Abstract
Cryptocellus canga sp. nov. is described from specimens collected in several caves at Carajás National Forest, Pará, Brazil. The new species differs from other species of the genus by the morphology of copulatory apparatus of the male leg III.
Brazilian Amazon; canga; Carajás, cave; Neotropics; taxonomy
TAXONOMY AND NOMENCLATURE
A new species of Cryptocellus (Arachnida: Ricinulei) from Eastern Amazonia
Ricardo Pinto-da-RochaI; Renata AndradeII
IDepartamento de Zoologia, Instituto de Biociências, Universidade de São Paulo. Rua do Matão, Travessa 14, 321, 05508-900 São Paulo, SP, Brazil
IIRua Paulo Orozimbo, 530, ap. 92B, 01535-000 São Paulo, SP, Brazil
ABSTRACT
Cryptocellus canga sp. nov. is described from specimens collected in several caves at Carajás National Forest, Pará, Brazil. The new species differs from other species of the genus by the morphology of copulatory apparatus of the male leg III.
Key words: Brazilian Amazon; canga; Carajás, cave; Neotropics; taxonomy.
Arachnids of the order Ricinulei occur in tropical forests and caves of New World and Africa, with 72 living species placed in the family Ricinoididae (HARVEY 2003, BONALDO & PINTO-DA-ROCHA 2003, COKENDOLPHER & ENRIQUEZ 2004, PINTO-DA-ROCHA & BONALDO 2007, TOURINHO & AZEVEDO 2007, BOTERO-TRUJILLO & PEREZ 2008, 2009, NASKRECKI 2008, PLATNICK & GARCIA 2008, TERUEL & ARMAS 2008, TOURINHO & SATURNINO 2010, TOURINHO et al. 2010, VALDEZ-MONDRAGÓN & FRANCKE 2011). Of the three genera included in this family, Cryptocellus is the most diversified genus with 36 species distributed from Central America to northern South America. Nine species of Cryptocellus occur in the Brazilian Amazon: Cryptocellus foedus Westwood, 1874, C. simonis Hansen and Sorensen, 1904, C. becki Platnick and Shadab, 1977, C. adisi Platnick, 1988, C. abaporu Bonaldo & Pinto-da-Rocha, 2003, C. tarsilae Pinto-da-Rocha & Bonaldo, 2007, C. icamiabas Tourinho & Azevedo, 2007, C. conori Tourinho & Saturnino, 2010 and C. iaci Tourinho et al., 2010.
In this paper we describe a new species of Cryptocellus from the Carajás National Forest, the second species found in caves of this region, besides C. tarsilae. The specimens available were collected in superficial caves formed in iron ore in areas of residual plateau, more specifically on the bases of outcrops of iron ore or 'canga'. The 'canga' is usually covered by open vegetation type called 'metalophylic vegetation', which is characterized by plants able to grow in soils rich in iron and other heavy metals. The new species described here does not show any troglomorphism.
MATERIAL AND METHODS
The specimens were collected by visual search and manual capture. The general terminology follows PLATNICK & SHADAB (1977), while that of male leg III follows COKENDOLPHER (2000). Measurements were taken according to COOKE & SHADAB (1973) and are given in millimetres (Tab. I). The specimens were covered with clay, and some setae had clay on the apex, giving them the false appearance of being clavate. They were partially cleaned by immersion in water with detergent and exposed to an ultrasound cleaner for about 15 minutes. The type material is deposited in the Museu de Zoologia da Universidade de São Paulo (MZSP, curator: R. Pinto-da-Rocha), Museu Paraense Emilio Goeldi (MPEG, curator: A.B. Bonaldo) and Instituto Butantan (IBSP, curator: D.M.B. Battesti).
Cryptocellus canga sp. nov.Figs 1-11

Types. Male holotype (MZSP 46554) and female paratype (MZSP 46555), BRAZIL, Pará: Flona Carajás (Gruta S11D-12, 6º23'45"S, 50º21'34'W), 13-30.I.2010, R. Andrade leg. Paratypes: same data for holotype, 2 males (MZSP 46556) and 1 female (MZSP 46557); (Gruta Cav19 S11, 6º24'20"S, 50º22'07'W), 2231.V.2010, 1 male (MZSP 46558); (Gruta Cav35 S11, 6º24'22"S, 50º23'06'W), 22-28.IX.2010, 1 female (MZSP 46559); (Gruta S11D-13; 6º24'06"S, 50º21'08'W), 1-14.VII.2010, 1 female (MZSP 46560); (Gruta S11D-24, 6º24'26"S, 50º21'14'W), 3-19.VIII.2010, 1 male (MZSP 46561); (Gruta S11D-37; 6º24'46"S, 50º21'31'W), 19-22.II.2010, 1 female (MZSP 46562); (Gruta S11D-40, 6º24'37"S, 50º19'29'W), 3-19.VIII.2010, 5 males and 1 female (MZSP 46563); (Gruta S11D-41, 6º24'38"S, 50º19'29'W), 3-19.VIII.2010, 1 female (MZSP 46564), 13-30.I.2010, 1 female (MZSP 46565); (Gruta S11D-80, 6º23'33"S, 50º18'57'W), 13-30.I.2010, 1 female (MZSP 46566); (Gruta S11A-05), 23.VIII-2.IX.2007, 2 males and 1 female (MPEG); (Gruta S11B-13), 23.VIII-2.IX.2007, 1 female (IBSP); (Gruta S11C-27), 23.VIII-2.IX.2007, 1 male (IBSP), all collected by R. Andrade.
Etymology. A lateritic hard crust of iron hydroxide with or without ore fragments, called "canga" develops covering the iron formations and high-grade ore bodies. The term "canga" was first used by local miners and originates from "Itapanhoacanga", Indian term, which means stone of a black man"s head
(ESCHWEGE 1822).
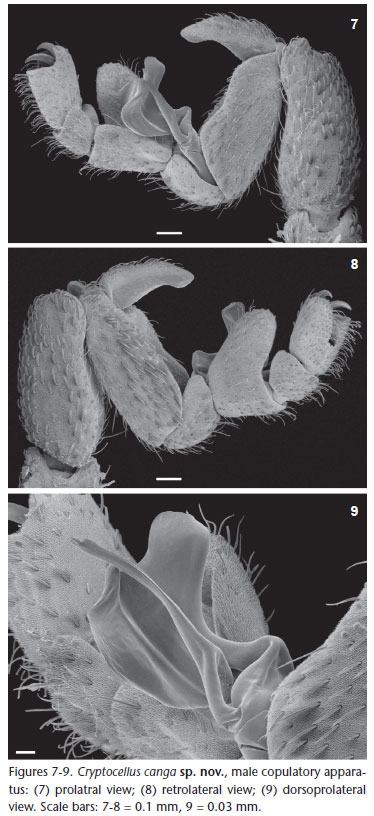
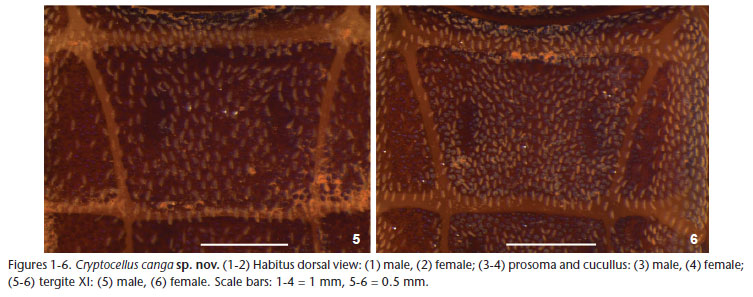
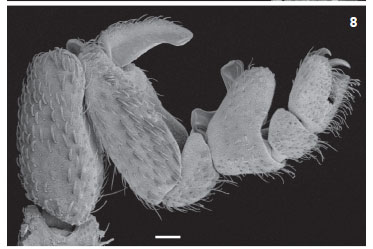
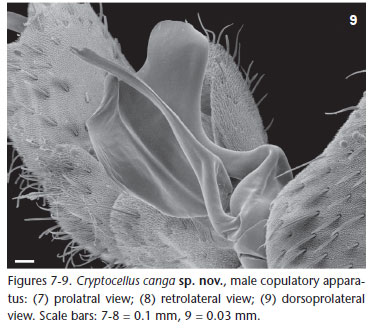
Diagnosis. C. canga sp. nov. differs from other species of the genus by the follow combination of characters: Body and legs with navicular setae (Figs 5 and 6); carapace without deep pits (Figs 3 and 4); copulatory apparatus with movable process with single apex and fixed process twisted as a spoon with two
apical, pronounced rounded lobes (Figs 7-9).
Description. Male (Holotype, MZSP 46554, Figs 1, 3, 5, and 7-9). Total length 5.46. Carapace 3.3 (including cucullus) long, 1.58 wide at leg III; general color brownish red, darker on margins, especially on posterior corner of carapace with small short white scale setae; with rounded eminence near posterior margin; anterior margin without a "U" notch; tubercles uniformly distributed, except near anterior margin where they are sparse (Fig. 3). Abdomen (Fig. 1) 3.0 long, 2.09 wide at tergite XII; color as in carapace; tergites with small short white setae densely covering dorsum; concentration of tubercles similar on tergites XI to XIII; median plate of tergite XI (Fig. 5) without wide central eminence; median plate of tergites XI-XIII uniformly colored (Fig. 1); median plate of tergites XI-XIII without median eminence decreasing posteriorly; median plate of tergites XI-XIII slightly wider than long; lateral margins of lateral plates of tergites lighter. Venter: sternal region with coxa I not meeting tritosternum; coxa II meeting along their entire length, their suture line about one-third longer than that of coxa III; coxa IV not touching it; coxae I-II densely tuberculated; III-IV with few scattered tubercles on anterior and posterior margins; sternites without tubercles, with small short thin white setae, except medially where they are more scattered. Pygidium with shallow notch on dorsal margin of basal segment (Fig. 1). Cucullus 0.92 long, 1.17 wide; entirely dark red, densely tuberculate on anterior half, with short white setae, longer setae scattered. Chelicera: fixed finger with three teeth (distal longer than others); movable finger with five equal-sized teeth. Pedipalp: covered with small short white scale setae; coxa, femur and tibia yellow, trochanter darker, posterior margin reddish; femur without tubercles. Leg formula II- III- IV- I; legs red, I-II darker, basitarsus and telotarsus lighter, densely covered with short white scale setae up to tibiae; basitarsus of legs I-II densely covered by tubercles on dorsal region; of leg III with few tubercles on dorsobasal half, basitarsal process with dorsal tubercles; tarsal claws thin, evenly curved; copulatory apparatus as on figures 7-9, fixed piece thin, wider and with bilobate apex, movable piece inserted on lateral of fixed piece. Legs with following sexual modifications: second leg not noticeable widened; femur I 2.8 times as long as wide; II 3.4 times longer than wide; tibia I without knob on middle of prolateral side of venter; trochanters without any remarkable features; leg III as in figures 7-9.
Female (Paratype, MZSP 46555, Figs 2, 4, 6, 10, and 11). Similar to male, except as follows. Total length 5.1. Carapace (Fig. 4) 3.0 (including cucullus) long, 1.53 wide at posterior edge of leg II; general color dark red; anterior margin slightly concave; tubercles scattered on anterior half and densely distributed on posterior half of carapace. Abdomen (Fig. 2) 3.01 long, 2.14 wide; median plate of tergite XI (Fig. 6) with anterior margin much wider than posterior, XII quadrate, XIII longer than wide; venter of coxae III-IV smooth. Pygidium with shallow "V" notch on dorsal margin of basal segment. Cucullus (Fig. 4) 0.82 long, 1.07 wide; dark red; uniformly tuberculate (except near carapace). Chelicera: fixed finger with 5 teeth (distal longer than others); movable finger with six teeth similar in size. Leg formula II-IV-III -I; femur I 2.9 times longer than wide; II 4.8 times longer than wide. Spermathecae as in figures 10-11.
Distribution. This species is known from caves located in S11 Plateau, Serra dos Carajás, Pará, Brazil.
Relationships. TOURINHO & SATUNINO (2010) summarized the known species group division of the genus Cryptocellus, proposing new arrangement and two new species groups (Adisi and Peckorum). The putative synapomorphy of Adisi-group (C. adisi Platnick, 1988, C. lisbethae Gozález-Sponga, 1998 and C. florezi Platnick & Garcia, 2008) is a single character, the fixed accessory piece of male leg III copulatory apparatus thin, curved and with rounded contour. This character is presented by C. canga sp. nov., although much wider and with a bilobate apex. C. canga sp. nov. shares with two species of Magnus-group (C. magnus Ewing, 1929, C. narino Platnick & Paz, 1979) the fixed tarsal process with bilobate apex, a feature not mentioned as diagnostic for the group, but differs from them by having a movable process thin (massive in Magnus-group, see PLATNICK & PAZ 1979). As some species of Magnus-group also shares with Adisi-group a fixed accessory piece curved and with rounded contour, such as C. narino (see PLATNICK & PAZ 1979, figs 10 and 11), and species of both groups present navicular setae that varies in shape, another remarkable feature not mentioned in diagnoses of Cryptocellus subgroups. We believed a review of species groups delimitation is needed.
ACKNOWLEDGMENTS
We are grateful to Vale do Rio Doce for having sponsored Renata Andrade and colleagues during the collecting trip, to IBAMA/ICMBio - National Forest of Carajás for collecting permits and to Luis B. Piló for help with the etymology. CNPq (472960-2008-3) and FAPESP (2008/06604-7) sponsored RPR.
LITERATURE CITED
Submitted: 17.IV.2012; Accepted: 27.VIII.2012.
Editorial responsibility: Antonio D. Brescovit
Corresponding author. E-mail: ricrocha@usp.br
- BONALDO, A.B. & R. PINTO-DA-ROCHA. 2003. On a new species of Cryptocellus from Brazilian Amazon (Arachnida, Ricinulei). Revista Iberica de Aracnologia 7:103-108.
- BOTERO-TRUJILLO, R. & G.A. PEREZ. 2008. A new species of Cryptocellus (Arachnida, Ricinulei) from northwestern Colombia. Journal of Arachnology 36:468-471.
- BOTERO-TRUJILLO, R. & G.A. PEREZ. 2009. A new species of Cryptocellus (Arachnida, Ricinulei) from the Kofán Territory in southwestern Colombia. Zootaxa 2050:56-64.
- COKENDOLPHER, J.C. 2000. First Cryptocellus from Suriname (Ricinulei). Memorie Societa Entomologica Italiana 78(2):515-520.
- COKENDOLPHER, J.C & T. ENRÍQUEZ, 2004. A new species and records of Pseudocellus (Arachnida: Ricinulei: Ricinoididae) from caves in Yucatán, Mexico and Belize. Texas Memorial Museum, Speleological Monographs 6:95-99.
- COOKE, J.A L. & M.U. SHADAB. 1973. New and little known ricinuleids of the genus Cryptocellus (Arachnida, Ricinulei). American Museum Novitates 2530:1-25.
- ESCHWEGE, W.L.V. 1822. Geognostisches Gemälde von Brasilien und wahrscheinliches Muttergestein der Diamanten. Weimar, 44p.
- HARVEY, M.S. 2003. Catalogue of the smaller arachnid orders of the world: Amblypygi, Uropygi, Schizomida, Palpigradi, Ricinulei and Solifugae. Collingwood, CSIRO Publishing, 385p.
- NASKRECKI, P. 2008. A new ricinuleid of the genus Ricinoides Ewing (Arachnida, Ricinulei) from Ghana. Zootaxa 1698:57-64.
- PINTO-DA-ROCHA, R. & A.B. BONALDO. 2007. A new species of Cryptocellus (Arachnida, Ricinulei) from Oriental Amazonia. Zootaxa 1386:47-51.
- PLATNICK, N.I. & L.F. GARCIA. 2008. Taxonomic notes on Colombian Cryptocellus (Arachnida, Ricinulei). Journal of Arachnology 36:145-149.
- PLATNICK, N.I. & M.U. SHADAB. 1977. On Amazonian Cryptocellus (Arachndia, Ricinulei). American Museum Novitates 2677:1-9.
- PLATNICK, N.I. & N. PAZ. 1979. On Cryptocellus magnus Group (Arachnida, Ricinulei). American Museum Novitates 2633:1-17.
- TERUEL, R. & L.F. ARMAS. 2008. Nuevo Pseudocellus Platnick 1980 de Cuba oriental y nuevos registros de Pseudocellus paradoxus (Cooke 1972) (Ricinulei: Ricinoididae). Boletín de la Sociedad Entomologica Aragonesa 43:29-33.
- TOURINHO, A.L & C.S. AZEVEDO. 2007. A new Amazonian Cryptocellus Westwood (Arachnida, Ricinulei). Zootaxa 1540:55-60.
- TOURINHO, A.L & R. SATURNINO. 2010. On the Cryptocellus peckorum and Cryptocellus adisi groups, and description of a new species of Cryptocellus from Brazil (Arachnida: Ricinulei). Journal of Arachnology 38:425-432.
- TOURINHO, A.L; N.F.L. MAN-HUNG & A.B. BONALDO. 2010. A new species of Ricinulei of the genus Cryptocellus Westwood (Arachnida) from northern Brazil. Zootaxa 2684:63-68.
- VALDEZ-MONDRAGÓN, A. & O.F. FRANCKE. 2011. Four new species of the genus Pseudocellus (Arachnida: Ricinulei: Ricinoididae) from Mexico. Journal of Arachnology 39:365-377.
Publication Dates
-
Publication in this collection
13 Nov 2012 -
Date of issue
Oct 2012
History
-
Received
17 Apr 2012 -
Accepted
27 Aug 2012