. Foram amostrados ao todo 1.680 indivíduos arbóreos (280 por área), perfazendo aproximadamente 1,35 ha de área equivalente. Utilizou-se como critério de inclusão aqueles indivíduos arbóreos que apresentaram CAP (Circunferência a Altura do Peito) > 15 cm, a 1,30 m do solo. Os indivíduos foram marcados com placas de alumínio, sendo estimados a altura da copa, o fuste (altura da primeira ramificação), com auxílio de podão telescópico, medido o CAP e coletadas amostras de material botânico.
Para o cálculo dos parâmetros fitossociológicos foram incluídas todas as árvores vivas amostradas, inclusive as espécies não identificadas. Estas foram numeradas e incluídas no levantamento como indeterminadas. Já para os cálculos de porcentagem de número de espécies, número de indivíduos e número de famílias, as indeterminadas foram excluídas da análise. Os parâmetros calculados na análise fitossociológica foram: densidade relativa (DR), frequência relativa (FR), dominância relativa (DoR), valor de importância (VI) e valor de cobertura (VC). Foi utilizado, na análise, o programa FITOPAC versão 1.0 (Shepherd 1994). Foram elaborados histogramas de frequência de classes de diâmetro e altura das árvores encontradas em cada área. Utilizou-se também o teste não-paramétrico de Kolmogorov-Smirnov (Sokal & Rohlf 1995) para verificar as diferenças estruturais entre áreas, considerando-se os valores de diâmetro e altura das espécies. Os cálculos foram realizados através do programa STATISTICA for Windows versão 4.3.
Foi calculado o índice de diversidade de Shannon (H'), na base logarítmica natural, e a equabilidade de Pielou correspondente (J) (Zar 1999). Para a análise de similaridade florística entre as áreas utilizou-se o coeficiente de Jaccard. A comparação entre as matrizes de distância geográfica e similaridade foi realizada através do teste de Mantel (Sokal & Rohlf 1995), usando um teste de Monte Carlo com 5.000 permutações para avaliar a significância. Os testes foram realizados utilizando-se o programa PC-ORD for Windows versão 5 (McCune & Mefford 1999).
Foram coletadas amostras simples de solo superficial (0-20 cm), na porção mediana de cada linha de amostragem, sendo posteriormente misturadas e enviadas ao Instituto Mineiro de Agropecuária (IMA), para a obtenção das variáveis: pH em água; teores K, P, Ca, Mg e Al; soma de bases (SB), saturação por bases (V), capacidade de troca de cátions (T), capacidade efetiva de troca de cátions (t), índice de saturação por alumínio (m), índice de saturação por bases (V), matéria orgânica (MO), C e N; e proporções de areia (grossa e fina), silte e argila.
Para o estudo das correlações entre a distribuição das espécies arbóreas e as variáveis ambientais realizou-se a análise de correspondência canônica, CCA (ter Braak 1995), utilizando-se o programa PC-ORD for Windows versão 5 (McCune & Mefford 1999). A matriz de abundância das espécies consistiu do número de indivíduos em relação ao somatório das linhas de amostragem de cada área, considerando como uma única parcela. Foram incluídas apenas as espécies com N > 10 indivíduos. A matriz de variáveis de solo por parcela incluiu inicialmente onze variáveis de solo (pH, H + Al, Al, t, m, V, M.O., areia grossa, areia fina, silte e argila). Após análise preliminar, seis destas foram eliminadas por apresentarem redundâncias ou correlações fracas (< 0,5 com os eixos 1 e 2). As variáveis remanescentes foram pH, m,V, areia grossa e silte. O teste de permutação de Monte Carlo (ter Braak 1988) foi aplicado para verificar a significância das correlações existentes nos dois primeiros eixos de ordenação.
Foram utilizadas curvas médias de acúmulo de espécies (Gotelli & Colwell 2001), calculadas para as seis áreas, com o objetivo de comparar a riqueza de espécies encontradas entre os diferentes locais amostrados. Foi utilizado o programa Ecosim - versão 7.0 (Gotelli & Enstminger 2006) que calculou também os intervalos de confiança (95%) das médias. As curvas foram construídas com as médias das riquezas obtidas em 1.000 re-amostragens de números pré-determinados de indivíduos (10, 20, 30, etc.), utilizando a opção de amostragem com reposição independente ("independent sampling").
As espécies ameaçadas de extinção foram relacionadas com base em listas de espécies ameaçadas da flora do Brasil (MMA 2008), Lista da Flora Ameaçada de Extinção com ocorrência no Brasil (IUCN 2012) e Lista de Espécies Ameaçadas de Extinção da Flora do Estado de Minas Gerais (Drummond et al. 2008).
Resultados
Considerando a diversidade florística das áreas estudadas, foram identificadas, no total, 373 espécies arbóreas, 194 gêneros e 60 famílias (ver Apêndice). Das 373 espécies listadas, 327 foram encontradas nos pontos amostrais. As famílias com maior número de espécies foram Fabaceae (67), Lauraceae (27), Myrtaceae (26), Rubiaceae (19), Euphorbiaceae, Meliaceae e Salicaceae (16), Sapotaceae (15), Moraceae (13), Annonaceae e Malvaceae (12), Bignoniaceae, Rutaceae e Sapindaceae (10). Juntas estas famílias representaram 60% do total de espécies inventariadas. Observou-se ainda que 30% das famílias (17) foram amostradas por uma única espécie. Fabaceae apresentou maior número de gêneros identificados (32) e maior riqueza de espécies em todas as áreas amostradas, seguida de Euphorbiaceae (13) e Rubiaceae (12). Lauraceae se destacou em três áreas (CAM, MACU e GAV) e Myrtaceae em duas (MACU e MACE).
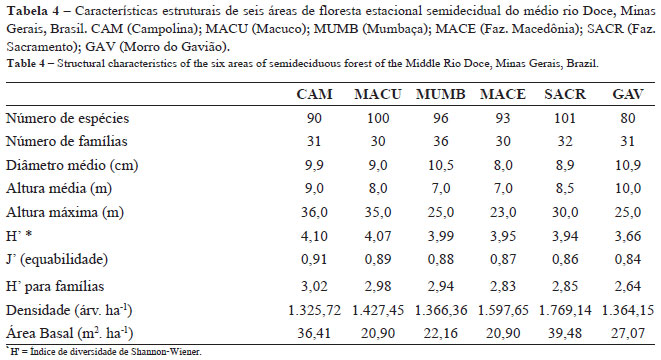
O número total de famílias variou de 33 (MACE) a 41 (MACU). Os gêneros com maior número de espécies foram: Ocotea (15), Casearia (12), Trichilia (10), Eugenia e Machaerium (9), Inga, Pouteria e Swartzia (7), Ficus e Marlierea (6). O número de famílias no levantamento fitossociológico variou de 30 (GAV e MACU) a 36 (MUMB) e o de espécies de 80 (GAV) a 101 (SACR) (ver Apêndice).
Joannesia princeps se destacou por ser mais frequente em cinco das seis áreas amostradas. Mereceram destaque ainda: Guarea macrophylla, Neoraputia alba e Virola gardneri por ocorrerem, com maior frequência, em três ou mais áreas estudadas. Três espécies foram encontradas em todos os remanescentes: Andradea floribunda, Carpotroche brasiliensis e G. macrophylla (Tab. 2). Esta última apresentou as maiores populações dos seis levantamentos realizados.
As espécies melhor representadas quanto ao número de indivíduos, considerando-se todas as áreas estudadas, foram: Senefeldera multiflora (72 indivíduos), Virola gardneri (51), Mabea fistulifera (48), Joannesia princeps (45), Sorocea guilleminiana (32) e G. macrophylla (29).
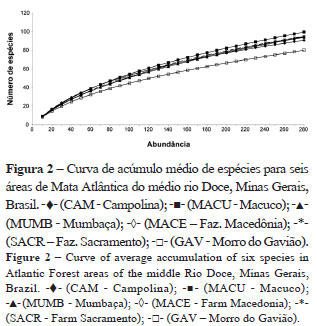
Foram construídas curvas de acúmulo médio de espécies para os locais estudados (Fig. 2). Os valores de riqueza observada e estimada, com os respectivos intervalos de confiança (95%), foram listados para uma amostra de 280 indivíduos (Tab. 3). Comparando-se os valores encontrados, duas comunidades destacaram-se, sendo que a primeira (MACU) apresentou riqueza estimada de espécies maior do que a das outras áreas. A segunda comunidade (GAV) apresentou riqueza estimada menor que as demais. As outras quatro áreas apresentaram riquezas estimadas semelhantes.
Os índices de diversidade de Shannon variaram de 3,66 (GAV) a 4,10 (CAM) para as espécies, e de 2,64 (GAV) a 3,02 (CAM) para as famílias (Tab. 4). Os valores de equabilidade (J') para as espécies variaram de 0,84 (GAV) a 0,91 (CAM), indicando pouca concentração de abundâncias relativas em espécies dominantes (Tab. 4).
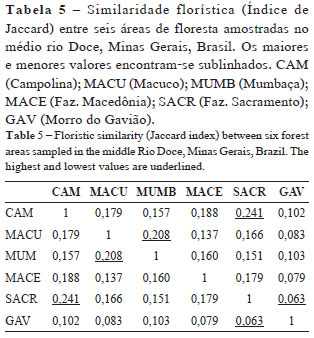
Os valores encontrados para o Índice de Jaccard mostraram similaridade florística entre áreas de 6,3 a 24,1% (Tab. 5), indicando que a composição florística arbórea destas comunidades é muito distinta, com poucas espécies em comum. A maior similaridade obtida entre áreas foi de 24,1% (entre CAM e SACR) e a menor 6,3% (entre SACR e GAV). A comparação das matrizes de distância geográfica e similaridade florística (Jaccard) pelo teste de Mantel não indicou correlação significativa (r = -0,16, t = -0,38 e p = 0,70).
As florestas estudadas apresentaram características florísticas distintas. Avaliando-se as doze principais espécies em termos de valor de importância (VI) (Tab. 2) verificou-se que apenas Virola gardneri foi comum a quatro levantamentos (CAM, MACU, MACE e SACR). Senefeldera multiflora ocorreu em três levantamentos (CAM, MUMB e SACR) e Neoraputia alba também se destacou em três áreas (MACU, MACE e SACR). Dalbergia nigra foi comum a dois levantamentos (GAV e SACR). Duas áreas (CAM e SACR) apresentaram o maior número de espécies em comum: N. alba, Pseudopiptadenia warmingii, S. multiflora e V. gardneri.
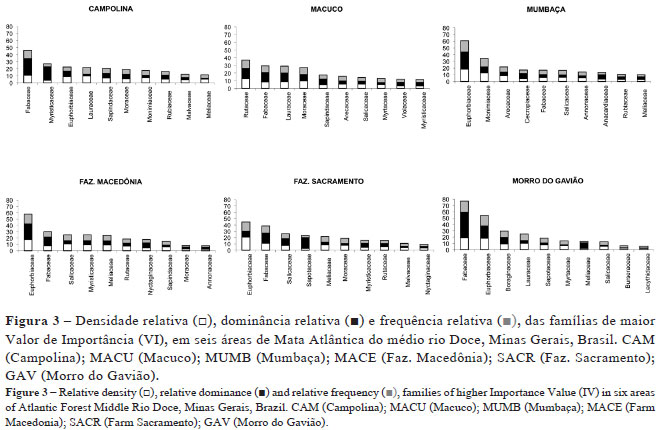
Foram observados os parâmetros fitossociológicos de densidade, dominância e frequência relativas das dez famílias com os maiores Valores de Importância (VI), nas seis áreas de Mata Atlântica do médio rio Doce (Fig. 3). Euphorbiaceae, Fabaceae e Rutaceae estão entre as mais representativas, considerando os valores elevados de densidade, dominância e frequência relativas quando comparadas às demais famílias. Euphorbiaceae destacou-se em três áreas (MUMB, MACE e SACR), Fabaceae em duas (CAM e GAV) e Rutaceae em uma (MACU).
Os dados de densidade, dominância e frequência relativa foram apresentados considerando as dez espécies com maior Valor de Importância (VI), nas seis áreas de Mata Atlântica do médio rio Doce (Fig. 4). Joannesia princeps se destacou em relação à dominância em três áreas (CAM, MUMB e MACE). Neoraputia alba destacou-se em apenas uma área (MACU), considerando os três parâmetros. Senefeldera multiflora apresentou maior densidade e frequência relativa em uma área (SACR). Mabea fistulifera, Cordia sellowiana e Pseudopiptadenia contorta sobressaíram em relação à dominância em outra área (GAV). Virola gardneri se destacou quanto à densidade e frequência em duas áreas (CAM e MACE). Chrysophyllum imperiale apresentou grande dominância relativa em uma única área fora do PERD (SACR).
O valor mediano dos diâmetros dos indivíduos das florestas estudadas variou de 8,0 (MACE) a 10,9cm (GAV) (Tab. 4). A maior proporção de indivíduos foi encontrada nas classes de menor diâmetro (<10 e 15-20 cm), em todas as áreas estudadas. Em duas florestas (MUMB e MACE) faltaram indivíduos nas classes de maior diâmetro (> 50cm) (Fig. 5). O teste de Kolmogorov-Smirnov para os valores de DAP foi significativo para as seguintes áreas: CAM e MACE (p < 0,001), MACU e GAV (p < 0,05), MUMB e MACE (p < 0,001), MUMB e SACR (p < 0,05), MACE e GAV (p < 0,001), SACR e GAV (p < 0,05). Uma área (GAV) apresentou grande quantidade de indivíduos com caules múltiplos, 28% do total amostrado, em comparação às outras florestas estudadas.
O valor mediano das alturas das árvores estudadas variou de 7 m (MACE e MUMB) a 10 m (GAV) (Tab. 4). Três florestas (CAM, MACU e SACR) apresentaram-se mais estruturadas com melhor distribuição dos indivíduos nas classes de altura, sendo que algumas árvores ultrapassaram os 50 m. A maior proporção dos indivíduos foi encontrada nas classes de 6-10 e 10-16 m. Árvores acima de 30 m de altura foram registradas em apenas duas áreas (CAM e MACU). Em três áreas (MUMB, MACE e GAV) não foram encontrados indivíduos de grande porte, entre as classes 28 e 30 m. (Fig. 5). O teste de Kolmogorov-Smirnov para altura foi significativo para as seguintes áreas: GAV e todas as outras; CAM, MUMB e MACE (p < 0,001); MACU e MACE (p < 0,001); MUMB e SACR (p < 0,001); MACE e SACR (p < 0,001).
A menor densidade por área foi 1.325,72 ind.ha-1 (CAM) e a maior 1.769,14 ind.ha-1 (SACR). Os maiores valores de área basal por hectare foram 36,41 m2.ha-1 (CAM) e 39,48 m2.ha-1 (SACR) e os menores foram encontrados para duas áreas (MACU e MACE), sendo 20,90 m2.ha-1, em cada (Tab. 4).
Considerando a análise dos solos das seis áreas estudadas, observou-se que duas delas (GAV e SACR) apresentaram as maiores variações em relação à composição química e granulométrica (Tab. 6). Na primeira o pH foi o mais baixo dentre os solos analisados, com maior acidez potencial (H + Al) e acidez trocável (Al+++) e maior teor de areia grossa. Na segunda o pH foi o mais alto de todas as áreas estudadas, com menores valores de acidez potencial e acidez trocável, além de maior porcentagem de areia fina.
As amostras de solo de três áreas (CAMP, MACE e SACR) apresentaram elevada saturação por bases (V superior a 50%) (EMBRAPA 1999), portanto, alta concentração de cátions trocáveis (Tab. 6). Dessa forma, estes solos podem ser classificados como eutróficos, ricos em nutrientes. Os solos de outras três florestas (MACU, MUMB e GAV) apresentaram valores elevados para o teor de alumínio (Al+++ superior a 0,5 cmolc.kg-1) e índice de saturação por alumínio, por isso estes solos podem ser considerados álicos (Resende et al. 2002). Em uma delas (MUMB) o solo apresentou índice de saturação de alumínio pouco menor que 50%.
A análise de correspondência canônica (CCA) indicou fortes correlações entre a distribuição das abundâncias das principais espécies na amostra e as variáveis de solo utilizadas (Figs. 6 e 7). A CCA produziu autovalores elevados, respectivamente 0,938 e 0,538 para os eixos de ordenação 1 e 2. Os dois primeiros eixos explicaram 39,3% e 22,6% da variância global (total acumulado de 62%), isto indicou pouca variância remanescente não explicada ("ruído" baixo na estrutura dos dados). Os testes de permutação de Monte Carlo indicaram gradientes significativos nos três eixos de ordenação (p = 0,01 para os autovalores) e correlações significativas com as variáveis de solo para os dois primeiros eixos (p = 0,01 para correlações espécie-ambiente). As variáveis com correlações internas (intra-set) mais fortes com o primeiro eixo foram em ordem decrescente: índice de saturação de alumínio (r = -0,764), silte (r = 0,662), areia grossa (r = -0,620), índice de saturação de bases (r = 0,569) e pH (r = 0,567). Para o segundo eixo, as variáveis mais fortemente correlacionadas foram: índice de saturação de bases (r = -0,622) e pH (r = -0,573). No terceiro eixo, a variável que se destacou foi silte, que apresentou um valor próximo de 0,500 (r = 0,476), as demais variáveis foram inferiores a 0,200.
Os diagramas gerados pelas análises de componentes principais indicaram a formação de três grupos de amostras (Fig. 7). O primeiro grupo foi constituído por espécies que ocuparam terrenos argilosos, próximos a sistemas lacrustres: Alchornea triplinervia, Euterpe edulis, Guarea macrophylla, Guatteria sp., Pouroma guianensis e Siparuna guianensis. Estas espécies foram típicas de uma área estudada (MUMB). O segundo grupo foi formado por espécies que ocuparam áreas secundárias, sem cursos d'água, com solos mais ácidos, com maiores teores de alumínio e acidez potencial: Anadenanthera colubrina, Carpotroche brasiliensis, Cordia sellowiana, Eugenia acutata, Mabea fistulifera e Pouteria pachycalyx. Tais espécies também foram encontradas em uma única área (GAV). O terceiro grupo foi formado por espécies correlacionadas aos solos com maior proporção de matéria orgânica e pH elevado (menos ácidos e ricos em nutrientes), localizados próximos aos rios. Estão incluídas neste grupo as espécies Amaioua guianensis, Actinostemon klotzchii, Casearia commersoniana, Neoraputia alba, Ocotea odorifera, Oxandra martiniana, Rinorea guianensis, Sorocea guilleminiana, Toulicia subsquamulata, Trichilia pallida, e Virola gardneri. Elas foram observadas em quatro áreas (CAM, MACU, MACE e SACR). Duas espécies, Jonnesia princeps e Senefeldera multiflora, ocuparam áreas onde os solos apresentaram maior porcentagem de silte.
Discussão
A diversidade florística no médio rio Doce coincide com estudos realizados em florestas por Oliveira-Filho & Fontes (2000). Nove das famílias também foram relacionadas pelos autores como predominantes nas florestas estacionais semideciduais do Domínio Mata Atlântica (Myrtaceae, Fabaceae, Rubiaceae, Lauraceae, Euphorbiaceae, Moraceae, Rutaceae, Sapindaceae e Bignoniaceae). Dentre elas, quatro se destacaram quanto à riqueza de espécies (Fabaceae, Myrtaceae, Lauraceae e Rubiaceae). A predominância de Fabaceae deve-se tanto a maior ciclagem e retenção de nutrientes de espécies desta família quanto a sua maior produtividade primária em ecossistemas florestais tropicais (Martins 1991).
Myrtaceae destaca-se no domínio Mata Atlântica, por apresentar maior número de espécies arbóreas em florestas semidecíduas (Oliveira-Filho & Fontes 2000). Lauraceae é apontada como característica de florestas montanas dos Andes e América Central (Gentry 1995) e tem presença marcante em comunidades florestais em São Paulo e Espírito Santo (Leitão-Filho 1982; Thomaz & Monteiro 1997).
Dentre os gêneros com maior número de espécies, sete também predominaram nos levantamentos realizados para a bacia do rio Doce (Eugenia, Ocotea, Machaerium, Ficus, Inga, Casearia e Pouteria). Eles estão entre os mais representativos de florestas semideciduais do sudeste brasileiro (Oliveira-Filho & Fontes 2000). Eugenia, Ocotea e Inga foram listados como mais ricos em florestas semideciduais, em Viçosa, MG (Meira-Neto & Martins 2002) e em Linhares, ES (Jesus & Rolim 2005; Rolim et al. 2006). Estes gêneros apresentam grande riqueza de espécies na Mata Atlântica, com elevado número de endemismos (Athayde 2005). Inga tem ampla distribuição no Brasil e é típico de florestas tropicais úmidas (Pennington 1997).
Dentre as vinte espécies mais frequentes nas áreas estudadas, algumas são de ampla ocorrência no domínio da Mata Atlântica, como Amaioua guianensis, Astronium fraxinifolium, Copaifera langsdorffii, Cordia trichotoma, Guarea guidonia, G. kunthiana e Siparuna guianensis (Oliveira Filho & Fontes 2000). Joannesia princeps e Virola gardneri, frequentes no presente estudo, têm sido relacionadas como típicas de floresta primária alta, desenvolvida, estratificada e com número representativo de árvores de grande porte. J. princeps é citada para florestas primárias e secundárias (Gentilini et al. 1981; Andrade et al. 1997; Lopes 2002). Neoraputia alba é frequente em um estudo fitossociológico realizado no PERD (Gentilini et al. 1981). A. fraxinifolium, C. langsdorffii, C. trichotoma, G. guidonia e G. kunthiana são típicas de florestas estacionais semideciduais de baixa altitude do Leste do Estado de Minas Gerais (Oliveira-Filho & Fontes 2000).
As diferenças observadas entre os valores de riqueza estimada e observada de espécies, em duas áreas (MACU e GAV), podem ser consequências de diversos processos históricos e ecológicos, que atuam ou atuaram em diferentes escalas de espaço e tempo (Tabarelli & Mantovani 1999a). A maior riqueza observada na primeira área (MACU) pode ser atribuída ao fato dela não ter sido afetada pelos distúrbios que ocorreram no médio rio Doce, como corte seletivo de madeira e grandes incêndios florestais. A menor riqueza observada na segunda área (GAV) deve-se a repetidos incêndios que tenderam a simplificar a composição de espécies e a estrutura (Silva et al. 2005). Segundo os mesmos autores, um incêndio florestal provoca a redução do número de árvores, da biomassa viva e do número de espécies por unidade de área, devido à morte das populações em parte ou no todo. O solo é outro fator importante a ser considerado na composição e riqueza de espécies (Oliveira-Filho et al. 2001; Botrel et al. 2002). Na área em questão ele distinguiu-se das demais pelo pH mais ácido e maior proporção de alumínio, limitando, aparentemente, o desenvolvimento de algumas espécies, e diminuindo a sua riqueza.
A redução da riqueza de espécies também é influenciada pela dominância de algumas pioneiras na composição florística da floresta. Mabea fistulifera e Cordia sellowiana, típicas de ambientes secundários, se destacaram em uma das áreas estudadas (GAV). M. fistulifera cresce naturalmente em capoeiras, clareiras de floresta e beira de estradas. Suas sementes germinam melhor em locais com alta temperatura e maior exposição do solo aos raios solares, com grande intensidade luminosa (Filho & Borges 1992).
Segundo Richards (1976), áreas que sofreram o corte seletivo apresentaram uma rica comunidade constituída de espécies de floresta primária e secundária, de estrutura bastante irregular, visto que as últimas ocuparam as clareiras formadas pela retirada dos indivíduos da comunidade. Estes fatores provavelmente afetaram duas florestas analisadas (MUMB e MACE) que apresentaram árvores de menor porte e diversidade elevada. O aumento da riqueza de espécies durante o processo de regeneração secundária também ocorre em florestas tropicais afetadas pela queima, onde são verificadas riquezas maiores nas florestas mais desenvolvidas, com espécies de diversas fases da sucessão, sendo observadas menos espécies nas florestas em estágios iniciais de sucessão secundária (Tabarelli & Mantovani 1999b; Pereira et al. 2007).
Nas florestas neotropicais, verifica-se relação direta entre a precipitação, a fertilidade dos solos e a riqueza de espécies. Ao contrário das florestas montanas, a elevada riqueza de árvores parece ser o padrão para as florestas neotropicais de terras baixas na América do Sul. Com o decréscimo da fertilidade, ou aumento da estação seca, observa-se redução da riqueza de espécies arbóreas (Tabarelli & Mantovani 1999a). Para as florestas analisadas no presente estudo estes fatores podem ter interferido na composição e riqueza de espécies.
Os índices de diversidade encontrados no presente estudo (3,66 a 4,10) são considerados elevados e estão em concordância com outros levantamentos realizados em florestas semidecíduas na região da bacia do rio Doce e no Estado de Minas Gerais (Meira-Neto et al. 1997 (4,23); Lopes et al. 2002 (3,98); Spósito & Stehmann 2006 (3,11 a 4,04); Lopes et al. 2009 (4,58) e Lacerda et al. 2011 (3,80).
A grande dessemelhança entre as comunidades arbóreas estudadas, mesmo aquelas muito próximas, indicou que as florestas podem apresentar diferenças estruturais e florísticas marcantes. Dessa forma, como abordado por Espírito-Santo et al. (2002), remanescentes pequenos espalhados na paisagem podem encerrar comunidades muito distintas, não devendo ser tratados como amostras semelhantes de uma totalidade antes homogênea. A região do médio rio Doce apresenta alta diversidade beta, mostrando que as comunidades florestais são distintas e necessitam de políticas de conservação mais rigorosas e de incentivos à preservação.
A maior similaridade florística entre duas florestas (CAM e SACR) deve-se provavelmente ao fato delas localizarem-se próximas a cursos d'água, apresentando características semelhantes às florestas ciliares da região. Isto provavelmente influenciou significativamente a composição do solo e a flora arbórea. A menor similaridade observada entre duas outras florestas (SACR e GAV) pode ser explicada por fatores como: diferenças de altitude (259 e 546 m respectivamente), presença e ausência de curso d'água, diferenças dos solos quanto à composição química (maior acidez e porcentagem de alumínio na segunda área) e granulométrica (maior porcentagem de areia fina na primeira e maior porcentagem de areia grossa na segunda), além do regime de distúrbios.
O destaque de Euphorbiaceae em três áreas estudadas (MUMB, MACE e SACR) pode ser atribuído à Joannesia princeps, que apresentou elevada dominância relativa, determinada pelo grande porte de seus indivíduos, e à Senefeldera multiflora, espécie com alta densidade e frequência relativas, encontrada nos diferentes estratos das florestas. Os maiores valores de importância de Euphorbiaceae, Rutaceae, Sapotaceae e Myrtaceae estão de acordo com estudos realizados em florestas estacionais semideciduais no PERD (Gentilini et al. 1981; Lopes et al. 2002; Lombardi & Gonçalves 2000; Gonçalves & Lombardi 2004; Pessoa & Oliveira 2006; Camargos et al. 2008; Souza 2008; Lacerda et al. 2011) e em Floresta Atlântica de Tabuleiro do Espírito Santo (Jesus & Rolim 2005).
A importância de Fabaceae, em cinco áreas estudadas, deve-se provavelmente aos valores elevados de dominância relativa de Anadenanthera colubrina, A. peregrina e Pseudopiptadenia contorta. Fabaceae e Euphorbiaceae estão entre as famílias mais ricas no estudo de uma Floresta Estacional Semidecidual Insular, em Ponte Nova, Alto Rio Doce (Meira-Neto et al. 1997). A. colubrina é relacionada neste trabalho entre as espécies de maior área basal e dominância.
Joannesia princeps e Virola gardneri se destacaram quanto ao valor de importância no presente estudo, por apresentarem elevada dominância. A primeira espécie também se destacou em outro trabalho realizado no PERD (Lopes et al. 2002), e a segunda teve elevado valor de importância na Floresta Atlântica de Tabuleiro, em Linhares, ES (Jesus & Rolim 2005). Outras duas espécies, Neoraputia alba e Senefeldera multiflora destacaram-se em duas florestas (MACU e SACR), por apresentarem características estruturais semelhantes, formando populações mais densas, ocupando os diferentes estratos e com frequência elevada nas unidades amostrais. A alta densidade e valor de importância de S. multiflora também foram observados em áreas de floresta submontana e aluvial na Reserva Biológica de Poço das Antas, RJ (Guedes-Bruni et al 2006; Pessoa & Oliveira 2006). Esta espécie se destacou ainda, quanto ao número de indivíduos e densidade absoluta, na floresta de Tabuleiro no Espírito Santo (Jesus & Rolim 2005).
Os elevados valores de importância (VI) de Mabea fistulifera e Cordia sellowiana encontrados no fragmento fora do PERD (GAV) podem ser relacionados à alta densidade, dominância e frequência das mesmas e por serem típicas de regiões que sofreram algum tipo de perturbação. O sucesso da primeira espécie deve-se, provavelmente, à grande capacidade de dispersão de suas sementes, à manutenção de um número significativo de plântulas e ao grande porte dos indivíduos adultos. C. sellowiana tem sido apontada como indicadora de floresta de altitude (Meira-Neto & Martins 2002). Apesar da área citada não atingir altitudes das florestas baixo e altomontanas, ela está situada em altitude maior do que as demais. Pseudopiptadenia contorta também se destacou quanto à dominância, devido à predominância de indivíduos de grande diâmetro e altura. Este fato está de acordo com alguns estudos fitossociológicos realizados em áreas semelhantes (Meira-Neto et al. 1997; Lopes et al. 2002).
A elevada dominância relativa de Chrysophyllum imperiale (SACR) deve-se ao grande porte dos seus indivíduos, apesar da baixa densidade e frequência relativa observadas. Esta espécie ocupou o segundo lugar em valor de importância. Por tratar-se de espécie ameaçada de extinção (Drumond et al. 2008; MMA 2008; IUCN 2012) e ser encontrada somente neste fragmento, fora da área do PERD, torna-se de suma importância a sua preservação.
A distribuição de frequência nas classes diamétricas, na maioria das áreas estudadas, indicou decréscimo acentuado do número de indivíduos, no sentido das menores para as maiores classes. Este padrão é o esperado para florestas tropicais, principalmente aquelas que sofreram ou sofrem perturbações antrópicas, além de ser característico de populações em "J" invertido (Harper 1990). Esta semelhança é apresentada em vários estudos de comunidades vegetais em florestas semidecíduas (Martins 1991; Lopes et al. 2002; Guedes-Bruni et al. 2006; Pessoa & Oliveira 2006; Spósito & Stehmann 2006; Lopes et al. 2009). Dessa forma, pode-se afirmar que as florestas analisadas estão em desenvolvimento na direção de estádios mais avançados de sucessão, visto que existiu um número considerável de jovens indivíduos que irão substituir aqueles mais velhos ou danificados. Somente duas florestas (CAMP e SACR) apresentaram uma frequência maior nas classes intermediárias e superiores, demonstrando serem áreas mais conservadas e melhor estruturadas.
As propriedades físicas e químicas dos solos das áreas estudadas contribuíram para analisar a heterogeneidade florística regional. Alguns trabalhos utilizam estas características como parâmetros ambientais para explicar a diferenciação de florestas no Estado de Minas Gerais, principalmente em áreas com grande heterogeneidade florística (Oliveira-Filho et al. 1994; Spósito & Stehmann 2006; Lopes et al 2009). Três áreas apresentaram solos do tipo eutrófico (CAMP, MACE e SACR), ricos em nutrientes, e três do tipo álico (MACU, MUMB e GAV), mais ácidos e pobres em nutrientes. As diferenças observadas provavelmente resultaram na diversidade de ambientes encontrados.
A proximidade ao ambiente lacustre também interferiu na composição de espécies de algumas florestas estudadas. Duas delas (MACU e MUMB) se diferenciaram das demais quanto à composição química (pH mais baixo) e granulométrica do solo (maior porcentagem de argila) e pela composição de espécies. Estes solos podem ser caracterizados como orgânicos, com baixa saturação por bases, elevada acidez e saturação por alumínio. Eles são formados por acumulações recentes de resíduos vegetais depositados sob excesso de água permanente, em áreas de várzeas, depressões e margens de cursos d'água encharcadas (Jacomine 2001). Richards (1976) comenta que nas regiões tropicais úmidas o caráter ácido ou básico é, frequentemente, de grande importância, visto que aproximadamente todos os solos são ácidos e marcadamente deficientes em bases. A presença de grandes quantidades de alumínio em solos tropicais é outro fator edáfico responsável pela diferenciação nas características estruturais da vegetação.
Três áreas estudadas (CAMP, MACE e SACR) localizam-se em regiões aluviais que recebem sedimentos carreados pelos cursos d'água, o que explica a grande quantidade de areia fina e matéria orgânica observada na composição dos solos. Estes são muito heterogêneos quanto à granulometria, estrutura, consistência e propriedades químicas. Eles podem ser ácidos, portanto com pH muito baixo e alta saturação por alumínio, ou básicos, com pH alto e alta saturação por bases (Jacomine 2001). Solos aluviais são muito variáveis a pequenas distâncias e normalmente são os mais ricos de uma paisagem. A disponibilidade de água e nutrientes afeta significativamente a distribuição das principais formas de vegetação; a deficiência de oxigênio (drenagem), por afetar o crescimento das raízes e a disponibilidade de nutrientes (toxidez), interfere no fornecimento de água e nutrientes. Estes dois fatores têm forte interação e nos lugares mais úmidos a ciclagem de nutrientes compensa a pobreza do solo (Rezende 2002).
Uma das áreas analisadas (GAV) apresentou características edáficas distintas das demais, com solo semelhante ao de Cerrado, Latossolo Vermelho Amarelo Álico, mais ácido, com alto teor de alumínio, rochoso e sem cursos d'água. Ele mostrou-se muito intemperizado, relação silte/argila próxima de 0,15, sendo que para a maioria dos Latossolos esta relação fica em torno de 0,7 (Rezende 2002). Segundo o autor, a fração silte serve como indicadora do grau de intemperização do solo ou do potencial dele de conter minerais primários facilmente intemperizáveis, isto é, de sua reserva em nutrientes. Estes fatores condicionaram o aparecimento de espécies adaptadas a esse ambiente e refletiram a baixa diversidade.
Considerando as correlações entre espécies e variáveis de solo, verificou-se que os autovalores intermediários para os eixos de ordenação 1 e 2 indicaram a ocorrência de gradientes longos, isto é, a maioria das espécies ocorre em regiões exclusivas de segmentos particulares (ter Braak 1995). A elevada variância (62%) contribuiu para maior significância das relações espécie-ambiente (ter Braak 1988). As correlações encontradas indicaram que as condições edáficas, representadas pelo índice de saturação de alumínio, quantidade de silte e areia grossa e valores de pH, provavelmente exerceram forte influência no padrão de distribuição das espécies no médio rio Doce.
As análises de componentes principais evidenciaram a heterogeneidade ambiental do médio rio Doce. Áreas mais conservadas, com características edáficas semelhantes, apresentaram valores elevados de área basal e altura (CAMP e SACR). A primeira área faz parte de um mosaico de vegetação representado pelo PERD e a segunda constitui reserva legal pertencente à Área de Proteção Ambiental (APA) Pingo d'Água. O PERD representa uma das maiores áreas protegidas de Mata Atlântica em Minas Gerais e está quase totalmente circundado por plantações de eucaliptos que compõem vários fragmentos do entorno.
Considerando as espécies arbóreas ameaçadas de extinção, seis constam nas listas de espécies ameaçadas da flora do estado de Minas Gerais e do Brasil (Drummond et al. 2008; MMA 2008). Na categoria vulnerável, estão presentes Astronium fraxinifolium (MACE, SACR e GAV), Dalbergia nigra e Melanoxylon brauna (CAMP, MACU, SACR e GAV). Da categoria em perigo, foram registradas Brosimum glaziouii (MUMB e GAV), Chysophyllum imperiale (SACR) e Ocotea odorifera (todas as áreas, exceto GAV). Dentre as espécies citadas três são encontradas na lista vermelha de espécies ameaçadas mundialmente: D. nigra, B. glaziouii e C. imperiale (IUCN 2012). Algumas destas espécies também ocorrem em áreas de preservação fora dos limites do PERD, o que demonstra a importância da conservação destas áreas disjuntas.
Agradecimentos
À CAPES, a bolsa de doutorado concedida ao primeiro autor; ao CNPq, o apoio financeiro ao projeto (PELD - Mata Atlântica e Sistema Lacustre do Médio Rio Doce) e a bolsa de produtividade em pesquisa concedida ao segundo autor; ao Prof. Dr. Francisco Antônio R. Barbosa, o suporte logístico; à Companhia Agroflorestal (CAF), à Prefeitura Municipal de Pingo d'Água, à empresa Celulose Nipo-Brasileira (CENIBRA), a colaboração e suporte nos trabalhos de campo; à administração do Parque Estadual do Rio Doce, a hospitalidade e colaboração; ao Prof. Dr. Júlio A. Lombardi, Drª Tereza Spósito e Dr. Marcos Sobral, que colaboraram nas identificações; à Renata M. Ferreira, Cíntia Tamara e Fernanda Raggi, a valiosa ajuda em campo e no laboratório; ao Sr. Geraldo dos Santos Adriano (Canela), o auxílio nas coletas.
Artigo recebido em 07/07/2012.
Aceito para publicação em 03/12/2012
Este artigo possui material adicional em sua versão eletrônica.
- Andrade, P.M; Pereira, M.C.A. & Silva, L. V. C. 1997. The vegetation of Rio Doce State Park. In: Tundisi, J.G. & Saijo, Y. (eds.). Limnological studies on the Rio Doce Valey lakes, Brazil. Academia Brasileira de Ciências, Rio de Janeiro. Pp. 15-21.
- Antunes, F.Z. 1986. Caracterização climática do estado de Minas Gerais. Informe Agropecuário 12:9-13.
- APG II. 2003. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. Botanical Journal of the Linnaean Society 141:399-436.
- Athayde, T. 2005. Ecologia e história natural da Mata Atlântica. Interciência, Rio de Janeiro. 197p.
- Bortoluzzi, R.L.C.; Carvalho-Okano, R.M.; Garcia, F.C.P. & Tozzi, A.M.G.A. 2004. Leguminosae, Papilionoideae no Parque Estadual do Rio Doce, Minas Gerais, Brasil II: árvores e arbustos escandentes. Acta Botanica Brasilica. 18:49-71.
- Botrel, R.T.; Oliveira-Filho, A.T.; Rodrigues, L.A. & Curi, N. 2002. Influência do solo e topografia sobre as variações da composição florística e estrutura da comunidade arbóreo-arbustiva de uma floresta estacional semidecidual em Ingaí, MG. Revista Brasileira de Botânica 25:195-213.
- Camargos, V.L.; Silva, A.F.; Meira Neto, J.A. & Martins, S.V. 2008. Influência de fatores edáficos sobre variações florísticas na Floresta Estacional Semidecídua no entorno da Lagoa Carioca, Parque Estadual do Rio Doce, MG, Brasil. Acta Botanica Brasilica 22:75-84.
- CETEC - Fundação Centro Tecnológico de Minas Gerais. 1981. Vegetação do Parque Estadual do Rio Doce. Vol. 2. Relatório Final. 288p.
- Drummond, G.M.; Machado, A.B.M.; Martins, C.S.; Mendonça, M.P. & Stehmann, J.R. (orgs.). 2008. Listas vermelhas das espécies da fauna e da flora ameaçadas de extinção em Minas Gerais. Fundação Biodiversitas, Belo Horizonte.
- EMBRAPA - Empresa Brasileira de Pesquisa Agropecuária. 1999. Atributos diagnósticos. In: EMBRAPA. Sistema brasileiro de classificação de solos. Centro Nacional de Pesquisa de Solos, Rio de Janeiro. 412p.
- EMBRAPA, 2007. Empresa Brasileira de Pesquisa Agropecuária. Mapa de solos do estado de Minas Gerais. Disponível em <http://www.cnps.embrapa.br/solosbr/conhecimentos>. Acesso em 8 Jul 2007.
- Espírito-Santo, F.D.B.; Oliveira-Filho, A.T.; Machado, E.L.M.; Souza, J.S.; Fontes, M.A. & Melo Marques, J.J.G.S. 2002. Variáveis ambientais e a distribuição de espécies arbóreas em um remanescente de floresta estacional semidecidual montana no campus da Universidade Federal de Lavras, MG. Acta Botanica Brasilica 16:331-356.
- Fidalgo, O. & Bononi, V.L.R. 1984. Técnicas de coleta, preservação e herborização de material botânico. Manual n ş 4. Instituto de Botânica, São Paulo. 62p.
- Filho, N.L. & Borges, E.E.L. 1992. Influência da temperatura e da luz na germinação de sementes de canudo de pito (Mabea fistulifera Mart.). Revista Brasileira de Sementes 14:57-60.
- Fonseca, G.A.B. 1997. Impactos antrópicos e biodiversidade terrestre. In: Paula, J.A. et al. (eds.). Biodiversidade, população e economia: uma região de Mata Atlântica. UFMG/Cedeplar, Belo Horizonte. Pp.455-468.
- Fundação SOS Mata Atlântica. Instituto Nacional de Pesquisas Espaciais. Instituto Socioambiental. 1998. Atlas da evolução dos remanescentes florestais e ecossistemas associados no domínio da Mata Atlântica no período de 1990-1995. Relatório Nacional - Síntese dos estados do Espírito Santo, Rio de Janeiro, Minas Gerais, Goiás, Mato Grosso do Sul, São Paulo, Paraná, Santa Catarina e Rio Grande do Sul. São Paulo. 55p.
- Fundação SOS Mata Atlântica. Instituto Nacional de Pesquisas Espaciais. 2010. Atlas dos remanescentes florestais da Mata Atlântica no período de 2008-2010. Relatório Nacional - Dados parciais dos estados avaliados até maio de 2010. São Paulo. Disponível em <http://mapas.sosma.org.br/dados>. Acesso em Dez 2010.
- Gentilini, E.; Carvalho, I.R. & Teixeira, M.C.B. 1981. Vegetação do Parque Estadual do Rio Doce. In: Programa de Pesquisas Ecológicas no Parque Estadual do Rio Doce. Vol. 2. Fundação Centro Tecnológico de Minas Gerais, Belo Horizonte. 285p.
- Gentry, A.H. 1995. Patterns of diversity and floristic composition in neotropical montane forests. In: Churchill, S.P.; Balslev, H.; Forero, E. & Luteyn, J.L. (eds.). Biodiversity and conservation of Neotropical Montane Forests. Neotropical Montane Forest biodiversity and conservation symposium 1.New York Botanical Garden, New York. Pp.103-126
- Gonçalves, M. & J. Lombardi. 2004. Adições ao conhecimento da composição florística de dois remanescentes de Mata Atlântica do sudeste de Minas Gerais, Brasil. Lundiana 5:3-8.
- Gotelli, N.J. & Colwell, R.K. 2001. Quantifying biodiversity: procedures and pitfalls in the measuring and comparison of species richness. Ecology letters 4:379-391.
- Gotelli, N.J. & Enstminger, G.L. 2006. Ecosim Null Models software for ecology. Version 7. Acquired Intelligence Inc. & Kesey. Jericho, VT 05465. Disponível em <http://garyentsminger.com/ecosim/index.htm>. Acesso em 13 Jun 2007.
- Guedes-Bruni, R.R.; Neto, S.J.S.; Morim, M.P. & Mantovani, W. 2006. Composição florística e estrutura de dossel em trecho de floresta ombrófila densa atlântica sobre morrote mamelonar na Reserva Biológica de Poço das Antas, Silva Jardim, Rio de Janeiro, Brasil. Rodriguésia 57:429-442.
- Guedes-Bruni, R.R.; Neto, S.J.S.; Morim, M.P. & Mantovani, W. 2006. Composição florística e estrutura de trecho de floresta ombrófila densa atlântica aluvial na Reserva Biológica de Poço das Antas, Silva Jardim, Rio de Janeiro, Brasil. Rodriguésia 57:413-428.
- Harper, J.L. 1990. Population biology of plants. Academic Press, London. 892p.
- IGA - Instituto de Geociências Aplicadas. 1982. Belo Horizonte (Mapa Geológico da Região Metropolitana de Belo Horizonte. Folha SE-23-Z-C-VI-3, escala 1: 50.000).
- Jacomine, P.K.T. 2001. Solos sob matas ciliares. In: Rodrigues, R.R.; Leitão-Filho, H.F.L. (eds.). Matas ciliares: conservação e recuperação. 2Ş ed. Ed. da Universidade de São Paulo, Fapesp, São Paulo. 320p.
- Jesus, R.M. & Rolim, S.G. 2005. Fitossociologia da Mata de Tabuleiro. Boletim técnico nş 19. SIF, Viçosa. Pp. 1-149.
- Lacerda, T.; Matos, F.A.R.; Gastauer, M.; Silva, M.C.N.A.; Moreira, L.C.B. & Meira-Neto, J.A.A. 2011. Fitossociologia de um fragmento de floresta estacional semidecidual limítrofe ao Parque Estadual do Rio Doce, Minas Gerais, Brasil. X Congresso de Ecologia do Brasil, São Lourenço.
- Leitão-Filho, H.F. 1992. A Flora arbórea da Serra do Japi. In: Morellato, L.P.C. (ed.). História natural da Serra do Japi. Ecologia e preservação de uma área florestal do sudeste do Brasil. Ed. da UNICAMP, Campinas. 40-62.
- Lombardi, J.A. & Gonçalves, M. 2000. Composição florística de dois remanescentes de Mata Atlântica do sudeste de Minas Gerais, Brasil. Revista Brasileira de Botânica 23:255-282.
- Lopes, W.P.; Silva, A.F.; Souza, A.L. & Meira Neto, J.A.A. 2002. Estrutura fitossociológica de um trecho de vegetação arbórea no Parque Estadual do Rio Doce - Minas Gerais, Brasil. Acta Botanica Brasilica 16:443-456.
- Lopes, R.M.F.; França, G.S.; Silva, F.RG.; Sposito, T.C.S. & Stehmann, J.R. 2009. Estrutura do componente arbóreo de uma floresta estacional semidecidual montana secundária no alto Rio Doce, Minas Gerais, Brasil. Rodriguésia 60:1037-1053.
- McCune, B. & M.J. Mefford. 1999. PC-ORD version 4.0., multivariate analysis of ecological data, users guide. MjM Software Design, Glaneden Beach.
- Martins, F.R. 1991. Estrutura de uma floresta mesófila. Ed. da UNICAMP, Campinas. 246p.
- Meira-Neto, J.A.A.; Souza, A.L.; Silva, A.F. & de Paula, A. 1997. Estrutura de uma floresta estacional semidecidual insular em área diretamente afetada pela usina hidrelétrica de pilar, Ponte Nova, Zona da Mata de Minas Gerais. Revista Árvore 21:493-500.
- Meira-Neto, J.A.A. & Martins, F.R. 2002. Composição florística de uma floresta estacional semidecidual montana no município de Viçosa - MG. Revista Árvore 26:437-446.
- MMA - Ministério do Meio Ambiente. 2008. Instrução normativa nº 06 de 23/09/2008. Lista nacional das espécies da flora brasileira ameaçadas de extinção. Disponível em <http://www.mma.gov.br>. Acesso em 15 Dez 2010.
- Nunes, S.R.D.F.S.; Garcia, F.C.P.; Lima, H.C. & Carvalho-Okano, R.M. 2007. Mimosoideae (Leguminosae) arbóreas do Parque Estadual do Rio Doce, Minas Gerais, Brasil: distribuição geográfica e similaridade florística na floresta atlântica no sudeste do Brasil. Rodriguésia 58:403-421.
- Oliveira-Filho, A.T.; Vilela, E.A.; Gavilanes, M.L. & Carvalho, D.A. 1994. Comparison of the woody flora and soils of six areas of montane semideciduos forest in southern Minas Gerais, Brazil. Edinburgh Journal of Botany 51:355-389.
- Oliveira-Filho, A.T. & Fontes, M.A.L. 2000. Patterns of floristic differentiation among Atlantic forests in southeastern Brazil and the influence of climate. Biotropica 32:793-810.
- Oliveira-Filho, A.T.; Curi, N.; Vitela, E.A. & Carvalho, D.A. 2001. Variation in tree community composition and structure with changes in soil properties within a fragmento for semideciduous forest in south-eastern Brazil. Edinburgh Journal of Botany 58:139-158.
- Oliveira-Filho, A.T.; Tameirão-Neto, E.; Carvalho, W.A.C.; Werneck, M.; Brina, A.E.; Vidal, C.V.; Resende, S.C. & Pereira, J.A.A. 2005. Análise florística do componente arbóreo de áreas de floresta atlântica sensu lato na região das Bacias do Leste (Bahia, Minas Gerais, Espírito Santo e Rio de Janeiro). Rodriguésia 56:185-235.
- Oliveira-Filho, A.T.; Jarenkow, J.A. & Rodal, M.J.N. 2006. Floristic relationships of seasonally dry forests of eastern south America based on tree species distribuition patterns. In: Pennington, R.T.; Lewis, G.P. & Ratter, J.A. (orgs.). Neotropical savannas and dry forests: plant diversity, biogeography and conservation. CRC Press, Boca Raton. Pp. 159-192.
- Oliveira-Filho, A. 2009a. Classificação das fitofisionomias da América do Sul cisandina tropical e subtropical: proposta de um novo sistema - prático e flexível - ou uma injeção a mais de caos. Rodriguésia 60:237-258.
- Oliveira-Filho, A. 2009b. TreeAtlan 1.0, flora arbórea da América do Sul cisandina tropical e subtropical: um banco de dados envolvendo biogeografia, diversidade e conservação. Universidade Federal de Minas Gerais. Disponível em <http://www.icb.ufmg.br/treeatlan/>. Acesso em 2 Jul 2012.
- Pedralli, G.; Teixeira, M.C.B. & França, E.G. 1986. Lauraceae do Parque Florestal Estadual do Rio Doce, Minas Gerais, Brasil. Ciência e Cultura 38:1414-1421.
- Pennington, T.D. 1997. The genus Inga: Botany. The Royal Botanic Gardens, Kew. 844p.
- Pereira, J.A.A.; Oliveira-Filho, A.T. & Lemos-Filho, J.P. 2007. Environmental heterogeneity and disturbance by humans control much of the tree species diversity of Atlantic montane forest fragments in SE Brazil. Biodiversity Conservation 16:1761-1784.
- Pessoa, S.V. & Oliveira, R.R. 2006. Análise estrutural da vegetação arbórea em três fragmentos florestais na Reserva Biológica de Poço das Antas, Rio de Janeiro, Brasil. Rodriguésia 57:391-411.
- Resende, M.; Curi; N.; Rezende, S.B. & Corrêa, G.F. 2002. Pedologia: base para distinção de ambientes. 4Ş ed. NEPUT, Viçosa. 338p.
- Ribeiro, M.C.; Metzger, J.P.; Martensen, A.C.; Ponzoni, F.J. & Hirota, M.M. 2009. The brazilian Atlantic Forest: how much is left, and how in the remaining forest distributed? Implications for conservation. Biological Conservation 142:1141-1153.
- Richards, P.W. 1976. The tropical rain forest, an ecological study. Facsimile da edição de 1952. Cambridge University Press, London. 450p.
- Rolim, S.G.; Ivanauskas, N.M.; Rodrigues, R.R.; Nascimento, M.T.; Gomes, J.M.L.; Folli, D.A. & Couto, H.T.Z. 2006. Composição florística do estrato arbóreo da floresta estacional semidecidual na planície aluvial do rio Doce, Linhares, ES, Brasil. Acta Botanica Brasilica 20:549-561.
- Shepherd, G.J. 1994. FITOPAC 1: manual do usuário. Universidade de Campinas, Campinas. 32p.
- Silva, V.F; Oliveira-Filho, A.T.; Venturin, W.A.C.C. & Gomes, J.B.V. 2005. Impacto do fogo no componente arbóreo de uma floresta estacional semidecídua no município de Ibituruna, MG, Brasil. Acta Botanica Brasilica 19:701-716.
- Sokal, R.R. & Rohlf, F.J. 1995. Biometry. W.H. Freeman, San Francisco. 887p.
- Souza, P.B. 2008. Diversidade florística e atributos pedológicos ao longo de uma encosta com floresta estacional semidecidual submontana, zona de amortecimento do Parque Estadual do Rio Doce, MG. Tese de Doutorado. Universidade Federal de Viçosa, Viçosa. 138p.
- Spósito, T.C. & Stehmann, J.R. 2006. Heterogeneidade florística e estrutural de remanescentes florestais da área de proteção ambiental ao sul da região metropolitana de Belo Horizonte (APA Sul-RMBH), Minas Gerais, Brasil. Acta Botanica Brasilica 20:347-362.
- Tabarelli, M. & Mantovani, W. 1999a. A riqueza de espécies arbóreas na floresta atlântica de encosta do estado de São Paulo, Brasil. Revista Brasileira de Botânica 22:217-223.
- Tabarelli, M. & Mantovani, W. 1999b. A regeneração de uma floresta tropical montana após corte e queima (São Paulo - Brasil). Revista Brasileira de Biologia 59:239-251.
- ter Braak, C.J.F. 1988. Canoco - a Fortran program for canonical community ordination by (Partial) (Detrended) (Canonical) correspondence analysis and redundancy analysis, version 2.1. Technical report LWA-88-2. TNO, Wageningen.
- ter Braak, C.J.F. 1995. Ordination. In: Jongman, R.H.G.; ter Braak, C.J.F. & van Tongeren, O.F.R. (eds.). Data analysis in community and landscape ecology. Cambrigde University Press, Cambrigde. Pp. 91-173.
- The Word Conservation Union - IUCN. 2012. Disponível em <http://www.iucn.org>. Acesso em 7 Dez 2012.
» link - Thomaz, L.D. & Monteiro, R. 1997. Composição florística da Mata Atlântica de encosta da estação biológica de Santa Lúcia, município de Santa Tereza-ES. Boletim do Museu de Biologia Mello Leitão 7:3-43.
- Zar, J.H. 1999. Biostatistical analysis. Prentice-Hall, New Jersey. 663p.
Datas de Publicação
-
Publicação nesta coleção
11 Nov 2013 -
Data do Fascículo
Set 2013
Histórico
-
Recebido
07 Jul 2012 -
Aceito
03 Dez 2012