Abstracts
Studies of plant responses to defoliation are important to develop pasture management strategies. The objective of this study was to evaluate the population density of basal, aerial and reproductive tillers, tiller appearance and mortality rates, forage accumulation and sward structure in Marandu grass pastures under different grazing intensities. The experimental period was from January to June 2006, divided in three seasons: summer, autumn and winter. The pastures were continuously grazed using variable stocking rates. The grazing intensities corresponded to 15, 30 and 45 cm of sward height. The experiment was arranged in a complete randomized block design with three treatments and two replicates. The sward heights were measured twice a week. The response variables were: forage accumulation, forage mass and its morphological components; and population densities of basal (TPDb), aerial (TPDa) and reproductive (TPDr) tillers. The highest TPDb (P > 0.05) was recorded for the shortest sward pasture, and the highest TPDr (P<0.05) for the tallest sward pasture. Swards showed a tiller size/density compensation mechanism and, consequently, the forage accumulation was similar (P > 0.05) among the grazing intensities. Pasture with a sward height of 35 cm had 94% of sun light interception. The highest variations in forage accumulation and sward structure were more influenced by seasonal differences than by grazing intensities. Pastures of Marandu grass showed large flexibility in grazing management, which allowed it to be maintained at sward heights between 15 and 35 cm.
Brachiaria brizantha; light interception; morphological components; sward height; tiller population density
Estudos das respostas das plantas à desfolhação podem ser úteis para estabelecer estratégias adequadas de manejo do pasto. Os objetivos deste trabalho foram avaliar a densidade populacional de perfilhos basilares, aéreos e reprodutivos, as taxas de aparecimento e de mortalidade de perfilhos, o acúmulo de forragem e a estrutura do dossel em pastos de capim-marandu, submetidos a três intensidades de pastejo. O período experimental foi de janeiro a junho de 2006, dividido em três estações do ano: verão, outono e inverno. O sistema de pastejo utilizado foi o de lotação contínua, com taxa de lotação variável. As intensidades de pastejo corresponderam a 15, 30 e 45 cm de altura do dossel. O delineamento experimental foi o de blocos casualizados, com três tratamentos e duas repetições. As alturas do pasto foram monitoradas duas vezes por semana. As variáveis respostas foram: o acúmulo de forragem, a massa de forragem e seus componentes morfológicos e as densidades populacionais de perfilhos basilares (DPPb), aéreos (DPPa) e reprodutivos (DPPr). A DPPb foi maior (P<0,05) em pastos mantidos mais baixos e a DPPr foi maior (P<0,05) nos pastos mantidos mais altos. As taxas de acúmulo de forragem foram semelhantes (P>0,05) entre as intensidades de pastejo. Pastos manejados com 35 cm de altura apresentaram 94% de interceptação da luz solar incidente. As maiores variações no acúmulo de forragem e na estrutura do dossel foram mais influenciadas pelas estações do ano do que pela intensidade de pastejo. O capim-marandu apresenta grande flexibilidade de manejo, podendo ser manejado, sob lotação contínua, com alturas entre 15 e 35 cm, sem prejuízo para a sua estrutura.
Altura do dossel; Brachiaria brizantha; componentes morfológicos; densidade populacional de perfilhos; interceptação luminosa
ANIMAL PRODUCTION AND REPRODUCTION PRODUÇÃO E REPRODUÇÃO ANIMAL
Tillering and forage accumulation in Marandu grass under different grazing intensities1 1 Parte da dissertação da primeira autora apresentada ao Programa de Pós-Graduação em Ciência Animal da Universidade Federal de Mato Grosso do Sul (UFMS) para a obtenção do título de Mestre.
Perfilhamento e acúmulo de forragem do capim-marandu submetido a diferentes intensidades de pastejo
Maria Paula Cavuto Abrão CalvanoI; Valéria Pacheco Batista EuclidesII; Denise Baptaglin MontagnerIII; Beatriz LemppIV; Gelson dos Santos DifanteV; Renata Santos FloresVI; Sandra GalbeiroVII
IZootecnista, Mestra. Departamento de Medicina Veterinária e Zootecnia, Universidade Federal de Mato Grosso do Sul. Cidade Universitária, Caixa Postal 549, 79070-900, Campo Grande, MS. (Autora para correspondência). cavutoabrao@hotmail.com
IIEngenheira Agrônoma, PhD. Embrapa Gado de Corte. Bolsista Produtividade CNPq. BR-262, Caixa Postal 154, 79002-970, Campo Grande, MS. val@cnpgc.embrapa.br
IIIZootecnista, Doutora. Embrapa Gado de Corte. BR-262, Caixa Postal 154, 79002-970, Campo Grande, MS. denise@cnpgc.embrapa.br
IVZootecnista, Doutora. Faculdade de Ciências Agrárias, Universidade Federal da Grande Dourados. Rua João Rosa Góes, 1761, Vila Progresso, Caixa Postal 322, 79825-070, Dourados, MS. blempp@ufgd.edu.br
VZootecnista, Doutor. Departamento de Agropecuária, Universidade Federal do Rio Grande do Norte. Caixa Postal 1524,- Campus Universitário Lagoa Nova, 59072-970, Natal,RN. gdifante@ufrnet.br
VIZootecnista, Mestra. Embrapa Gado de Corte. Bolsista DTI-CNPq. BR-262, caixa postal 154, 79002-970, Campo Grande, MS. renata@cnpgc.embrapa.br
VIIZootecnista, Doutora. Departamento de Zootecnia, Universidade Estadual de Maringá. Av. Colombo, 5780, 87020-900, Maringá, PR. sgalbeiro@gmail.com
ABSTRACT
Studies of plant responses to defoliation are important to develop pasture management strategies. The objective of this study was to evaluate the population density of basal, aerial and reproductive tillers, tiller appearance and mortality rates, forage accumulation and sward structure in Marandu grass pastures under different grazing intensities. The experimental period was from January to June 2006, divided in three seasons: summer, autumn and winter. The pastures were continuously grazed using variable stocking rates. The grazing intensities corresponded to 15, 30 and 45 cm of sward height. The experiment was arranged in a complete randomized block design with three treatments and two replicates. The sward heights were measured twice a week. The response variables were: forage accumulation, forage mass and its morphological components; and population densities of basal (TPDb), aerial (TPDa) and reproductive (TPDr) tillers. The highest TPDb (P > 0.05) was recorded for the shortest sward pasture, and the highest TPDr (P<0.05) for the tallest sward pasture. Swards showed a tiller size/density compensation mechanism and, consequently, the forage accumulation was similar (P > 0.05) among the grazing intensities. Pasture with a sward height of 35 cm had 94% of sun light interception. The highest variations in forage accumulation and sward structure were more influenced by seasonal differences than by grazing intensities. Pastures of Marandu grass showed large flexibility in grazing management, which allowed it to be maintained at sward heights between 15 and 35 cm.
Key words:Brachiaria brizantha; light interception; morphological components, sward height, tiller population density.
RESUMO
Estudos das respostas das plantas à desfolhação podem ser úteis para estabelecer estratégias adequadas de manejo do pasto. Os objetivos deste trabalho foram avaliar a densidade populacional de perfilhos basilares, aéreos e reprodutivos, as taxas de aparecimento e de mortalidade de perfilhos, o acúmulo de forragem e a estrutura do dossel em pastos de capim-marandu, submetidos a três intensidades de pastejo. O período experimental foi de janeiro a junho de 2006, dividido em três estações do ano: verão, outono e inverno. O sistema de pastejo utilizado foi o de lotação contínua, com taxa de lotação variável. As intensidades de pastejo corresponderam a 15, 30 e 45 cm de altura do dossel. O delineamento experimental foi o de blocos casualizados, com três tratamentos e duas repetições. As alturas do pasto foram monitoradas duas vezes por semana. As variáveis respostas foram: o acúmulo de forragem, a massa de forragem e seus componentes morfológicos e as densidades populacionais de perfilhos basilares (DPPb), aéreos (DPPa) e reprodutivos (DPPr). A DPPb foi maior (P<0,05) em pastos mantidos mais baixos e a DPPr foi maior (P<0,05) nos pastos mantidos mais altos. As taxas de acúmulo de forragem foram semelhantes (P>0,05) entre as intensidades de pastejo. Pastos manejados com 35 cm de altura apresentaram 94% de interceptação da luz solar incidente. As maiores variações no acúmulo de forragem e na estrutura do dossel foram mais influenciadas pelas estações do ano do que pela intensidade de pastejo. O capim-marandu apresenta grande flexibilidade de manejo, podendo ser manejado, sob lotação contínua, com alturas entre 15 e 35 cm, sem prejuízo para a sua estrutura.
Palavras-chave: Altura do dossel, Brachiaria brizantha, componentes morfológicos, densidade populacional de perfilhos, interceptação luminosa.
INTRODUCTION
Marandu grass (Brachiaria brizantha Hochst. Bhp ex A. Rich cv. Marandu) is the most used forage species in areas of cultivated pastures in the Brazilian Savana (Macedo, 2005). According to Da Silva (2004), although this cultivar is one of the most studied, information on their responses to grazing management practices, growth patterns and forage production is still limited. In recent years, numerous studies have been conducted related to the control of grazing of tropical grasses, aiming to indentify the impacts of management strategies on the production and persistence of pastures.
For Lemaire & Chapman (1996), the responses of plants to disturbances promoted by defoliation can be seen as mechanisms to restore and maintain homeostatic patterns of growth, under which all resources are used to attain optimal plant growth. Therefore, changes in tiller density occur as a result of phenotypic plasticity, compensating for the different grazing managements used. Changes in tiller density allow the plant community to maintain a certain stability of production, within a considerable range of sward conditions (Hodgson & Da Silva, 2002). However, it causes changes in the arrangement and distribution of morphological components and therefore in the structural composition of forage mass, which, in turn, may result in changes in the grazing behavior and, consequently, in animal performance (Da Silva & Carvalho, 2005).
The objectives of this study were to evaluate the population density of basal, aerial and reproductive tillers, tiller appearance and mortality rates, forage production and canopy structure in Marandu grass pastures subjected to three grazing intensities under continuous stocking.
MATERIALS AND METHODS
The experiment was conducted at Embrapa Gado de corte, Campo Grande, MS (20º27' S latitude, 54º 37' W longitude and 530 m altitude). The experimental period was from January to July 2006, divided into three seasons: summer (January, February and March), autumn (April, May) and winter (June and July).
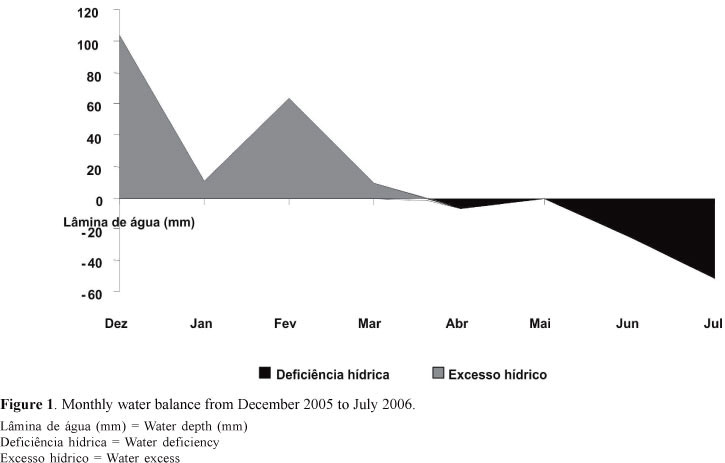
The climate is of the subtype Aw (Köppen classification), wet tropical savanna. The temperature and precipitation data used for calculating the monthly water balance during the experimental period were recorded by the weather station of Embrapa Gado de corte, distant about 4 km from the experimental area (Figure 1).
The soil of the experimental area was classified as a Dystrophic Red Latosol (Oxisol) with clayey texture, acidic pH, low base saturation, high levels of aluminum and low phosphorus (EMBRAPA, 2006). The pastures were established in November 2000 and, from March 2001 to September 2005, were grazed under continuous stocking. In November 2005, the maintenance fertilization was carried out, by applying 200 kg/ha of the formula 0-20-20 (NPK). Nitrogen fertilization was also performed, using 90 kg/ha of nitrogen in the form of urea, split in 2 applications: the first on 20/12/05 and the second on 28/02/06.
Each paddock (0.67 ha) was grazed by three cross steers (testers), with average initial weight of 250 kg, which were distributed at random. Additional animals (put & take) were placed and removed in the paddocks to maintain the sward at the target heights. Close to the experimental paddocks, areas were reserved for the maintenance of the regulator animals when they were not needed in the experimental area. The sanitary management of the put & take followed the schedule used by Embrapa Gado de corte.
The treatments consisted of pastures of Brachiaria brizantha cv. Marandu subjected to three grazing intensities, with target canopy heights of 15, 30 and 45 cm, maintained by continuous stocking with variable stocking rate. The experiment was arranged in a randomized block design with three treatments and two repetitions.
In December 2005, the average sward height was approximately 40 cm. Thus, a high stocking rate was used to reach the target heights of 15 and 30 cm rapidly. The sward height was measured twice a week at 60 points per paddock, distributed over four imaginary lines, with 15 points each, with a centimeter-graduaded ruler, measuring the distance from the ground to the average height of the curvature of grass leaves.
Light interception (LI) by the canopy and leaf area index (LAI) were measured by an AccuPAR Linear PAR/LAI ceptometer, Model - LP 80 (Decagon Devices). In each experimental unit, 60 readings were taken above the sward canopy and 60 readings at the base of the canopy, totaling 120 readings per paddock. LI was calculated as the difference between the incident radiation above the canopy and at the base of the sward canopy. LAI readings (leaf, stem and dead material) were provided by the ceptometer.
The tiller population density (TPD, tiller/m2) was obtained by counting the tillers inside a rectangular iron frame (0.75 x 0.25 cm), every 28 days. Eight areas, representative of the pasture condition, were chosen in each experimental unit, which remained fixed throughout the evaluation period. During each count, basal (TPDb), aerial (TPDa) and reproductive (TPDr) tillers were identified.
To evaluate the tillering demographic patterns, eight points of 0.0625 m2 (0.25 x 0.25 m) were marked at random, in each experimental unit. In the first evaluation (January 2006), all tillers in each section were marked with a color string tag, identifying the generation of reference tillers. Every 28 days, a new color was used to identify the new generations of tillers that appeared. At each count and labeling, the tillers were identified according to the location of their points of origin, either as basal or aerial tillers. Vegetative and reproductive tillers were also identified. The dead tillers were counted in each generation, which allowed the calculation of appearance (TAR) and mortality (TMR) rates of basal and aerial tillers (tillers/tiller.day). The missing string tags in each generation were not considered in the total amount of tillers per point. The stability index (P1/P0) of the tiller population (basal and aerial) was calculated according to Bahmani et al. (2003), using the equation:
P1/P0 = TSR (1 + TAR),
where: TSR (tiller survival rate) = 1-TMR.
For estimation of forage mass available (FM), 15 samples of 1 m2 were cut at ground level in each paddock, at random, every 28 days. The samples were subsampled twice: one subsample was dried at 65 °C and weighed to estimate the total dry matter availability, and the other was separated into leaves, culms (culms + sheath) and dead material. These data were used to estimate the masses of leaf blade (LBM), culm (CM) and dead material (DMM), as well as percentages of leaf blades (PLB), culm (PC) and dead material (PDM), in addition to the leaf:culm ratio (LCR). The forage mass (FM) and population density of basal tillers were used to estimate the weight of basal tillers (g DM/tiller), because they are usually more representative of the forage mass than the aerial tillers (Difante et al., 2008). For estimating forage accumulation, three grazing exclusion cages (1 m2) were used per paddock. Every 28 days, the cages were placed at points representative of the mean canopy height, with mass and morphological composition similar to the areas under grazing. Forage was cut at ground level for determination of dry masses. The total forage accumulation (kg DM/ha) was obtained by the difference between the forage mass from inside the cage (the cut at that day) and from the outside of the cage (the previous cut). After each cut, the cages were placed elsewhere in the paddock, using the same methodology. The forage accumulation rate (FAR: kg/ha/day of DM) was calculated by dividing the total forage accumulation by 28.
Data was grouped by season (summer, autumn and winter) and analyzed by a mathematical model containing the random effect of block and the fixed effects of canopy height, season and their interactions. Analyses were performed using the procedure "Mixed" (SAS Institute, 1996). Means were compared by the t test, at 5% probability level. In the case of significant interactions, means were compared by the probability of difference (PDIFF) option of SAS and the Tukey test at 5% probability level.
RESULTS AND DISCUSSION
The mean canopy heights remained below the target heights of 15, 30 and 45 cm, except in the pastures maintained at 15 cm. Thus, grazing intensities actually imposed corresponded to the mean heights of 15, 25 and 35 cm (Table 1). The reduction in forage accumulation over the experimental period and the small size of the paddocks (0.67 ha) made it difficult to maintain the three tester animals during the experiment; then, it was required to remove these animals gradually from the experimental area. At the end of the experiment, only one tester animal was allowed per paddock. The stocking rates used during the experiment were reported and discussed by Flores et al. (2008).
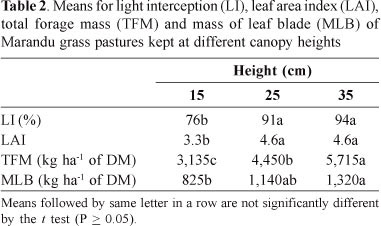
Higher leaf area indices (LAI) (P<0.01) were obtained in higher sward heights, resulting in higher light interception (LI) (P<0.01) from the top to the bottom of the canopy, and higher forage mass (P<0.01) (Table 2). Note that the LAI was estimated by the Accupar LP 80 ceptometer, which is usually different from the LAI estimated by the direct method (Molan, 2004). In this case, the LAI was not related only to leaf area, but also with the quantity and position of other parts of the plant within the canopy, suggesting that caution should be used in estimating LAI with the ceptometer. Molan (2004) also reported decreases in LAI from 5.7 to 1.7 and in the LI from 99.6 to 81.9%, as the height of Marandu grass was reduced from 40 to 10 cm under continuous stocking.
The management of Marandu grass at 35 cm allowed the canopy to reach the critical LAI at which point 95% of incoming light is intercepted (Brougham, 1957; Brown & Blaser, 1968). Parsons et al. (1988) showed that the maximum rate of forage accumulation occurred consistently associated with a value of LAI at which 95% of incident light was intercepted in pastures of perennial ryegrass (Lolium perenne L.), a result similar to that of Pedreira & Pedreira (2007) in Brachiaria brizantha cv. Xaraés.
There was no interaction (P > 0.05) between the effects of time and season for LI and LAI. LI did not vary between seasons (Pe"0.05), however, LAI was higher (P<0.05) in the summer compared with those observed in the autumn and winter: means were respectively 5.1, 3.9 and, 3.6. Verhagen et al. (1963) reported that there were many aspects involved in the light interception by crops. Some were related to the spatial arrangement of the leaves, with the horizontal and vertical distributions of leaves and leaf angle. Others were related to the functional characteristics dependent on factors of the plant and the environment, including age, type and size of leaves, light saturation and fluctuations in intensity and quality of light. Thus, the LAI only partially explains the light interception by the sward. Considering only the pastures that reached 95% of LI, the result for the LAI was the same as described above, with an average of 5.8, 4.5 and 4.2 for summer, autumn and winter, respectively. Molan (2004) also noted that the optimal LAI varied with the season, and the highest values were recorded in the summer, progressively reducing to early spring.
LI was lower than those found by Molan (2004) for Marandu grass under continuous stocking. The author found that starting from 20 cm, the LI by the canopy was 98%. Trinidad et al. (2007) reported for the same grass, but under intermittent grazing, LIs of 95 and 98% for the heights of 24 and 32 cm, respectively. The higher LIs with lower canopy heights found by these authors was probably the result of the high natural soil fertility and increased nitrogen fertilization used (167 and 190 kg N/ha, respectively), leading to different canopy structures. Positive correlation was found (P <0.01, r2 = 0.69) between canopy height and forage mass, corroborating the reports by other authors (Carnevalli et al., 2006; Barbosa et al. 2007; Pedreira et al., 2007; Trindade et al. 2007) for tropical grasses.
The population density of basal tillers (TPDb) was influenced by canopy height (P<0.01) and season (P<0.01). However, there was no interaction between these effects (P > 0.05). Pastures kept at 15 cm had higher TPDb, while those at 25 and 35 cm had lower TPDb (Figure 2A). Similar results were obtained by Sbrissa & Da Silva (2008) and Sbrissa et al. (2001). Leaf area is influenced by grazing intensity (Matthew et al., 2001) and determines the quantity and quality of radiation that reaches the base of the canopy and can activate axillary buds, producing new tillers (Deregibus et al., 1983). Therefore, the decrease in TPDb of pastures kept at higher heights is related to the competition for light among tillers, since there was greater light interception by the canopy (Table 2). In pastures kept at 15 cm, the incident light at the base of the canopy was read at 251 µMol/m2/s (P>0.01), while in pastures kept at 25 and 35 cm, the readings were 76 and 62 µMol/m2/s, respectively. In this way, a greater amount of incident light on the base of the canopy in pastures with more grazing intensities, resulted in the activation of axillary buds, which gave rise to new basal tillers (Figure 2A).
The highest TPDbs (P <0.01) were found in the summer; the intermediate in the autumn, and the lowest in the winter (Figure 2B). In relative terms, during the winter the TPDb was 76% of those observed during summer (Figure 2B). Environmental variations may be as important as hormonal factors in the development of buds and, consequently, interfere with the appearance of new tillers (Murphy & Brisk, 1992).The reduction in light intensity may have contributed to the reduction in TPDb along the seasons, since the highest incident light at the base of the canopy (P<0.01) was recorded during summer (201 µMol/m2/s); the intermediate value was recorded in the autumn (131 µMol/m2/s); and the lowest in the winter (57 µMol/m2/s). Besides, water deficiency during the winter can also be responsible for the reduction in TPDb (Figure 1). According to Nabinger & Bridges (2001), the water deficit hinders the development of both new and existing tillers.
There were no interactions (P > 0.05) between the effects of time and season for the rates of appearance of basal tillers (TARb) and mortality of basal tillers (TMRb) and the population stability index of basal tillers (ESTIb) of Marandu grass. Also, no differences were observed among the canopy heights (P > 0.05) for TARb and TMRb and for the ESTIb: the means were 0.60 tillers per 100 tillers.day; 0.34 tillers per 100 tillers.day; and 1.002 tillers per 100 tillers.day, respectively. However, there was effect (P<0.01) of the season for these variables (Table 3). The highest TARb was found during the summer, resulting not only of the high water and light availability, but also of the nitrogen fertilizations carried out in this season, hence ensuring a greater tiller production (Table 3). On the other hand, TMRb was higher (P<0.05) in the winter (Table 3), mainly because of the water deficit (Figure 1), shorter days and also the higher TPD in the summer (Figure 2B), a consequence of the development of the Marandu grass pastures, preparing for the peak bloom, which occurs mainly in the autumn.
The rates of appearance and mortality of tillers are important to understand the mechanisms involved in the perpetuation of the plants (Sbrissa & Da Silva, 2008). The balance between TARb and TMRb provides maintenance of stability and the size of the tiller population in the pasture and is described by ESTIb (Nabinger, 2002). ESTIb above 1.0 were found by Difante et al. (2008), throughout the experimental period, in Marandu grass pastures subjected to different intensities and frequencies of cutting. However, the decrease (P<0.05) in ESTIb, observed during the experimental period (Table 3), may be due to the beginning of the reproductive phase of the grass, which modifies the relationship between TARb and TMRb, besides the climatic constraints.
Stability indices below 1.0 indicate that the survival, together with the appearance of tillers, is not enough to counterbalance the mortality rate, hence the population tends to decrease. On the other hand, stability indices equal to 1.0 indicate a stable population of tillers, in which the number of tillers hardly varies, resulting in the balance of the tiller population (Bahmani et al. 2003). This index can indicate the persistence of a pasture, because the reduction in the number of tillers usually indicates the existence of processes of pasture degradation (Dias-Filho, 2007). Regardless of the canopy heights, even during the winter, with major climatic constraints, the ESTIb was very close to 1.0.
Pastures maintained at 15 cm had the lowest forage mass (Table 2), even with a larger number of basal tillers (Figure 2A). This phenomenon is known as a compensation mechanism between size (mass/tiller) and tiller population density (Sbrissa et al., 2001, 2003; Sbrissa & Da Silva, 2008). Size/density compensation is represented by reversible morphogenetic changes in individual plants in order to optimize their LAI, to ensure the sustainability of the pasture canopy and photosynthetic efficiency (Matthew et al., 1995). Tiller mass decreased (P<0.01) with the increase in grazing intensity. Hence, pastures kept high had small number of tillers (Figure 2A), but with greater mass (0.74 g DM/tiller), while pastures under high stocking rate showed a great number of tillers (Figure 2A), but with lower mass (0.30 g DM/tiller). However, pastures kept at 25 cm had tillers with intermediate mass (0.51 g DM/tiller).
There was interaction (P<0.01) between canopy height and season for population density of aerial tiller (TPDa). During the summer, pastures maintained at 35 cm had lower TPDa compared with those at other heights. In the autumn, the TPDa was similar among the different heights, while in the winter, pastures at 35 cm had TPDa higher than those maintained at 25 and 15 cm (Table 4). The higher TPDa during the summer, in pastures kept at 15 and 25 cm (Table 4), may be related to the rapid lowering of Marandu grass at the beginning of the experiment to obtain the target sward heights, by the decapitation of adult tillers, which promotes breaking of apical dominance and activation of axillary buds that can give rise to aerial tillers.
On the other hand, the increase in TPDa during the autumn and winter, in pastures kept at 35 cm (Table 4) was probably a result of stem elongation. Stem elongation allows the leaves to reach the top of the canopy in search of light, but the tiller becomes more susceptible to grazing, which removes the apical meristem and activates the axillary buds, giving rise to aerial tillers. According to Taiz & Zeiger (2004), the hormone auxin produced by the apical meristem maintains the apical dominance, inhibiting the development of axillary buds and therefore new tillers are not produced. When a tiller is decapitated, the apical dominance is released and aerial tillers are produced, ensuring the maintenance of leaf area and thus the photosynthetic rates. This was what happened in pastures lowered to 15 and 25 cm, as well as in pastures kept higher, at 35 cm, in different seasons (Table 4).
There was no interaction (P > 0.05) between height and season for rates of appearance of aerial tillers (TARa) and mortality (TMRa). Also, there were no differences between canopy heights (P > 0.05) for both variables. The TARa was higher (P<0.05) in the summer and the TMRa (P<0.05) was higher in winter, compared with those observed in other seasons (Table 5).
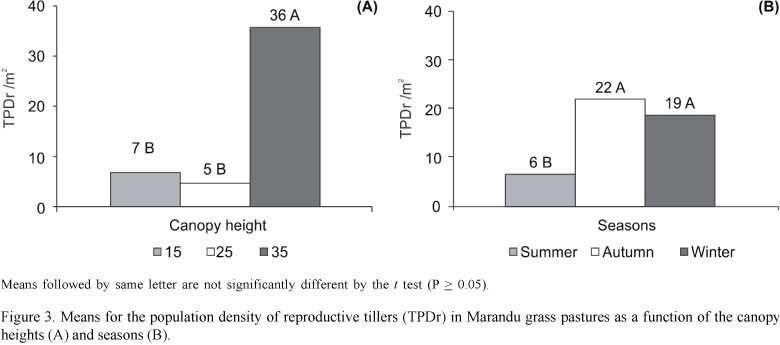
Pastures kept at 35 cm had higher (P<0.01) population density of reproductive tillers (TPDr) than those kept at 15 and 25 cm (Figure 3A). Maintaining the Marandu grass at higher heights allowed stem elongation because of the competition for light. The stem elongation together with the low grazing frequency, which usually occurs in pastures kept higher and under continuous stocking, allows these tillers, free of grazing, to reach the reproductive stage. Sbrissa (2004) also found a larger number of reproductive tillers in Marandu grass at 40 cm high when compared with those kept at 10 cm. Thus, grazing management can be an important tool for controlling the Marandu grass flowering.
The lowest (P<0.01) TPDr was recorded in the summer (Figure 3B), and the peak of flowering occurred in the Autumn. Reproductive tillers remain alive for most of the winter, going into senescence and decomposition only when the first rains start. This season is marked by the rise in temperature and humidity, which promote high tissue renewal and emergence of new tillers (Skinner & Nelson, 1992). A similar result was reported by Sbrissa (2004) in Marandu grass pastures maintained between 10 and 40 cm high.
Independent of the season, the forage accumulation rates (FAR) were similar among the pastures kept at different heights, with mean of 61 kg/ha of DM. Comparable forage accumulation rates were reported by Fagundes et al. (1999), Carvalho et al. (2000), and Bortolo et al. (2001) in pastures maintained at different heights. The use of grazing exclusion cages is probably responsible for the non-detection of differences in FARs at different heights, as they can cause overestimation of FAR in lower canopies and underestimation of those kept higher (Bortolo et al. 2001; Molan, 2004). However, Grant et al. (1983) and Bircham & Hodgson (1983) reported that the tiller size/density compensation in plant communities subjected to defoliation can make large variations between defoliation regimes result in small differences in the total amount of leaves produced per unit area. This compensation mechanism has been confirmed in several studies with tropical grasses kept under continuous stocking at different canopy heights (Carvalho et al. 2000; Sbrissa et al. 2001, 2003; Sbrissa & Da Silva, 2008).
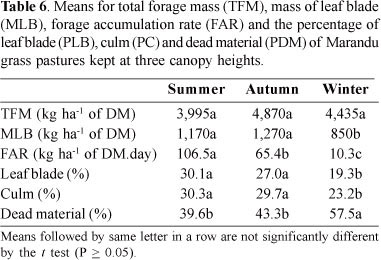
The mass of the leaf blade (MLB) was higher (P <0.05) for the pasture kept at 35 cm, intermediate at 25 cm and lower at 15 cm (Table 1). Because the FARs were similar among canopies of different heights, the different MLBs found for the pastures were the result of the grazing intensities.
There were no differences (P > 0.05) among the canopy heights for morphological components. The mean percentages were 25.6% for leaf blades (PLB); 27.7% for culms (PC); and 46.7% for dead material (PDM). No interaction was found (P > 0.05) between canopy height and season for total forage mass (TFM), mass of the leaf blade (MLB), forage accumulation rate (FAR) and morphological components. But, there were differences among seasons (P<0.05) for FAR, MLB and morphological components (Table 6). The highest FAR was recorded in the summer, intermediate in the autumn and the lowest in the winter (Table 6). Seasonal variations in forage accumulation were related to population density of basal tillers (Figure 2B). Similar behaviors were observed for the net accumulation of leaf blades in Marandu grass under continuous stocking (Molan, 2004) and under rotational grazing (Trinidad et al., 2007). Regardless of the canopy heights, the PLB and PC were higher (P<0.01) and PDM was lower (P <0.01) in the summer and autumn, when compared with those in the winter (Table 6). Consequently, the MLBs were higher in the summer and autumn (Table 6) than in the winter. Despite the higher population density of basal tillers (TPDb) (Figure 2B), resulting in higher FAR (Table 6) in the summer, this did not lead to increase in MLB, because of the higher stocking rate required to maintain the canopy height during this period, when compared with other seasons (Flores et al., 2008). On the other hand, the increase in dead material (PDM) throughout the year is typical of tropical grass pastures under continuous stocking (Euclides et al., 1999, 2000; Molan, 2004; Fagundes et al. 1999, 2006), because, during the growing season, there is accumulation of dead material associated with the natural senescence of grasses, which is accelerated by water stress. This was confirmed by the increase in mortality rates of basal (Table 3) and aerial (Table 5) tillers during the winter.
CONCLUSIONS
The morphogenetic and structural characteristics and, consequently, the forage accumulation in Marandu grass are strongly influenced by the seasons.
The use of high-intensity grazing, represented by the grazing height at 15 cm, is effective in controlling the flowering in Marandu grass pastures.
Marandu grass pastures show great management flexibility and can be kept at heights between 15 and 35 cm, under continuous stocking.
Recebido para publicação em 05/03/2009 e aprovado em 25/07/2011
Experimento financiado pela Embrapa Gado de Corte e CNPq.
- Bahmani I, Thom ER, Matthew C, Hopper RJ & Lemaire G (2003) Tiller dynamics of perennial ryegrass cultivars derived from different New Zeland ecotypes: effects of cultivar, season, nitrogen fertiliser, and irrigation. Australian Journal of Agricultural Research, 54:803-817.
- Barbosa RA, Nascimento Junior D, Euclides VPB, Da Silva SC, Zimmer AH & Torres Junior RAA (2007) Capim-tanzânia submetido a combinações entre intensidade e freqüência de pastejo. Pesquisa Agropecuária Brasileira, 42:329-340.
- Bircham JS & Hodgson J (1983) The influence of sward conditionon rates of herbage growth and senescence in mixed swardsunder continuous stocking management. Grass and Forage Science, 38:323-331.
- Bortolo M, Cacato U, Martins EM, Cano CCP, Coalho MR, Canto MW & Santos GT (2001) Avaliação de uma pastagem de coastcross-1 (Cynodon dactylon (L.) Pers) sob diferentes níveis de matéria seca residual. Revista Brasileira de Zootecnia, 30:627-635.
- Brougham RM (1957) Interceptation of light by foliage of pure and mixed swards of pasture plants. Australian Journal of Agricultural Research, 9:39-52.
- Brown RH & Blaser RE (1968) Leaf area index in pasture regrowth. Herbage Abstracts, 38:1-9.
- Carnevalli RA, Da Silva SC, Bueno AAO, Uebele MC, Bueno FO, Hodgson J, Silva GN, Morais JPG (2006) Herbage production andgrazin glosses in Panicummaximumcv. Mombaça under four grazing managements. Tropical Grasslands, 40:165-176.
- Carvalho CAB, Da Silva SC, Sbrissia AF, Pinto LFM, Carnevalli RA, Fagundes JL & Pedreira CGS (2000) Demografia do perfilhamento e taxas de acúmulo de matéria seca em capim tifton 85 sob pastejo. Scientia Agricola, 57:591-600.
- Da Silva SC (2004) Fundamentos para o manejo de pastagens de plantas forrageiras dos gêneros Brachiaria e Panicum In: 2º Simpósio Sobre Manejo Estratégico da Pastagem, Viçosa. Anais, Siprema, p. 347-685.
- Da Silva SC & Carvalho PCF (2005) Foraging behaviour and herbage intake in the favourable tropics/sub-tropics. In: Mcgilloway, D.A. (Ed.). Grassland: a global resource. Wageningen, Wageningen Academic, p. 81-95.
- Deregibus VA, Sanchez RA & Casal JJ (1983) Effects of light quality on tiller production in Lolium spp. Plant Physiology, 72:900-912.
- Dias-Filho MB (2007) Degradação de pastagens: processos, causas e estratégias de recuperação. 3Ş. ed, Belém, Embrapa Amazônia Oriental, 190p.
- Difante GS, Nascimento Júnior D, Da Silva SC, Euclides VPB, Zanine AM & Adese B (2008) Dinâmica do perfilhamento do capim-marandu cultivado em duas alturas e três intervalos de corte. Revista Brasileira de Zootecnia, 37: 189-196.
- Empresa Brasileira de Pesquisa em Agropecuária - Embrapa (2006) Centro Nacional de Pesquisa de Solos. Sistema Brasileiro de Classificação de Solos. 2. ed. Rio de Janeiro: CNPS, 2006. 306p.
- Euclides VPB, Cardoso EG, Macedo MCM & Oliveira MP (2000) Consumo voluntário de Brachiaria decumbens cv. Basilisk e Brachiaria brizantha cv. Marandu sob pastejo. Revista Brasileira de Zootecnia, 29:2200-2208.
- Euclides VPB, Thiago LRL, Macedo MCM & Oliveira MP (1999) Consumo voluntário de forragem de três cultivares de Panicum maximum sob pastejo. Revista Brasileira de Zootecnia, 28:1177-1185.
- Fagundes JL, Da Silva SC, Pedreira CGS, Sbrissia AF, Carnevalli RA, Carvalho CAB & Pinto LFM (1999) Índice de área foliar, interceptação luminosa e acúmulo de forragem em pastagem de Cynodon spp. Sob diferentes intensidades de pastejo. Scientia Agricola, 56: 187-195.
- Fagundes JL, Fonseca DM, Mistura C; Morais RV, Vitor CMT, Gomide JA, Nascimento Junior D, Casagrande DR & Costa LT (2006) Características morfogênicas e estruturais do capim-braquiária em pastagem adubada com nitrogênio avaliadas nas quatro estações do ano. Revista Brasileira de Zootecnia, 35:21-29.
- Flores RS, Euclides VPB, Abrão MPC, Galbeiro S, Difante GS & Barbosa RA (2008). Desempenho animal, produção de forragem e características estruturais dos capins marandu e xaraés submetidos a intensidades de pastejo. Revista Brasileira de Zootecnia, 37:1355-1365.
- Grant SA, Barthram GT, Torvell L, King J & Elston A (1983) Sward management, lamina turnover and tiller population-density in continuously stocked lolium-perenne-dominated swards. Grass and Forage Science, 38:333-344.
- Hodgson J & Da Silva SC (2002) Options in tropical pasture management. In: 39Ş Reunião Anual da Sociedade Brasileira de Zootecnia, Recife, Anais, SBZ, p.180-202.
- Lemaire G & Chapman D (1996) Tissue flows in grazed plant communities. In: Hodgson, J., Illius, A. W. (Eds.) The ecology and management of grazing systems. Guildford, Cab international. p.03-36.
- Macedo MCM (2005) Pastagens no ecossistema cerrados: evolução das pesquisas para o desenvolvimento sustentável. In: 2º Simpósio Sobre Pastagens nos Ecossitemas Brasileiros, Goiânia, Anais, SBZ. p.56-84.
- Matthew C, Lemaire G, Hamilton NRS & Hernández-Garay AH (1995) A modified self-thinning equation to describe size/density relationships for defoliated swards. Annals of Botany, 76: 579-587.
- Matthew C, Van Loo EN & Thom ER (2001). Understanding shoot and root development. In: 19º International Grassland Congress, Piracicaba, Proceedings, FEAQ, p.19-27.
- Molan LK (2004) Estrutura do dossel, interceptação luminosa e acúmulo de forragem em pastos de Capim-Marandu submetidos a alturas de pastejo por meio de lotação contínua. Dissertação de Mestrado. Piracicaba, Escola Superior de Agricultura Luiz de Queiroz. 159p. disponível em: http://www.teses.usp.br/teses/disponiveis/11/11139/tde-16092004-162818/ Acesso em: 16 de janeiro de 2009.
- Murphy JS & Briske DD (1992) Regulation of tillering by apical dominance - chonology, interpretive value, and current perspectives. Journal of Range Management, 45:419-429.
- Nabinger C & Pontes LS (2001) Morfogênese de plantas forrageiras e estrutura do pasto. In: 38Ş Reunião Anual da Sociedade Brasileira de Zootecnia, 38, 2001, Piracicaba, Anais, SBZ, p.755-771.
- Parsons AJ, Johnson IR & Williams JHH (1988) Leaf age struture and canopy photosyntesis in rotationally and continuously grazed swards. Grass and Forage Science, 43: 1-14.
- Pedreira BC, Pedreira CGS & Da Silva SC (2007) Estrutura do dossel e acúmulo de forragem de Brachiaria brizantha cultivar Xaraés em resposta a estratégias de pastejo. Pesquisa Agropecuária Brasileira, 42: 281-287.
- Pedreira C & Pedreira CGS (2007) Fotossíntese foliar do capim-xaraés Brachiaria brizantha (A. Rich.) Stapf. cv. Xaraés e modelagem da assimilação potencial de dosséis sob estratégias de pastejo rotativo. Revista Brasileira de Zootecnia, 36: 773-779.
- SAS INSTITUTE. SAS/STAT. (1996). User's guide statistics, versão 6, 4. ed., Cary, USA: v.1,2.
- Sbrissia AF (2004) Morfogênese, dinâmica do perfilhamento e do acúmulo de forragem em pastos de capim-marandu sob lotação contínua. Tese de doutorado. Piracicaba, escola Superior de Agricultura Luiz de Queiroz. 171p. Disponível em: http://www.teses.usp.br/teses/disponiveis/11/11139/tde-25052004-143513/ Acesso em: 16 de janeiro de 2009.
- Sbrissia AF & Da Silva SC (2008) Compensação tamanho/densidade populacional de perfilhos em pastos de capim-marandu. Revista Brasileira de Zootecnia, 37:35-47.
- Sbrissia AF, Da Silva SC, Carvalho CAB, Carnevalli RA, Pinto LFM, Fagundes JL & Pedreira CGS (2001) Tiller size/population density compensation in grazed coastcross bermudagrass swards. Scientia Agrícola, 58:655-665.
- Sbrissia AF, Da Silva SC, Matthew C, Carvalho CAB, Carnevalli RA, Pinto LFM, Fagundes JL & Pedreira CGS (2003) Tiller size/density compensation in grazed Tifton 85 bermudagrass swards. Pesquisa Agropecuária Brasileira, 38:1459-1468.
- Skinner RH & Nelson C J (1992) Estimation of potential tiller production and site usage during tall fescue canopy development. Annals of Botany, 70:493-499.
- Taiz L & Zeiger E (2004) Fisiologia vegetal. Santarém ER (Tradução) 3Ş ed. Porto Alegre:Artmed, 719 p.
- Trindade JK, Da Silva SC, Souza Juniour SJ, Giacomini AA, Zeferino CV, Guarda VDA & Carvalho PCF (2007) Composição morfológica da forragem consumida por bovinos de corte durante o rebaixamento do capim-marandu submetido a estratégias de pastejo rotativo. Pesquisa Agropecuária Brasileira, 42:883-890.
- Verhangen AMW, Wilson JH & Britten EJ (1963) Plant production in relation to foliage illumation. Annals of Botany, 27:626-640.
Publication Dates
-
Publication in this collection
17 Jan 2012 -
Date of issue
Dec 2011
History
-
Accepted
25 July 2011 -
Received
05 Mar 2009