
Abstract
In 2015 a chance discovery of the beach moon flower in coastal Brazil led to an investigation of the global occurrence, distribution, and abundance of this pantropical littoral plant species. We here document new distribution records for coastal Brazil and West Tropical Africa; postulate a human-mediated long-distance dispersal for this species from the Indo-Pacific to the Atlantic, followed by local distribution via ocean currents; and provide historical context on the name confusion with other species. We also point out the risks inherent in using specimen information available on the internet without adequate verification for the identity of the specimens as a necessary first step.
Key words:
biogeography; drift seed; global change biology; pantropical; plant dispersal
Resumo
Em 2015, uma descoberta acidental da “beach moon flower” no litoral do Brasil levou a uma investigação sobre a ocorrência, distribuição e abundância dessa espécie de planta litorânea pantropical. Aqui documentamos novos registros de distribuição para o litoral brasileiro e a África tropical ocidental, postulamos uma dispersão de longa distância mediada por humanos para essa espécie, do Indo-Pacífico ao Atlântico, seguida da distribuição local através de correntes oceânicas e contextualizamos acercada confusão do nome com outras espécies. Apontamos também os riscos inerentes do uso de informações sobre espécimes disponíveis na internet, sem que haja a devida identificação desses espécimes como um primeiro passo necessário.
Palavras-chave:
biogeografia; semente de deriva; mudanças globais biológicas; pantropical; dispersão de plantas
Introduction
In 2015 a routine plant collecting trip in coastal Pernambuco state, Brazil, found a thriving population of Ipomoea violacea L. (Convolvulaceae) growing in the littoral strand in a private nature reserve (Reserva Particular do Patrimônio Natural [RPPN] Fazenda Tabatinga), in Goiana Municipality. This did not seem significant at first, because I. violacea, the beach moon flower, is known to grow on tropical beaches almost world-wide, but later research showed that this pantropical coastal twiner had not, in fact, been documented anywhere in Brazil except the Fernando de Noronha islands (Flora do Brasil 2020), some 350 kilometres offshore from mainland Pernambuco. For that reason - what appeared to be the first mainland record of this species in Brazil - the specimen collected at Fazenda Tabatinga began to capture our interest. As further research was carried out, the details became more intriguing and led us deeper into some biogeographical puzzles.
Soon after our discovery of I. violacea at Fazenda Tabatinga, herbarium visits to several Brazilian herbaria by GS during 2016 (ALCB, CEN, CGMS, HUEFS, INPA, IPA, UB) followed up with consultation of virtual herbaria (SpeciesLink 2017; REFLORA - Virtual Herbarium 2019) by JA and GS led to further specimen evidence that Ipomoea violacea is more widespread in Brazil than previously thought or documented in floras, checklists, and online references. At a later time, we expanded the geographic scope of study from Brazil to western coastal Africa because similar discontinuities were noted by AB for I. violacea in the observed versus reported distribution in that part of the world.
This paper sets out to explore the global distribution for Ipomoea violacea L., put these new distribution records in context, provide some necessary caveats about identification and naming, which have broader ramifications for information abstracted from the literature as well as for online specimen records. We synthesize the information available about I. violacea and propose a dispersal route and mechanisms that account for the current global distribution as documented by voucher specimens identified for this study.
Taxonomic synonyms and misapplied names
The beach moon flower has had a complex and confusing nomenclatural history even though the species is distinctive and easily recognizable everywhere it grows. The currently accepted scientific name for it is Ipomoea violacea L., a name that only began to gain acceptance after Manitz (1977) provided a lectotype for the Linnean name and unambiguously fixed its application. Prior to this typification the botanical and ecological literature mentioning the beach moon flower used names such as I. longiflora R. Br., I. macrantha Roem. & Schultes, and I. tuba (Schltdl.) G.Don (Austin 1975, 1980, 1982a, b, c; Berhaut 1967, 1975; Fosberg & Sachet 1977; Gunn 1972; Khan 1985; Ooststroom 1940; Ooststroom & Hoogland 1953; Powell 1979; Ridley 1890; Sachet 1975; Verdcourt 1958), all now regarded as synonyms, and sometimes the confused name “I. grandiflora”, which has been applied to multiple species by different authors (Powell et al. 1978). Manitz’ (1977) lectotypification clarified the taxonomy and synonymy for this widespread beach plant and began to stabilize the scientific name for it.
Complicating matters is a pernicious confusion in the published literature that wrongly uses the name Ipomoea violacea for an entirely different species. This confusion began with ethnobotanical studies of two narcotic species of Convolvulaceae used in Mexico and the taxonomic identifications for the plant species involved. MacDougall (1960) correctly identified one of these species, used among the Zapotec people for medicinal and ritual purposes because the seeds are hallucinogenic, as Ipomoea tricolor Cav. Schultes (1964) took exception to this usage: he conflated two very different species and applied the name “Ipomoea violacea” to this Mexican narcotic plant. For the rest of his career Schultes insisted this hallucinogenic plant should be called “Ipomoea violacea”; however, the rules of nomenclature do not allow for this usage after the lectotypification made by Manitz (1977). The hallucinogenic species is correctly known today as Ipomoea tricolor Cav. (Fig. 1). There is now a vast chemical and pharmaceutical literature about the secondary compounds and their biological properties for I. tricolor, which is also of horticultural importance: I. tricolor is grown in gardens throughout the world under cultivar names such as ‘Heavenly Blue’, ‘Pearly Gates’, and ‘Flying Saucers’. Sadly, the misapplication of the name “Ipomoea violacea” continues and even recent papers that describe fungal symbionts (Ahimsa-Müller et al. 2007) of ergoline alkaloid-positive Convolvulaceae used the wrong scientific name for the host plant. To be clear, our study concerns only the beach moon flower, the genuine Ipomoea violacea L., which has no known hallucinogenic properties, and we do not further discuss Ipomoea tricolor Cav., which has been so inextricably confused with it as far as the scientific names go.

a-c. Comparative diagnostic morphology for three Ipomoea species often confused under the same name - a. Ipomoea violacea flowers - inset: immature fruit showing rounded sepal shape; b. Ipomoea alba flowers; inset - immature fruit showing distinctly awned sepals; c. Ipomoea tricolor flowers - inset: calyx from a herbarium voucher showing triangular sepal shape and distinctive paler margins. Photo credits: a. Ipomoea violacea - flower: Imin Kamin [Imin K. et al. FRI-70039 (KEP)]; inset of fruit: G. Staples, not vouchered; b. Ipomoea alba - flower: G. Staples, cultivated, not vouchered; inset of fruit: Koykarn Kanjana, not vouchered; c. Ipomoea tricolor - flower: G. Staples, cultivated, not vouchered; inset: Gaumer & sons 23456 (GH).
In addition to the confusion with I. tricolor, Ipomoea violacea is not infrequently confused with, and wrongly identified as, I. alba L., which is also a night-flowering species with a similar corolla morphology and coloration (Fig. 1). Ipomoea alba also now has a pantropical distribution, but is intolerant of saline habitats (ocean beaches, mangroves, etc.) and is more typically found growing in inland habitats that are moist to wet; it has been introduced to the tropics globally (including much of Africa) as an ornamental and is listed as an invasive species in a number of different references (Randall 2017; IUCN ISSG 2019; CABI 2019). Ipomoea alba has the potential to become a serious weed of rainforest gaps and margins due to its capacity to spread aggressively from cultivated sources into surrounding natural areas. Figure 1 compares the floral morphology for the three species and Table 1 summarizes the diagnostic morphological characters that can be used to distinguish them.
Oceanic dispersal of seeds/fruits and Ipomoea violacea
Transport of fruits and seeds via oceanic currents is a well-documented plant dispersal phenomenon (Ridley 1930). In fact, collecting drift seeds on tropical beaches is a popular past time for many people (Gunn & Dennis 1976). The principle ocean currents that are responsible for transport from source areas to collection sites are also well-documented and understood (Fig. 2). In general, viability for tropical drift seeds and fruits is low: many of these propagules are dead when they drift for long periods of time (months, years) in salt water and are exposed to intense ultraviolet radiation by the time they reach beaches far outside the tropical climate regions (Gunn 1977). However, localized dispersal over shorter distances and time frames can be a highly effective method for some tropical plant species to increase their distribution and establish new populations. It is this dispersal via seawater flotation that provides the standard explanation for understanding the distribution of Ipomoea violacea on tropical beaches world-wide.

Global marine currents. Warm currents are shown in red; cold currents are shown in blue; isothermal currents are in black. This image is downloaded from Windows to the Universe® <http://windows2universe.org> © 2010 National Earth Science Teachers Association.This image is licensed under a Creative Commons Attribution- NonCommercial-ShareAlike 3.0 Unported License.
Seeds float by a variety of different mechanisms (Gunn 1972, 1977); in the case of I. violacea flotation is due to a cavity inside the seed that forms as the embryo matures (Fig. 3). This condition varies from seed to seed: as Ridley (1930) noted, some seeds sink, others float. The ability to float is entirely dependent on whether the cavity develops inside the testa during seed maturation and how large the cavity becomes. In terms of duration of flotation Ridley (1930: 303, quoting Guppy) stated that the seeds of I. violacea might sink in seawater after six or seven days, but can float for as long as 42 days (six weeks). Ridley stated that Guppy successfully germinated a seed of I. violacea after it floated in seawater for one year. We traced this claim to Guppy (1891: 306) in a post-script to his article about plant dispersal as exemplified by the flora of Cocos-Keeling island in the Pacific. Guppy collected various seeds and fruits in 1888, both fresh and in the beach drift on Cocos-Keeling, and brought them back to London where the flotation and subsequent germination experiments were conducted in 1890-1891. In the post-script to his published article he mentions the single seed of I. violacea germinated after floating one year in seawater; the flotation experiments were subjected to three weeks of winter cold at below freezing temperature. It is remarkable that a seed of a tropical plant species could survive not only such prolonged exposure to seawater but a degree of cold that would never occur in the wild, yet remain viable.

Seed of Ipomoea violacea showing central cavity (air space) inside the testa. Photo: J. Alencar; [Staples et al. 1715 (PEUFR)].
According to Nelson’s (1978) definitions for the transport of drift seeds/fruits, three broad categories are recognizable:
-
local dispersal - seeds/fruits spread in the immediate vicinity of the beach where they are found;
-
refuse dispersal - those discarded by humans onto the beach, washed overboard from ships, washed into the sea from refuse dumps;
-
peregrine dispersal - seeds that are transported long-distance by drifting in ocean currents.
Not included in Nelson’s categories is ballast water: modern ships rely on ballast water for maintaining trim and steerage. This is a relatively recent technological advance; more will be said about ballast employed in sailing ships in the Discussion. When referring to the third category, Darwin (1859) affirmed that this mode of transport would not occur in a random way, because the ocean currents are not random; he called this mode of distribution “casual”. In his famous “On the Origin of Species” Darwin (1859) calculated that such transport of seeds would be possible for distances of, at most, a few hundred kilometers, and consequently he believed that intercontinental dispersal was impossible by this means.
With this background on seed dispersal via ocean currents we attempt to account for recent discoveries for Ipomoea violacea along the coast of Brazil and the west coast of tropical Africa and to put these new distribution records into a larger context for the global distribution.
Material and Methods
When the new collections of Ipomoea violacea were made at Goiana, Pernambuco, we began our analysis of the species’ distribution by combining taxonomically verified specimen records from three geographically organized databases in BRAHMS software (Brahms 2002) compiled by GS between 2002-2016 into a single global distribution map for this species (Fig. 4). Maps were generated directly from the BRAHMS specimen data using the DIVA-GIS mapping software interface (DIVA-GIS 2014). These three databases for Convolvulaceae specimens examined in the course of taxonomic, floristic, and systematic studies of the family are broadly organized for Asia and the Pacific; Africa and Madagascar; and the Americas (North, Middle, Caribbean, and South). After consolidating the bona fide records for Ipomoea violacea into a global distribution (Fig. 4), some apparent gaps in the distribution were noted: Pacific west coast of the Americas; north and east coasts of South America; Atlantic west coast of tropical Africa. We had to then determine if the perceived gaps represented genuine absences of the species, or whether the gaps were due to lack of specimen data in our databases.

To test this hypothesis we turned to virtual herbaria, firstly for Brazil (SpeciesLink 2017; REFLORA - Virtual Herbarium 2019), and then later for several large herbaria: K, L, MO, NY, P, and WAG each of which maintains an institutional “virtual herbarium” displaying a large number of digital records for plant specimens from its own collections, with or without images of the specimen sheets. We searched each of these virtual herbaria for specimen records using the accepted name, Ipomoea violacea, as well as the principal synonyms: I. longiflora, I. macrantha, and I. tuba. We searched specifically by geographical criteria for countries/regions in the perceived gap areas. There being no practical method to search records that were misidentified, we did not attempt this systematically but did randomly discover some misidentified sheets of I. violacea while examining digital images. Records were created in GS’ databases for these newly discovered specimens of I. violacea examined as digital images.
We then developed a species profile for genuine Ipomoea violacea that allowed us to filter out specimen records returned in these searches that could be other species. The following morphological and ecological criteria apply: genuine Ipomoea violacea is a littoral species and typically grows near saltwater or brackish water bodies - it is seldom found far from sea beaches or coastal mangrove habitats, at altitude less than 100 m above sea level (Fang & Staples 1995; Staples 2010; Ooststroom 1940; Staples & Syahida-Emiza 2015), where it often grows over rocky outcrops or twines in littoral thickets (Ridley 1930) or on top of coastal cliffs (Fosberg & Sachet 1977); I. violacea rarely or never trails on sandy beaches or dunes (Ridley 1930). The flowers are large, showy and have been described as yellowish white; the flowers are nocturnal and odorless (contrasting with other night-flowering Ipomoea species such as I. alba L., I. muricata (L.) Jacq., and I. aspera Choisy, for example, which have night-fragrant flowers). Label data that do not correspond closely with this profile were considered suspect and required close inspection of the specimen or a digital image, if available, to verify the identity for the specimen sheets.
Three possible outcomes could be anticipated for records returned from virtual herbaria searches:
-
genuine Ipomoea violacea;
-
misapplications of Ipomoea violacea that are actually I. tricolor;
-
misidentifications that are some other species, neither I. violacea nor I. tricolor.
When records were returned from searches, it was then essential to determine whether the specimens were bona fide Ipomoea violacea, the misapplications of that name for what is actually I. tricolor, or a specimen of some other species that is wrongly identified. Preference was given to records with an image attached, so that the identity of the specimen could be taxonomically verified. Where an image was available the species (I. violacea, I. tricolor) are easily told apart (Fig. 1); when no specimen image was linked to the database record we excluded records with plant descriptors that did not correspond to I. violacea (such as diurnal flowers with bright blue corollas) or because the habitat reported by the collector was wrong (inland habitats far from ocean beaches).
The georeferenced data for collections of Ipomoea violacea validated by this study are made available for download as supplementary data to this article at <https://doi.org/10.6084/m9.figshare.10061948.v2>.
Results
In evaluating specimen records found in online searches we found several records called Ipomoea violacea that are suspect and were discarded: these exclusions are explained in the Discussion. Based on specimen records we believe are bona fide I. violacea, there are no trustworthy reports for I. violacea anywhere on the Pacific coast of the American continents. In marked contrast, I. violacea is widespread on the circum-Caribbean coasts of the Americas, with one long gap in Mexico, discussed below, and throughout the islands of the Greater and Lesser Antilles. A literature report (Hatch et al. 1990) for I. violacea in extreme southern Texas, USA, proved to be accurate: a voucher specimen was located in SMU that documents this occurrence. We will say more about these Caribbean populations in the Discussion section.
We also found supporting voucher specimens to document the distribution of I. violacea along the northern coast of South America. Literature reported the species as present in Venezuela (Austin 1982a), and the Guianas (Austin 1997); we found corroborating vouchers from Guyana and Surinam, but not for French Guiana. A request to the curator at CAY did locate a single recent collection (Girault et al. 1569, in 2016) that documents the presence of I. violacea on the coast of French Guiana. Furthermore, we did find several records that appear to be genuine I. violacea for eastern and northern Brazil and west tropical Africa, and those are discussed here. All supporting voucher specimens that document new distribution records are cited in the List of Exsicates at the end of this paper.
1. New distribution records for Ipomoea violacea in Brazil
The presence of I. violacea in the continental portion of Brazil seems to be recent, based on the collection dates for voucher specimens. The earliest documented collection in Brazilian territory was made in 1873, during the Challenger Expedition that visited several islands of the Fernando de Noronha archipelago (Fig. 5a). It was collected again at Fernando de Noronha by Ridley in 1887 during a study of the phanerogamic flora of the archipelago (Ridley 1890). For several decades after that no other occurrences were recorded. More recent records of I. violacea on the islands date from 1955-2003 (Freitas 2007).

a. (top) Archipelago of Fernando de Noronha, Pernambuco, Brazil. Localities where Ipomoea violacea was collected between 1873-2003; vouchers are cited in the List of Voucher Specimens at the end of this paper. b. (bottom) Baía de Todos os Santos, Bahia, Brazil. Localities where Ipomoea violacea was collected in 2010 and 2012, both within the Salvador metropolitan region; vouchers are cited in the List of Voucher Specimens at the end of this paper.
In 2015, when our team conducted field work along the coastal area of Pernambuco, we found a thriving population of I. violacea in the municipality of Goiana. It was in the beach area, twining in coastal thickets and climbing to 3-4 m high in the trees. Later we discovered that the same population had been documented previously in 2011.
When cataloging the collections of the project “Convolvuláceas da América do Sul” we verified that this was the only collection of I. violacea for mainland Brazil made by our team. In 2016, while identifying herbarium specimens in the Herbário Alexandre Leal da Costa (ALCB), GS discovered two collections of I. violacea from Bahia that were misidentified as Operculina. These specimens showed that the known distribution of I. violacea extended by at least 800 km further south in coastal eastern Brazil, without any records between these two points.
Interestingly, the location of the two Bahian collections (Fig. 5b) is within the bay of Baía de Todos os Santos, in the Metropolitan Region of Salvador, an important port area. This bay extends 80 km into the interior of the continent, is bordered by approximately 300 km of continental shoreline, and the entrance of the Baía is flanked by the ports of Aratu and Barra. For a better understanding of the hydrographic formation see Figure 5b.
In 2017 photos of living plants were shared on social media, asking for an identification of an Ipomoea found in Refúgio da Vida Silvestre (RVS) Santa Cruz, a federal conservation unit (UC) located in the municipality of Aracruz, state of Espírito Santo. This plant was identified by JA as I. violacea, based on its striate and glabrous stems, cordiform leaves, persistent glabrous sepals and salverform whitish corolla. We contacted the management of the RVS Santa Cruz and requested more photos and geographic coordinates of the exact location of the plant. A voucher specimen was requested from this population, but this requires collection authorization issued by the Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio). To date permission has not been given for a voucher specimen to be collected so this Espírito Santo population is being documented by the photos and coordinates provided (L.M. Coser, personal communication 2017).
In early 2020, late in the revision of this manuscript, a specimen record was found in SpeciesLink (2020) for a genuine collection of I. violacea from Amapá state; the specimen is misidentified as Ipomoea alba but there is a digital image attached to the record that allowed us to verify the correct identity. In addition to a voucher specimen from French Guiana (newly documented here) this Amapá record extends the distribution for I. violacea along the northern coast of the South American continent. There remains a sizable gap - ca 2,000 km - along the northern coast of Brazil where I. violacea does not occur, so far as known, between Amapá (newly documented here) and Paraíba state (Fig. 6), littoral zone where I. violacea might be found. While it is possible that ocean currents here (Fig. 2) are unfavorable for dispersal of I. violacea seeds from either the Caribbean population or the Fernando de Noronha population, it seems more likely that this gap is actually due to lack of collections. We point out the gap in hopes of encouraging plant collectors to search for I. violacea on ocean beaches and in mangrove ecosystems along the northern Brazilian coastline. We might confidently expect them to find thriving populations of I. violacea there.

2. New distribution records for Ipomoea violacea on the Atlantic coast of West Africa
When we began to explore the global distribution for Ipomoea violacea the initial mapping of specimen data in GS’ database presented a gap for the entire west coast of Africa (Fig. 4). This was in sharp contrast to the eastern coast of Africa, where several distribution points were noted on the continent, on Madagascar, and scattered across the Indian Ocean islands as far as India and Sri Lanka. This led us to investigate literature reports and specimen records to determine whether the West African gap was genuine, or simply due to lack of data in our database.
Verdcourt (1958: 216) first recorded Ipomoea violacea L. in West Tropical Africa, under the synonym I. tuba (Schlecht.) G.Don, citing a specimen collected by Irvine in Ghana in 1934 (K!, see the List of Voucher Specimens at the end of this paper). Verdcourt stated “It does not appear to have been recorded from W. Tropical Africa before”. He also pointed out the widespread and pernicious confusion of I. tuba with Ipomoea species in sect. Calonyction (e.g., I. alba, I. muricata).
Heine (1963: 352) subsequently mentioned I. tuba in the second edition of the “Flora of West Tropical Africa”, including it in a list of Ipomoea species cultivated in the region. In the “Useful Plants of West Tropical Africa” (Burkill 1985), this species is listed as “a perennial twiner of the American tropics, widely distributed as an ornamental”. See below for comments on cultivation of I. violacea.
Ipomoea violacea was recorded by Berhaut (1967, as I. tuba) in the “Flore du Sénégal” with the first voucher specimen collected in 1960. Berhaut (1967: 273) cited three of his own collections: 5668, 5671, 5673 that document the occurrence in Sénégal. We were able to examine a digital image for Berhaut 5668 in the BR herbarium; the other two collections were not located. In the expanded family account for Convolvulaceae that appeared in “Flore Illustrée du Sénégal” (Berhaut 1975: 167) he described the plants as forming large stands and climbing over the mangrove trees.
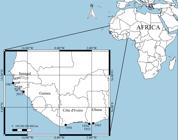
Searches in the Kew herbarium by AB located voucher specimens from Ghana collected more or less contemporaneously (1952 and 1965) with these literature reports. The Ghanaian specimens were cited by Gunn (1972: 160, as I. macrantha Roem. & Schultes), with one erroneously being stated as collected from Guinea. Online searches of virtual herbaria with strong African collections (BR, WAG) located additional specimens from Cote d’Ivoire and Sénégal that could be verified as genuine I. violacea.
In 2013, a specimen of Ipomoea was collected in Guinea; this specimen from Taide Island in Boké prefecture was subsequently identified at Kew as Ipomoea violacea L. The specimen was collected in an area where the plant was unlikely to have been dispersed by man (C. Couch, personal communication). In fact, it was this recent collection of I. violacea that prompted further research into the presence of the species in West Africa, an independent development parallel with the investigations for Brazil.
Except for the first record from Ghana, all West African specimens were collected after 1950, with the majority of records coming in the 1950s and 1960s (Fig. 7). These cover a stretch of coastline in excess of 2,000 km and there is a large distance between distribution points. All locations align well with sites of mangroves in the West African region and this is reinforced by Berhaut’s (1975) observations that it grows in the mangrove ecosystem and climbs on the species found there.
Discussion
These investigations demonstrated that some of the perceived distribution gaps for I. violacea (continental mainland of Brazil, West Tropical Africa) were actually due to absence of specimen data. Dedicated searching in herbaria and online virtual herbaria located specimens that filled those gaps. The collection dates for specimens from mainland Brazil and West Tropical Africa hint at a recent introduction for I. violacea into both areas, followed by local dispersal via seed flotation in the ocean, driven by prevailing currents. We propose a hypothesis for the long-range dispersal of I. violacea into the northern and eastern Brazil and West Tropical African regions, and then speculate about the global pantropical distribution for this species.
Is the pantropical distribution for Ipomoea violacea natural?
The majority of botanical references we examined stated that I. violacea (or one of its synonyms) is pantropical in distribution (for example, Gunn 1972; Sachet 1975; Fosberg & Sachet 1977; Austin 1975, 1980, 1982a, b; Powell 1979; Heine 1984; Bosser & Heine 2000; Deroin 2001; Staples & Syahida-Emiza 2015). Other publications made no comment on the status of I. violacea in the local flora (for example, Ooststroom & Hoogland 1953; Khan 1985; Gonçalves 1987; Liogier 1994; Staples 2010). Only one reference (Acevedo-Rodríguez 2005) provided a more precise statement regarding the distribution, indicating that I. violacea is native in the Caribbean and is “introduced in the tropics of the Old World”. But was this species naturally and genuinely pantropical, or has its contemporary global distribution been effected by human dispersal?
The reported pantropical distribution rests on the widespread presence of I. violacea throughout the tropical Indo-Pacific as well as in the Caribbean - at the present time two geographically isolated bodies of water - with several noteworthy gaps elsewhere around the globe, as discussed herein. How one interprets this pattern is the crux of the matter: most botanical authors have interpreted the reported distribution as a natural one for a coastal littoral species; only one (Acevedo-Rodríguez 2005), offers a different interpretation for the observed distribution. If the pantropical distribution is natural and not human-influenced, then two possible scenarios could account for it: either I. violacea was present in both the Indo-Pacific and Caribbean before the closure of the Central American Seaway by the rising land mass that is now the Isthmus of Panama (there is no common agreement on when this happened: estimates vary from 16 million years ago (MYA) (Montes et al. 2015) to 3.5 MYA (Hoorn et al. 2010) to as recently as 2.8 MYA (O’Dea et al. 2016)); or, alternatively, I. violacea dispersed by natural means from some starting point to the places where we find it today, making its way around the continental land masses (presumably via ocean currents) to establish populations that are widely disjunct. If the latter scenario is true, then the place of origin could be the Caribbean, as stated by Acevedo-Rodríguez (2005), or the tropical Indo-Pacific, with equal likelihood. However, it seems highly anomalous that if I. violacea were originally a Caribbean native, and dispersed from there to the Old World, the gap along the north coast of Brazil, the demonstrably recent arrival on the east coast of Brazil, and the total absence of the species along the western coast of the Americas are very hard to explain. One would reasonably expect thriving populations in all coastal littoral areas with suitable habitat between the Caribbean and the tropical Indo-Pacific. These enormous gaps in the distribution point toward another explanation for the observed global distribution.
In light of our examination of the recent distribution changes for I. violacea in Brazil and the Atlantic coast of Africa, we have come to wonder whether another explanation for the observed pantropical distribution is possible. After consideration of some relevant questions, we propose a hypothesis to explain the global distribution for the species.
How did Ipomoea violacea reach Brazil?
Theoretically at least, it seems possible that seeds of I. violacea could have reached the Brazilian coast by drifting with the ocean currents. We know that the Fernando de Noronha archipelago is subject to the influence of the South Equatorial Current (Barcellos et al. 2016), a cold and deep current that originates on the west coast of Africa (Fig. 2). With the east-west bifurcation of the South Equatorial Current, one branch, the Brazil Current [BC], a warm and superficial current, reaches the northeastern coast of Brazil; the other branch, the Guiana Current, flows northwestward along the coast of South America and then into the Caribbean Sea. The BC then flows along the eastern coast of Brazil towards the south until reaching the Zone of Subtropical Convergence where the BC converges with the Malvinas Current and then moves away from the Brazilian coast (Silveira et al. 2000).
We note here a curious fact: the records of I. violacea from Fernando de Noronha are all collected from the beaches of “Mar de Dentro” the side of the islands facing the North Atlantic and the coast of Brazil (Levy et al. 2016) and not from the “Mar de Fora” the side of the islands facing the African continent. Ridley (1890: 46, as I. tuba) pointed out that all the known populations of I. violacea at Fernando de Noronha were found on the north side of the islands and he presumed that the seeds had drifted onto the islands from the north.
Perhaps more significantly Fernando de Noronha receives, through these Atlantic Ocean currents, a great amount of floating debris; this marine trash can travel long distances and form sizable agglomerates. Among this trash are various types of waste and according to a study by Levy et al. (2016), most of this trash originates from countries of West Africa and even as far away as central, southwest and southeast Asia. According to the authors the anti-clockwise rotation of waters in the South Atlantic Ocean would be responsible for carrying agglomerations of debris from Africa (along with floating marine algae) toward the Fernando de Noronha archipelago.
Thus, it is theoretically possible for seeds of I. violacea originating from West Africa to have floated across the Atlantic via the South Equatorial Current to reach Fernando de Noronha. Yet the dates of first collection do not support this dispersal method, because I. violacea was documented from Fernando de Noronha as early as 1873, while the first collection from West Africa (Ghana) was only made in 1934. Given the long history of European exploration of the west coast of Africa - from 1434 onward - with botanical collecting beginning from the late 17th century and becoming more concerted from the 19th century onward, it seems unlikely that I. violacea could have been overlooked by botanical collectors for centuries, if it were actually there. The evidence available from dates of first voucher specimen collection suggests that I. violacea arrived later in West Tropical Africa, after it was already established at Fernando de Noronha.
Has cultivation been a factor in the current distribution of Ipomoea violacea?
With regard to the distribution of I. violacea in Africa, there are questions about whether this species has been present but not collected or is a recent colonist, either via natural means or by human introduction. The “Useful Plants of West Tropical Africa” (Burkill 1985) indicated that this species was used as an ornamental, but the paucity of detail surrounding its use and contradictory references make this uncertain, especially as it was generally unsuccessful in cultivation. Given that I. violacea is often confused with I. alba, this statement may be due to a case of mistaken identity or supposition that this species was also desirable as an ornamental.
Although some publications (Heine 1963; Burkill 1985) mention cultivation as responsible for the presence of I. violacea in Africa, this seems suspicious: in our experience (spanning more than 30 years for GS) I. violacea has seldom been cultivated anywhere. We explored several horticultural encyclopedias and reference manuals to learn whether I. violacea is typically included as a cultivated species. Although the genus Ipomoea was nearly always included there were few mentions of this species: I. violacea, or one of its synonyms, was included by Bailey (1935), Verdcourt (1970), Liberty Hyde Bailey Hortorium (1976), Huxley (1992); significantly, some of these authors commented that the species “… is not very successful in cultivation.” (Verdcourt 1970: 152) or “is not widely cultivated” (Huxley 1992). Ipomoea violacea was not mentioned at all in other horticultural references from the twentieth century: Bailey (1901); Synge (1956); Philip & Lord (1988, 1990, 1994); Griffiths (1992); Cullen et al. (2000). Gunn (1972: 161) stated (with reference to I. violacea) that “… its ornamental value has apparently not been explored.” These same horticultural references regularly included I. alba as an ornamental. Our conclusion is that the mention of cultivation as a factor in the introduction of I. violacea into West Africa is probably erroneous and is likely based on confusion with I. alba, as pointed out by Verdcourt (1958), Gunn (1972), and others. The moonflower, Ipomoea alba L., has been globally dispersed by humans as an ornamental flowering climber and then has naturalized virtually everywhere it has been introduced.
Berhaut (1975) also stated his belief that if I. violacea is American in origin it was probably dispersed naturally to Sénégal, in contrast to the views expressed by Heine (1963). Gunn (1972) expressed the same belief when he stated that it “has had little, if any, assistance from man except in recent years” in its dispersal.
In the Brazilian case, there is some specimen evidence to document that deliberate introduction by humans for ornamental purposes did occur, at least once: Dárdano de Andrade-Lima collected I. violacea in 1955 on the island of Fernando de Noronha (voucher: Andrade-Lima 55-2202) and seeds (or seedlings, it is not clear) from this plant were brought to, and cultivated in, Parque Estadual de Dois Irmãos, an Atlantic Forest reserve located near Recife, Pernambuco. Three years later (1958) Andrade-Lima collected a herbarium voucher from these cultivated plants (voucher: Andrade-Lima 58-3312); but since that time the plants either died or were extirpated: multiple site visits between 2015-2017 did not locate any plants of I. violacea. However, while the specimen evidence demonstrates a deliberate introduction from Fernando de Noronha to Recife, it does not account for the introduction of I. violacea to the archipelago in the first place.
Is Ipomoea violacea really absent from the West coast of the Americas?
Dedicated searching in herbaria and virtual herbaria failed to locate bona fide specimens of I. violacea from any country on the Pacific coast of the Americas. We investigated Mexico, the Central American countries, Colombia, Ecuador, Peru, and Chile. The absence of I. violacea from the Pacific coasts of Mexico and Central America stands in sharp contrast with the Caribbean coasts of Mexico and Central America, where we found numerous specimens of genuine I. violacea.
Recently, Villaseñor (2016: 705) included I. violacea in a comprehensive checklist of Mexican flora and cited for it the Pacific-coast state of Jalisco as well as several inland states (Durango, Morelos, Zacatecas) that have no coastline; he did not cite voucher specimens to document these distributions. We believe this reported distribution could represent another case of confusion between I. violacea and I. tricolor Cav. (also included by Villaseñor). We found many cases of specimens collected from Mexico that were identified as I. violacea but which were actually I. tricolor: to cite one example, G.B. Hinton collected extensively in Mexico (principally Guerrero and Michoacan) and several of his collections we have seen (e.g., 9722, 9723, 9842, 12496, 12501, 12838) were distributed as I. violacea; every one of them proved to be misidentified specimens of I. tricolor. This pattern of misidentifications in Mexican Ipomoea specimens repeated frequently and to date we found no genuine specimens of I. violacea from the Pacific coast of Mexico; we are doubtful that genuine I. violacea occurs there. Nor did we find any genuine specimens of I. violacea from any country southward along the Pacific coast of Middle and South America.
We did find specimens from Ecuador cited in TROPICOS (Tropicos 2019) and REFLORA - Virtual Herbarium (2019) as “I. violacea”; we believe these are almost certainly misidentified. As with the Mexican literature reports and specimen label data, the Ecuadorian label data mentions the corolla color is blue. This suggests strongly that these specimens are actually I. tricolor Cav. or another species. In only one case is there a digital image linked to the online specimen record: J. Hudson 744 from Esmeraldas, Ecuador, a coastal location appropriate for I. violacea. The duplicate of Hudson 744 in RB does have an image linked to the REFLORA record; this shows vegetative characters similar to I. violacea, but the sepals are shaped wrongly for that species and the label states the corolla was blue with a white center. We think this collection has been misidentified and is actually another species, possibly I. calantha Griseb. or I. clavata (G.Don) Ooststr. Unfortunately, insects have damaged the corolla so it is not possible to key out the digital image in the Flora of Ecuador key for Ipomoea species (Austin 1982c). Furthermore, Ipomoea violacea was not included in the Flora of Ecuador account (Austin 1982c) nor in the comprehensive checklist for the Ecuadorian flora (Jørgensen & León-Yánez 1999). As in the case of Mexico, we are highly skeptical that genuine I. violacea occurs on the Pacific coast of Ecuador.
This situation presents a biogeographic enigma: there is certainly suitable littoral oceanic habitat where I. violacea could thrive on the Pacific coast of the Americas, yet we have not found any trace of it there based on specimens or digital images examined. This gap in the distribution seems to be genuine and not an artifact of sampling. One oceanographic factor that could be responsible for this is the Peru (or Humboldt) Current (Fig. 2), a superficial, cold-water current running northward along the western coast of South America. The Peru Current would effectively prevent dispersal of floating seeds for I. violacea from the tropical Indo-Pacific from reaching beaches on the Pacific South American coast, because this current moves in a contrary direction.
A hypothesis for how Ipomoea violacea reached Brazil, West Africa and possibly the Caribbean
What the two cases presented here suggest to us is that I. violacea arrived relatively recently in Atlantic waters, at Fernando de Noronha and independently at Ghana, and once populations established at those localities the species spread by seed flotation in ocean currents to other nearby areas.
Both Fernando de Noronha and Ghana were stopping points for sailing ships traveling between Europe and the tropical Indo-Pacific from the early fifteenth century onward. Once Portuguese navigators reached India (in 1498) and soon thereafter Indonesia (in 1512), and China (in 1513) the stage was set for seeds of I. violacea to be transported from the Indo-Pacific to the Atlantic. The likely mechanism for the seeds to be moved is in ballast used by sailing ships. Ridley explains ballast as it relates to plant dispersal:
“Ballast is sand or gravel carried about in ships for the purpose of weighting them. … It was usually taken from near the shore whence the ship started, and was discharged at the port of destination. Hence the seeds or fruits which happened to be lying in the sand or shingle on the shore might be, and often were, transported by ocean traveling ships to great distances, and usually discharged on ground similar to that from which they came.” Ridley (1930: 645-646).
Ballast was also used in sailing ships that traversed the tropical Indo-Pacific for almost a millennium before the first Europeans arrived there. It has been well documented that a trading network spanned the vast region from eastern Africa to China and Japan (Mabberley 1998; Keay 2010): Arab traders sailed between ports on the east African coast and the Middle East to the Indian subcontinent, and onward from there to SE Asia. Chinese merchant-traders met their Arab counterparts at various trading entrepots in modern day Malaysia, Indonesia and Singapore. It is entirely possible that the vast distribution for I. violacea across the tropical Indo-Pacific has been aided by seeds transported in ballast of sailing ships from this pre-European era, but there is no means to document this now, centuries after the fact.
How Ipomoea violacea came to be a pantropical species
Our hypothesis is that I. violacea has become a globally distributed, pantropical littoral species today by the following historical routes and mechanisms:
-
Origin in the tropical Indo-Pacific;
-
Long-distance dispersal by humans (seeds carried in ballast aboard sailing ships) - possibly within Indo-Pacific; definitely between Indo-Pacific and Atlantic-Caribbean;
-
Establishment of new populations in three disjunct geographical areas:
-
the Caribbean Basin - before 1689;
-
Fernando de Noronha archipelago - before 1870;
-
Ghana / West tropical Africa - before 1930;
-
-
Local dispersal of seeds via ocean currents to other nearby locales.
In this scenario the Caribbean populations of I. violacea could have been introduced there by long-distance, human-mediated dispersal. Because the original description for I. violacea was based on plants collected in the Caribbean the presumption has long been that the species is native there. However, under the hypothesis proposed here, it is equally possible that the species could have been brought to the Caribbean from the Indo-Pacific by European sailing ships any time after Columbus “discovered” the New World in 1492. Two centuries later, I. violacea was in turn “discovered” by Charles Plumier, who botanized in the islands of the West Indies in 1689, 1693, and 1695 (Stafleu & Cowan 1983). Linnaeus based his description for I. violacea on a drawing made by Plumier in Jamaica, copied by the Dutch botanist Boerhaave, and studied by Linnaeus when he visited the Netherlands (1735-1738). Given the fairly rapid local dispersal documented here for the Brazilian and West African populations, two centuries seem ample time for I. violacea to become widely dispersed and naturalized in the Caribbean Basin. As Mabberley (1998) has pointed out for the Pacific, simply because a species was growing in a place where Europeans “discovered” it does not mean the species is native there: prior human activity had greatly altered the natural distributions for many plant species, particularly those found in coastal areas near ports and towns, centuries before botanical collecting began to provide permanent evidence for their presence. Ipomoea violacea perfectly fits this profile.
What is peculiar about the distribution of this species in the Caribbean is the long gap along the eastern Mexican coast between the Yucatan Peninsula and the single, isolated collection from Texas. This stretch of tropical ocean beach spans more than 1,800 km and there is no obvious reason why I. violacea is absent there. Yet diligent searching in herbaria and virtual herbaria failed to discover any specimens of bona fide I. violacea from the Mexican states of Tamaulipas, Veracruz, or Tabasco that would fill this gap. Nor was I. violacea included in the Flora de Veracruz account for Ipomoea (McDonald 1994). Possibly I. violacea does not occur on the eastern coast of Mexico, or perhaps this gap is due to collecting bias. Similar to the discussion for the western coast of the Americas, the absence of I. violacea from this long stretch of seemingly suitable oceanic beachfront is an enigma.
Also requiring brief comment is the possibility for I. violacea to have reached West Tropical Africa by natural means: seeds floating in oceanic currents. Although a superficially plausible scenario, this seems unlikely to us based on available specimen evidence. Certainly the prevailing oceanic currents are favorable to carrying seeds from the Indian Ocean southward via the Agulhas Current around the Cape of Good Hope and then northward along the Atlantic coast of Africa via the Benguela Current (Fig. 2). Secondly, Guppy’s flotation experiments (Guppy 1891) demonstrated that a seed of I. violacea could float in seawater for one year and also survive freezing temperatures to remain viable and germinate. However, if seeds were carried by oceanic currents around the Cape of Good Hope and along the western coast of Africa, then why are there no populations of I. violacea documented between the Cape and Ghana? There is abundant tropical, coastal littoral habitat along several thousand kilometers of Atlantic beach and one might reasonably expect plants of I. violacea to thrive there. But despite several centuries of botanical collecting along this coastline, populations of I. violacea have only recently been documented, with the earliest known collection dating from 1934. On this basis, we think it unlikely that seeds of I. violacea arrived in West Tropical Africa by natural means.
The current global distribution for I. violacea as we document it here is shown in Figure 8. The supplementary files linked to this publication (< https://doi.org/10.6084/m9.figshare.10061948.v2 >)provide the geographic coordinates based on verified voucher specimen data that support this map.

Conclusions
Several observations and insights were gained from consideration of the distribution for I. violacea on a global basis, in light of what we discovered about the regional distribution for coastal Brazil and West Tropical Africa. First, the online specimen data available from various aggregator sites (GBIF, etc.) and institutional virtual herbaria was not taxonomically trustworthy. Unless we could confirm the identity for each specimen record - either by study of the physical specimen or a high-resolution digital scan of the herbarium sheet, or the identification had been provided by a taxonomist with expertise in Convolvulaceae - it was impossible to rely on the identifications. There was simply too high a proportion of misidentified specimens and misapplied names without a preliminary taxonomic verification step. As an indication of how large this proportion of misidentifications can be Goodwin et al. (2015) stated that “…more than 50% of tropical specimens, on average, are likely to be incorrectly named.” Specimen data stored in GBIF for the genus Ipomoea was one of the global datasets these authors examined to arrive at this estimation.
The gaps we perceive in 2020 might be filled by targeted collecting. It would require dedicated collecting effort to visit beaches in the perceived gaps, to search for plants / populations of I. violacea there. In our collecting experience, field teams often bypass or spend limited time in coastal littoral zones, because the species richness is lower there compared to inland habitats and ecosystems. Plus, the flowers of I. violacea are nocturnal; botanists are seldom active in the field during the night and so they are unlikely to see the plants when in bloom. This combination of field work biases works against finding I. violacea without making a special effort to seek it out.
It should be possible, using selected molecular techniques, to test the hypothesis that I. violacea originated in the Old World and was dispersed to the New World and the Atlantic coast of Africa by human-mediated dispersal. If this can be done, then it opens the possibility to investigate the origin and dispersal for several other species of Convolvulaceae that are now pantropical, but were almost certainly assisted by human activity in achieving that distribution. Among those species are I. cairica (L.) Sweet, I. indica (Burm.) Merr., and I. pes-caprae (L.) R.Br.; for each of them, we do not know where they originated and how they spread around the world to achieve the pantropical distribution they now have.
Other Convolvulaceae with contemporary pantropical distributions - I. alba, I. asarifolia (Desr.) Roem. & Schultes, I. batatas (L.) Lam., I. nil (L.) Roth, I. purpurea (L.) Roth, I. triloba L., for example - are all believed to be natives of tropical America and were dispersed from there by human activity during the Columbian Exchange. It should be possible to apply molecular tools to these species to test this hypothesis for their origin and dispersal routes.
-
Supplementary dataSupplementary data to this paper are made available at <https://doi.org/10.6084/m9.figshare.10061948.v2>. There are 4 supplementary files that include geographical coordinates for all verified specimens of Ipomoea violacea. These specimen records provide the foundation for the global distribution described herein and were used to generate the distribution maps in Figures 4-8.
-
List of Voucher Specimens:Voucher specimens and photographic reports for Ipomoea violacea cited in the text for Brazil, French Guiana, U.S.A. and West Tropical Africa.BRAZIL. AMAPÁ: Sucurijú, lago Jaburu, 30.X.2007, fl. and fr., M.P.M. Menezes et al. 264 (HBRA, IAN); 31.X.2007, fl., M.P.M. Menezes et al. 274 (HBRA). BAHIA: Salvador, Região Metropolitana, Ilha das Fontes, São Francisco do Conte, 30.VIII.2010, fl. and fr., M.S. Lisboa & M.L. Guedes 111 (ALCB); Ilha das Frades, Fragmento Costa, 25.VII.2012, fr., M.L. Guedes et al. 19920 (ALCB). PARAÍBA: Conde, Área de Preservação Permanente de Tambaba, 23.VIII.2011, fl., L.A. Pereira & J. C. Silva 299 (JPB). PERNAMBUCO: Fernando de Noronha, Ilha da Rata, 1.VI.1993, fl., A.M. Miranda et al. 842 (ALCB, HUEFS); Ilha da Rata, St. Michael’s Mount, IX.1873, fl. and fr., Moseley (E, K, P); Ilha de São José, 18.VI.2003, fl. and fr., A.M. Miranda 4123 (HUEFS); 21.III.2000, fl. and fr., A.M. Miranda 3678 (HST, HUEFS); Ilha de Fernando de Noronha, próximo Santo Antonio, 19.X.1955, fl., Andrade-Lima 55-2202 (IPA); Chaloupe Bay, 1887, H.N. Ridley et al. 92 (P); Alagados, 19.VI.2003, fl., A.M. Miranda 4130 (RB). Goiana, R.P.P.N. Fazenda Tabatinga, 30.V.2011, fl., D. Cavalcanti 521 (UFP); R.P.P.N. Fazenda Tabatinga, ocean beach at edge of forest, 10.XI.2015, fl. and fr., G. Staples et al. 1715 (PEUFR). Recife, cultivated at Dois Irmãos, 20.XI.1958, fl., Andrade-Lima 58-3312 (IPA). Photographic voucher: ESPIRÍTO SANTO: Aracruz, interior do Refúgio da Vida Silvestre (R.V.S.) de Santa Cruz, 2017, L.M. Coser (no voucher). FRENCH GUIANA. CAYENNE COMMUNE: Îlets Dupont (îlet ouest), 3.5 m elev., 4.XI.2016, fr., R. Girault, S. Aurélien & J. Cippe 1569 (CAY). COTE D’IVORE. SASSANDRA-MARAHOUÉ: on beach near Sassandra, 21.VIII.1956, fl., J.J.F.E. de Wilde 329 (WAG). GHANA. WESTERN REGION: mouth of Ancobra River, towards Axim, IV.1952, fl., J.K. Morton 6604 (K); beach near Axima, III.1934, fr., F.R. Irvine 2565 (K); without region: Shama, edge of a mangrove swamp, 15.V.1965, fl., J.B. Hall 2982 (K). GUINEA-CONAKRY. BOKÉ PREFECTURE: Taide Island, Kamsar at SW of island, 26.XI.2013, fr., K. Guilavogui et al. 682 (HNG, K). SÉNÉGAL. ZIGUINCHOR: Parc Nationale Basse-Casamance, 2.XI.1987, fl., C. van den Berghen 8036 (WAG); Fatick, Mar Saloum, Ile de Mar, IX.1960, fl., J. Berhaut 5668 (BR). UNITED STATES OF AMERICA. TEXAS: Jefferson Co., 21 miles NE of High Island, in moist sand on beach, 19.XI.1945, fl., V.L. Cory 50993 (SMU).
Acknowledgements
The authors are grateful for the cooperation of the listed herbaria and their curators, to CNPq and also to CAPES for awarding a study grant to the first author; CNPq funded the project “Convolvuláceas da América do Sul” (PVE 314725/2014-8) from 2015-2017. Financial support: Harvard University Herbaria funded a herbarium research travel award to Staples in 2018. We are grateful to Lígia Coser for her collaboration. Sophie Gonzalez (CAY) kindly located recent specimens of I. violacea that documented the species presence in French Guyana and sent digital images for confirmation of the identification. Fernanda Satori Petrongari (SP) kindly located one of the Berhaut specimens in BR and sent digital images for confirmation of the identification. Tiana Rehman (BRIT) located and digitized the only known Texas voucher for I. violacea. The Harvard Botany Libraries obtained critical references for us. We thank Marcelo Freire Moro, Martin Cheek, and the anonymous reviewer for their feedback on the manuscript; M.F. Moro also helped us prepare the supplementary data files for uploading.
References
- Acevedo-Rodríguez P (2005) Vines and climbing plants of Puerto Rico and the Virgin Islands. Contributions from the United States National Herbarium 51: 1-483.
- Ahimsa-Müller MA, Markert A, Hellwig S, Knoop V, Steiner U, Drewke C & Leistner E (2007) Clavicipitaceous fungi associated with ergoline alkaloid-containing Convolvulaceae. Journal of Natural Products 70: 1955-1960.
- Austin DF (1975) Convolvulaceae. In: Flora of Panama. Annals of the Missouri Botanical Garden 62: 157-224.
- Austin DF (1980) Convolvulaceae. In: Dassanayake MD & Fosberg FR (eds.) A revised handbook to the Flora of Ceylon 1: 288-363.
- Austin DF (1982a) Convolvulaceae. In: Febres ZL & Steyermark JA (eds.) Flora de Venezuela 8: 15-239.
- Austin DF (1982b) Convolvulaceae. In: Correll DS & Correll HB (eds.) Flora of the Bahama Archipelago. J. Cramer, Vaduz. Pp. 1161-1190.
- Austin DF (1982c) Convolvulaceae. In: Harling G & Sparre B (eds.) Flora of Ecuador 15: 1-99.
- Austin DF (1997) Convolvulaceae. In: Boggan J, Funk V, Kelloff C, Hoff M, Cremers G & Feuillet C (eds.) Checklist of the plants of the Guianas (Guyana, Suriname, French Guiana). 2nd ed. National Museum of Natural History, Smithsonian Institution, Washington D.C. Pp. 87-88.
- Bailey LH (1901) Cyclopedia of American horticulture. Ipomoea Vol. 2. Macmillan, New York. Pp. 814-819
- Bailey LH (1935) The standard cyclopedia of horticulture. Ipomoea Vol. 2. Macmillan, New York. Pp. 1656-1662.
- Barcellos RL, Lins SRRM, Coelho-Júnior C, Travassos PEPF & Camargo PB (2016) Sistema sedimentar do Sueste: caracterização morfossedimentar e geoquímica sazonal. Mar de fora, Fernando de Noronha (PE). In: Mello TJ, Vasoncelos GJL & Micheletti T (orgs.) Anais, I encontro de pesquisa de Fernando de Noronha, São Pedro e São Paulo e Atol das Rocas. ICMBio, Fernando de Noronha. Pp. 98-101.
- Berhaut J (1967) Flore du Sénégal. Éditions Clairafrique, Dakar. 487p.
- Berhaut J (1975) Convolvulaceae. In: Flore Illustrée du Sénégal 3: 43-208.
- Bosser J & Heine H (2000) Convolvulaceae. In: Bosser J, Cadet T, Guého J & Marais W (eds.) Flore des Mascareignes 127: 1-63.
-
Brahms (2002) Brahms-management of natural history. Available at <https://dps007.plants.ox.ac.uk/bol/>. Accessed numerous times from 2002 onward.
» https://dps007.plants.ox.ac.uk/bol/ - Burkill HM (1985) The useful plants of West tropical Africa. Royal Botanic Gardens, Kew. 960p.
-
CABI (2019) Invasive Species Compendium. Wallingford, UK. Available at <https://www.cabi.org/isc>. Access on 09 January 2018.
» https://www.cabi.org/isc - Cullen J, Alexander JCM, Brickell CD, Edmondson JR, Green PS, Heywood VH, Jørgensen P-M, Jury SL, Knees SG, Maxwell HS, Miller DM, Robson NKB, Walters SM & Yeo PF (2000) The European Garden Flora. Vol. 6. Dicotyledons part IV. Cambridge Univ. Press, New York. 739p.
- Darwin C (1859) On the origin of species by means of natural selection. John Murray, London. 502p.
- Deroin T (2001) Convolvulaceae. In: Morat P (ed.) Flore de Madagascar et des Comores 117: 11-287.
-
DIVA-GIS (2014) A free computer program for mapping and geographic data analysis. Available at <https://www.diva-gis.org/>. Accessed numerous times from 2014 onward.
» https://www.diva-gis.org/ - Fang R-C & Staples G (1995) Convolvulaceae. In: Wu C-Y & Raven P (eds.) Science Press, Beijing, and Missouri Botanical Garden Press, St. Louis. Flora of China 16: 271-325.
-
Flora do Brasil 2020 em construção (2016) Instituto de Pesquisas Jardim Botânico do Rio de Janeiro. Available at <http://floradobrasil.jbrj.gov.br/>. Accessed 2016.
» http://floradobrasil.jbrj.gov.br/ - Fosberg FR & Sachet M-H (1977) Flora of Micronesia 3, Convolvulaceae. Smithsonian Contributions to Botany 36: 1-34.
- Freitas ÄMM (2007) A flora fanerogâmica atual do Arquipélago de Fernando de Noronha - Brasil. PhD dissertation, Universidade Estadual de Feira de Santana. 296p
- Gonçalves ML (1987) Convolvulaceae and Cuscutaceae. In: Launert E (ed.) Flora Zambesiaca 8: 1-142.
-
Goodwin ZA, Harris DJ, Filer D, Wood JRI & Scotland RW (2015) Widespread mistaken identity in tropical plant collections. November 16, 2015. Current Biology 25: R1057-R1069. DOI: <https://doi.org/10.1016/j.cub.2015.10.002>.
» https://doi.org/10.1016/j.cub.2015.10.002 - Griffiths M (1992) The new RHS Index of garden plants. Macmillan Press, London. 1234p.
- Gunn CR (1972) Moonflowers, Ipomoea section Calonyction, in temperate North America. Brittonia 24: 150-168.
- Gunn CR (1977) Merremia discoidesperma: its taxonomy and capacity of its seeds for ocean drifting. Economic Botany 31: 237-252.
- Gunn CR & Dennis JV (1976) World guide to tropical drift seeds and fruits. The New York Times Book Co. Demeter Press, New York. 240p.
- Guppy HB (1891 [title page 1890]) The dispersal of plants as illustrated by the flora of the Keeling or Cocos islands. Journal of the Transactions of the Victoria Institute 24: 267-306.
- Hatch SL, Gandhi KN & Brown LE (1990) Checklist of the vascular plants of Texas (MP-1655). Texas Agricultural Experiment Station, College Station. 158p.
- Heine H (1963) Convolvulaceae. In: Hepper FN (ed.) Crown agents for oversea governments and administrations, London. 2nd ed. Flora of West Tropical Africa 2: 335-352.
- Heine H (1984) Convolvulaceae. Flore de la Nouvelle-Calédonie et dépendances 13: 1-91.
-
Hoorn C, Wesselingh FP, Steege H ter, Bermudez MA, Mora A, Sevink J, Sanmartín I, Sanchez-Meseguer A, Anderson CL, Figueiredo JP, Jaramillo C, Riff D, Negri FR, Hooghiemstra H, Lundberg J, Stadler T, Särkinen T & Antonelli A (2010) Amazonia through time: Andean uplift, climate change, landscape evolution, and biodiversity. Science 330: 927-931. DOI: <https://doi.org/10.1126/science.1194585>.
» https://doi.org/10.1126/science.1194585 - Huxley A (1992) The new RHS dictionary of gardening. 4 vols. Macmillan Press, London & Stockton Press, New York.
-
IUCN ISSG (2019) Global Register of Introduced and Invasive Species (GRIIS). Available at <http://www.griis.org/>. Access on 09 January 2019.
» http://www.griis.org/ - Jørgensen PM & León-Yánez S (1999) Catalogue of the vascular plants of Ecuador. Monographs in Systematic Botany from the Missouri Botanical Garden 75: 1169.
- Keay J (2010) India, a history: from the earliest civilisations to the boom of the twenty-first century. Harper Press, London. 658p.
- Khan MS (1985) Convolvulaceae. Flora of Bangladesh 30: 1-59.
- Levy RP, Cantareli CV, Macedo EC & Mello TJ (2016) Ocorrência de lixo marinho associado ao Sargassum sp. no Arquipélago de Fernando de Noronha - PE - Brasil. In: Mello TJ, Vasoncelos GJL & Micheletti T (orgs.) Anais, I encontro de pesquisa de Fernando de Noronha, São Pedro e São Paulo e Atol das Rocas. ICMBio, Fernando de Noronha. Pp. 167-174.
- Liberty Hyde Bailey Hortorium (1976) Hortus Third. Macmillan, New York. 1290p.
- Liogier AH (1994) Convolvulaceae. La flora de la Española 6: 49-114.
- Mabberley DJ (1998) Where are the wild things? Paradisus: Hawaiian plant watercolors by Geraldine King Tam. Honolulu Academy of Arts, Honolulu. Pp. 1-11.
- MacDougall T (1960) Ipomoea tricolor a hallucinogenic plant of the Zapotecs. Boletin del Centro de Investgaciones Antropologicas de Mexico 6: 6-8.
- Manitz H (1977) “Was ist Ipomoea violacea L.?” Feddes Repertorium 88: 265-271.
- McDonald A (1994) Convolvulaceae II, Ipomoea Flora de Veracruz 77: 1-133.
- Montes C, Cardona A, Jaramillo C, Pardo A, Silva JC, Valencia V, Ayala C, Pérez-Angel LC, Rodriguez-Parra LA, Ramirez V & Niño H (2015) Middle Miocene closure of the Central American Seaway. Science 348: 226-229.
- Nelson EC (1978) Tropical drift fruits and seeds on coasts in the British Isles and western Europe, 1. Irish beaches. Watsonia 12: 103-112.
- O’Dea A, Lessios HA, Coates AG, Eytan RI, Restrepo-Moreno SA, Cione AL, Collins LS, Queiroz A, Farris DW, Norris RD, Stallard RF, Woodburne MO, Aguilera O, Aubry M-P, Berggren WA, Budd AF, Cozzuol MA, Coppard SE, Duque-Caro† H, Finnegan S, Gasparini GM, Grossman EL, Johnson KG, Keigwin LD, Knowlton N, Leigh EG, Leonard-Pingel JS, Marko PB, Pyenson ND, Rachello-Dolmen PG, Soibelzon E, Soibelzon L, Todd JA, Vermeij GJ & Jackson JBC (2016) Formation of the isthmus of Panama. Science Advances 2016;2: e1600883.
- Ooststroom SJ van (1940) The Convolvulaceae of Malaysia, III. The genus Ipomoea Blumea 3: 481-582.
- Ooststroom SJ van & Hoogland RD (1953) Convolvulaceae. In: Van Steenis CGGJ (ed.) Noordhoff-Kolff N.V., Jakarta. Flora Malesiana 4: 388-512.
- Philip C & Lord T (1988) The Plant Finder. 2nd ed. Headmain Ltd. for the Hardy Plant Society, Whitbourne. 480p.
- Philip C & Lord T (1990) The Plant Finder. 4th ed. Headmain Ltd. for the Hardy Plant Society, Whitbourne. 635p.
- Philip C & Lord T (1994) The RHS Plant Finder. 8th ed. Headmain Ltd. for the Royal Horticultural Society, Whitbourne. 832p.
- Powell DA (1979) Convolvulaceae of the Lesser Antilles. Journal of the Arnold Arboretum 60: 219-271.
- Powell DA, Nicolson DH & Austin DF (1978) Convolvulus grandiflorus Jacq. (Convolvulaceae) re-examined. Brittonia 30: 199-202.
- Randall RP (2017) A global compendium of weeds. 3rd ed. Published by the author, Mount Helena. 3654p.
-
REFLORA - Virtual Herbarium (2019) Available at <http://reflora.jbrj.gov.br/reflora/herbarioVirtual/ConsultaPublicoHVUC/ConsultaPublicoHVUC.do>. Accessed 16 October 2019.
» http://reflora.jbrj.gov.br/reflora/herbarioVirtual/ConsultaPublicoHVUC/ConsultaPublicoHVUC.do - Ridley HN (1890 [title page 1891]) Notes on the botany of Fernando Noronha. Journal of the Linnean Society, Botany 27: 1-95.
- Ridley HN (1930) The dispersal of plants throughout the world. L. Reeve & Co., Ashford. 744p.
- Sachet M-H (1975) Flora of the Marquesas, 1: Ericaceae-Convolvulaceae. Smithsonian Contributions to Botany 23: 1-34.
- Schultes RE (1964) The nomenclature of two Mexican narcotics. Taxon 13: 65-66.
- Silveira ICA, Schmidt ACK, Campos EJD, Godoi SS & Ikeda Y (2000) A corrente do Brasil ao largo da costa leste brasileira. Revista Brasileira de Oceanografia 48: 171-183.
-
SpeciesLink (2002 and ongoing) Available at <http://www.splink.org.br/index>. Accessed numerous times from 2017-2020.
» http://www.splink.org.br/index - Stafleu FA & Cowan RS (1983) Taxonomic Literature (second edition). Vol. IV: P-Sak. Regnum Vegetabile 110. Bohn, Scheltema & Holkema, Amsterdam. 1214p.
- Staples G (2010) Convolvulaceae. In: Smitinand T & Larsen K (eds.) Forest Herbarium, Royal Forest Department, Bangkok. Flora of Thailand 10: 330-468.
- Staples G W & Syahida-Emiza S (2015) Convolvulaceae. In: Kiew R, Chung RCK, Saw LG & Soepadmo E (eds.) Forest Research Institute Malaysia, Kepong. Flora of Peninsular Malaysia: Seed Plants 5: 55-198.
- Synge PM (1956) The Royal Horticultural Society Dictionary of Gardening. Ipomoea 2nd ed. Vol. 2. Clarendon Press, Oxford. Pp 1059-1061.
-
Tropicos (2019) Missouri Botanical Garden. Available at <http://www.tropicos.org>. Access on 03 June 2019.
» http://www.tropicos.org - Verdcourt B (1958) Notes from the East African Herbarium: VII. Notes on African Convolvulaceae (Part 3). Kew Bulletin 1958: 199-217.
- Verdcourt B (1970) Convolvulaceae. In: Menninger EA Flowering vines of the world; an encyclopedia of climbing plants. Hearthside Press, New York. Pp. 142-158.
- Villaseñor JL (2016) Checklist of the native vascular plants of Mexico. Revista Mexicana de Biodiversidade 87: 559-902.
Edited by
-
Area Editor: Dr. Marcelo Moro








