ABSTRACT
The importance of amphibian bioacoustics is widely documented in ecological, taxonomical and evolutionary studies, as calls might act as a primary mechanism of reproductive isolation. The influence of air temperature and male size on the variation of the advertisement calls in anurans is widely recognized but still insufficiently analyzed in Brazilian species. Our goals were to (1) describe the advertisement call of Aplastodiscus albosignatus (Lutz & Lutz, 1938), (2) test the influence of temperature and body size on its acoustic signals and (3) evaluate the variation of within and between-male acoustic signals. Advertisement calls of A. albosignatus consist of an unpulsed note with four harmonics. In most cases, the dominant frequency is the third harmonic but, in some calls, it was the minimum frequency (first harmonic). The average duration of calls was 0.191 s, the interval between calls was 2.08 s and the repetition rate was 33 calls per minute. On average, the minimum frequency was 550.15 Hz, the maximum frequency was 3531.70 Hz and the third harmonic was 2498.9 Hz. To evaluate the effect of air temperature, and body size on the variation of call parameters, we performed generalized linear models. The most explanatory model for spectral parameters was temperature plus body size. Concerning temporal variables, the best model that explains the variation in call duration was body size, while for the interval between calls was air temperature. The maximum frequency and the frequency of the third harmonic had little variation in the calls of both the same male and different males. Thus, these parameters were considered important in species recognition.
KEY WORDS:
Albosignatus group; Atlantic Forest; coefficient of variation; dominant frequency; harmonics
INTRODUCTION
Acoustic signals represent the main form of intra- and inter-specific communication in anurans. These signals are a fundamental characteristic of the natural history of this group and are mainly associated with sexual selection, reproduction, rival assessment and recognition (Wells 2010Wells KD (2010) The ecology and behavior of amphibians. Chicago, University of Chicago Press, 1148 pp., Toledo et al. 2014Toledo LF, Martins IA, Bruschi DP, Passos MA, Alexandre C, Haddad CFB (2014) The anuran calling repertoire in the light of social context. Acta Ethologica 18: 87-99. https://doi.org/10.1007/s10211-014-0194-4
https://doi.org/10.1007/s10211-014-0194-...
, Bee et al. 2016Bee MA, Reichert MS, Tumulty J (2016) Assessment and recognition of rivals in anuran contests. Advances in the Study of Behavior 48: 161-249.). The most common vocalizations emitted by males are the advertisement calls, which are species-specific and function as a mechanism of pre-zygotic reproductive isolation (Wells 2010Wells KD (2010) The ecology and behavior of amphibians. Chicago, University of Chicago Press, 1148 pp., Gambale and Bastos 2014Gambale PG, Bastos RP (2014) Vocal repertoire and bioacoustic analyses in Physalaemus cuvieri (Anura, Leptodactylidae) from southern Brazil. Herpetological Journal 24: 31-39., Guerra et al. 2018Guerra V, Llusia D, Gambale PG, Morais AR, Marquez R, Bastos RP (2018) The advertisement calls of Brazilian anurans: Historical review, current knowledge and future directions. PLoS ONE 13: 1-22. https://doi.org/10.1371/journal.pone.0191691
https://doi.org/10.1371/journal.pone.019...
). These acoustic signals contain important information for females and possible competitors, indicating, for example, the individual’s position, body size, body temperature and energetic condition (Wells 2010Wells KD (2010) The ecology and behavior of amphibians. Chicago, University of Chicago Press, 1148 pp., Toledo et al. 2014Toledo LF, Martins IA, Bruschi DP, Passos MA, Alexandre C, Haddad CFB (2014) The anuran calling repertoire in the light of social context. Acta Ethologica 18: 87-99. https://doi.org/10.1007/s10211-014-0194-4
https://doi.org/10.1007/s10211-014-0194-...
).
Since anurans are ectothermic animals, their acoustic signals can be influenced by abiotic factors such as temperature, resulting in differences between individuals (Robertson 1984Robertson GM (1984) Acoustic Spacing by Breeding Males of Uperoleia rugosa (Anura: Leptodactylidae). Zeitschrift für Tierpsychologie 297: 283-297. https://doi.org/10.1111/j.1439-0310.1984.tb00364.x
https://doi.org/10.1111/j.1439-0310.1984...
, Giacoma et al. 1997Giacoma C, Zugolaro C, Beani L (1997) The Advertisement Calls of the Green Toad (Bufo viridis): Variability and Role in Mate Choice. Herpetologica 53: 454-464., Navas and Bevier 2001Navas CA, Bevier CR (2001) Thermal dependency of calling performance in the eurythermic frog Colostethus subpunctatus. Herpetologica 57: 384-395., Lingnau and Bastos 2007Lingnau R, Bastos RP (2007) Vocalizations of the Brazilian torrent frog Hylodes heyeri (Anura: Hylodidae): Repertoire and influence of air temperature on advertisement call variation. Journal of Natural History 41: 17-20. https://doi.org/10.1080/00222930701395626
https://doi.org/10.1080/0022293070139562...
). Some species reduce the duration of the call at higher temperatures, which may be related to an attempt to avoid increased metabolism and energy expenditure (Lingnau and Bastos 2007Lingnau R, Bastos RP (2007) Vocalizations of the Brazilian torrent frog Hylodes heyeri (Anura: Hylodidae): Repertoire and influence of air temperature on advertisement call variation. Journal of Natural History 41: 17-20. https://doi.org/10.1080/00222930701395626
https://doi.org/10.1080/0022293070139562...
, Boschetti et al. 2019Boschetti JP, De Bastiani VIM, Lingnau R, Lucas EM (2019) Bioacoustics of Pithecopus rusticus (Anura, Phyllomedusidae): a rare species possibly threatened with extinction. South American Journal of Herpetology 14: 196-203. https://doi.org/10.2994/SAJH-D-17-00071.1
https://doi.org/10.2994/SAJH-D-17-00071....
). For example, the temperature can influence the rate of call emissions and pulse repetition, while spectral parameters (e.g., minimum frequency), which are not related to muscular contraction, can be less affected by temperature (Wong et al. 2004Wong BBM, Cowling ANN, Cunningham RB, Donnelly CF, Cooper PD (2004) Do temperature and social environment interact to affect call rate in frogs (Crinia signifera)? Austral Ecology 29: 209-214. https://doi.org/10.1111/j.1442-9993.2004.01338.x
https://doi.org/10.1111/j.1442-9993.2004...
).
Body size is also a factor that can influence a species’ acoustic signals (Castellano et al. 2002Castellano S, Cuatto B, Rinella R, Rosso A (2002) The advertisement call of the European treefrogs, Hyla arborea: a multi-level study of variation. Ethology 108: 75-89. https://doi.org/10.1046/j.1439-0310.2002.00761.x
https://doi.org/10.1046/j.1439-0310.2002...
, Bernal et al. 2005Bernal XE, Guarnizo CE, Luddecke H (2005) Geographic variation in advertisement call and genetic structure of Colostethus palmatus (Anura, Dendrobatidae) from the Colombian Andes Herpetologists’ League. Herpetologica 61: 395-408. https://doi.org/10.1655/04-87.1
https://doi.org/10.1655/04-87.1...
, Wells 2010Wells KD (2010) The ecology and behavior of amphibians. Chicago, University of Chicago Press, 1148 pp.). In general, spectral signals such as the dominant frequency are more influenced by the size of the individual, while temporal parameters tend to be more influenced by air temperature (e.g., Lemes et al. 2012Lemes P, Tessarolo G, Morais AR, Bastos RP (2012) Acoustic Repertoire of Barycholos ternetzi (Anura: Strabomantidae) in Central Brazil. South American Journal of Herpetology 7: 157-164. https://doi.org/10.2994/057.007.0205
https://doi.org/10.2994/057.007.0205...
, Morais et al. 2012Morais AR, Batista VG, Gambale PG, Signorelli L, Bastos RP (2012) Acoustic communication in a Neotropical frog (Dendropsophus minutus): vocal repertoire, variability and individual discrimination. Herpetological Journal 22: 249-257., Bee et al. 2013Bee MA, Suyesh R, Biju SD (2013) The vocal repertoire of Pseudophilautus kani, a shrub frog (Anura: Rhacophoridae) from the Western Ghats of India. Bioacoustics 22: 67-85. https://doi.org/10.1080/09524622.2012.712750
https://doi.org/10.1080/09524622.2012.71...
). This occurs due to the influence of the size of the vocal cords and other components of the larynx (Wong et al. 2004Wong BBM, Cowling ANN, Cunningham RB, Donnelly CF, Cooper PD (2004) Do temperature and social environment interact to affect call rate in frogs (Crinia signifera)? Austral Ecology 29: 209-214. https://doi.org/10.1111/j.1442-9993.2004.01338.x
https://doi.org/10.1111/j.1442-9993.2004...
). Understanding how these variations occur in the advertisement calls of species contributes to a better taxonomic delimitation and can generate an ecological basis for hypotheses about the change in acoustic traits over time (Castellano et al. 2002Castellano S, Cuatto B, Rinella R, Rosso A (2002) The advertisement call of the European treefrogs, Hyla arborea: a multi-level study of variation. Ethology 108: 75-89. https://doi.org/10.1046/j.1439-0310.2002.00761.x
https://doi.org/10.1046/j.1439-0310.2002...
).
The calls formed by a single type of note are called simple calls, while those formed by different notes are called complex calls (Köhler et al. 2017Köhler J, Jansen M, Rodríguez A, Kok PJR, Toledo LF, Emmrich M, Glaw F, Haddad CFB, Rödel MO, Vences M (2017) The use of bioacoustics in anuran taxonomy: Theory, terminology, methods and recommendations for best practice. Zootaxa 4251: 001-124. https://doi.org/10.11646/zootaxa.4251.1.1
https://doi.org/10.11646/zootaxa.4251.1....
). Simple calls are common in many species, such as those of the genera Physalaemus (e.g., Baêta et al. 2007Baêta D, Lourenço ACC, Pezzuti TL, Pires MRS (2007) The tadpole, advertisement call, and geographic distribution of Physalaemus maximus Feio, Pombal & Caramaschi, 1999 (Amphibia, Anura, Leiuperidae). Arquivos do Museu Nacional 65: 27-32., Gambale and Bastos 2014Gambale PG, Bastos RP (2014) Vocal repertoire and bioacoustic analyses in Physalaemus cuvieri (Anura, Leptodactylidae) from southern Brazil. Herpetological Journal 24: 31-39., Hepp et al. 2020Hepp F, Pombal J Jr (2020) Review of bioacoustical traits in the genus Physalaemus Fitzinger, 1826 (Anura: Leptodactylidae: Leiuperinae). Zootaxa 4725(1): 1-106. https://doi.org/10.11646/zootaxa.4725.1.1
https://doi.org/10.11646/zootaxa.4725.1....
) and Aplastodiscus (e.g., Abrunhosa et al. 2005Abrunhosa PA, Pimenta BVS, Cruz CAG, Haddad CFB (2005) Advertisement Calls of Species of the Hyla Albosignata Group (Amphibia, Anura, Hylidae). Arquivos do Museu Nacional 63: 275-282., Zina and Haddad 2006Zina J, Haddad CFB (2006) Acoustic repertoire of Aplastodiscus arildae and A. leucopygius (Anura: Hylidae) in Serra do Japi, Brazil. South American Journal of Herpetology 1: 227-236. https://doi.org/10.2994/1808-9798(2006)1[227:AROAAA]2.0.CO;2
https://doi.org/10.2994/1808-9798(2006)1...
). Complex calls, with a larger number of notes, and greater variation of these notes, are found in other Neotropical species such as Dendropsophus minutus (Peters, 1872) and Bokermannohyla ibitiguara (Cardoso, 1983) (Nali and Prado 2014Nali RC, Prado CP (2014) Complex call with different messages in Bokermannohyla ibitiguara (Anura, Hylidae), a gladiator frog of the Brazilian Cerrado. Journal of Herpetology 48: 407-414., Turin et al. 2018Turin RAF, Nali RC, Prado CPA (2018) Intraspecific call variation in a Neotropical gladiator frog with a complex advertisement call. Amphibia Reptilia 39: 31-39. https://doi.org/10.1163/15685381-00003147
https://doi.org/10.1163/15685381-0000314...
).
Among the species that present simple calls, Aplastodiscus Lutz, 1950 (Hylidae) currently comprises 16 described species (Berneck et al. 2016Berneck BVM, Haddad CFB, Lyra ML, Cruz CAG, Faivovich J (2016) The Green Clade grows: A phylogenetic analysis of Aplastodiscus (Anura; Hylidae). Molecular Phylogenetics and Evolution 97: 213-223. https://doi.org/10.1016/j.ympev.2015.11.014
https://doi.org/10.1016/j.ympev.2015.11....
, Frost 2021Frost DR (2021) Amphibian Species of the World: an Online Reference. Version 6.1 (29/06/2021). Available online at Available online at https://amphibiansoftheworld.amnh.org/index.php
https://amphibiansoftheworld.amnh.org/in...
, Pinheiro et al. 2021Pinheiro PD, Pezzuti TL, Berneck BV, Lyra ML, Lima RC, Leite FS (2021) A new cryptic species of the Aplastodiscus albosignatus group (Anura: Hylidae). Salamandra 57: 27-43.) all with simple advertising calls (Abrunhosa et al. 2005Abrunhosa PA, Pimenta BVS, Cruz CAG, Haddad CFB (2005) Advertisement Calls of Species of the Hyla Albosignata Group (Amphibia, Anura, Hylidae). Arquivos do Museu Nacional 63: 275-282., Zina and Haddad 2006Zina J, Haddad CFB (2006) Acoustic repertoire of Aplastodiscus arildae and A. leucopygius (Anura: Hylidae) in Serra do Japi, Brazil. South American Journal of Herpetology 1: 227-236. https://doi.org/10.2994/1808-9798(2006)1[227:AROAAA]2.0.CO;2
https://doi.org/10.2994/1808-9798(2006)1...
, Berneck et al. 2016Berneck BVM, Haddad CFB, Lyra ML, Cruz CAG, Faivovich J (2016) The Green Clade grows: A phylogenetic analysis of Aplastodiscus (Anura; Hylidae). Molecular Phylogenetics and Evolution 97: 213-223. https://doi.org/10.1016/j.ympev.2015.11.014
https://doi.org/10.1016/j.ympev.2015.11....
, Bezerra et al. 2020Bezerra AM, Passos LO, De Luna-Dias C, Quintanilha AS, De Carvalho-e-Silva SP (2020) A missing piece of the puzzle: Re-encounter of Aplastodiscus musicus, its call, and phylogenetic placement (Anura: Hylidae: Cophomantini). Herpetologica 76: 74-82. https://doi.org/10.1655/Herpetologica-D-18-00061
https://doi.org/10.1655/Herpetologica-D-...
). Aplastodiscus albosignatus (Lutz & Lutz, 1938) is endemic to the Atlantic Forest and associated with streams in mountainous regions (< 850 m). The species’ distribution was originally considered to extend from the state of Minas Gerais to Santa Catarina. A recent taxonomic study (Berneck et al. 2016Berneck BVM, Haddad CFB, Lyra ML, Cruz CAG, Faivovich J (2016) The Green Clade grows: A phylogenetic analysis of Aplastodiscus (Anura; Hylidae). Molecular Phylogenetics and Evolution 97: 213-223. https://doi.org/10.1016/j.ympev.2015.11.014
https://doi.org/10.1016/j.ympev.2015.11....
), however, suggests that the populations previously recognized as A. albosignatus that occur in southern São Paulo, as well as in Paraná and Santa Catarina, consist of an undescribed species (Aplastodiscus sp. 5). The advertisement call of A. albosignatus was described by Abrunhosa et al. (2005Abrunhosa PA, Pimenta BVS, Cruz CAG, Haddad CFB (2005) Advertisement Calls of Species of the Hyla Albosignata Group (Amphibia, Anura, Hylidae). Arquivos do Museu Nacional 63: 275-282.) based on 10 calls of only one individual. To date, there are no other analyzed recordings of the vocalization of this species (Guerra et al. 2018Guerra V, Llusia D, Gambale PG, Morais AR, Marquez R, Bastos RP (2018) The advertisement calls of Brazilian anurans: Historical review, current knowledge and future directions. PLoS ONE 13: 1-22. https://doi.org/10.1371/journal.pone.0191691
https://doi.org/10.1371/journal.pone.019...
). In the present study, we extend previous knowledge on the calls of A. albosignatus by: 1) providing a more robust description of the advertisement call; 2) examining the influence of abiotic (temperature) and biotic (body size) factors on the acoustic signals and (3) evaluating the intra-individual and inter-individual variation of the calls. To fulfill these goals, we expect that, as already described for other Neotropical species, 1) air temperature will be negatively associated with variation of temporal parameters of the advertisement calls (Lingnau and Bastos 2007Lingnau R, Bastos RP (2007) Vocalizations of the Brazilian torrent frog Hylodes heyeri (Anura: Hylodidae): Repertoire and influence of air temperature on advertisement call variation. Journal of Natural History 41: 17-20. https://doi.org/10.1080/00222930701395626
https://doi.org/10.1080/0022293070139562...
. Köhler et al. 2017Köhler J, Jansen M, Rodríguez A, Kok PJR, Toledo LF, Emmrich M, Glaw F, Haddad CFB, Rödel MO, Vences M (2017) The use of bioacoustics in anuran taxonomy: Theory, terminology, methods and recommendations for best practice. Zootaxa 4251: 001-124. https://doi.org/10.11646/zootaxa.4251.1.1
https://doi.org/10.11646/zootaxa.4251.1....
), 2) body size will be negatively associated with spectral parameters (Lingnau and Bastos 2007Lingnau R, Bastos RP (2007) Vocalizations of the Brazilian torrent frog Hylodes heyeri (Anura: Hylodidae): Repertoire and influence of air temperature on advertisement call variation. Journal of Natural History 41: 17-20. https://doi.org/10.1080/00222930701395626
https://doi.org/10.1080/0022293070139562...
, Köhler et al. 2017Köhler J, Jansen M, Rodríguez A, Kok PJR, Toledo LF, Emmrich M, Glaw F, Haddad CFB, Rödel MO, Vences M (2017) The use of bioacoustics in anuran taxonomy: Theory, terminology, methods and recommendations for best practice. Zootaxa 4251: 001-124. https://doi.org/10.11646/zootaxa.4251.1.1
https://doi.org/10.11646/zootaxa.4251.1....
) and 3) the variation of acoustic signals will be greater between males (inter-individual) than the individual variation itself (intra-individual).
MATERIAL AND METHODS
The study was carried out in lotic environments of a subtropical forest (25°29’46”S 48°58’58”W, 1000 a.s.l.) of the Atlantic Forest, municipality of Piraquara, state of Paraná, southern Brazil (Figs 1-3). The collections of individuals occurred at Mananciais da Serra, more specifically on the Ipiranga Trail, which is about seven kilometers long. The sampled region is located in a transition zone between the phytophysiognomies Mixed Ombrophilous Forest, dominated by Araucaria angustifolia (Araucaria Forest) and Dense Ombrophilous Forest (Reginato and Goldenberg 2007Reginato M, Goldenberg R (2007) Análise florística, estrutural e fitogeográfica da vegetação em região de transição entre as Florestas Ombrófilas Mista e Densa Montana, Piraquara, Paraná, Brasil. Hoehnea 34: 349-360. https://doi.org/10.1590/S2236-89062007000300006
https://doi.org/10.1590/S2236-8906200700...
). Annual rainfall ranges from 1400 to 1700 mm, with rainfall concentrated from October to March (Paraná 1987Paraná (1987) Atlas do estado do Paraná. Curitiba, Secretaria do Estado da Agricultura e do Abastecimento, Intitutos de Terras, Cartografias e Florestas, 486 pp.). The average temperature is 22 °C in the hottest months and 18 °C in the coldest months (Ferreira 1996Ferreira JCV (1996) O Paraná e seus municípios. Curitiba, Editora Memória Brasileira, 728 pp.).

Location of the study area: (1) map of Brazil highlighting the state of Paraná; (2) map of Paraná indicating the study area; (3) map of Paraná highlighting the physiognomy of the vegetation.
We recorded 22 males of A. albosignatus between January 2019 and February 2020. When vocalizing males were located, at first their vocalization was recorded. The distance between the microphone and the individual was approximately 50 cm. After recording, we captured and individualized the amphibian and recorded air temperature with a thermometer (0.1 °C precision). We recorded the vocalizations with a CSR HT81 - YOGA unidirectional microphone coupled to a Tascam DR-40 recorder. After the collections, we weighed and measured the snout-vent length (SVL) of the recorded males with a scale (0.01 g precision) and digital caliper (0.05 mm precision). Afterward, we euthanized individuals with xylocaine, fixed them in 10% formalin and conserved them in 70% ethanol. The collections were carried out with the competent Federal Organ authorization, SISBIO (authorization #66853-2) and also with authorization of the Ethics Committee for Animal Use from the Biological Sciences Section of the Federal University of Paraná (CEUA/BIO, UFPR, process number 23075.043325/2019-27). The collected individuals were stored in the Herpetological Collection of the Universidade Tecnológica Federal do Paraná, Francisco Beltrão Campus (RLUTF 251, 252, 341, 343, 467, 468, 469, 489, 715, 716, 718, 732, 733, 734, 735, 736, 737, 749, 1218, 1219, 1220 and 1221). Call recordings were deposited at the Fonoteca Neotropical Jacques Vielliard, where they can be accessed under the codes FNJV 45561 - 45582.
We analyzed 220 calls from 22 males with Raven pro 1.6 software for Windows (Cornell Lab of Ornithology, Bioacoustics Research Program 2011), using FFT (Fast Fourier Transformation) = 256. There was no need to use filters. The following acoustic parameters were analyzed: minimum frequency (Hz), maximum frequency (Hz), frequency of the third harmonic (Hz), call duration (s), interval between calls (s) and repetition rate (calls/min). Spectral measurements were obtained by selecting three variables in the “choose measurements” option in Raven 1.6: (1) low frequency, (2) high frequency and (3) peak frequency. The bioacoustic terms were used according to definitions proposed by Köhler et al. (2017Köhler J, Jansen M, Rodríguez A, Kok PJR, Toledo LF, Emmrich M, Glaw F, Haddad CFB, Rödel MO, Vences M (2017) The use of bioacoustics in anuran taxonomy: Theory, terminology, methods and recommendations for best practice. Zootaxa 4251: 001-124. https://doi.org/10.11646/zootaxa.4251.1.1
https://doi.org/10.11646/zootaxa.4251.1....
), with an adaptation for calculating the dominant frequency. The dominant frequency values in A. albosignatus can be in the first or third harmonic (see data below), and therefore an average of the dominant frequency values could not be provided, which would be an uninformative value. Thus, we inform the values of minimum frequency (first harmonic) and third harmonic that better illustrate the location of the dominant frequency.
We generated oscillograms and spectrograms using R software (R Development Core Team 2016R Development Core Team (2016) R: A language and environment for statistical computing. Version 3.3.2, available online at http://www.R-project.org
http://www.R-project.org...
), with a window width of 512 samples, 90% overlap and FFT of 512 samples. We used the packages TuneR version 1.0 (Ligges et al. 2013Ligges U, Krey S, Mersmann O, Schnackenberg S (2013) Tuner: Analysis of music. Available online at Available online at http://r-forge.r-project.org/projects/tuner [Accessed 15/06/2020]
http://r-forge.r-project.org/projects/tu...
) and Seewave version 1.7.3 (Sueur et al. 2008Sueur J, Aubin T, Simonis C (2008) Seewave: a free modular tool for sound analysis and synthesis. Bioacoustics 18: 213-226. https://doi.org/10.1080/09524622.2008.9753600
https://doi.org/10.1080/09524622.2008.97...
).
To evaluate the effect of air temperature and body size on call duration, the interval between calls, fundamental frequency, maximum frequency and peak frequency of the third harmonic, we performed generalized linear models using Gauss distribution in R software (version 4.1.0) and the following packages: corrplot (Wei et al. 2017Wei T, Simko V, Levy M, Xie Y, Jin Y, Zemla J (2017). Package ‘corrplot’. Statistician 56: e24. https://cran.r-project.org/web/packages/corrplot/corrplot.pdf
https://cran.r-project.org/web/packages/...
) fBasics (Wuertz et al. 2007Wuertz D, Wuertz MD, Team RC (2007) The fBasics Package. https://cran.r-project.org/web/packages/fBasics/index.html
https://cran.r-project.org/web/packages/...
), DescTools (Signorell et al. 2016Signorell A, Aho K, Alfons A, Anderegg N, Aragon T (2016) DescTools: Tools for descriptive statistics. R package, v. 0.99.18, Vienna, R Foundation for Statistical Computing. https://cran.rproject.org/web/packages/DescTools/index.html
https://cran.rproject.org/web/packages/D...
). We multiplied mass by snout-vent length (mass * SVL) to obtain a ‘body size’ variable that was used for hypothesis testing. We checked the normality of the variables using the Shapiro Wilk test. We found that the interval between calls and peak frequency of the third harmonic did not have a normal distribution. Thus, all variables were transformed by square root. We tested five hypotheses, where the dependent variables (call duration, interval between calls, minimum frequency, maximum frequency and peak frequency of the third harmonic) are influenced by body size plus temperature (e.g. call duration ~ body size + temperature). To choose the best model, we used Akaike’s Information Criterion (AIC, Arnold 2010Arnold TW (2010) Uninformative parameters and model selection using Akaike’s Information Criterion. The Journal of Wildlife Management 74: 1175-1178. https://doi.org/10.2193/2009-367
https://doi.org/10.2193/2009-367...
).
The variability of the acoustic parameters of the calls of each male (intra-individual) and between males (inter-individual) was analyzed from the variation coefficient for each parameter, as proposed by Gerhardt (1991Gerhardt HC (1991) Female mate choice in treefrogs: static and dynamic acoustic criteria. Animal Behaviour 42: 615-635. https://doi.org/10.1016/S0003-3472(05)80245-3
https://doi.org/10.1016/S0003-3472(05)80...
), where . The intra-individual variation coefficient (CVintra) considers the mean and standard deviation of the calls of each individual. The interindividual coefficient of variation (CVinter) is considered the mean and standard deviation of the parameters of all individuals. When the variation coefficient is equal to or less than 5%, the evaluated parameter is considered to be static, and when it is greater than or equal to 12%, it is considered to be dynamic (Gerhardt 1991Gerhardt HC (1991) Female mate choice in treefrogs: static and dynamic acoustic criteria. Animal Behaviour 42: 615-635. https://doi.org/10.1016/S0003-3472(05)80245-3
https://doi.org/10.1016/S0003-3472(05)80...
).
To assess whether there is variability between male vocalizations, the ratio of the two variation coefficients was calculated (CVinter/CVintra). When the value obtained is CVinter/CVintra > 1, the evaluated acoustic signal assists in individual recognition (Márquez and Eekhout 2006Márquez R, Eekhout X (2006) Advertisement calls of six species of anurans from Bali, Republic of Indonesia. Journal of Natural History 40: 9-10. https://doi.org/10.1080/00222930600712129
https://doi.org/10.1080/0022293060071212...
).
RESULTS
The individuals had an average size of 42.6 mm ± 1.8 mm (39.4-45.8 mm) and an average mass of 4.66 ± 0.5 g (3.8-5.7 g). The average temperature recorded during the samplings was 16.5 ± 1.7 °C (13-19 °C). Individuals were found vocalizing on tree branches, usually more than two meters high.
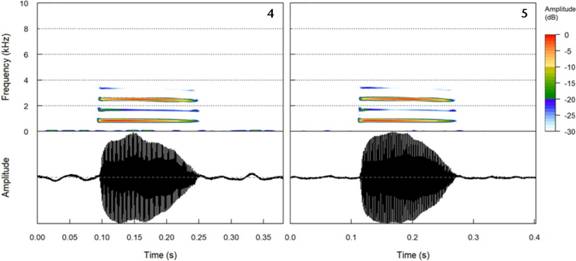
The advertisement call of A. albosignatus is formed by a single note and a single pulse, with four visible harmonics. In general, the dominant frequency occurs in the third harmonic but, in some calls, the dominant frequency was recorded at the minimum frequency (first harmonic) (Table 1, Figs 4, 5). In a total of 220 analyzed calls, the dominant frequency was in the third harmonic in 145 calls (66%), and in the first harmonic in 75 calls (34%). The same male could present calls with the dominant frequency in the first or third harmonic. The mean duration of the calls was 0.191 ± 0.028 s (0.12-0.25), the interval between calls was 2.08 ± 1.20 s (0.56-9.61) and the repetition rate was 33 ± 14.73 calls per minute (15-66) (Table 1). On average, the minimum frequency was 550.15 ± 42 Hz (421.09-689.06), the maximum frequency 3531.70 ± 144.20 Hz (3253.9-3828.12) and the third harmonic 2498.9 ± 100.3 Hz (2239.45-2670.12) (Table 1).

Spectrogram and oscillogram of two advertisement calls of the same male of Aplastodiscus albosignatus: (4) call with the dominant frequency in the first harmonic (= minimum frequency); (5) call with the dominant frequency in the third harmonic. Male recorded on January 14, 2020, voucher of specimen RLUTF 1220, voucher of recording FNJV 45563, 4.7 g, 44.8 mm, 18.1 °C.
Main acoustic parameters of the advertisement call of Aplastodiscus albosignatus in Mananciais da Serra, Paraná, Brazil. Legend: X = average; ± SD = standard deviation; Min = minimum recorded value; Max = maximum recorded value. All data refer to 10 analyzed calls of 22 males, totaling 220 analyzed calls.
According to the AIC criterion, the most explanatory model for the three spectral parameters (minimum frequency, maximum frequency and third harmonic frequency) was temperature plus body size, conditioning jointly these variables (Table 2). Concerning temporal variables, the best model that explains the variation in call duration was body size, while for the interval between calls the best model was the air temperature (Table 2).
The variation coefficient indicated that the minimum frequency has an intermediate variation. The maximum frequency and the frequency of the third harmonic were considered static parameters (Table 3). Regarding temporal parameters, the call duration was considered as a static intra-individual and dynamic inter-individual parameter, while the interval between calls was dynamic in both comparisons. The ratio between CVinter and CVintra of all the analyzed acoustic signals was greater than 1, indicating that there is greater variation between males than at the individual level (Table 3).
Most supported models (ΔAIC < 4) for each dependent variable considering two independent variables: temperature and body size. The values highlighted in bold represent the best model for the variable in question.
Variation of the main acoustic signals of the vocalizations of Aplastodiscus albosignatus in Mananciais da Serra, Paraná, Brazil. Legend: CVINTRA = intra-individual variation coefficient; CVINTER = interindividual variation coefficient.
DISCUSSION
The species belonging to the A. albosignatus group have non-pulsed calls consisting of a single note (Abrunhosa et al. 2005Abrunhosa PA, Pimenta BVS, Cruz CAG, Haddad CFB (2005) Advertisement Calls of Species of the Hyla Albosignata Group (Amphibia, Anura, Hylidae). Arquivos do Museu Nacional 63: 275-282.). A difference observed between the previous description of A. albosignatus and the present description is the dominant frequency since Abrunhosa et al. (2005Abrunhosa PA, Pimenta BVS, Cruz CAG, Haddad CFB (2005) Advertisement Calls of Species of the Hyla Albosignata Group (Amphibia, Anura, Hylidae). Arquivos do Museu Nacional 63: 275-282.) proposed that the frequency with the highest peak energy always occurs in the third harmonic, between 2.61 and 2.66 kHz. In the recordings analyzed for the population of Piraquara (PR), we observed that the dominant frequency can occur in either the third or first harmonic, between 0.4 and 0.6 kHz. This difference is likely to be related to the fact that Abrunhosa et al. (2005Abrunhosa PA, Pimenta BVS, Cruz CAG, Haddad CFB (2005) Advertisement Calls of Species of the Hyla Albosignata Group (Amphibia, Anura, Hylidae). Arquivos do Museu Nacional 63: 275-282.) analyzed only 10 calls of a single male, while we analyzed the calls of a larger number of males. Most of the analyzed calls had a dominant frequency in the third harmonic (66%) but, in some cases, the dominant frequency was in the first harmonic (34%). Sometimes, the same male had dominant frequencies in the third harmonic and, in another call, in the first harmonic. This pattern can also be seen in Aplastodiscus callipygius (Cruz & Peixoto, 1985) (Abrunhosa et al. 2005Abrunhosa PA, Pimenta BVS, Cruz CAG, Haddad CFB (2005) Advertisement Calls of Species of the Hyla Albosignata Group (Amphibia, Anura, Hylidae). Arquivos do Museu Nacional 63: 275-282.) and A. leucopygius (Zina and Haddad 2006Zina J, Haddad CFB (2006) Acoustic repertoire of Aplastodiscus arildae and A. leucopygius (Anura: Hylidae) in Serra do Japi, Brazil. South American Journal of Herpetology 1: 227-236. https://doi.org/10.2994/1808-9798(2006)1[227:AROAAA]2.0.CO;2
https://doi.org/10.2994/1808-9798(2006)1...
).
Aplastodiscus leucopygius has calls with three harmonics and the dominant frequency can be present in the first, second and third harmonics (Zina and Haddad 2006Zina J, Haddad CFB (2006) Acoustic repertoire of Aplastodiscus arildae and A. leucopygius (Anura: Hylidae) in Serra do Japi, Brazil. South American Journal of Herpetology 1: 227-236. https://doi.org/10.2994/1808-9798(2006)1[227:AROAAA]2.0.CO;2
https://doi.org/10.2994/1808-9798(2006)1...
). The authors observed on two occasions that, when the same male was recorded alone, he emitted the dominant frequency in the third harmonic, but, when recorded in antiphony with another male, he emitted the dominant frequency in the first harmonic (Zina and Haddad 2006Zina J, Haddad CFB (2006) Acoustic repertoire of Aplastodiscus arildae and A. leucopygius (Anura: Hylidae) in Serra do Japi, Brazil. South American Journal of Herpetology 1: 227-236. https://doi.org/10.2994/1808-9798(2006)1[227:AROAAA]2.0.CO;2
https://doi.org/10.2994/1808-9798(2006)1...
). The dominant frequency can transmit information about the male body size and, in most species, this acoustic signal is inversely proportional to the size of the calling male (Wagner 1989Wagner WE (1989) Graded aggressive signals in Blanchard’s cricket frog: vocal responses to opponent proximity and size. Animal Behaviour 38: 1025-1038. https://doi.org/10.1016/S0003-3472(89)80141-1
https://doi.org/10.1016/S0003-3472(89)80...
, Bee et al 1999Bee MA, Perrill SA, Owen PC (1999) Size assessment in simulated territorial encounters between male green frogs (Rana clamitans). Behavioral Ecology and Sociobiology 45: 177-184. https://doi.org/10.1007/s002650050551
https://doi.org/10.1007/s002650050551...
, Vilaça et al 2011Vilaça TRA, dos Santos Silva JR, Solé M (2011) Vocalization and territorial behaviour of Phyllomedusa nordestina Caramaschi, 2006 (Anura: Hylidae) from southern Bahia, Brazil. Journal of Natural History 45: 1823-1834. https://doi.org/10.1080/00222933.2011.561018
https://doi.org/10.1080/00222933.2011.56...
, Morais et al. 2012Morais AR, Batista VG, Gambale PG, Signorelli L, Bastos RP (2012) Acoustic communication in a Neotropical frog (Dendropsophus minutus): vocal repertoire, variability and individual discrimination. Herpetological Journal 22: 249-257.). Studies show that larger males vocalize at lower dominant frequencies (Guimarães and Bastos 2003Guimarães LD, Bastos RP (2003) Vocalizações e interações acústicas em Hyla raniceps (Anura, Hylidae) durante a atividade reprodutiva. Iheringia, Série Zoologia 93: 149-158. https://doi.org/10.1590/S0073-47212003000200005
https://doi.org/10.1590/S0073-4721200300...
, Toledo and Haddad 2009Toledo LF, Haddad CFB (2009) Defensive Vocalizations of Neotropical Anurans. South American Journal of Herpetology 4: 25-42. https://doi.org/10.2994/057.004.0104
https://doi.org/10.2994/057.004.0104...
, Morais et al. 2012Morais AR, Batista VG, Gambale PG, Signorelli L, Bastos RP (2012) Acoustic communication in a Neotropical frog (Dendropsophus minutus): vocal repertoire, variability and individual discrimination. Herpetological Journal 22: 249-257.) due to the morphological structure of the components of the larynx (Martin 1971Martin WF (1971) Mechanics of sound production in toads of the genus Bufo: Passive elements. Journal of Experimental Zoology 176: 273-293. https://doi.org/10.1002/jez.1401760304
https://doi.org/10.1002/jez.1401760304...
). The vibration of the vocal cords, which depends on the size of the individuals, mainly influences the minimum frequency of the calls (Giacoma et al. 1997Giacoma C, Zugolaro C, Beani L (1997) The Advertisement Calls of the Green Toad (Bufo viridis): Variability and Role in Mate Choice. Herpetologica 53: 454-464.). This correlation can provide information about the body size of the calling male to other competing males, which avoids physical combat (Bastos et al. 2011Bastos RP, Alcantara MB, Morais AR, Lingnau R, Signorelli L (2011) Vocal behaviour and conspecific call response in Scinax centralis. Herpetological Journal 21: 43-50.). Small males of Acris crepitans Baird, 1854, during an experiment with playback, decreased the dominant frequency of their calls (Wagner 1989Wagner WE (1989) Graded aggressive signals in Blanchard’s cricket frog: vocal responses to opponent proximity and size. Animal Behaviour 38: 1025-1038. https://doi.org/10.1016/S0003-3472(89)80141-1
https://doi.org/10.1016/S0003-3472(89)80...
). Wagner (1992Wagner WE (1992) Deceptive or honest signalling of fighting ability? A test of alternative hypotheses for the function of changes in call dominant frequency by male cricket frogs. Animal Behaviour 44: 449-462. https://doi.org/10.1016/0003-3472(92)90055-E
https://doi.org/10.1016/0003-3472(92)900...
) proposed the hypothesis that males can change the frequency of their calls so their opponents receive wrong information about the emitting male’s fighting ability. Subsequently, Bee et al. (2000Bee MA, Perrill SA, Owen PC (2000) Male green frogs lower the pitch of acoustic signals in defense of territories: A possible dishonest signal of size? Behavioral Ecology 11: 169-177. https://doi.org/10.1093/beheco/11.2.169
https://doi.org/10.1093/beheco/11.2.169...
) performed playback experiments with Rana clamitans Latreille in Sonnini de Manoncourt and Latreille, 1801 and their results support the theory of emission of dishonest signals by the frogs. Thus, the variation in the dominant frequency in the call of A. albosignatus may be socially dependent. However, experiments with playback are necessary to evaluate this interaction more precisely.
In general, the size of the males is considered a factor that influences mainly the spectral parameters of the call, such as dominant and minimum frequency (Toledo and Haddad 2009Toledo LF, Haddad CFB (2009) Defensive Vocalizations of Neotropical Anurans. South American Journal of Herpetology 4: 25-42. https://doi.org/10.2994/057.004.0104
https://doi.org/10.2994/057.004.0104...
, Vilaça et al. 2011Vilaça TRA, dos Santos Silva JR, Solé M (2011) Vocalization and territorial behaviour of Phyllomedusa nordestina Caramaschi, 2006 (Anura: Hylidae) from southern Bahia, Brazil. Journal of Natural History 45: 1823-1834. https://doi.org/10.1080/00222933.2011.561018
https://doi.org/10.1080/00222933.2011.56...
, Turin et al. 2018Turin RAF, Nali RC, Prado CPA (2018) Intraspecific call variation in a Neotropical gladiator frog with a complex advertisement call. Amphibia Reptilia 39: 31-39. https://doi.org/10.1163/15685381-00003147
https://doi.org/10.1163/15685381-0000314...
). It is possible to observe a pattern in which smaller individuals present higher frequencies. Our result may indicate that heavier males invest more energy to emit the advertisement call since the energy costs also depend on the size of the individuals (Gerhardt 1994Gerhardt HC (1994) The Evolution of Vocalization in Frogs and Toads Author. Annual Review of Ecology and Systematics 25: 293-324., Turin et al. 2018Turin RAF, Nali RC, Prado CPA (2018) Intraspecific call variation in a Neotropical gladiator frog with a complex advertisement call. Amphibia Reptilia 39: 31-39. https://doi.org/10.1163/15685381-00003147
https://doi.org/10.1163/15685381-0000314...
). However, in the present study, the variation in spectral parameters was better explained by the body size plus temperature model. This is an interesting result, as air temperature tends to influence the temporal acoustic parameters, while spectral parameters are considered mostly thermally independent among frogs (Navas and Bevier 2001Navas CA, Bevier CR (2001) Thermal dependency of calling performance in the eurythermic frog Colostethus subpunctatus. Herpetologica 57: 384-395.). Nevertheless, there are reports of some contrasting results, of temperature influencing spectral parameters (Ziegler et al. 2016Ziegler L, Arim M, Bozinovic F (2016). Intraspecific scaling in frog calls: the interplay of temperature, body size and metabolic condition. Oecologia 181: 673-681. https://doi.org/10.1007/s00442-015-3499-8
https://doi.org/10.1007/s00442-015-3499-...
and references therein), indicating that these patterns are still not fully understood.
Regarding temporal acoustic parameters, the interval between calls was influenced mainly by temperature, as expected according to the literature. The influence of temperature on temporal parameters has been recorded in many anuran species (Lingnau and Bastos 2007Lingnau R, Bastos RP (2007) Vocalizations of the Brazilian torrent frog Hylodes heyeri (Anura: Hylodidae): Repertoire and influence of air temperature on advertisement call variation. Journal of Natural History 41: 17-20. https://doi.org/10.1080/00222930701395626
https://doi.org/10.1080/0022293070139562...
, Morais et al. 2012Morais AR, Batista VG, Gambale PG, Signorelli L, Bastos RP (2012) Acoustic communication in a Neotropical frog (Dendropsophus minutus): vocal repertoire, variability and individual discrimination. Herpetological Journal 22: 249-257., Boschetti et al. 2019Boschetti JP, De Bastiani VIM, Lingnau R, Lucas EM (2019) Bioacoustics of Pithecopus rusticus (Anura, Phyllomedusidae): a rare species possibly threatened with extinction. South American Journal of Herpetology 14: 196-203. https://doi.org/10.2994/SAJH-D-17-00071.1
https://doi.org/10.2994/SAJH-D-17-00071....
). Temperature has a great influence on the metabolism of amphibians, mainly in acoustic signals affected by active muscle contraction, which reflects in the amount of energy spent in the emission of the calls (Navas and Bevier 2001Navas CA, Bevier CR (2001) Thermal dependency of calling performance in the eurythermic frog Colostethus subpunctatus. Herpetologica 57: 384-395., Wells 2010Wells KD (2010) The ecology and behavior of amphibians. Chicago, University of Chicago Press, 1148 pp.). Regarding call duration, the model that best explained the variation was body size. We can interpret this result as meaning that investing in longer calls can increase male reproductive success through sexual selection. Another possible explanation is that larger males can store larger volumes of air in their lungs, producing positive correlations with temporal parameters (Bernal et al. 2005Bernal XE, Guarnizo CE, Luddecke H (2005) Geographic variation in advertisement call and genetic structure of Colostethus palmatus (Anura, Dendrobatidae) from the Colombian Andes Herpetologists’ League. Herpetologica 61: 395-408. https://doi.org/10.1655/04-87.1
https://doi.org/10.1655/04-87.1...
).
The maximum frequency and the peak frequency of the third harmonic were acoustic signals considered static, that is, they have little variability and mainly affect the quality of the call (Gerhardt 1991Gerhardt HC (1991) Female mate choice in treefrogs: static and dynamic acoustic criteria. Animal Behaviour 42: 615-635. https://doi.org/10.1016/S0003-3472(05)80245-3
https://doi.org/10.1016/S0003-3472(05)80...
). Static acoustic signals are important for species recognition since they tend to remain constant among males (Gerhardt 1991Gerhardt HC (1991) Female mate choice in treefrogs: static and dynamic acoustic criteria. Animal Behaviour 42: 615-635. https://doi.org/10.1016/S0003-3472(05)80245-3
https://doi.org/10.1016/S0003-3472(05)80...
, Márquez and Eekhout 2006Márquez R, Eekhout X (2006) Advertisement calls of six species of anurans from Bali, Republic of Indonesia. Journal of Natural History 40: 9-10. https://doi.org/10.1080/00222930600712129
https://doi.org/10.1080/0022293060071212...
). Temporal parameters are usually dynamic factors in relation to intra-individual variation (Tárano 2001Tárano Z (2001) Variation in Male Advertisement Calls in the Neotropical Frog Physalaemus enesefae. Copeia 2001: 1064-1072. https://doi.org/10.1643/0045-8511(2001)001[1064:VIMACI]2.0.CO;2). In this study, the call duration and the interval between calls were considered dynamic parameters. Characteristics of the call with wide variation, which are considered dynamic, are important in sexual selection (Gerhardt 1991Gerhardt HC (1991) Female mate choice in treefrogs: static and dynamic acoustic criteria. Animal Behaviour 42: 615-635. https://doi.org/10.1016/S0003-3472(05)80245-3
https://doi.org/10.1016/S0003-3472(05)80...
). Temporal parameters, in general, are more variable than spectral parameters because they tend to be more influenced by changes in ambient temperature and social conditions (Gerhardt and Huber 2002Gerhardt HC, Huber F (2002) Acoustic Communication in Insects and Anurans: Common Problems and Diverse Solutions. Chicago, The University of Chicago Press, 542 pp., Wong et al. 2004Wong BBM, Cowling ANN, Cunningham RB, Donnelly CF, Cooper PD (2004) Do temperature and social environment interact to affect call rate in frogs (Crinia signifera)? Austral Ecology 29: 209-214. https://doi.org/10.1111/j.1442-9993.2004.01338.x
https://doi.org/10.1111/j.1442-9993.2004...
).
The ratio between CVinter and CVintra indicated that there is greater variation between males than the individual variation itself, as found in other species (Morais et al. 2012Morais AR, Batista VG, Gambale PG, Signorelli L, Bastos RP (2012) Acoustic communication in a Neotropical frog (Dendropsophus minutus): vocal repertoire, variability and individual discrimination. Herpetological Journal 22: 249-257., Forti et al. 2017Forti LR, Lingnau R, Bertoluci J (2017) Acoustic variation in the advertisement call of the Lime treefrog Sphaenorhynchus caramaschii (Anura: Hylidae). Vertebrate Zoology 67: 197-205.). There are several possible explanations for the variation between males, such as genetic variation, variation in morphology and physiology, and directed sexual selection (Morais et al. 2012Morais AR, Batista VG, Gambale PG, Signorelli L, Bastos RP (2012) Acoustic communication in a Neotropical frog (Dendropsophus minutus): vocal repertoire, variability and individual discrimination. Herpetological Journal 22: 249-257.). The frequency of the third harmonic and call duration were the acoustic signals with the highest value of the CVinter/CVintra ratio among the analyzed parameters. This may suggest a potential for male identification by females and competing males (Morais et al. 2012Morais AR, Batista VG, Gambale PG, Signorelli L, Bastos RP (2012) Acoustic communication in a Neotropical frog (Dendropsophus minutus): vocal repertoire, variability and individual discrimination. Herpetological Journal 22: 249-257., Turin et al. 2018Turin RAF, Nali RC, Prado CPA (2018) Intraspecific call variation in a Neotropical gladiator frog with a complex advertisement call. Amphibia Reptilia 39: 31-39. https://doi.org/10.1163/15685381-00003147
https://doi.org/10.1163/15685381-0000314...
). Temporal parameters, such as call duration, have already been considered important in individual recognition (Gambale et al. 2014Gambale P, Signorelli L, Bastos R (2014) Individual variation in the advertisement calls of a Neotropical treefrog (Scinax constrictus). Amphibia-Reptilia 35: 271-281. https://doi.org/10.1163/15685381-00002949
https://doi.org/10.1163/15685381-0000294...
, Gasser et al. 2009Gasser H, Amézquita A, Hödl W (2009) Who is Calling? Intraspecific Call variation in the aromobatid frog Allobates femoralis. Ethology 115: 596-607. https://doi.org /10.1111/j.1439-0310.2009.01639.x
https://doi.org /10.1111/j.1439-0310.200...
). Since both parameters are correlated with male size, it is plausible to consider that females can identify larger males from these call features (Turin et al. 2018Turin RAF, Nali RC, Prado CPA (2018) Intraspecific call variation in a Neotropical gladiator frog with a complex advertisement call. Amphibia Reptilia 39: 31-39. https://doi.org/10.1163/15685381-00003147
https://doi.org/10.1163/15685381-0000314...
).
With this work, we have extended the description of the advertisement call of A. albosignatus, providing information based on a larger number of males and calls than previously reported in the literature. For Aplastodiscus, a similar number of call analyses is available only for A. arildae and A. leucopygius (Zina and Haddad 2006Zina J, Haddad CFB (2006) Acoustic repertoire of Aplastodiscus arildae and A. leucopygius (Anura: Hylidae) in Serra do Japi, Brazil. South American Journal of Herpetology 1: 227-236. https://doi.org/10.2994/1808-9798(2006)1[227:AROAAA]2.0.CO;2
https://doi.org/10.2994/1808-9798(2006)1...
). Increased knowledge about the variation in advertisement calls contributes to a finer taxonomic delimitation and can provide a basis for hypotheses about the change in acoustic features over time (Castellano et al. 2002Castellano S, Cuatto B, Rinella R, Rosso A (2002) The advertisement call of the European treefrogs, Hyla arborea: a multi-level study of variation. Ethology 108: 75-89. https://doi.org/10.1046/j.1439-0310.2002.00761.x
https://doi.org/10.1046/j.1439-0310.2002...
). Future efforts should try to obtain recordings of A. albosignatus near its type locality, in Paranapiacaba (=Alto da Serra), São Paulo (Werner C. Bokermann, unpublished data), for a better definition of the species’ advertisement calls due to the discussions of Berneck et al. (2016Berneck BVM, Haddad CFB, Lyra ML, Cruz CAG, Faivovich J (2016) The Green Clade grows: A phylogenetic analysis of Aplastodiscus (Anura; Hylidae). Molecular Phylogenetics and Evolution 97: 213-223. https://doi.org/10.1016/j.ympev.2015.11.014
https://doi.org/10.1016/j.ympev.2015.11....
) of a possible new species under this name. Since we notice that both temperature and body size are factors that influence the calls of this species, this should also be taken into strong consideration when comparing the calls of species in the genus Aplastodiscus.
ACKNOWLEDGMENTS
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brasil, Finance Code 001, and by Fundação Araucária and Conselho Nacional de Desenvolvimento Científico e Tecnológico (421298/2017-0). Field work at the study site was authorized by the Environmental Institute of Paraná (IAP). We want to thank the Associate Editor Fabricius Domingos for the kind review and accessibility in answering our questions. We would also like to thank the biologist and translator Piter Keo for reviewing the text.
LITERATURE CITED
- Arnold TW (2010) Uninformative parameters and model selection using Akaike’s Information Criterion. The Journal of Wildlife Management 74: 1175-1178. https://doi.org/10.2193/2009-367
» https://doi.org/10.2193/2009-367 - Abrunhosa PA, Pimenta BVS, Cruz CAG, Haddad CFB (2005) Advertisement Calls of Species of the Hyla Albosignata Group (Amphibia, Anura, Hylidae). Arquivos do Museu Nacional 63: 275-282.
- Baêta D, Lourenço ACC, Pezzuti TL, Pires MRS (2007) The tadpole, advertisement call, and geographic distribution of Physalaemus maximus Feio, Pombal & Caramaschi, 1999 (Amphibia, Anura, Leiuperidae). Arquivos do Museu Nacional 65: 27-32.
- Bastos RP, Alcantara MB, Morais AR, Lingnau R, Signorelli L (2011) Vocal behaviour and conspecific call response in Scinax centralis Herpetological Journal 21: 43-50.
- Bee MA, Perrill SA, Owen PC (1999) Size assessment in simulated territorial encounters between male green frogs (Rana clamitans). Behavioral Ecology and Sociobiology 45: 177-184. https://doi.org/10.1007/s002650050551
» https://doi.org/10.1007/s002650050551 - Bee MA, Perrill SA, Owen PC (2000) Male green frogs lower the pitch of acoustic signals in defense of territories: A possible dishonest signal of size? Behavioral Ecology 11: 169-177. https://doi.org/10.1093/beheco/11.2.169
» https://doi.org/10.1093/beheco/11.2.169 - Bee MA, Reichert MS, Tumulty J (2016) Assessment and recognition of rivals in anuran contests. Advances in the Study of Behavior 48: 161-249.
- Bee MA, Suyesh R, Biju SD (2013) The vocal repertoire of Pseudophilautus kani, a shrub frog (Anura: Rhacophoridae) from the Western Ghats of India. Bioacoustics 22: 67-85. https://doi.org/10.1080/09524622.2012.712750
» https://doi.org/10.1080/09524622.2012.712750 - Bernal XE, Guarnizo CE, Luddecke H (2005) Geographic variation in advertisement call and genetic structure of Colostethus palmatus (Anura, Dendrobatidae) from the Colombian Andes Herpetologists’ League. Herpetologica 61: 395-408. https://doi.org/10.1655/04-87.1
» https://doi.org/10.1655/04-87.1 - Berneck BVM, Haddad CFB, Lyra ML, Cruz CAG, Faivovich J (2016) The Green Clade grows: A phylogenetic analysis of Aplastodiscus (Anura; Hylidae). Molecular Phylogenetics and Evolution 97: 213-223. https://doi.org/10.1016/j.ympev.2015.11.014
» https://doi.org/10.1016/j.ympev.2015.11.014 - Bezerra AM, Passos LO, De Luna-Dias C, Quintanilha AS, De Carvalho-e-Silva SP (2020) A missing piece of the puzzle: Re-encounter of Aplastodiscus musicus, its call, and phylogenetic placement (Anura: Hylidae: Cophomantini). Herpetologica 76: 74-82. https://doi.org/10.1655/Herpetologica-D-18-00061
» https://doi.org/10.1655/Herpetologica-D-18-00061 - Boschetti JP, De Bastiani VIM, Lingnau R, Lucas EM (2019) Bioacoustics of Pithecopus rusticus (Anura, Phyllomedusidae): a rare species possibly threatened with extinction. South American Journal of Herpetology 14: 196-203. https://doi.org/10.2994/SAJH-D-17-00071.1
» https://doi.org/10.2994/SAJH-D-17-00071.1 - Castellano S, Cuatto B, Rinella R, Rosso A (2002) The advertisement call of the European treefrogs, Hyla arborea: a multi-level study of variation. Ethology 108: 75-89. https://doi.org/10.1046/j.1439-0310.2002.00761.x
» https://doi.org/10.1046/j.1439-0310.2002.00761.x - Ferreira JCV (1996) O Paraná e seus municípios. Curitiba, Editora Memória Brasileira, 728 pp.
- Forti LR, Lingnau R, Bertoluci J (2017) Acoustic variation in the advertisement call of the Lime treefrog Sphaenorhynchus caramaschii (Anura: Hylidae). Vertebrate Zoology 67: 197-205.
- Frost DR (2021) Amphibian Species of the World: an Online Reference. Version 6.1 (29/06/2021). Available online at Available online at https://amphibiansoftheworld.amnh.org/index.php
» https://amphibiansoftheworld.amnh.org/index.php - Gambale PG, Bastos RP (2014) Vocal repertoire and bioacoustic analyses in Physalaemus cuvieri (Anura, Leptodactylidae) from southern Brazil. Herpetological Journal 24: 31-39.
- Gambale P, Signorelli L, Bastos R (2014) Individual variation in the advertisement calls of a Neotropical treefrog (Scinax constrictus). Amphibia-Reptilia 35: 271-281. https://doi.org/10.1163/15685381-00002949
» https://doi.org/10.1163/15685381-00002949 - Gasser H, Amézquita A, Hödl W (2009) Who is Calling? Intraspecific Call variation in the aromobatid frog Allobates femoralis Ethology 115: 596-607. https://doi.org /10.1111/j.1439-0310.2009.01639.x
» https://doi.org /10.1111/j.1439-0310.2009.01639.x - Gerhardt HC (1991) Female mate choice in treefrogs: static and dynamic acoustic criteria. Animal Behaviour 42: 615-635. https://doi.org/10.1016/S0003-3472(05)80245-3
» https://doi.org/10.1016/S0003-3472(05)80245-3 - Gerhardt HC (1994) The Evolution of Vocalization in Frogs and Toads Author. Annual Review of Ecology and Systematics 25: 293-324.
- Gerhardt HC, Huber F (2002) Acoustic Communication in Insects and Anurans: Common Problems and Diverse Solutions. Chicago, The University of Chicago Press, 542 pp.
- Giacoma C, Zugolaro C, Beani L (1997) The Advertisement Calls of the Green Toad (Bufo viridis): Variability and Role in Mate Choice. Herpetologica 53: 454-464.
- Guerra V, Llusia D, Gambale PG, Morais AR, Marquez R, Bastos RP (2018) The advertisement calls of Brazilian anurans: Historical review, current knowledge and future directions. PLoS ONE 13: 1-22. https://doi.org/10.1371/journal.pone.0191691
» https://doi.org/10.1371/journal.pone.0191691 - Guimarães LD, Bastos RP (2003) Vocalizações e interações acústicas em Hyla raniceps (Anura, Hylidae) durante a atividade reprodutiva. Iheringia, Série Zoologia 93: 149-158. https://doi.org/10.1590/S0073-47212003000200005
» https://doi.org/10.1590/S0073-47212003000200005 - Hepp F, Pombal J Jr (2020) Review of bioacoustical traits in the genus Physalaemus Fitzinger, 1826 (Anura: Leptodactylidae: Leiuperinae). Zootaxa 4725(1): 1-106. https://doi.org/10.11646/zootaxa.4725.1.1
» https://doi.org/10.11646/zootaxa.4725.1.1 - Köhler J, Jansen M, Rodríguez A, Kok PJR, Toledo LF, Emmrich M, Glaw F, Haddad CFB, Rödel MO, Vences M (2017) The use of bioacoustics in anuran taxonomy: Theory, terminology, methods and recommendations for best practice. Zootaxa 4251: 001-124. https://doi.org/10.11646/zootaxa.4251.1.1
» https://doi.org/10.11646/zootaxa.4251.1.1 - Lemes P, Tessarolo G, Morais AR, Bastos RP (2012) Acoustic Repertoire of Barycholos ternetzi (Anura: Strabomantidae) in Central Brazil. South American Journal of Herpetology 7: 157-164. https://doi.org/10.2994/057.007.0205
» https://doi.org/10.2994/057.007.0205 - Ligges U, Krey S, Mersmann O, Schnackenberg S (2013) Tuner: Analysis of music. Available online at Available online at http://r-forge.r-project.org/projects/tuner [Accessed 15/06/2020]
» http://r-forge.r-project.org/projects/tuner - Lingnau R, Bastos RP (2007) Vocalizations of the Brazilian torrent frog Hylodes heyeri (Anura: Hylodidae): Repertoire and influence of air temperature on advertisement call variation. Journal of Natural History 41: 17-20. https://doi.org/10.1080/00222930701395626
» https://doi.org/10.1080/00222930701395626 - Márquez R, Eekhout X (2006) Advertisement calls of six species of anurans from Bali, Republic of Indonesia. Journal of Natural History 40: 9-10. https://doi.org/10.1080/00222930600712129
» https://doi.org/10.1080/00222930600712129 - Martin WF (1971) Mechanics of sound production in toads of the genus Bufo: Passive elements. Journal of Experimental Zoology 176: 273-293. https://doi.org/10.1002/jez.1401760304
» https://doi.org/10.1002/jez.1401760304 - Morais AR, Batista VG, Gambale PG, Signorelli L, Bastos RP (2012) Acoustic communication in a Neotropical frog (Dendropsophus minutus): vocal repertoire, variability and individual discrimination. Herpetological Journal 22: 249-257.
- Nali RC, Prado CP (2014) Complex call with different messages in Bokermannohyla ibitiguara (Anura, Hylidae), a gladiator frog of the Brazilian Cerrado. Journal of Herpetology 48: 407-414.
- Navas CA, Bevier CR (2001) Thermal dependency of calling performance in the eurythermic frog Colostethus subpunctatus Herpetologica 57: 384-395.
- Paraná (1987) Atlas do estado do Paraná. Curitiba, Secretaria do Estado da Agricultura e do Abastecimento, Intitutos de Terras, Cartografias e Florestas, 486 pp.
- Pinheiro PD, Pezzuti TL, Berneck BV, Lyra ML, Lima RC, Leite FS (2021) A new cryptic species of the Aplastodiscus albosignatus group (Anura: Hylidae). Salamandra 57: 27-43.
- R Development Core Team (2016) R: A language and environment for statistical computing. Version 3.3.2, available online at http://www.R-project.org
» http://www.R-project.org - Reginato M, Goldenberg R (2007) Análise florística, estrutural e fitogeográfica da vegetação em região de transição entre as Florestas Ombrófilas Mista e Densa Montana, Piraquara, Paraná, Brasil. Hoehnea 34: 349-360. https://doi.org/10.1590/S2236-89062007000300006
» https://doi.org/10.1590/S2236-89062007000300006 - Robertson GM (1984) Acoustic Spacing by Breeding Males of Uperoleia rugosa (Anura: Leptodactylidae). Zeitschrift für Tierpsychologie 297: 283-297. https://doi.org/10.1111/j.1439-0310.1984.tb00364.x
» https://doi.org/10.1111/j.1439-0310.1984.tb00364.x - Signorell A, Aho K, Alfons A, Anderegg N, Aragon T (2016) DescTools: Tools for descriptive statistics. R package, v. 0.99.18, Vienna, R Foundation for Statistical Computing. https://cran.rproject.org/web/packages/DescTools/index.html
» https://cran.rproject.org/web/packages/DescTools/index.html - Sueur J, Aubin T, Simonis C (2008) Seewave: a free modular tool for sound analysis and synthesis. Bioacoustics 18: 213-226. https://doi.org/10.1080/09524622.2008.9753600
» https://doi.org/10.1080/09524622.2008.9753600 - Tárano Z (2001) Variation in Male Advertisement Calls in the Neotropical Frog Physalaemus enesefae Copeia 2001: 1064-1072. https://doi.org/10.1643/0045-8511(2001)001[1064:VIMACI]2.0.CO;2
- Toledo LF, Haddad CFB (2009) Defensive Vocalizations of Neotropical Anurans. South American Journal of Herpetology 4: 25-42. https://doi.org/10.2994/057.004.0104
» https://doi.org/10.2994/057.004.0104 - Toledo LF, Martins IA, Bruschi DP, Passos MA, Alexandre C, Haddad CFB (2014) The anuran calling repertoire in the light of social context. Acta Ethologica 18: 87-99. https://doi.org/10.1007/s10211-014-0194-4
» https://doi.org/10.1007/s10211-014-0194-4 - Turin RAF, Nali RC, Prado CPA (2018) Intraspecific call variation in a Neotropical gladiator frog with a complex advertisement call. Amphibia Reptilia 39: 31-39. https://doi.org/10.1163/15685381-00003147
» https://doi.org/10.1163/15685381-00003147 - Vilaça TRA, dos Santos Silva JR, Solé M (2011) Vocalization and territorial behaviour of Phyllomedusa nordestina Caramaschi, 2006 (Anura: Hylidae) from southern Bahia, Brazil. Journal of Natural History 45: 1823-1834. https://doi.org/10.1080/00222933.2011.561018
» https://doi.org/10.1080/00222933.2011.561018 - Wagner WE (1989) Graded aggressive signals in Blanchard’s cricket frog: vocal responses to opponent proximity and size. Animal Behaviour 38: 1025-1038. https://doi.org/10.1016/S0003-3472(89)80141-1
» https://doi.org/10.1016/S0003-3472(89)80141-1 - Wagner WE (1992) Deceptive or honest signalling of fighting ability? A test of alternative hypotheses for the function of changes in call dominant frequency by male cricket frogs. Animal Behaviour 44: 449-462. https://doi.org/10.1016/0003-3472(92)90055-E
» https://doi.org/10.1016/0003-3472(92)90055-E - Wei T, Simko V, Levy M, Xie Y, Jin Y, Zemla J (2017). Package ‘corrplot’. Statistician 56: e24. https://cran.r-project.org/web/packages/corrplot/corrplot.pdf
» https://cran.r-project.org/web/packages/corrplot/corrplot.pdf - Wells KD (2010) The ecology and behavior of amphibians. Chicago, University of Chicago Press, 1148 pp.
- Wong BBM, Cowling ANN, Cunningham RB, Donnelly CF, Cooper PD (2004) Do temperature and social environment interact to affect call rate in frogs (Crinia signifera)? Austral Ecology 29: 209-214. https://doi.org/10.1111/j.1442-9993.2004.01338.x
» https://doi.org/10.1111/j.1442-9993.2004.01338.x - Wuertz D, Wuertz MD, Team RC (2007) The fBasics Package. https://cran.r-project.org/web/packages/fBasics/index.html
» https://cran.r-project.org/web/packages/fBasics/index.html - Ziegler L, Arim M, Bozinovic F (2016). Intraspecific scaling in frog calls: the interplay of temperature, body size and metabolic condition. Oecologia 181: 673-681. https://doi.org/10.1007/s00442-015-3499-8
» https://doi.org/10.1007/s00442-015-3499-8 - Zina J, Haddad CFB (2006) Acoustic repertoire of Aplastodiscus arildae and A. leucopygius (Anura: Hylidae) in Serra do Japi, Brazil. South American Journal of Herpetology 1: 227-236. https://doi.org/10.2994/1808-9798(2006)1[227:AROAAA]2.0.CO;2
» https://doi.org/10.2994/1808-9798(2006)1[227:AROAAA]2.0.CO;2
ADDITIONAL NOTES
-
Zoobank register
http://zoobank.org/730D30AE-4DA7-4733-9B9D-4CCD87BDF4A6 -
How to cite this article
Moser CF, Schuck LK, Olmedo GM, Lingnau R (2022) Individual variation in the advertisement call of Aplastodiscus albosignatus (Anura: Hylidae) is correlated with body size and environmental temperature. Zoologia (Curitiba) 39: e21008. https://doi.org/10.1590/S1984-4689.v39.e21008 -
Published by
Sociedade Brasileira de Zoologia at Scientific Electronic Library Online (https://www.scielo.br/zool)
Edited by
Editorial responsibility
Data availability
Data citations
Frost DR (2021) Amphibian Species of the World: an Online Reference. Version 6.1 (29/06/2021). Available online at Available online at https://amphibiansoftheworld.amnh.org/index.php
Publication Dates
-
Publication in this collection
21 Mar 2022 -
Date of issue
2022
History
-
Received
29 June 2021 -
Accepted
03 Jan 2022