Abstract
The male holotype of Hapalopus nondescriptus Mello-Leitão, 1926 is redescribed, illustrated and compared with freshly collected specimens from the type locality. The only difference noted among the holotype and the new material concerns the development of the subapical keel. Its taxonomic position is reinterpreted and discussed, resulting in its transfer to the genus Vitalius Lucas, Silva Junior & Bertani, 1993, and thus making the new combination Vitalius nondescriptus (Mello-Leitão, 1926) comb. nov. The female is described for the first time and the morphological variations in two males, born from the female used in the description, is presented and illustrated. The male differs from those of other Vitalius species by the palpal bulb with short apical keel and bifid tibial spur with narrow prolateral branch and almost straight retrolateral branch. The female differs from those of other Vitalius species by urticating hair of 'type I' having the region 'a' shorter than region 'b'. Hapalopus nondescriptus has a confusing taxonomic history, since the holotype specimen was also used to describe another theraphosid species (Cyclosternum melloleitaoi Bücherl, Thimoteo & Lucas, 1971) which was, consequently, considered its objective synonym. Thus, we consider it a clear example of theraphosid taxonomical chaos.
Brazil; Cyclosternum; Hapalopus; objeticve synonym; taxonomy
TAXONOMY AND NOMENCLATURE
Vitalius nondescriptus comb. nov. (Araneae: Theraphosidae: Theraphosinae): an example of theraphosid taxonomic chaos
Rogério Bertani** Corresponding author. E-mail: rbertbutantan.gov.br; Roberto Hiroaki Nagahama; Caroline Sayuri Fukushima
Laboratório Especial de Ecologia e Evolução, Instituto Butantan. Avenida Vital Brazil 1500 05503-900 São Paulo, SP, Brazil
ABSTRACT
The male holotype of Hapalopus nondescriptus Mello-Leitão, 1926 is redescribed, illustrated and compared with freshly collected specimens from the type locality. The only difference noted among the holotype and the new material concerns the development of the subapical keel. Its taxonomic position is reinterpreted and discussed, resulting in its transfer to the genus Vitalius Lucas, Silva Junior & Bertani, 1993, and thus making the new combination Vitalius nondescriptus (Mello-Leitão, 1926) comb. nov. The female is described for the first time and the morphological variations in two males, born from the female used in the description, is presented and illustrated. The male differs from those of other Vitalius species by the palpal bulb with short apical keel and bifid tibial spur with narrow prolateral branch and almost straight retrolateral branch. The female differs from those of other Vitalius species by urticating hair of 'type I' having the region 'a' shorter than region 'b'. Hapalopus nondescriptus has a confusing taxonomic history, since the holotype specimen was also used to describe another theraphosid species (Cyclosternum melloleitaoi Bücherl, Thimoteo & Lucas, 1971) which was, consequently, considered its objective synonym. Thus, we consider it a clear example of theraphosid taxonomical chaos.
Key words: Brazil; Cyclosternum; Hapalopus; objeticve synonym; taxonomy.
The mygalomorph Theraphosidae comprises most of the largest living spiders, commonly called "tarantulas". With almost one thousand species described (PLATNICK 2012), it is one of the richest spider families. The taxonomic difficulties of the family are also well known. RAVEN (1990) called attention to the lack of taxonomic revisions in Theraphosidae, "either in its entirety or in any region"; and said that "a group so long left without revision becomes a nomenclatural and taxonomic nightmare". Despite the progress in the taxonomy of Theraphosidae in the last twenty years, much is yet to be done.
In this work we elucidate the identity of the species Hapalopus nondescriptus Mello-Leitão, 1926, which has a confusing history. It was described based on a single male from Ouro Preto, state of Minas Gerais, Brazil. The author considered it close to Hapalopus flovohirtus Simon, 1889 (now Catanduba flavohirta). Many years later, BÜCHERL et al. (1971) reviewed various types established by Mello-Leitão housed at the Museu Nacional, Rio de Janeiro. They found a specimen labeled "Hapalopus sp.", from Ouro Preto, but did not recognize it as a Mello-Leitão type. They considered it an undescribed species of Cyclosternum Ausserer, 1871 and decided to describe this new species, naming it Cyclosternum melloleitaoi Bücherl, Timotheo & Lucas, 1971.
Recently, a catalog of spider types described by Mello-Leitão was published (SILVA-MOREIRA et al. 2010) and the authors stated that there is no doubt that both species were described using the same specimen as a holotype, making Cyclosternum melloleitaoi an objective synonym of Hapalopus nondescriptus. However, PLATNICK (2012) considered that SILVA-MOREIRA et al. (2010) "provided no evidence justifying a placement of the species in either genus" and that "their conclusion should be checked by a theraphosid systematist before any change is accepted". Both species are thus listed as valid in Platnick's Catalog (PLATNICK 2012).
The case involving H. nondescriptus and C. melloleitaoi is a clear example of the theraphosid "taxonomic chaos" to which RAVEN (1990) referred. All efforts to clarify the identity of one or more theraphosid species are important to reverse this taxonomic chaos. We redescribe the species, describe the female for the first time, and reinterpret its taxonomic position after studying the holotype and additional specimens.
MATERIAL AND METHODS
The general description format follows BERTANI (2001) and RAVEN (2005) with some modifications. Terminology of male palpal bulb follows BERTANI (2000), of urticating hairs follows COOKE et al. (1972) and of leg spination follows PETRUNKEVITCH (1925) with modifications proposed by BERTANI (2001).
All measurements are in millimeters and were obtained with a Mitutoyo® digital caliper with an error of 0.005 mm, rounded up to two significant decimals. Leg and palp measurements were taken from dorsal aspect of right side (unless appendages were lost or obviously regenerated). A Nikon® SMZ1500 dissecting microscope was used for illustrations (with camera lucida attachment). A Bioval® 2000 microscope and a Canon® T3i digital camera were used to obtain urticating hair images. Abbreviation: (A) apical keel, (ALE) anterior lateral eyes, (AME) anterior median eyes, (ap) apical, (ITC) inferior tarsal claw, (PI) prolateral inferior keel, (PLE) posterior lateral eyes, (PME) posterior median eyes, (PMS) posterior median spinnerets, (p) prolateral, (PS) prolateral superior keel, (r) retrolateral, (R) retrolateral keel, (SA) subapical keel, (STC) superior tarsal claws, (v) ventral.
Specimens of the following institutions were examined: MNRJ - Museu Nacional, Rio de Janeiro; MZSP - Museu de Zoologia da Universidade de São Paulo, São Paulo.
Geographical coordinates: primary sources are between round brackets, secondary sources (Google Earth©) between square brackets.
TAXONOMY
Vitalius nondescriptus (Mello-Leitão, 1926) comb. nov. Figs 1-11
Hapalopus nondescriptus Mello-Leitão, 1926: 319, figs 7-8; Silva-Moreira et al., 2010: 69; Platnick, 2012.
Cyclosternum melloleitaoi Bücherl, Timotheo & Lucas, 1971: 125, figs 29-31; First synonymized by Silva-Moreira et al., 2010: 69; Platnick, 2012.
Diagnosis. The male differs from those of other Vitalius species by the palpal bulb with short apical keel (Figs 1-2 and 5-8) and bifid tibial spur with narrow prolateral branch and almost straight retrolateral branch (Fig. 3). The female differs from those of other Vitalius species by urticating hair of type I having the region 'a' shorter than region 'b' (Fig. 9).
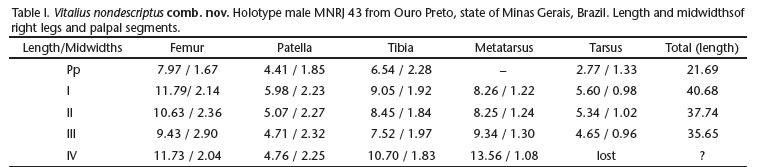
Redescription. Holotype male. MNRJ 43. Carapace 12.47 long, 11.42 wide, chelicerae 4.72. Leg measurements in Table I. Abdomen 12.70 long, 7.97 wide. Spinnerets: PMS, 1.61 long, wide, 0.26 apart; PLS, 1.55 basal, 1.07 middle, 2.55 distal; midwidths 0.94, 0.85, 0.43, respectively.
Carapace. Length to width 1.09; cephalic area moderately raised, thoracic striae shallow. Fovea: shallow, straight, wide. Carapace covered with short, slender, dense setae, bordered with stiff setae pointing out. Eyes and eye tubercle. Tubercle 0.56 high, length 1.24, width 2.00. Clypeus 0.17 wide. Anterior row procurved, posterior slightly recurved. Sizes and inter-distances: AME 0.40, ALE 0.55, PME 0.26, PLE 0.50, AME- AME 0.31, AME-ALE 0.36, AME-PME 0.35, ALE-ALE 1.07, ALE- PME 0.45, PME-PME 0.83, PME-PLE 0.14, PLE-PLE 1,62, ALE-PLE 0.19, AME-PLE 0.44. Ratio of eye group width to length 2.17. Maxillae. Length to width: 1.59. Cuspules: ca. 120 spread over ventral inner heel. Lyra absent. Labium: length 2.10, width 2.69, with ca. 105 cuspules spaced by less than one diameter from each other on anterior third center. Labio-sternal groove shallow, flat, without visible sigilla.
Chelicerae. Rastellum absent, basal segments with 12 and 11 teeth on right and left side, respectively, both with series of basal denticles. Sternum. Length 5.75, width 5.19. Posterior angle rounded, not separating coxae IV. Sigilla: 3rd and 2nd pairs ovals, one diameter from margin, 1st rounded, close to margin. Legs: leg formula: IV I II III. Clavate trichobothria: on distal 2/3 of tarsi I-IV. Leg coxae: with sparse soft setae; stridulatory or modified setae lacking. Scopula. Retrolateral femur IV scopulate. Tarsi I-III fully scopulate. Tarsus IV of both sides not found in the vial. Metatarsi I 4/5, II 2/3; III 1/3 scopulate, IV ascopulate. Spination in Table II. Claws: ITC absent from all legs; STC with small teeth. Urticating hairs. Type I with region 'a' shorter than region 'b' on the abdomen dorsum (Fig. 9).
Palp (Figs 1-2). Palpal bulb pyriform, embolus short, slightly flattened laterally at distal region. Prolateral keels present. PS forming embolus edge distally. A present, short. R present, sharp. SA present, but weakly developed. Bifid tibial spur (Fig. 3) with branches originating from common base, retrolateral longest, and with slight distal curvature; prolateral straight. Metatarsus I slightly curved, when folded touches retrolateral tibial spur branch laterally. Color pattern. Carapace dark brown with light brown hairs on its border. Coxae, labium, sternum, maxila and legs brown. Leg rings and stripes inconspicuous. Abdomen dorsally black, ventrally brown (Fig. 10).
Description. Female (Fig. 11). MZSP 465351. Carapace 12.55 long, 11.87 wide, chelicerae 7.80. Leg measurements in Table III. Abdomen 19.0 long, 10.56 wide. Spinnerets: PMS, 2.0 long, 0.97 wide, 1.22 apart; PLS, 2.96 basal, 2.21 middle .80 distal; midwidths 1.33, 1.06, 0.83, respectively. All characteristics as in the male, except:
Carapace. Length to width 1.06. Fovea: procurved, 2.89 wide. Eyes and eye tubercle. Tubercle 0.61 high, length 1.69, width 2.13. Clypeus 0.15 wide. Sizes and inter-distances. AME 0.50, ALE 0.54, PME 0.28, PLE 0.50, AME-AME 0.11, AME-ALE 0.24, AME-PME 0.10, ALE-ALE 1.18, ALE-PME 0.48, PME-PME 0.95, PME-PLE 0.16, PLE-PLE 1,56, ALE-PLE 0.22, AME-PLE 0.50. Ratio of eye group width to length 2.0. Maxillae. Length to width: 1.44. Cuspules: ca. 100 spread over ventral inner heel. Labium: length 2.53, width 2.70, with ca. 142 cuspules. Sternum. Length 6.73, width 5.88. Scopula. Tarsus IV divided by a wide row of setae. Metatarsi I 3/4, II 1/2; III 1/3 scopulate, IV ascopulate. Spination in Table IV. Genitalia. Two short spermathecae separated by short area heavily sclerotized, spermathecal stalk narrower than spermathecal bulb (Fig. 4).
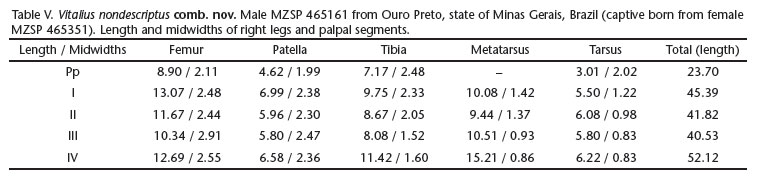
Variation. Male MZSP 465361. Carapace 14.87 long, 13.24 wide, chelicerae 4.21. Leg measurements in Table V. Abdomen 19.72 long, 17.44 wide. Spinnerets: PMS, 2.02 long, 0.70 wide, 2.13 apart; PLS, 2.68 basal, 2.06 middle, 2.69 distal; midwidths 0.87, 0.76, 0.70, respectively. Carapace. Length to width 1.12; Fovea: 2.99 wide. Eyes and eye tubercle. Tubercle 0.80 high, length 1.38, width 2.30. Clypeus 0.09 wide. Sizes and interdistances. AME 0.52, ALE 0.57, PME 0.40, PLE 0.59, AME-AME 0.13, AME-ALE 0.29, AME-PME 0.05, ALE-ALE 1.09, ALE-PME 0.42, PME-PME 0.89, PME-PLE 0.16, PLE-PLE 1.51, ALE-PLE 0.21, AME-PLE 0.49. Ratio of eye group width to length 1.84. Maxillae. Length to width: 1.29. Cuspules: ca.100 spread over ventral inner heel. Labium: length 1.81, width 2.53, with ca. 142 cuspules. Chelicerae. Rastellum absent, basal segments with 10 and 11 teeth on right and left side, respectively, both with series of basal denticles. Sternum. Length 6.72, width 6.00. Scopula. Tarsus IV divided by a wide row of setae. Spination in Table VI. Palp (Figs 5-6). SA present, developed.
Male MZSP 465371. Carapace 12.66 long, 11.01 wide, chelicerae 4.73. Leg measurements in Table VII. Abdomen 15.34 long, 10.72 wide. Spinnerets: PMS, 1.46 long, 0.61 wide, 1.01 apart; PLS, 2.88 basal, 2.21 middle, 2.58 distal; midwidths 0.92, 0.77, 0.55, respectively. Carapace. Length to width 1.15; Fovea: 2.75 wide. Eyes and eye tubercle. Tubercle 0.79 high, length 1.22, width 2.15. Clypeus 0.16 wide. Sizes and inter-distances: AME 0.47, ALE 0.55, PME 0.27, PLE 0.42, AME-AME 0.16, AME-ALE 0.19, AME-PME 0.05, ALE-ALE 1.14, ALE-PME 0.27, PME-PME 0.79, PME-PLE 0.14, PLE-PLE 1.32, ALE-PLE 0.13, AME-PLE 0.40. Ratio of eye group width to length 1.95. Maxillae. Length to width: 1.37. Cuspules: ca.110 spread over ventral inner heel. Labium: length 1.78, width 2.21, with ca. 130 cuspules. Chelicerae: basal segments with 12 and 11 teeth on right and left side, respectively, both with series of basal denticles. Sternum. Length 5.31, width 5.32. Scopula. Tarsus IV divided by a wide row of setae. Spination in Table VIII. Palp (Figs 7-8). SA present, developed. Urticating hairs. Mostly with type I with region 'a' shorter than region 'b' (Fig. 9) and a small patch of type III on postero-dorsal area on the abdomen dorsum.
Type material. Holotype male from BRAZIL, Minas Gerais: Ouro Preto (MNRJ 43) (examined).
Additional material examined. BRAZIL: Minas Gerais: Ouro Preto (P.E. Itacolomi, 20º26'0.96"S, 43º30'7.48"W, 1.318 m a.s.l.), 1 female found with egg sac, 11 January 2005, R. Bertani, C.S. Fukushima, M.P.M. Pavani, R. Martins & K.M. Faria leg., MZSP 465351; 2 males, captive born from female MZSP 465351, MZSP 465161 and MZSP 465171; Caeté (Serra da Gandarela, 20º05'S, 43º38'W, 1.100 m a.s.l.), 1 male, pit-fall, 25/XI/2011, MNRJ 4431; Santa Bárbara (Serra do Caraça, 20º05'S, 43º29'W, 1.200 m a.s.l.), 1 male, 12-23/XI/1961, U.M. Lenko-Kloss leg., MZSP 14.895.
Distribution. Brazil, State of Minas Gerais, in the highlands (up 1100 m a.s.l.) of Serras do Itacolomi, Caraça, and Gandarela.
Natural History. The female MZSP 465351 was found with egg sac on 11 January 2005 under a fallen tree trunk. The mother and the egg sac were maintained in captivity and on 28 January 2005, 52 spiderlings emerged from the egg sac. Of these, two males reached maturity, in January 2011 and February 2012.
DISCUSSION
When comparing the holotype of H. nondescriptus with freshly collected specimens from the type locality, we found agreement in most morphological characteristics such as size, color pattern, legs and palpal spination, tibial spur shape, folding of metatarsus I contacting the retrolateral spur branch, the short embolus with short apical keel (Figs 1-2), and the morphology of type I urticating hair with region 'a' shorter than region 'b' (Fig. 9). On the other hand, the subapical keel in the new examined material is well-developed, similar to those seen in Vitalius Lucas, Silva Junior & Bertani, 1993, Nhandu Lucas, 1982 and Lasiodora C.L. Koch, 1850 species (Figs 5-8). Thus, the only difference noted among the holotype and the new material concerns the development of the subapical keel, which can vary for some taxa, e.g. Vitalius vellutinus (Mello-Leitão, 1923) from small to vestigial (BERTANI 2001). It is more plausible to consider that the holotype presents developmental variation of the subapical keel rather than having two very similar, sympatric species share all the characters shown above, yet only differing by a minor variance in bulb morphology.
The species herein under study is a theraphosine with type I urticating hair on the abdomen dorsum, spermathecae fused at a small area (Fig. 4), absence of long hairs on female's carapace, a tibial spur with two branches originating from a common base (Fig. 3), presence of retrolateral scopulae on femur IV, absence of stridulatory setae on trochanter and coxae of all legs and palp, and presence of the following keels on male bulb: prolateral superior, prolateral inferior, retrolateral, apical and subapical keel (Figs 1-2 and 5-8). This set of characters indicates close affinities with Vitalius. Therefore, we transfered Hapalopus nondescriptus to Vitalius, making the new combination Vitalius nondescriptus (Mello-Leitao, 1926) comb. nov.
However, in specimens of V. nondescriptus comb. nov. the apical keel is small (Figs 1-2 and 5-8) and the tibial spur has a straighter retrolateral branch and a narrow prolateral branch (Fig. 3), unlike other species of the genus. Most Vitalius species present a medially developed apical keel and tibial spur with curved retrolateral branch and thickened prolateral branch (BERTANI 2001). Furthermore, the type I urticating hair with a short region 'a', when comparing with region 'b' (Fig. 9), is found in two species of Proshapalopus Mello-Leitão, 1923 - Proshapalopus anomalus Mello-Leitão, 1923 and Proshapalopus multicuspidatus (Mello-Leitão, 1929), in species of Ami Pérez-Miles, 2008, and in Citharacanthus livingstoni Schmidt & Weinmann, 1996 (PÉREZ-MILES et al. 2008), but not in Vitalius (BERTANI 2001). The presence of this urticating hair type I modification is clearly homoplasy among the three genera (above) and V. nondescriptus comb. nov., because they are not closely related (see cladograms of PÉREZ-MILES et al. 1996 for Citharacanthus, BERTANI 2001 for Proshapalopus, and PÉREZ-MILES et al. 2008 for Ami). On the other hand, the characters short apical keel and tibial spur shape could represent homoplasious or plesiomorphic characters in relation to the related Lasiodora (short apical keel and tibial spur shape) and Nhandu (tibial spur shape).
ACKNOWLEDGMENTS
We thank Instituto Estadual de Florestas (IEF) of the state of Minas Gerais and Alberto V.M. Matos, Park Manager of the Parque Estadual do Itacolomi, for collection permits and aiding in the field trip; Rosana Martins, Maurício P.M. Pavani and Kátia M. Faria for aiding in field work. We also thank Adriano Kury and Ricardo Pinto-da-Rocha for specimens' loan and for providing a repository for collected specimens. Support: FAPESP 03/12587-4 for RB.
LITERATURE CITED
Submitted: 16.IV.2012; Accepted: 23.V.2012.
Editorial responsibility: Walter A.P. Boeger
- BERTANI, R. 2000. Male palpal bulbs and homologous features in Theraphosinae (Araneae, Theraphosidae). The Journal of Arachnology 28:29-42. doi: 10.1636/0161-8202(2000)028[0029:MPBAHF]2.0.CO;2
- BERTANI, R. 2001. Revision, cladistic analysis, and zoogeography of Vitalius, Nhandu, and Proshapalopus; with notes on other theraphosine genera (Araneae, Theraphosidae). Arquivos de Zoologia 36:265-356.
- BÜCHERL, W.; A. TIMOTHEO DA COSTA & S. LUCAS. 1971. Revisão de alguns tipos de aranhas caranguejeiras (Orthognatha) estabelecidos por Candido de Mello-Leitão e depositados no Museu Nacional do Rio. Memórias do Instituto Butantan 35:117-138.
- COOKE J.A.L.; V.D. ROTH & F.H. MILLER. 1972. The urticating hairs of theraphosid spiders. American Museum Novitates 2498:1-43.
- MELLO-LEITÃO, C.F. 1926. Algumas Theraphosoideas novas do Brasil. Revista do Museu Paulista 14:307-324.
- PÉREZ-MILES, F.; S.M. LUCAS; P.I. SILVA JUNIOR & R. BERTANI. 1996. Systematic revision and cladistic analysis of Theraphosinae (Araneae: Theraphosidae). Mygalomorph 1:33-68.
- PÉREZ-MILES, F.; R. GABRIEL; L. MIGLIO; A. BONALDO; R. GALLON; J. J. JIMENEZ & R. BERTANI. 2008. Ami, a new theraphosid genus from Central and South America, with the description of six new species (Araneae: Mygalomorphae). Zootaxa 1915:54-68.
- PETRUNKEVITCH, A. 1925. Arachnida from Panama. Transactions of Connecticut Academy of Arts and Sciences 27:51-248.
- PLATNICK, N.I. 2012. The world spider catalog version 12.5. American Museum of Natural History. Available online at: http://research.amnh.org/entomology/spider/catalog/index.html [Acessed: February 2012]
- RAVEN, R.J. 1990. Comments on the proposed precedence of Aphonopelma Pocock, 1901 (Arachnida, Araneae) over Rhechostica Simon, 1892. Bulletin of Zoological Nomenclature 47:126-127.
- RAVEN, R.J. 2005. A new tarantula species from northern Australia (Araneae, Theraphosidae). Zootaxa 1004:15-28.
- SILVA-MOREIRA, T. DA; R.L.C. BAPTISTA; A.B. KURY; A.P.L. GIUPPONI; E.H. BUCKUP & A.D. BRESCOVIT. 2010. Annotated check list of Arachnida type specimens deposited in the Museu Nacional, Rio de Janeiro. II-Araneae. Zootaxa 2588:1-91.
Publication Dates
-
Publication in this collection
13 Nov 2012 -
Date of issue
Oct 2012
History
-
Received
16 Apr 2012 -
Accepted
23 May 2012