Resumos
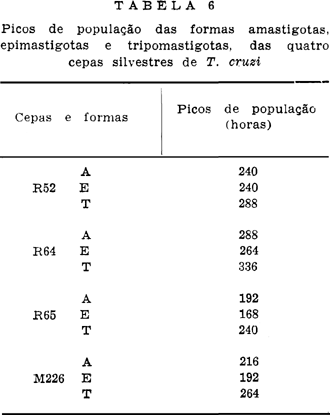
Foram estudadas três cepas de T. cruzi isoladas de Didelphis albiventris (R52, R64 e R65) e uma isolada de Calomys callosus (M226), quanto ao comportamento "In vitro" no meio LIT. A evolução da população dos tripanossomas com relação ao crescimento e morfogênese foi acompanhada por um período de 13 dias (312 horas), em intervalos regulares. As contagens diferenciais, separando-se formas amastigotas, epimastigotas e tripomastigotas, foram realizadas na câmara de Neubauer. Os resultados obtidos levaram à conclusão de que as 4 cepas estudadas têm comportamento distintos. O melhor crescimento obtido ocorreu para a cepa M226, seguindo-se por ordem decrescente a R65, R52 e R64. Os picos das populações ocorreram como segue: M226, entre 192-264 horas; R65 entre 168-240 horas; R52 às 240 horas e R64 entre 264-312 horas.
Trypanosoma cruzi; Didelphis albiventris; Calomys callosus
In the course of this work, four strains of sylvatic Trypanosoma cruzi were studied in regard to their growth and differentiation in LIT medium. R52, R64, and R65 were isolated from individual Didelphis albiventris (Marsupialia) and M226 from Calomys callosus (Rodentia). The evolution of the numerical incresase and morphogenesis of the trypanosome populations in the culture was observed at regular intervals during a period of 312 hours. Differential counts were made of amastigotes, epimastigotes, and trypomastigotes in Neubauer chambers. Results showed that the four strains had characteristic behavior patterns. M226 increased the most and was followed by R65, R52, and then R64.
Trypanosoma cruzi; Didelphis albiventris; Calomys callosus
Crescimento e diferenciação "In vitro" de cepas de Trypanosoma cruzi, isoladas de animais silvestres* * Trabalho realizado com auxílio do Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq PDE/02-1-06, FUB-CNPq, cadastro n o 81.961)
Growth and differentiation "In vitro" of strains of Trypanosoma cruzi isolated from wild animals
Dalva A. MelloI; Monamaris M. BorgesII; Lúcia Helena ChiariniIII
IDo Departamento de Medicina Complementar da Faculdade de Ciências da Saúde da Universidade de Brasília 70910 Brasília, DF Brasil
IIBolsista do CNPq
IIIDo Departamento de Estatística do Instituto de Ciências Exatas da Universidade de Brasília 70910 Brasília, DF Brasil
RESUMO
Foram estudadas três cepas de T. cruzi isoladas de Didelphis albiventris (R52, R64 e R65) e uma isolada de Calomys callosus (M226), quanto ao comportamento "In vitro" no meio LIT. A evolução da população dos tripanossomas com relação ao crescimento e morfogênese foi acompanhada por um período de 13 dias (312 horas), em intervalos regulares. As contagens diferenciais, separando-se formas amastigotas, epimastigotas e tripomastigotas, foram realizadas na câmara de Neubauer. Os resultados obtidos levaram à conclusão de que as 4 cepas estudadas têm comportamento distintos. O melhor crescimento obtido ocorreu para a cepa M226, seguindo-se por ordem decrescente a R65, R52 e R64. Os picos das populações ocorreram como segue: M226, entre 192-264 horas; R65 entre 168-240 horas; R52 às 240 horas e R64 entre 264-312 horas.
Unitermos:Trypanosoma cruzi, cepas silvestres. Didelphis albiventris. Calomys callosus.
ABSTRACT
In the course of this work, four strains of sylvatic Trypanosoma cruzi were studied in regard to their growth and differentiation in LIT medium. R52, R64, and R65 were isolated from individual Didelphis albiventris (Marsupialia) and M226 from Calomys callosus (Rodentia). The evolution of the numerical incresase and morphogenesis of the trypanosome populations in the culture was observed at regular intervals during a period of 312 hours. Differential counts were made of amastigotes, epimastigotes, and trypomastigotes in Neubauer chambers. Results showed that the four strains had characteristic behavior patterns. M226 increased the most and was followed by R65, R52, and then R64.
Uniterms:Trypanosoma cruzi, wild strains. Didelphis albiventris. Calomys callosus.
INTRODUÇÃO
Chagas8 (1909), foi o primeiro autor a verificar morfogênese do Tripanosoma cruzi em meio de cultura acelular. Posteriormente, numerosos trabalhos foram realizados sobre cultivo de T. cruzi "In vitro" em diferentes meios acelulares, sendo analisados vários aspectos do comportamento deste protozoário. Entre outros trabalhos sobre estes assuntos, chama atenção os de: Muniz e Freitas11 (1946), Noble 12 (1955), Pick15 (1959), Camargo6 (1964), Brener e Chiari4 (1965), Castellani e col.7 (1967), Pan14 (1971), Chiari9 (1974).
No que se refere particularmente a estudos com cepas de T. cruzi isoladas de animais silvestres salientam-se os trabalhos de Norman e col.13 (1959) e de Ribeiro e Barretto16 (1975). O último autor tentou relacionar o tipo de crescimento de cepas de T. cruzi, cultivadas no meio Warren, com o comportamento em animais e polimorfismo das formas sanguícolas baseando-se nas observações realizadas por Brener2,3 (1965 e 1977), Brener e Chiari5 (1963) e Andrade1 (1976).
Com o objetivo de contribuir acrescentando novos dados experimentais ao assunto, este trabalho apresenta resultados obtidos sobre crescimento e diferenciação "In vitro" de cepas de T. cruzi, isoladas de animais silvestres.
MATERIAL E MÉTODOS
As cepas utilizadas neste trabalho tiveram as seguintes origens: R52 foi isolada de uma fêmea de Didelphis albiventris (Marsupialia) capturada no norte do município de Formosa, Goiás, em 17 de setembro de 1975; R64 foi isolada também de uma fêmea de D. albiventris capturada no município de Mambaí, Goiás, em 20 de setembro de 1975; R65 teve a mesma origem, data de captura e procedência da R52; M226 foi isolada de uma fêmea de Calomys callosus (Rodentia) coletada também no norte do município de Formosa em 10 de dezembro de 1975. Estas cepas eram mantidas em camundongos albinos e em C. callosus nascidos em laboratório, através de inoculações com fezes positivas de barbeiros alimentados nestes animais.
Para os isolamentos em meio de cultura acelular e para os estudos sobre o crescimento e diferenciação dos tripanossomas, foi utilizado o meio LIT. O volume do meio de cultura em cada tubo foi de 8 ml. A proporção de antibiótico por tubo com LIT foi de 0,8 mg de estreptomicina e 800 unidades de penicilina. As culturas eram mantidas na estufa a 28°C.
O isolamento inicial das cepas no LIT foi feito a partir de animais que apresentavam níveis elevados de parasitemia. Estes eram anestesiados com éter e por punção cardíaca em câmara assética retirava-se sangue destinado ao meio de cultura.
Após 8-20 dias seguidos ao semeio do sangue infectado, foi realizado o primeiro repique. Deste primeiro repique, procedia-se novos semeios semanais até se obter um crescimento rico, sendo então iniciados os estudos quantitativos. O inóculo para cada tubo variou entre 5.093-5.950 parasitos/ml; a idade das cepas após o primeiro isolamento foi: R52 = 67 dias, R64 = 63 dias, R65 = 50 dias e M226 = 37 dias. Foram utilizados para cada cepa cinco tubos com cultura. Antes de se proceder às contagens os tubos eram bem homogeneizados e o resultado final correspondia a média das cinco contagens para cada cepa.
A evolução da população dos tripanosomas, quanto ao seu crescimento numérico e morfogênese, foi acompanhada através da contagem destes em câmara de Neubauer, como está descrito em Camargo6 (1964) com aumento de 500 (12,5x40). Estas contagens eram feitas separando-se as formas amastigotas, epimastigotas e tripomastigotas. A evolução das quatro cepas de T. cruzi foi acompanhada durante 13 dias (Tabela 1). Das culturas em diferentes períodos foram feitas lâminas coradas com Giemsa-MayGrunwald e destas lâminas foram tiradas microfotografias com objetiva x 100.
A análise estatística dos dados prendeu-se ao estudo do comportamento das curvas de crescimento da população dos tripanosomas nas quatro cepas e formas. Para cada um dos fenômenos analisados, foi verificada a função que melhor se ajustava às observações e a taxa média de crescimento.
As taxas médias de crescimento foram calculadas a partir de uma média aritmética, média quadrática ou média geométrica dependendo da função ajustante ser uma reta, parábola do 2o grau, exponencial ou logística. Tais funções foram estimadas através do critério dos mínimos quadrados exceto a logística que foi ajustada a partir do método dos pontos selecionados.
RESULTADOS
Análise qualitativa
As quatro cepas de T. cruzi estudadas neste trabalho sempre apresentaram nos repiques iniciais, abundantes aglomerados de formas amastigotas (Fig. 1). Quando a população das cepas atingiam o ponto de saturação no tubo de cultura, praticamente estes aglomerados desapareciam, cedendo lugar aos epimastigotas e tripomastigotas (Fig. 2), embora raros amastigotas livres ainda pudessem ser vistos. Em várias ocasiões foi possível constatar tripomastigotas como se estivessem em processo de divisão (Fig. 3). Tais aspectos foram observados somente a partir da elevação da população destas formas. A manutenção destas cepas "In vitro" no LIT, por um período de 12-13 meses após o isolamento inicial do T. cruzi do sangue, e sucessivos repiques (perfazendo um total de 29), mostrou que havia um decréscimo gradativo e substancial das formas amastigotas.
Quanto à sobrevivência no LIT não renovado, os resultados encontrados foram diferentes para cada cepa: R52 morria com 41 dias de manutenção neste meio; R64 morria com 31 dias; R65 com 60 dias; e finalmente M226 com 70 dias.
Análise Quantitativa
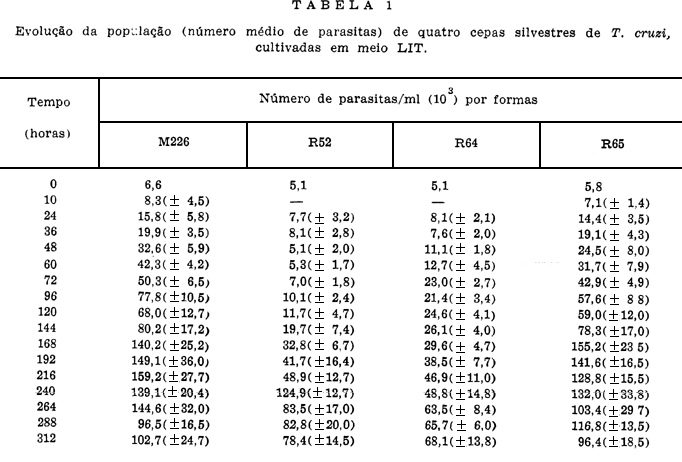
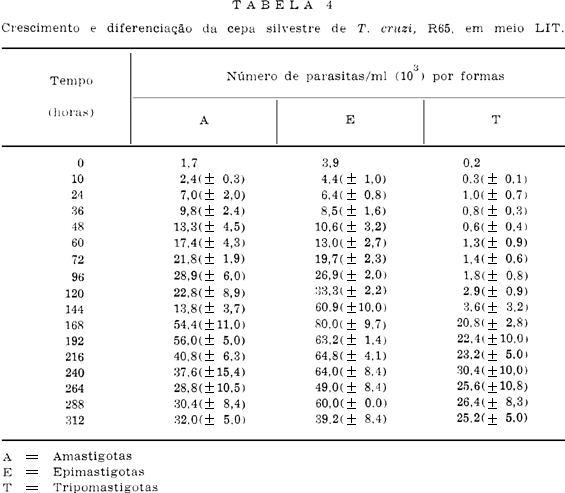
A Tabela 1 apresenta os resultados obtidos sobre os níveis de crescimento no meio LIT das quatro cepas de T. cruzi estudadas neste trabalho.
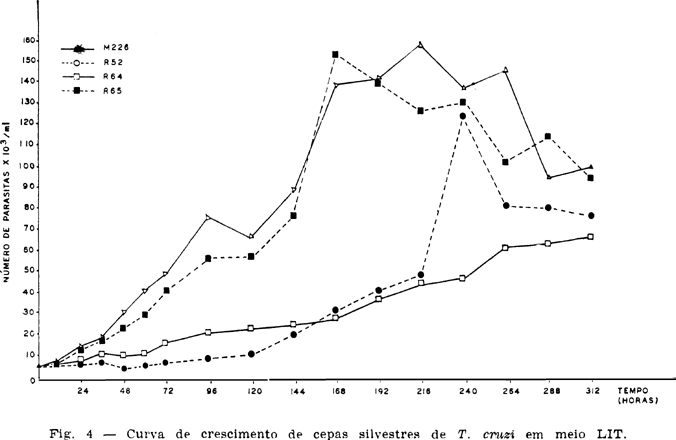
A cepa que apresentou melhor crescimento numérico total foi a M226, vindo por ordem decrescente a R65, R52 e R64 (Tabela 1 e Fig. 4).
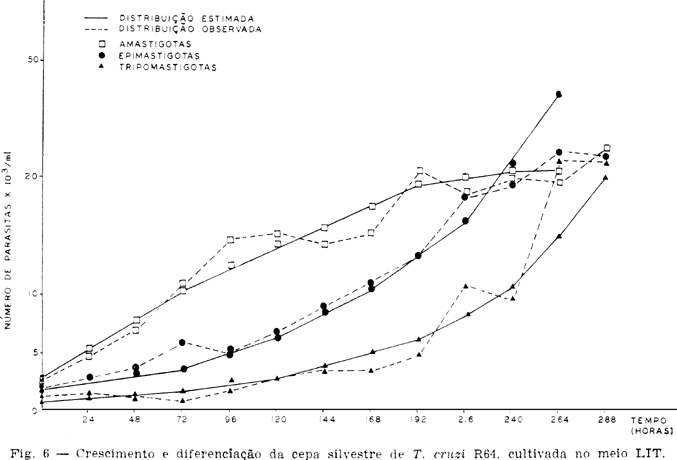
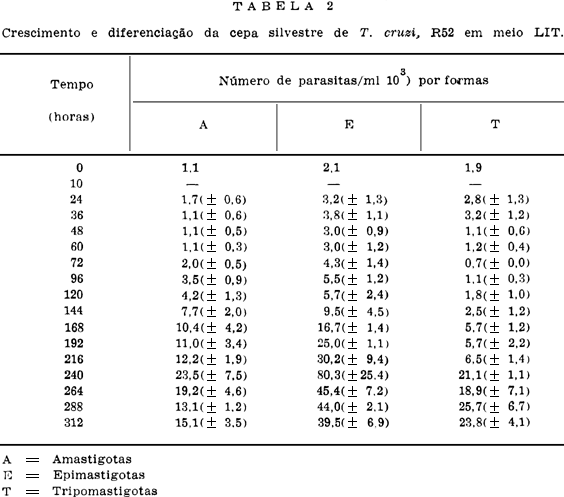
As Tabelas e Figs. 5, 6, 7 e 8 mostram os resultados sobre o crescimento numérico e diferenciação de cada cepa estudada. A produção de amastigotas para a cepa R52 (Tabela 2), durante o período de observação em todo o tempo foi inferior a de epimastigotas e superior as de tripomastigotas até as 264 h. Para a R64 (Tabela 3), o crescimento de amastigotas foi superior às epimastigotas e tripomastigotas até as 192 h se equilibrando numericamente ao final das observações (288 h, 312 h e 336 h). Com relação às cepas R65 e M226 (Tabelas 4 e 5), verificou-se que aquelas mesmas formas tiveram um crescimento numérico maior do que as epimastigotas até as 96 h e 72 h respectivamente, sendo entretanto sempre superior as formas tripomastigotas.
Os picos de população das formas e cepas estudadas em relação ao tempo, encontra-se na Tabela 6.
As curvas de crescimento para cada forma e cepa estão representadas nas Figs. 5, 6, 7 e 8. As taxas de crescimento e tipo de curvas obtidas estão apresentadas na Tabela 7.
Enquanto as cepas R52 e R65 apresentaram um crescimento exponencial para as três formas, a cepa R64 só o apresentou para as epimastigotas e tripomastigotas. A forma amastigota desta cepa apresentou um crescimento representado por uma parábola do segundo grau. Para cepa M226 as formas amastigotas, epimastigotas e tripomastigotas mostraram, respectivamente, um crescimento representado por uma reta, uma logística e uma reta.
A maior taxa média de crescimento das formas amastigotas deu-se na cepa M226, seguida pela R65, R52 e R64. Para as epimastigotas a cepa R65 foi a que apresentou a maior taxa média de crescimento, seguindo-se as cepas R52, M226 e R64. Finalmente, as tripomastigotas, que apresentaram a maior taxa média de crescimento foram as da cepa M226, seguindo-se as R65, R64 e R52.
COMENTÁRIOS E CONCLUSÕES
Norman e col.13 (1959) estudando sete cepas de T. cruzi isoladas de animais silvestres em meio bifásico, verificaram que todas elas apresentavam formas amastigotas que apareciam entre o 4° e 10° dia após incubação. O número destas formas dependia, segundo estes autores, da quantidade do inóculo. A partir do 14° dia as culturas apresentavam as formas epimastigotas. Os tripomastigotas só eram observados em baixo número, após várias transferências e em culturas com 28 dias de idade. Das sete cepas estudadas, apenas uma, isolada de Procyonlotor, apresentou mudanças de comportamento durante os experimentos.
Brener e Chiari 2 (1965), ao estudar o comportamento "In vitro" no meio LIT, de quatro cepas isoladas de casos humanos e três isoladas de Triatoma infestans, verificaram diferentes comportamentos. As cepas isoladas de triatomíneo cujas formas sanguícolas eram bem largas deram origem nos repiques iniciais a massas coloniais de amastigotas puras ou com epimastigotas. Estas cepas, após repiques a cada 10 dias por um período de dois meses, apresentavam gradual desaparecimento das amastigotas, sendo substituídas por epimastigotas. Nas cepas de origem humana, cujas formas sanguícolas eram predominantemente delgadas (Y e Berenice), as amastigotas rapidamente desapareciam e após dez dias de incubação só havia epimastigotas livres ou em rosetas. As outras duas cepas, de origem humana (ABC e PNM), com formas sanguícolas largas, embora apresentassem "In vitro" epimastigotas em dominância, no entanto raras massas de amastigotas poderiam ainda ser encontradas. Concluíram esses autores que a presença ou ausência das amastigotas estaria relacionada diretamente com características intrínsecas das cepas.
Ribeiro e Barretto 16 (1975) estudaram comportamento "In vitro" no meio Warren, de 10 cepas de T. cruzi de diferentes origens comparando-o com infecções experimentais em camundongos. Evidenciaram estes autores comportamentos diferentes, que levou-os a inserir estas cepas em três grupos: I (uma amostra isolada de D. azarae e uma de Triatoma infestans) as formas sanguícolas dominantes nos camundongos infectados eram largas; na cultura as colônias de amastigotas persistiam após quatro subculturas e praticamente desapareciam dando lugar às formas flageladas; II (duas amostras de Tadarida laticaudata e duas humanas, Indio e Y) nestas raramente encontravam-se amastigotas; III (três amostras de roedores Akodon sp. e Sciurus sp. e uma de Cebus apella) morfologia sanguícola heterogênea; as culturas recentes apresentavam amastigotas grupadas ou isoladas, as quais desapareciam paulatinamente para dar lugar só às formas epimastigotas e tripomastigotas.
Os resultados de Ribeiro e Barretto16 (1975) apresentaram concordância em parte com os de Brener e Chiari4 (1965) no que se refere aos aspectos da morfologia das formas sanguícolas e o comportamento "In vitro", discordando entretanto no que se refere a patogenicidade.
Ao se analisar do ponto de vista qualitativo, os dados obtidos neste trabalho, verifica-se que o comportamento das amastigotas no LIT foi homogêneo para as quatro cepas estudadas. Todas apresentaram aglomerados numerosos de formas amastigotas nas culturas jovens, os quais diminuiam progressivamente nos repiques sucessivos para ceder lugar às epimastigotas e tripomastigotas (Figs, 1 e 2). Chama atenção o encontro das tripomastigotas aos pares como se estivessem em divisão (Fig. 3). Achado similar está relatado no trabalho de Camargo6 (1964). As quatro cepas tinham formas sanguícolas predominantemente largas sendo pouco patogênicas para camundongos e Calomys callosas (Mello e col.10 1979). Pode-se concluir portanto, referindo-se ao comportamento das amastigotas, ao tipo das formas sanguícolas e à patogenicidade, que os resultados aqui obtidos não seguem os mesmos padrões daqueles encontrados por Brener e Chiari4 (1965) e Ribeiro e Barretto16 (1975).
Camargo6 (1964), estudando o comportamento de cepa Y no meio LIT, verificou que havia uma fase de crescimento que correspondia a um período de quatro dias. O número de metacíclicos se desenvolvia no final da fase exponencial e atingia o maior número na fase estacionaria. Quando este autor mantinha as culturas com índice logarítimo permanente obtido em subculturas, o pico metacíclico jamais ocorria. A fase logarítimica caracterizava-se por uma predominância de epimastigotas.
Castellani e col.7 (1967) mostraram que a diferenciação de epimastigotas em tripomastigotas seria estimulada por acumulação de ácidos orgânicos no meio, no final do crescimento exponencial. Esses resultados foram confirmados por Pan14 (1971), ao estudar o comportamento de cepa Brasil em dois meios diferentes com três variações de temperatura (24,5°C, 29,5°C e 35°C). Concluiu esse autor que a composição do meio é que seria importante na produção numérica e diferenciação do parasito.
Chiari9 (1974) analisou crescimento e diferenciação nos meios LIT e HIL, das cepas humanas de T. cruzi, Y e MR, mantidas "In vitro" por diferentes períodos de tempo após isolamento do hospedeiro. Estas cepas haviam sido estudadas anteriormente por Brener e Chiari5 (1963), Camargo6 (1964) e Brener e Chiari4 (1965). No trabalho de Chiari6 (1974) os resultados encontrados não mostraram diferenças significantes quanto ao crescimento das duas cepas guardadas por longo tempo. As modificações observadas referiam-se a metaciclogênese.
Do ponto de vista quantitativo, os resultados aqui obtidos mostraram diferenças de crescimento estatisticamente significantes para cada cepa e forma (Tabelas 2, 3, 4, 5 e 7). Comparar estes resultados com aqueles de Camargo6 (1964) e Chiari4 (1974) em relação aos aspectos quantitativos, seria discrepante urna vez que as cepas aqui estudadas foram isoladas recentemente do hospedeiro vertebrado (37-67 dias). Comentários entretanto poderiam ser conduzidos no que se refere ao tipo de crescimento obtido com estas cepas de T. cruzi. Como mostra as Figs. 5, 6, 7 e 8 e a Tabela 7, as curvas de crescimento nem sempre foram as mesmas para todas as formas e cepas. Embora as cepas R52 e R65 tenham tido diferenças de crescimento distintas, evoluiram entretanto dentro de um mesmo padrão, i.e., as curvas de crescimento obtidas foram do tipo exponencial (Tabela 6 e Figs. 5 e 7). As formas amastigotas da cepa R64 evoluiram segundo uma parábola do segundo grau, enquanto as epimastigotas e tripomastigotas segundo uma exponencial (Tabela 6, Fig. 6). As formas amastigotas e tripomastigotas da cepa M226 deram uma reta enquanto as epimastigotas deram uma logística.
Os resultados obtidos na pesquisa aqui realizada levaram a conclusão de que as quatro cepas de T. cruzi estudadas têm comportamento distintos. Estes resultados devem no entanto ser encarados considerando que: estas cepas foram isoladas de hospedeiros vertebrados silvestres; foram estudadas logo depois de isolamento "In vitro"; têm pouca patogenicidade para camundongos (Mello e col.10 1979), e as formas sanguícolas são predominantemente largas.
Recebido para publicação em 02/09/1980
Aprovado para publicação em 22/09/1980
- 1. ANDRADE, S. Tentative for growping different Trypanosoma cruzi strains in some types. Rev. Inst. Med. trop. S. Paulo, 18:140-1, 1976.
- 2. BRENER, Z. Comparative studies of different strains of Trypanosoma cruzi. Ann. trop. Med. Parasit., 59:19-26, 1965.
- 3. BRENER, Z. Intraspecific variations in Trypanosoma cruzi: two types of parasite populations presenting distinct characteristics. Washington, D. C., Pan American Health Organization, 1977. p. 11. (PAHO-Scient. Publ., 347).
- 4. BRENER, Z. & CHIARI, E. Aspects of early growth of different Trypanosoma cruzi strains in culture medium. J. Parasit., 51:922-6, 1965.
- 5. BRENER, Z. & CHIARI, E. Variações morfológicas observadas em diferentes amostras de Trypanosoma cruzi. Rev. Inst. Med. trop. S. Paulo, 5:220-4, 1963.
- 6. CAMARGO, E. P. Growth and differentiation in Trypanosoma cruzi. I. Origin of metacyclic trypanosoma in liquid media. Rev. Inst. Med. trop. S. Paulo, 6:93-100, 1964.
- 7. CASTELLANI, O. et al. Differentiation of Trpanosoma cruzi in culture. J. Protozool., 14:447-51, 1967.
- 8. CHAGAS, C. Nova Tripanossomíase humana. Estudos sobre a morfologia e ciclo evolutivo do Schisotrypanum cruzi n. gen, n. sp.; Agente etiológico de nova entidade mórbida do homem. Mem. Inst. Oswaldo Cruz, 1:159-217, 1909.
- 9 CHIARI, E. Growth and differentiation of Trypanosoma cruzi culture forms kept in laboratory for different periods of time. Rev. Inst. Med. trop. S. Paulo, 16:81-7, 1974.
- 10. MELLO, D. A. et al. Alguns aspectos do comportamento de cepas silvestres de Trypanosoma cruzi em camundongos e Calomys callosus (Rodentia). Rev. Saúde públ., S. Paulo, 13:314-25, 1979.
- 11. MUNIZ, J. & FREITAS, G. Realização "In vitro" do ciclo do S. cruzi no vertebrado, em meios de caldo líquido peritoneal. Rev. bras Biol., 6:467-84, 1946.
- 12. NOBLE, E. R. The morphology and life cycles of trypanosomes. Quart. Rev. Biol., 30:1-28, 1955.
- 13. NORMAN, L. et al. Morphology and virulence of Trypanosoma cruzi like hemoflagellates isolated from wild mammals in Georgia and Florida. J. Parasit., 45:457-63, 1959.
- 14. PAN, C. T. Cultivation and morphogenesis of Trypanosoma cruzi in improved liquid media. J. Protozool., 18:556-60, 1971.
- 15. PICK, F. Sur un nouveau milieu au sang permettant la transmission rapide de T. cruzi (formes sanguines et metacyclique) en forme leismanienes. Bull. Soc. Path. exot., 50:217-19, 1959.
- 16. RIBEIRO, R. D. & BARRETO, M. P. Estudos sobre reservatórios e vetores silvestres do Trypanosoma cruzi LXI. Aspectos do comportamento de amostras diversas de T. cruzi em cultura. Rev. bras. Biol., 35:301-7, 1975.
Datas de Publicação
-
Publicação nesta coleção
27 Jan 2006 -
Data do Fascículo
Dez 1980
Histórico
-
Aceito
22 Set 1980 -
Recebido
02 Set 1980