Abstracts
Trichomes reduce herbivore attack on plants by physically and/or chemically inhibiting movement or other activities. Despite evidence that herbivores are negatively affected by trichomes there also reports of insect counter-adaptations that circumvent the plant’s defense. This paper reports on a study that investigated the likely mechanisms employed by larvae of the nymphalid butterfly, Heliconius charithonia (L.), that allow it to feed on a host that is presumably protected by hooked trichomes (Passiflora lobata (Killip) Hutch). Evidence were gathered using data from direct observations of larval movement and behavior, faeces analysis, scanning electron microscopy of plant surface and experimental analysis of larval movement on plants with and without trichomes (manually removed). The latter involved a comparison with a non specialist congener, Heliconius pachinus Salvin. Observations showed that H. charithonia larvae are capable of freeing themselves from entrapment on trichome tips by physical force. Moreover, wandering larvae lay silk mats on the trichomes and remove their tips by biting. In fact, trichome tips were found in the faeces. Experimental removal of trichomes aided in the movement of the non specialist but had no noticeable effect on the specialist larvae. These results support the suggestion that trichomes are capable of deterring a non specialist herbivore (H. pachinus). The precise mechanisms that allow the success of H. charithonia are not known, but I suggest that a blend of behavioral as well as physical resistance mechanisms is involved. Future studies should ascertain whether larval integument provides physical resistance to trichomes.
Herbivory; mechanical defense; insect-plant interaction
Apesar de as evidências mostrarem que herbívoros são negativamente afetados pelos tricomas, há também relatos de contra-adaptações que sobrepujam as defesas das plantas. Este estudo busca os prováveis mecanismos usados pelas larvas da borboleta ninfalídea Heliconius charithonia (L.) que permitem que elas se alimentem de uma planta hospedeira que é, presumivelmente, protegida por tricomas uncinados (curvados) (Passiflora lobata (Killip) Hutch.). Para isso realizou-se observação direta de movimento e comportamento da larva, análise de fezes, microscopia eletrônica de varredura da superfície foliar e análise experimental do movimento de larvas em plantas com e sem tricomas (removidos manualmente). O experimento foi feito comparando o comportamento dessas larvas com o de larvas de um não-especialista, Heliconius pachinus Salvin. As larvas de H. charithonia são capazes de se desvencilhar do aprisionamento pelos tricomas usando força física. Além disso, ao movimentar-se, a larva espalha fios de seda sobre os tricomas e retira suas pontas com as mandíbulas. De fato, pontas de tricoma foram encontradas nas fezes das larvas. A remoção experimental dos tricomas auxiliou o movimento da larva não-especialista, mas não teve efeitos notáveis sobre a larva especialista. Os resultados confirmam que os tricomas são capazes de deter um herbívoro não especializado (H. pachinus). Os exatos mecanismos responsáveis pelo sucesso de H. charithonia ainda são desconhecidos, mas sugere-se que a combinação de mecanismos comportamentais e de resistência física estejam envolvidos e estudos futuros necessitam verificar a possibilidade de resistência física no tegumento das larvas.
Herbivoria; defesa mecânica; interação inseto-planta
ECOLOGY, BEHAVIOR AND BIONOMICS
Herbivore handling of a Plants trichome: the case of Heliconius charithonia (L.) (Lepidoptera: Nymphalidae) and Passiflora lobata (Killip) Hutch. (Passifloraceae)
Interação herbívoro-tricoma: o caso de Heliconius charithonia (L.) (Lepidoptera: Nymphalidae) e Passiflora lobata (Killip) Hutch. (Passifloraceae)
Márcio Z. Cardoso
Depto. Botânica, Ecologia e Zoologia, Centro de Biociências, Univ. Federal do Rio Grande do Norte, 59072-970 Natal, RN, mzc@cb.ufrn.br
ABSTRACT
Trichomes reduce herbivore attack on plants by physically and/or chemically inhibiting movement or other activities. Despite evidence that herbivores are negatively affected by trichomes there also reports of insect counter-adaptations that circumvent the plants defense. This paper reports on a study that investigated the likely mechanisms employed by larvae of the nymphalid butterfly, Heliconius charithonia (L.), that allow it to feed on a host that is presumably protected by hooked trichomes (Passiflora lobata (Killip) Hutch). Evidence were gathered using data from direct observations of larval movement and behavior, faeces analysis, scanning electron microscopy of plant surface and experimental analysis of larval movement on plants with and without trichomes (manually removed). The latter involved a comparison with a non specialist congener, Heliconius pachinus Salvin. Observations showed that H. charithonia larvae are capable of freeing themselves from entrapment on trichome tips by physical force. Moreover, wandering larvae lay silk mats on the trichomes and remove their tips by biting. In fact, trichome tips were found in the faeces. Experimental removal of trichomes aided in the movement of the non specialist but had no noticeable effect on the specialist larvae. These results support the suggestion that trichomes are capable of deterring a non specialist herbivore (H. pachinus). The precise mechanisms that allow the success of H. charithonia are not known, but I suggest that a blend of behavioral as well as physical resistance mechanisms is involved. Future studies should ascertain whether larval integument provides physical resistance to trichomes.
Key Words: Herbivory, mechanical defense, insect-plant interaction
RESUMO
Apesar de as evidências mostrarem que herbívoros são negativamente afetados pelos tricomas, há também relatos de contra-adaptações que sobrepujam as defesas das plantas. Este estudo busca os prováveis mecanismos usados pelas larvas da borboleta ninfalídea Heliconius charithonia (L.) que permitem que elas se alimentem de uma planta hospedeira que é, presumivelmente, protegida por tricomas uncinados (curvados) (Passiflora lobata (Killip) Hutch.). Para isso realizou-se observação direta de movimento e comportamento da larva, análise de fezes, microscopia eletrônica de varredura da superfície foliar e análise experimental do movimento de larvas em plantas com e sem tricomas (removidos manualmente). O experimento foi feito comparando o comportamento dessas larvas com o de larvas de um não-especialista, Heliconius pachinus Salvin. As larvas de H. charithonia são capazes de se desvencilhar do aprisionamento pelos tricomas usando força física. Além disso, ao movimentar-se, a larva espalha fios de seda sobre os tricomas e retira suas pontas com as mandíbulas. De fato, pontas de tricoma foram encontradas nas fezes das larvas. A remoção experimental dos tricomas auxiliou o movimento da larva não-especialista, mas não teve efeitos notáveis sobre a larva especialista. Os resultados confirmam que os tricomas são capazes de deter um herbívoro não especializado (H. pachinus). Os exatos mecanismos responsáveis pelo sucesso de H. charithonia ainda são desconhecidos, mas sugere-se que a combinação de mecanismos comportamentais e de resistência física estejam envolvidos e estudos futuros necessitam verificar a possibilidade de resistência física no tegumento das larvas.
Palavras-Chave: Herbivoria, defesa mecânica, interação inseto-planta
The surface of many plant species are covered with protective trichomes that can negatively impact wandering herbivores by physically obstructing the animals movement or releasing protective chemicals (Levin 1973, Van Dam & Hare 1998, Fordyce & Agrawal 2001). Plants of the genus Passiflora L. (Passifloraceae) are better known for the employment of cyanogenic-based chemical defense (Spencer 1988) than by the use of physical defense against herbivores. Yet, in the Pseudodysosmia Harms section [subgenus Decaloba (DC) Rchb.)] of the Passiflora all 18 species bear hollow, hook-like structures known as uncinate trichomes (MacDougal 1994), that are very effective in deterring caterpillars of the specialist herbivore Heliconius Kluk (Gilbert 1971). Among the Passifloraceae, uncinate trichomes are a unique feature of the Pseudodysosmia group and are found in no other species in the Passiflora genus (MacDougal 1994).
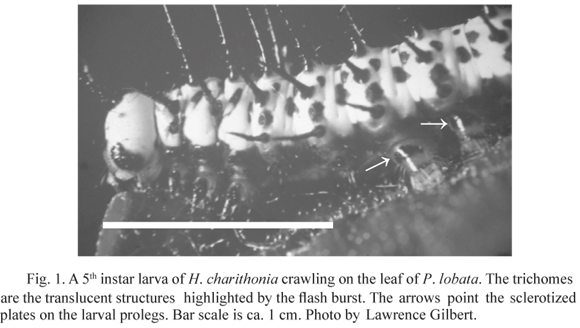
Although butterflies in the Heliconiiti group (Heliconius and related genera) are the major herbivores to attack Passiflora (Benson et al. 1975), Gilbert (1971) has shown that Heliconius larvae die from entrapment in the hooks of the trichomes of Passiflora adenopoda DC. In fact, in view of the seemingly insurmountable trichome defense, he stated that "it is difficult to imagine how heliconiines might circumvent the highly effective and specific mechanical defense of P. adenopoda without drastic developmental alterations". In fact, later studies have found that hooked trichome Passiflora species are virtually free of herbivores aside from a small subset of heliconiiti species (Benson et al. 1975, MacDougal 1994). Indeed, only two species are commonly listed as herbivores: Heliconius charithonia (L.) (Fig. 1) and Dione moneta Hübner (Lepidoptera: Nymphalidae) (Benson et al. 1975, MacDougal 1994). Therefore, although trichomes seem to be highly effective as a defense, some species have developed the ability to somehow circumvent the physical barrier, perhaps through developmental alterations as suggested by Gilbert (1971).
In view of the fact that, in general, some larvae of Heliconius are unable to cope with the trichomes while others are apparently unharmed, I set out to examine the likely mechanisms employed by H. charithonia in order to survive where other caterpillars perish. This report presents observational as well as experimental data with the goal of shedding light on this mechanistical question.
Material and Methods
The study was conducted using individuals collected from populations housed in glass greenhouses (4 x 6.5 m) at the University of Texas, Austin. Butterflies and the host plant derive from founders collected in Sirena Station, Corcovado National Park, Costa Rica. In Sirena, H. charithonia feeds on Passiflora lobata (Killip) Hutch., a member of the hooked trichome clade (Gilbert 1984, MacDougal 1994).
Larval behavior. Observations on larval behavior were made in the greenhouses directly on a full grown host plant, and in the laboratory. Larval observations in the laboratory were made on leaves collected in the greenhouse and kept in a vial with water to prevent wilting. Larval behavior was observed with the naked eye or under a scope, with the leaf slightly tilted so as to see the contact between the caterpillar and the leaf surface. The goals of these observations were to examine the general behavior of the caterpillars when moving or foraging on a leaf with trichomes. Most of these behavioral observations were made with the specialist herbivore, H. charithonia. A sample of faeces of H. charithonia larvae was diluted in distilled water and examined for leaf remains, a technique commonly employed in studying food choice in grasshoppers (Mulkern 1967). Additionally, opportunistic observations were made with larvae of other heliconiiti species: Dryas julia (Fabricius), Agraulis vanillae (L.) and H. erato (L.).
Experimental removal of trichomes. In order to ascertain whether larval movement is arrested by the trichomes, late instars (4th and 5th) of the specialist herbivore, H. charithonia, and of the non-specialist, H. pachinus Salvin, were placed on isolated P. lobata leaves and followed for several hours. Selection of H. pachinus was made to increase phylogenetic independence, because it belongs to a clade distinct from H. charithonia (Gilbert 1991).
Prior to larval placement, experimental P. lobata leaves were subjected to a shaving treatment for trichome removal. First, peeling was done by putting a stick tape on the leaf surface and pulling it out repeatedly, until the investigator deemed necessary. To ensure maximal trichome removal a shaver was also used. After peeling, it was passed as close to the surface as possible. Control leaves did not have their trichomes removed. Treated and untreated leaves were paired according to their size (as measured by the length of the main vein). The place where the larva was initially put was recorded on the leaf surface with a marking pen. Subsequently, larval movement was monitored for 5h at intervals of 1.5h.
A larval movement was recorded as such every time the larva moved away from the point where it was last seen. No movement was recorded when the larva did not move from the release point or from the point where it was last recorded. The number of moves made by a larva was counted and transformed into proportion of moves made (out of four possible moves), ranging from zero (no moves) to one (moved in all occasions). Since these data were not normally distributed I used a Wilcoxon non parametric test and compared movement rate for each species separately.
Trichome density. Observations of trichome characteristics were made under a Bausch & Lomb Stereo zoom scope, with the petiole immersed in water and with the light source aimed at a 90o angle from the leaf blade. Because trichomes are translucent and difficult to see under direct light, visibility was improved by spraying a light powder onto the surface of the leaf. Trichome density was estimated on a representative leaf by counting the number of trichomes in five randomly chosen areas (10 x 10 mm quadrats) on both the upper and lower leaf surfaces. Leaf area was estimated by scanning the leaf and using imaging software to compute size (NIH Image for the Macintosh).
Leaf SEM. Leaf samples of P. lobata kept in Sorensen buffer were fixed in 2.5% glutaraldehyde and rinsed in a buffer with distilled water solution three times. Subsequently, samples were dehydrated by an alcohol graded series (25-100%), followed by critical point dry, stub mounting and metal coating for SEM in a Hitachi S-340 scanning electron microscope. Pictures were taken to characterize trichome morphology and also to study damage caused by the herbivore. The areas of the leaf where a larva had been observed under the scope were marked and cut out for direct inspection.
Results
Larval behavior. In general terms, there is nothing special about the larval behavior of H. charithonia that sets it apart from other larvae. The crawling larva lays silk as it moves on the leaf and, at times, a trichome would seem to disturb the movement. For example, a leg would get stuck and, yet, the larva would simply pull it away. Apart from that, the most striking behavior was that the larva seemed to cut away some trichome tips as it moved. While performing this task their mandibles would on occasion get caught by the tip of the trichomes. Nevertheless, the larvae would pull their mandibles without any visible harm. Sometimes, that would make them spit gut contents. Most interestingly, though, H. charithonia would bit the tips of the trichomes in front of it. As a consequence, several trichome tips were found in the faeces sample, confirming the larvaes ability to handle the trichomes.
In some opportunistic observations, larvae of D. julia, A. vanillae and H. erato were followed on the leaf of P. lobata. D. julia moved as fast as H. charithonia. Leg pulling ability was seen on Dryas and Agraulis but not on H. erato. In fact, the single H. erato larva observed was trapped and dead 24h later after being put on the leaf.
Experimental removal of trichomes. Removal of trichomes increased movement of H. pachinus larvae (Fig. 2). On average, larvae on control (intact) plants moved 12.5%, while those on peeled leaves moved 75%. These differences were significant (Wilcoxon, c2 approximation = 5.25, df = 1, P = 0.022). In contrast, H. charithonia larvae moved equally well in both treatments with a tendency to move more in the treated leaf (62.5% vs. 83%, control vs. treatment; Wilcoxon, c2 approximation = 0.42, df = 1, P = 0.52) (Fig. 2)
Trichome density. Trichomes cover the whole plant in P. lobata. Trichome distribution on leaves varies between upper and lower surfaces, with more trichomes on the upper than on the lower surface. On a representative leaf, the average trichome density on the upper side was 15.8 ± 2.40 trichomes.mm-2 (n = 5 quadrats), while on the under side the density was 4.4 ± 1.73 trichomes.mm-2 (n = 5), a significant difference (t-test, t = 8.6, df = 8, P < 0.0001). For an estimated leaf area of 7.09 cm2, this gives an estimate of 11,202 trichomes on the upper surface and 3,120 on the lower surface.
Leaf SEM. The SEM pictures show a striking landscape (Fig. 3 A-I). The whole leaf blade is covered with menacing hooked trichomes. Although trichome measurements were not made, one can see at least two size classes with the same orientation (Fig. 3 A,B). Trichomes seem to be densely spread on the leaf blade (see section on estimates of trichome density), both on the blade itself (Fig. 3 A,B) and on veins (Fig. 3 A,C,E-I). The pictures in Fig. 3 also show the areas where the larvae crawled (Fig. 3 C-I), revealed by the presence of fine thread lines. These fine lines are larval silk spun by the caterpillars as a support for movement, and cover most of the surface, including the trichomes. In fact, perhaps adding insult to injury, trichomes may be used as supports for laying down the silk thread (Fig. 3 D-F). One can also see that many trichomes have their tips removed (Fig. 3 G-I), confirming observations of larval behavior made under the scope. Many trichomes present on the main vein from the lower surface, where the larvae H. charithonia move frequently, had their tips taken away.


Discussion
The role of trichomes as effective mechanical barriers to herbivores is well established (Levin 1973, Valverde et al. 2001, Hanley et al. 2007). In fact, both Gilbert (1971) and Pillemer & Tingey (1976) demonstrated with stunning scanning electron images of dead herbivores the dramatic consequences of entrapment by hooked trichomes. Many other studies have shown the negative effect of trichomes on herbivores (e.g., Hoffman & McEvoy 1986, Wilkens et al 1996, Van Dam & Hare 1998, Medeiros & Moreira 2002) by traits such as trichome presence, density, shape, length and glandular nature (Voigt et al. 2007). In my study, trichome presence and shape affected larval movement of non specialist Heliconius, confirming its defensive role. On the other hand, movements by the specialist H. charithonia were not significantly affected by them, yet larvae tended to wander more on the treated leaf. In fact, even for specialists, trichomes may increase handling time and/or time spent moving on a leaf (Fordyce & Agrawal 2001) and they therefore incur a cost to the herbivore.
My observations on free ranging caterpillars showed that they usually stay on the under surface of the leaf, where trichome density is lower, crawling along the main vein up to the tip of the leaf. The fact that many hookless trichomes were found in this area suggests that hook removal may be an important adaptation that enables H. charithonia to avoid getting trapped by the trichomes, a behavior that seems to be restricted to H. charithonia. Moreover, the fact that the larval faeces contained trichome hooks (and not the whole trichome) suggests that trichome trimming may be needed in order to handle trichomes. Finally, the potential harm trichomes may incur on a specialist was unknowingly demonstrated when I dropped a H. charithonia larvae on a P. lobata leaf and accidentally injured it; the larvae subsequently died.
Silk weaving may also be important because it provides a surface to which the crochets can connect that is independent of leaf anatomy (e.g., Rathcke & Poole 1975). However this alone cannot be considered a key characteristic given that larvae in general are known to weave in order to create a stable surface on which to crawl (Alexander 1961, Craig 1997, Sugiura & Yamazaki 2006). Quite possibly, morphology (eg Medeiros & Moreira 2002, Medeiros & Bolignon 2007) may be an important component of the suite that enables H. charithonia to handle the plants trichomes. For example, I noticed that crawling larva of H. charithonia can actually pull their legs from the hold of the trichome hook, something that larvae of D. julia and A. vanillae larvae were also capable of doing. A preliminary inspection on the legs of several Heliconius larvae did not reveal any noticeable difference in crochet size, number or arrangement that would explain this ability. However, the lateral sclerotized proleg plate (Fig. 1) of H. charithonia is more pigmented, which may indicate a tougher plate, and is similar in appearance to the ones in Dryas and Agraulis. Could this help these larvae avoid entrapment? This hypothesis clearly deserves further investigation.
Although no specific characteristic can be pinpointed as to how H. charithonia overcomes the hosts defenses, it seems likely that a suite of behavioral (eg hook removal, silk spinning) and mechanical traits (eg ability to detach from trichome hook) allow it do so. It would be interesting to ascertain whether there are costs associated with the seemingly ample advantage of exploiting a host that is unavailable to all other Heliconius. The herbivore offense by H. charithonia larvae (Karban & Agrawal 2002) suggests that it forages with finesse (sensu Dussourd 1993).
Acknowledgments
I would like to thank Larry Gilbert for enthusiastic discussions regarding the project and for collecting P. lobata for the experiments; R. Riess for helping in leaf tissue preparation and SEM operation; J. MacDougal for providing publication and discussions on the hooked Passiflora; G. Sword for suggesting the fecal analysis and A. Almeida, A. Freitas and J. Vasconcellos-Neto for improvements to the manuscript. Butterflies and plants were collected under permits from the Costa Rican government to Larry Gilbert. Thanks to CAPES for providing a doctoral fellowship.
Received 17/X/06. Accepted 20/III/08.
- Alexander, A.J. 1961. A study on the biology and behavior of caterpillars, pupae and emerging butterflies of the subfamily Heliconiinae in Trinidad, West Indies. Part I. Some aspects of larval behavior. Zoologica 46: 1-24.
- Benson, W.W., K.S. Brown & L.E. Gilbert. 1975. Coevolution of plants and herbivores: Passion flower butterflies. Evolution 29: 659-680.
- Craig, C.L. 1997. Evolution of arthropod silks. Annu. Rev. Entomol. 42: 231-267.
- Dussourd, D.E. 1993. Foraging with finesse: Caterpillar adaptations for circumventing plant defenses, p.92-131. In N.E. Stamp & T.M. Casey (eds.), Caterpillars, ecological and evolutionary constraints on foraging. Chapman & Hall, New York, 548p.
- Fordyce, J.A. & A. Agrawal. 2001. The role of plant trichomes and caterpillar group size on growth and defence of the pipevine swallowtail Battus philenor J. Anim. Ecol. 70: 997-1005.
- Gilbert, L.E. 1971. Butterfly-plant coevolution: Has Passiflora adenopoda won the selectional race with heliconiine butterflies? Science 172: 585-586.
- Gilbert, L.E. 1984. The biology of butterfly communities, p.41-54. In R. Vane-Wright & P.R. Ackery (eds.), The biology of butterflies. Academic Press, London, 429p.
- Gilbert, L.E. 1991. Biodiversity of a Central American Heliconius community: Patterns, process, and problems, p 403-427. In P.W. Price, T.M. Lewinsohn, G.W. Fernandes & W.W. Benson (eds.), Plant-Animal Interactions: Evolutionary ecology in tropical and temperate regions. Wiley-Interscience, New York, 639p.
- Hanley, M. E., R.B. Lamont, M.M. Fairbanks & C.M. Rafferty. 2007. Plant structural traits and their role in anti-herbivore defence. Perspect. Plant Ecol. Evol. Syst. 8: 157-178.
- Hoffman, G.D. & P.B. McEvoy. 1986. Mechanical limitations on feeding by meadow spittlebugs Philaenus spumarius (Homoptera: Cercopidae) on wild and cultivated host plants. Ecol. Entomol. 11: 415-426.
- Karban, R & A. Agrawal. 2002. Herbivore offense. Annu. Rev. Ecol. Syst. 33: 641-664.
- Levin, D. 1973. The role of trichomes in plant defense. Q. Rev. Biol. 48: 3-15.
- MacDougal, J.M. 1994. Revision of Passiflora subgenus Decaloba section Pseudodysosmia (Passifloraceae). Syst. Bot. Monogr. 41: 1-146.
- Medeiros, L. & D.S. Bolignon. 2007. Adaptations of two specialist herbivores to movement on the hairy leaf surface of their host, Solanum guaraniticum Hassl (Solanaceae). Rev. Brasil. Entomol. 51: 210-216.
- Medeiros, L. & G.R.P. Moreira. 2002. Moving on hairy surfaces: modifications of Gratiana spadicea larval legs to attach on its host plant Solanum sisymbriifolium Entomol. Exp. Appl. 102: 295-305.
- Mulkern, G.B. 1967. Food selection by grasshoppers. Annu. Rev. Entomol. 12: 59-78.
- Pillemer, E.A. & W.M. Tingey. 1976. Hooked trichomes: a physical plant barrier to a major agricultural pest. Science 193: 482-484.
- Rathcke, B.J. & R.W. Poole. 1975. Coevolutionary race continues: Butterfly larval adaptation to plant trichomes. Science 187: 175-176.
- Spencer, K.C. 1988. Chemical mediation of coevolution in the Passiflora-Heliconius interaction, p.167-240. In K.C. Spencer (ed.), Chemical mediation of coevolution. Academic Press, San Diego, 609p.
- Sugiura, S. & K. Yamazaki. 2006. The role of silk threads as lifelines for caterpillars: Pattern and significance of lifeline-climbing behaviour. Ecol. Entomol. 31: 52-57.
- Valverde, P.L., J. Fornoni & J. Núñez-Farfán. 2001. Defensive role of leaf trichomes in resistance to herbivorous insects in Datura stramonium J. Evol. Biol. 14: 424-432.
- Van Dam, N. & J.D. Hare. 1998. Biological activity of Datura wrightii glandular trichome exudates against Manduca sexta larvae. J. Chem. Ecol. 24: 1529-1549.
- Voigt, D., E. Gorb & S. Gorb. 2007. Plant surface-bug interactions: Dicyphus errans stalking along trichomes. Arthropod-Plant Inter. 1: 221-243.
- Wilkens, R.T., G.O. Shea, S. Halbreich & N.E. Stamp. 1996. Resource availability and the trichome defense of tomato plants. Oecologia 106: 181-191.
Publication Dates
-
Publication in this collection
14 July 2008 -
Date of issue
June 2008
History
-
Accepted
20 Mar 2008 -
Received
17 Oct 2006