Resumos
Neste estudo foi determinado a cobertura vegetal e a composição florística em 38 parcelas de 3,75 ha (250 m x 150 m), distribuídas por 30.000 ha na savana de Alter do Chão, Município de Santarém, Pará. Nas 38 parcelas foram registradas 130 espécies em 45 famílias. As únicas espécies de dicotiledôneas que cobriram 1% ou mais da área no estrato herbáceo-arbustivo foram Dioclea bicolor Benth. e Lafoensia pacari A. St.-Hil. A maior parte da área foi coberta pelas gramíneas Paspalum carinatum Humb. & Bonpl. ex Flügge (16%) e Trachypogon plumosus (Humb. & Bonpl. ex Willd.) Nees (22%). A gramínea Axonopus canescens (Nees ex Trin.) Pilg. e a ciperácea Rhyncospora hirsuta Vahl também cobriram mais que 1% da área. Apenas oito espécies, Anacardium occidentale L., Himatanthus fallax (Müll. Arg.) M. M. Plumel, Lafoensia pacari A. St.-Hil., Byrsonima coccolobifolia Kunth, Byrsonima crassifolia (L.) Kunth, Pouteria ramiflora (Mart.) Radlk., Qualea grandiflora Mart. e Salvertia convallariodora A. St.-Hil. tinham copas no estrato arbóreo que projetaram sobre mais que 1% da área. Destas, somente B. crassifolia (5,7%), S. convallariodora (6,0%) e P. ramiflora (2,1%) projetaram sobre mais que 2% da área. Cinqüenta e três por cento da área não tinha cobertura de arbustos, gramíneas ou ciperáceas, e 45% também não tinha cobertura de copas de árvores. Gramíneas e ciperáceas cobriram em média 39,2% das parcelas, e arbustos 11,0%. As correlações entre matrizes de similaridade para as espécies em diferentes estratos e grupos taxonômicos da vegetação foram geralmente baixas e houve pouca correlação entre matrizes baseadas em dados quantitativos e matrizes baseadas em dados de presença/ausência. Portanto, deve-se ter cautela em comparações entre áreas de savana baseadas somente em um estrato vegetativo ou em um grupo taxonômico.
Amazônia; cobertura arbórea; cobertura de ciperáceas; cobertura herbácea-arbustiva; cobertura de gramíneas
In this study, we determined the floristic composition in 38 plots of 3.75 ha (250 m x 150 m) distributed throughout 30.000 ha of savannas of " Alter do Chão" , Santarém Municipality. Our surveys revealed 130 species in 45 families. The only species of dicotoledons that covered 1% or more of the area in the herb-shrub layer were Dioclea bicolor Benth. and Lafoensia pacari A. St.-Hil. Most of the area was covered by the grasses Paspalum carinatum Humb. & Bonpl. ex Flügge (16%) and Trachypogon plumosus (Humb. & Bonpl. ex Willd.) Nees (22%). The grass Axonopus canescens (Nees ex Trin.) Pilg. and the sedge Rhyncospora hirsuta Vahl also covered slightly more than 1% of the area. Only eight species, Anacardium occidentale L., Himatanthus fallax (Müll. Arg.) M. M. Plumel, Lafoensia pacari A. St.-Hil., Byrsonima coccolobifolia Kunth, Byrsonima crassifolia (L.) Kunth, Pouteria ramiflora (Mart.) Radlk., Qualea grandiflora Mart. and Salvertia convallariodora A. St.-Hil. had canopies in the tree layer which projected over more than 1% of the area. Of these, only B. crassifolia (5.7%), S. convallariodora (6.0%) and P. ramiflora (2.1%) projected over more than 2% of the area. Most of the area (mean = 53%) did not have any grass, bush or sedge cover and 45% also had no tree-canopy cover. Grasses and sedges covered a mean of 39.2% of the plots, and shrubs 11.0%. The correlations between similarity matrices based on species in different vegetation strata and taxonomic groups were generally low and there was little correlation between matrices based on quantitative data and matrices based on presence/absence. Therefore, caution should be exercised in comparisons among savanna areas based on only one vegetative stratum or on only one taxonomic group.
Amazonia; grass cover; herb-shrub cover; sedge cover; tree cover
ARTIGOS
Composição florística e cobertura vegetal das savanas na região de Alter do Chão, Santarém PA
Floristic composition and vegetation cover of the savannas in the " Alter do Chão" region, Santarém PA
William E. MagnussonI,1 1 . Autor para correspondência: bill@inpa.gov.br ; Albertina P. LimaI; Ana L.K.M. AlbernazII; Tânia M. SanaiottiI; Jean-Louis GuillaumetIII
IInstituto Nacional de Pesquisas da Amazônia, Coordenação de Pesquisas em Ecologia, Caixa Postal 478, 69.011-970 Manaus AM, Brasil
IIMuseu Paraense Emílio Goeldi, Campus de Pesquisa/Coordenação de Ciências da Terra e Ecologia, Av. Perimetral 1901, 66.077-530 Terra Firme, Belém PA, Brasil
IIIMuséum National d'Histoire Naturelle, Laboratoire de Phanérogamie, 16, rue Buffon, 75005 Paris, França
RESUMO
Neste estudo foi determinado a cobertura vegetal e a composição florística em 38 parcelas de 3,75 ha (250 m x 150 m), distribuídas por 30.000 ha na savana de Alter do Chão, Município de Santarém, Pará. Nas 38 parcelas foram registradas 130 espécies em 45 famílias. As únicas espécies de dicotiledôneas que cobriram 1% ou mais da área no estrato herbáceo-arbustivo foram Dioclea bicolor Benth. e Lafoensia pacari A. St.-Hil. A maior parte da área foi coberta pelas gramíneas Paspalum carinatum Humb. & Bonpl. ex Flügge (16%) e Trachypogon plumosus (Humb. & Bonpl. ex Willd.) Nees (22%). A gramínea Axonopus canescens (Nees ex Trin.) Pilg. e a ciperácea Rhyncospora hirsuta Vahl também cobriram mais que 1% da área. Apenas oito espécies, Anacardium occidentale L., Himatanthus fallax (Müll. Arg.) M. M. Plumel, Lafoensia pacari A. St.-Hil., Byrsonima coccolobifolia Kunth, Byrsonima crassifolia (L.) Kunth, Pouteria ramiflora (Mart.) Radlk., Qualea grandiflora Mart. e Salvertia convallariodora A. St.-Hil. tinham copas no estrato arbóreo que projetaram sobre mais que 1% da área. Destas, somente B. crassifolia (5,7%), S. convallariodora (6,0%) e P. ramiflora (2,1%) projetaram sobre mais que 2% da área. Cinqüenta e três por cento da área não tinha cobertura de arbustos, gramíneas ou ciperáceas, e 45% também não tinha cobertura de copas de árvores. Gramíneas e ciperáceas cobriram em média 39,2% das parcelas, e arbustos 11,0%. As correlações entre matrizes de similaridade para as espécies em diferentes estratos e grupos taxonômicos da vegetação foram geralmente baixas e houve pouca correlação entre matrizes baseadas em dados quantitativos e matrizes baseadas em dados de presença/ausência. Portanto, deve-se ter cautela em comparações entre áreas de savana baseadas somente em um estrato vegetativo ou em um grupo taxonômico.
Palavras-chave: Amazônia, cobertura arbórea, cobertura de ciperáceas, cobertura herbácea-arbustiva, cobertura de gramíneas
ABSTRACT
In this study, we determined the floristic composition in 38 plots of 3.75 ha (250 m x 150 m) distributed throughout 30.000 ha of savannas of " Alter do Chão" , Santarém Municipality. Our surveys revealed 130 species in 45 families. The only species of dicotoledons that covered 1% or more of the area in the herb-shrub layer were Dioclea bicolor Benth. and Lafoensia pacari A. St.-Hil. Most of the area was covered by the grasses Paspalum carinatum Humb. & Bonpl. ex Flügge (16%) and Trachypogon plumosus (Humb. & Bonpl. ex Willd.) Nees (22%). The grass Axonopus canescens (Nees ex Trin.) Pilg. and the sedge Rhyncospora hirsuta Vahl also covered slightly more than 1% of the area. Only eight species, Anacardium occidentale L., Himatanthus fallax (Müll. Arg.) M. M. Plumel, Lafoensia pacari A. St.-Hil., Byrsonima coccolobifolia Kunth, Byrsonima crassifolia (L.) Kunth, Pouteria ramiflora (Mart.) Radlk., Qualea grandiflora Mart. and Salvertia convallariodora A. St.-Hil. had canopies in the tree layer which projected over more than 1% of the area. Of these, only B. crassifolia (5.7%), S. convallariodora (6.0%) and P. ramiflora (2.1%) projected over more than 2% of the area. Most of the area (mean = 53%) did not have any grass, bush or sedge cover and 45% also had no tree-canopy cover. Grasses and sedges covered a mean of 39.2% of the plots, and shrubs 11.0%. The correlations between similarity matrices based on species in different vegetation strata and taxonomic groups were generally low and there was little correlation between matrices based on quantitative data and matrices based on presence/absence. Therefore, caution should be exercised in comparisons among savanna areas based on only one vegetative stratum or on only one taxonomic group.
Key words: Amazonia, grass cover, herb-shrub cover, sedge cover, tree cover
Introdução
Savanas amazônicas constituem uma formação restrita, ocorrendo por toda a bacia Amazônica, que no Brasil, abrange uma superfície de 150.000 km2, cerca de 7% do total da Amazônia brasileira (Braga 1979). Ocorrem também em outros países amazônicos, como Suriname, Guiana, Guiana Francesa, Venezuela, Colômbia e Bolívia. A vegetação de savanas consiste de dois tipos funcionais: as gramíneas e as dicotiledôneas (Boutton et al. 1983, Scholes & Archer 1997). No entanto, a dinâmica de dicotiledôneas herbáceas e arbustivas em relação às queimadas é bastante diferente à das árvores (Sanaiotti & Magnusson 1995, Davis et al. 1998). Modelos de funcionamento de savanas geralmente consideram dicotomias entre gramíneas e outras formas de vida, ou entre o estrato herbáceo-arbustivo e o estrato arbóreo (e.g. Gignaux et al. 1996, Jeltsch et al. 1998). Fisionomicamente, as savanas são formações vegetais abertas com um estrato herbáceo sempre presente, estratos arbustivos e, ou, arbóreos mais ou menos desenvolvidos, sujeitos a queimadas.
A vegetação aberta ocorrente nos arredores de Santarém, localmente chamada de campo, é conhecida na literatura como " savana arenosa" (Bouillenne 1930 apud Aubréville 1961), " savana amazônica" (Beard 1953, Eiten 1977, 1978), " floresta seca semi-decídua" (Aubréville 1961), " campos" (Hueck 1972) e " cerrado" (Araujo et al. 1986). O Projeto Radambrasil (1974) utiliza o termo geral " savana" como equivalente de " cerrado" com duas faces dominantes " savana arbórea densa" (cerradão) e " savana parque" . Apesar desta variedade de terminologia, as savanas brasileiras são equivalentes às variações de savana descritas na nomenclatura internacional (Ferri 1983, Ribeiro et al. 1983, Sillbauer-Gottsberger & Gottsberger 1984, Durigan et al. 1987, Veloso et al. 1991, Ratter & Dargie 1992).
Do ponto de vista da composição florística, as savanas amazônicas, apesar de serem menos ricas que as savanas do Brasil central, apresentam várias espécies em comum, tanto em relação às herbáceas quanto às plantas lenhosas (Kubitski 1979, Kubitski 1983, Eiten 1984). Informações publicadas sobre a composição florística de savanas amazônicas estão disponíveis em Magnani (1952), Andrade-Lima (1958), Egler (1960), Takeuchi (1960), Azevedo (1967), Ledoux (1968, 1969), Dantas & Rodrigues (1982), Bastos (1984), Gottsberger & Morawetz (1986), Milliken & Ratter (1989), Miranda (1993, 1995), Miranda & Carneiro Filho (1994), Sanaiotti & Magnusson (1995) e Miranda et al. (2002). No entanto, é difícil fazer generalizações sobre a composição florística porque todos os estudos apresentam uma ou mais das seguintes limitações: (1) Não apresentam dados quantitativos de abundância, (2) não incluem espécies de todos os estratos da vegetação, (3) não descrevem a variação dentro de sub-regiões, (4) não definem claramente os limites da vegetação considerada " savana" .
Os objetivos deste estudo são descrever a composição florística e cobertura da vegetação de todas as áreas de savanas de região de Alter do Chão e avaliar a covariação entre os estratos herbáceo, arbustivo e arbóreo para determinar se a cobertura de um estrato pode ser utilizada para prever a cobertura dos outros estratos.
Material e métodos
Área de estudo A área de estudo está localizada na região entre Santarém e a Vila de Alter do Chão (2º31' S, 55º00' W) na margem direita do Rio Tapajós, Pará. A precipitação anual média na região é de 1.950 mm e a temperatura média é de 27,5 ºC (registro do Aeroporto de Santarém, 1970-1988), com uma estação seca pronunciada entre os meses de junho e dezembro, e uma estação chuvosa de janeiro a maio. A área estudada é uma savana que caracteriza-se por um estrato inferior essencialmente herbáceo de altura e densidade variáveis, um estrato arbustivo de 60-80 cm de altura e um estrato arbóreo que pode atingir até 10 metros de altura. Em algumas áreas são encontrados agrupamentos destes três estratos formando grupo espesso de plantas (moitas) de formas e tamanhos diferentes (Sanaiotti & Magnusson 1995). As savanas formam bordas abruptas com as formações florestais, e são facilmente reconhecidas em loco. Os levantamentos foram realizados no final da época chuvosa e início de seca entre abril e julho de 1998. Neste período, a vegetação normalmente já rebrotou e a biomassa vegetativa está recuperada das queimadas que ocorrem entre agosto e dezembro (obs. pessoal dos autores). A maior parte dos solos das parcelas é arenoso (Magnusson et al. 2002). Existem poucas áreas de savana com solos argilosos e, ou drenagem impedida na região, e estas não foram incluídas em nossas amostras.
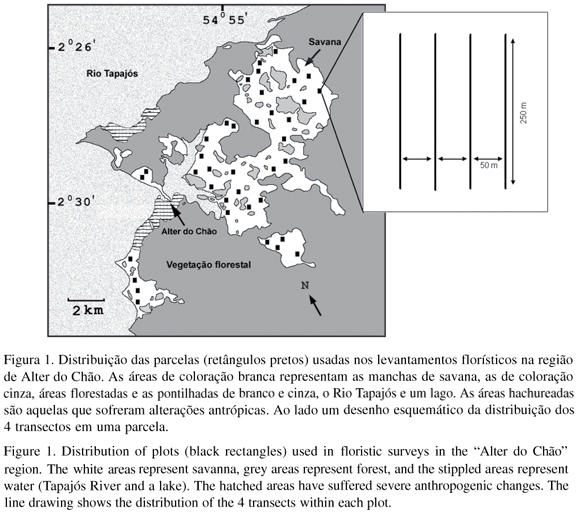
Transectos e parcelas A unidade amostral básica foi uma parcela que cobriu uma área de 3,75 ha (250 m x 150 m), contendo 4 transectos de 250 m, espaçados 50 m entre si (figura 1), e estabelecidos no sentido norte-sul. Trinta e oito parcelas foram distribuídas de maneira sistemática dentro de uma área de 30.000 ha visando cobrir de forma mais uniforme possível às áreas de savana delimitadas por meio de imagens de Landsat. Uma distância de 30 m foi mantida entre as parcelas e as áreas de floresta mais próximas, mas em uma parcela a extremidade de um dos transectos se estendeu até uma distância inferior a 10 m da floresta.
Registro e identificação das espécies nas parcelas Muitas espécies na savana têm índices de cobertura menor que 0,1%, e estas podem não ser registradas usando o método de pontos (Bullock 1996). Para incluir espécies raras e manter o mesmo esforço em cada parcela, foram registradas todas as espécies que ocorreram numa faixa de um metro do lado direito de cada linha. As espécies foram incluídas se alguma parte aérea da planta sobrepôs a faixa. Portanto, esta medida não é totalmente independente da cobertura média da espécie, mas nossas buscas fora desta faixa indicaram que poucas espécies presentes dentro da parcela não foram detectadas.
O sistema de classificação adotado segue o modelo de Cronquist (1988). A maioria dos indivíduos foi identificada por meio de um guia de campo que foi baseado no material anteriormente coletado por D. Ackerly, W. E. Magnusson, I. S. Miranda e T. M. Sanaiotti, identificado por especialistas e depositado no Herbário do Instituto Nacional de Pesquisas da Amazônia (INPA), Herbário da Universidade de Brasília (UB) e Herbário do Jardim Botânico de Edimburgo (E).
As espécies não identificadas em campo foram coletadas e identificadas por comparação com os espécimes do Herbário INPA. No entanto, no campo não foi possível diferenciar as espécies de Davilla (D. nitida e D. pedicellaris) no estádio estéril, portanto, registramos todos os indivíduos como Davilla spp. Também não foi possível diferenciar algumas espécies de Myrcia, que foram classificadas em dois morfotipos: morfotipo 1 consistindo principalmente de Myrcia lanceolata e, ou, Myrcia silvatica; morfotipo 2 de Myrcia amazonica e, ou, Myrcia obtusa. Amostras férteis de todas as espécies estão depositadas no herbário INPA. As amostras foram herborizadas após serem dessecadas em estufa seguindo técnicas descritas em Mori et al. (1989).
Definição de estratos Foram definidos dois estratos: o estrato inferior herbáceo-arbustivo com altura inferior a 2 m e o estrato arbóreo com altura superior a 2 m. O estrato herbáceo-arbustivo apresenta-se como um mosaico de manchas herbáceas, essencialmente de monocotiledôneas, manchas de arbustos, manchas mistas e outras associadas a grupos espesso de plantas (moitas). A dinâmica das dicotiledôneas com altura inferior a 2 m é muito influenciada por queimadas, independente da forma de vida ser herbácea, arbustiva ou escandente (Sanaiotti & Magnusson 1995). Portanto, as dicotiledôneas com altura inferior a 2 m integraram o estrato herbáceo-arbustivo.
Todas as plantas no estádio fértil foram classificadas em função das suas ocorrências nos estratos arbóreo (A), herbáceo-arbustivo (B) e quando elas possuíam potencial de ser adulta nos dois estratos (AB). Nas análises estatísticas envolvendo dados quantitativos, foram considerados os seguintes conjuntos funcionais: 1. todas as espécies de dicotiledôneas e monocotiledôneas menores que dois metros de altura (TODOS < 2 m); 2. todas as espécies de monocotiledôneas (MONOCOT); 3. ervas dicotiledôneas e arbustos de espécies que somente ocorrem no estrato B (DICOT < 2 m); 4. jovens e adultos de espécies de árvores que reproduzem no estrato A e no estrato B com altura menor que dois metros (ARV < 2 m); 5. espécies do estrato A com altura maior que dois metros (ARV > 2 m). Nas análises estatísticas envolvendo presença/ausência das espécies, foram considerados os seguintes conjuntos funcionais: 1. todas as espécies encontradas (TODAS); 2. todas as espécies de monocotiledôneas (MONOCOT); 3. ervas dicotiledôneas e arbustos de espécies que somente ocorrem no estrato B (DICOT) e 4. espécies do estrato A (ÁRVORES). Todas as lianas foram incluídas somente no estrato < 2 m.
Cobertura de indivíduos com altura menor que 2 m A cobertura relativa das espécies foi estimada com quadrados de ponto (Mantovani & Martins 1990, Bullock 1996) colocados a cada 2 m ao longo dos transectos, resultando em 500 pontos por parcela. Uma distância de 2 m entre os pontos garantiu independência entre os registros porque poucos arbustos nesta área têm diâmetro maior que 2 m. Os pontos foram localizados com uma trena de 50 m esticada e suspensa ao longo do transecto. Todas as espécies tocadas por uma vara vertical de metal de 2 mm de diâmetro, e de até 2 m de altura, foram registradas. Cada espécie foi registrada somente uma vez em cada ponto, independente do número de vezes que foi tocada pela vara.
Cobertura de espécies com altura maior que 2 m Nas savanas da área de estudo, o efeito do fogo resulta em uma diferenciação abrupta entre o estrato herbáceo-arbustivo e o estrato arbóreo a aproximadamente 2 m (Sanaiotti & Magnusson 1995). Normalmente não é viável determinar a cobertura arbórea usando o método de quadrados de ponto (Bullock 1996). Portanto, foi registrado somente se o ponto estava contido dentro da área geral da copa da árvore. Este método é subjetivo porque pessoas diferentes delimitam a posição da copa de maneiras diferentes. A questão de onde está o limite da copa é um exercício em geometria fractal (Mandelbrot 1967) e provavelmente não existe uma resposta global correta. Além disso, duas árvores com formas de copa semelhantes podem ter densidades foliares diferentes e, portanto, coberturas reais muito diferentes. Por esta razão, os valores de cobertura das árvores devem ser considerados somente como índices de cobertura, que não podem ser comparados diretamente com os valores de cobertura do estrato inferior, que são valores absolutos. Epífitas e hemiparasitas arborícolas não foram incluídas nos levantamentos.
Similaridade entre parcelas As análises foram baseadas em medidas de cobertura. Amostragens baseadas no número de indivíduos não permitem comparações entre espécies de estratos diferentes e, na savana, nem sempre é fácil separar os indivíduos, muitos dos quais resultam da reprodução vegetativa (Hoffmann & Moreira 2002). Todos os elementos da vegetação foram amostrados, permitindo a avaliação de covariação entre os estratos considerados.
Para determinar quanto das informações florísticas de um estrato podem ser usadas para prever as dissimilaridades florísticas entre parcelas baseadas em outro estrato, foram comparadas as matrizes de associação das parcelas, baseadas na composição das espécies, usando o teste de Mantel (McCune & Grace 2002). A dissimilaridade entre as parcelas usada na matriz de associação foi calculada para os dados quantitativos usando o índice de Bray e Curtis, e a similaridade baseada em dados de presença/ausência foi calculada usando o índice de Jaccard (McCune & Grace 2002) no programa PATN (Belbin 1992). As correlações de Pearson entre as matrizes de associação baseadas nas diferentes formas de vida foram calculadas no programa PATN e testadas usando o teste de Mantel com 1.000 permutações (Belbin 1992).
Resultados
Nos levantamentos foram registradas 130 espécies botânicas de 45 famílias. Destas, 26 espécies não foram detectadas nos levantamentos quantitativos e suas coberturas relativas não foram estimadas. Doze das espécies não detectadas nos levantamentos quantitativos foram detectadas em apenas uma parcela (tabela 1).
Sessenta e sete porcento das espécies foram encontradas reproduzindo no estrato abaixo de 2 m e 21% no estrato acima de 2 m (tabela 1). Somente 16 espécies foram encontradas reproduzindo freqüentemente em ambos estratos, nove destas espécies (Hirtella ciliata, Vismia glaziovii, Copaifera martii, Lafoensia pacari, Myrcia fallax, duas espécies não identificadas de Myrcia, Ouratea acuminata e Tocoyena formosa) são árvores pequenas ou arbustos grandes na área de estudo. As demais 7 espécies (Mandevilla hirsuta, Odontadenia lutea, Memora allamandiflora, Pleonotoma briottonii, Dioscorea melastomatifolia, Passiflora sp., Bredemeyera floribunda) consistem de cipós de crescimento rápido.
As únicas espécies de dicotiledôneas que cobriram 1% ou um pouco mais da área no estrato inferior a 2 m foram o arbusto Dioclea bicolor (1,1%) e a árvore Lafoensia pacari (1,2%). A maior parte da área foi coberta pelas gramíneas Paspalum carinatum (16%) e Trachypogon plumosus (22%). A gramínea Axonopus canescens (1,3%) e a Cyperaceae, Rhyncospora hirsuta (1,2%) também cobriram cerca de 1% da área. No estrato arbóreo (> 2 m), apenas oito espécies, Anacardium occidentale, Himatanthus fallax, Lafoensia pacari, Byrsonima coccolobifolia, Byrsonima crassifolia, Pouteria ramiflora, Qualea grandiflora e Salvertia convallariodora, tiveram copas que projetaram sobre mais que 1% da área (tabela 1). Destas, somente B. crassifolia (5,7%), P. ramiflora (2,1%) e S. convallariodora (6,0%) cobriram mais que 2% da área.
A maior parte da área (média = 53%, s = 9,9%) não tinha cobertura de arbustos, gramíneas ou ciperáceas. Gramíneas e ciperáceas cobriram em média 39,2%, s = 1,1% das parcelas, e arbustos 11,0%, s = 3,8%. Estas áreas não são mutuamente exclusivas porque alguns pontos tinham as duas formas de vida como cobertura. As copas das árvores cobriram em média 20,7%, s = 6,4% das parcelas.
Referente aos dados quantitativos, a correlação de Pearson entre a matriz de associação baseada em todas as espécies com altura < 2 m e a matriz de associação baseada somente em monocotiledôneas (gramíneas e ciperáceas) foi alta (r = 0,964) e apresentou uma probabilidade estatisticamente significativa (tabela 2), indicando que é a variação em gramíneas e ciperáceas que domina a associação baseada em todas as espécies com altura < 2 m. Em contraste, para dados de presença/ausência, a matriz de associação baseada em todas as espécies foi altamente correlacionada (r = 0,958) com as dicotiledôneas (herbáceas e arbustivas) e apresentou uma correlação mais fraca (r = 0,598) com as monocotiledôneas (tabela 3). Apesar de todas as matrizes de associação serem estatisticamente significantes correlacionadas entre si (parte superior das tabelas 2 e 3), a maioria das correlações de Mantel foram menores que 0,5 (parte inferior das tabelas 2 e 3), indicando que informações de um estrato têm pouco poder de previsão sobre outros estratos.
As correlações entre matrizes de associação baseada em dados numéricos e em dados de presença/ausência também foram baixas para as monocotiledôneas (r = 0,29), e o estrato herbáceo-arbustivo (r = 0,40). A presença/ausência de árvores nas parcelas também foi pouco relacionada com a cobertura das mesmas espécies no estrato herbáceo-arbustivo (r = 0,20) e no estrato arbóreo (r = 0,11). Tampouco, houve alta correlação entre a cobertura no estrato herbáceo-arbustivo e a cobertura no estrato arbóreo (r = 0,30).
Discussão
O método dos quadrados de ponto é mais usado em pequenas parcelas com vegetação herbácea (Mantovani & Martins 1990, Bullock 1996). No entanto, foi fácil adaptar a técnica para amostragens da camada vegetativa com altura inferior a 2 m. O método oferece alta precisão e registra as áreas abertas dentro das áreas foliares de plantas individuais. Isto é importante quando a distribuição geral de formas de vida não varia entre parcelas, mas as densidades ou áreas foliares divergem. Nas savanas da região de Alter do Chão, 53% da área apresentou solo descoberto, desprovido de cobertura de arbustos e gramíneas, e 45% também não tinha cobertura arbórea.
Apesar da alta precisão do método, comparações com outras áreas devem ser feitas com cautela. A vegetação na camada inferior a 2 m é muito variável sazonalmente devido aos efeitos das queimadas. Nós fizemos os levantamentos no final da época da chuva quando a cobertura vegetativa da maioria das espécies está no máximo. No entanto, a cobertura de algumas espécies anuais como Polycarpea corymbosa e Gymnopogon foliosus é muito menor nesta época que na época seca.
Apesar de algumas espécies arbóreas serem caducifólias (Miranda 1993), a cobertura de árvores é estável sazonalmente. A medida de cobertura de árvores é mais subjetiva e menos precisa que a medida de cobertura de plantas menores, pois o método de pontos não é acurado para árvores porque, usando-se uma vara, é difícil determinar se as folhas estão diretamente acima do ponto. Provavelmente, seria possível desenvolver um sistema óptico mais preciso usando um suporte e lentes de precisão. Nós somente determinamos se o ponto estava diretamente abaixo da área geral da copa. Este método inclui áreas sem folhas no meio da copa como parte da copa, e a determinação dos limites gerais de copa é um exercício em geometria fractal (Mandelbrot 1967). No entanto, o método é tão ou mais preciso quanto outros métodos geralmente usados para estimar a cobertura de árvores (Bullock 1996), e os índices de cobertura relativa são úteis para comparar as espécies.
Nossa definição de savana é paisagística e inclui somente as áreas dominadas por gramíneas, e que são facilmente identificáveis como savanas em imagens Landsat. Todas seriam consideradas savanas sazonais na classificação de Sarmiento e Monasterio (1985). Não incluimos as zonas de transição ou áreas florestadas, apesar de algumas espécies serem comuns a estes três habitats (por ex. Haridasan 1992, Longman & Jenik 1992).
Baseado na cobertura do estrato arbóreo, Salvertia convallariodora e Byrsonima crassifolia são as espécies de árvores dominantes na savana com coberturas de copas de 6,0% e 5,7% respectivamente. Miranda (1993), utilizando estimativas de densidade de indivíduos, chegou à mesma conclusão. Das 30 espécies com altura maior que 2 m em levantamentos quantitativos, três eram cipós e seis espécies (Hirtella ciliata, Casearia javitensis, C. sylvestris, Myrcia morfotipo 1, Palicourea rigida e Tocoyena formosa) ocorreram com maior freqüência, e reproduziram, no estrato herbáceo-arbustivo. Todas as 16 espécies arbóreas encontradas em levantamentos quantitativos por Miranda (1993) foram encontradas nos levantamentos quantitativos neste estudo.
Os levantamentos de presença/ausência acrescentaram somente quatro espécies de árvores aos levantamentos quantitativos (Tapirira guianensis, Miconia rufescens, Matayba guianensis, Cybistax antisyphilitica) das quais T. guianensis e M. guianensis são espécies abundantes nas matas da região (W. E. Magnusson obs. pess.). Em contraste, os levantamentos de presença/ausência revelaram 23 espécies dos estratos mais baixos que não foram detectadas nos levantamentos quantitativos e 56% destas ocorrem em áreas florestadas.
Este estudo mostra que as similaridades entre parcelas nesta escala dependem do estrato amostrado. A similaridade entre parcelas baseada em dados quantitativos para todas as espécies é determinada principalmente pelas monocotiledôneas. Isto é esperado porque as espécies do grupo de monocotiledôneas têm as maiores coberturas (Belbin 1992). As baixas correlações entre as árvores no estrato arbóreo e no estrato arbustivo não eram esperadas. Estes dois estratos muitas vezes são considerados em conjunto em modelos de funcionamento de savanas, porque ambos são dominados por dicotiledôneas e, na ausência de queimadas, muitas espécies do estrato arbustivo podem fazer parte do estrato arbóreo. Para a região das savanas de Alter do Chão, a abundância de arbustos e a abundância de árvores são determinadas por diferentes fatores.
Análises de presença/ausência atribuem o mesmo peso para todas as espécies, e espécies comuns que ocorrem em quase todas as parcelas, como as gramíneas Trachypogon plumosus e Paspalum carinatum, contribuem pouco ou nada para diferenças entre as parcelas. Nestas análises, as dicotiledôneas, que são menos abundantes, porém mais ricas em espécies, contribuem mais para as diferenças entre as parcelas. As correlações entre as matrizes de associação para diferentes estratos baseadas em presença/ausência foram geralmente mais altas que para os dados quantitativos, mas não suficientemente altas para permitir que um grupo seja usado como indicador de outro.
Em algumas associações vegetais, informações de um estrato prevêem bem a similaridade entre parcelas baseada em dados de espécies de outros estratos (por ex. Ruokolainen et al. 1997). No entanto, nossas análises indicam padrões de similaridade baseados em espécies de um estrato não devem ser generalizados para toda a vegetação de savanas amazônicas. Nas florestas, em escala local, todos os estratos estão relacionados a fatores edáficos. Nas savanas, a freqüência de queimadas adiciona um outro fator, e possivelmente isto explique a falta de co-variação entre os diferentes estratos.
Agradecimentos Devemos muito à hospitalidade dos moradores de Alter do Chão, especialmente Carlinhos Coelho e Jucinéia de Souza. A SUDAM concedeu o uso do Posto-Alojamento Alter do Chão. O estudo foi financiado pelo CNPq através de Auxílio integrado nº CA-05/95. Este estudo não teria sido possível sem o esforço inicial de coleta e identificação das plantas por David Ackerly e Izildinha S. Miranda, e pela determinação dos gêneros e espécies feitas pelos especialistas C. Proença, D. Zappi, G. Lewis, G. Prance, J. J. Wurdack, W. Punt, T. Filgueiras, D. A. Simpson, A. M. Giulietti, T. Pennington, A. S. da F. Vaz, R. A. P. Andreata, T. B. Cavalcanti, W. M. Ferreira, V. C. Souza, C. Farney, K. K. Athoff. Jakes Câmara foi responsável pelas comparações das amostras coletadas durante o presente estudo com material do Herbário do INPA.
(recebido: 22 de dezembro de 2005; aceito: 03 de março de 2008)
- ANDRADE-LIMA, D. 1958. Viagem aos campos de Monte Alegre, PA. Boletim Técnico do Instituto Agronômico do Norte 36:99-149.
- ARAÚJO, A.P., JORDY FILHO, S. & FONSECA, W.E. 1986. A vegetação da Amazônia. In Anais do Primeiro Simpósio do Trópico Úmido, II Flora e Floresta (E.S. Cruz, coord.). EMBRAPA-CPATU, Belém, v.1, p.135-152.
- AUBREVILLE, A. 1961. Étude écologique des principales formations végétales du Brésil, et contribution à la connaissance des forêts de l'Amazonie brésilienne. Nogent-sur-Marne, Centre Technique Forestier Tropical, França.
- AZEVEDO, L.G. 1967. Tipos eco-fisionômicos de vegetação do Território Federal do Amapá. Revista Brasileira de Geografia 29:25-51
- BASTOS, M.N.C. 1984. Levantamento florístico dos campos do estado do Pará: I Campo de Joanes (Ilha de Marajó). Boletim do Museu Paraense Emílio Goeldi, Série Botânica 1:67-68.
- BEARD, J.S. 1953. The savanna vegetation of northern tropical America. Ecological Monographs 23:149-215.
- BELBIN, L. 1992. PATN: pattern analysis package. CSIRO, Canberra.
- BOUTTON, T.W., ARSHAD, M.A. & TIESZEN, L.L. 1983. Stable isotope analysis of termite food habits in East African grasslands. Oecologia 59:1-6.
- BRAGA, P.I.S. 1979. Subdivisão fitogeográfica, tipos de vegetação, conservação e inventário florístico na Amazônia. Acta Amazonica 9:53-80.
- BULLOCK, J. 1996. Plants. In Ecological Census Techniques (W.F. Sutherland, ed.) Cambridge University Press, Cambridge, p.111-138.
- CRONQUIST, P.E.R. 1988. The evolution and classification of flowering plants. New York Botanical Garden, New York.
- DANTAS, M. & RODRIGUES, I.A. 1982. Estudos fitoecológicos do trópico úmido brasileiro. IV Levantamentos botânicos em campos do Rio Branco EMBRAPA-CPATU. Boletim de Pesquisa 40:1-31.
- DAVIS, M.A., WRAGE, K.J. & REICH, P.B. 1998. Competition between tree seedlings and herbaceous vegetation: support for a theory of resource supply and demand. Journal of Ecology 86:652-661.
- DURIGAN, G., SARAIVA, I.R., GURGEL GARRIDO, L.M.A., GARRIDO, M.A.O. & PECHE FILHO, A. 1987. Fitossociologia e evolução da densidade da vegetação do cerrado, Assis, SP. Boletim Técnico do Instituto Florestal. São Paulo 41:37-58
- EITEN, G. 1977. Delimitação do conceito de cerrado. Arquivo do Jardin Botânico do Rio de Janeiro 21:125-134.
- EITEN, G. 1978. Delimitations of the cerrado concept. Vegetatio 36:169-178.
- EITEN, G. 1984. Vegetation of Brasília, Brazil. Phytocoenologia 12:271-292.
- EGLER, W.A. 1960. Contribuições ao conhecimento dos campos da Amazônia. I. Os campos de Ariramba. Boletim do Museu Paraense Emílio Goeldi, nova série Botânica 4:1-36.
- FERRI, M.G. 1983. Os cerrados, um grupo de formas de vegetação semelhantes às savanas. Revista do Serviço Público, Brasília, 40:57-61.
- GIGNAUX, J., MENAUT, J.C., NOBLE, I.R. & DAVIES, I.D. 1996. A spatial model of savanna function and dynamics: model description and preliminary results. In Dynamics of tropical communities (D.M. Newberry, H.H.T. Prins & N.D. Brown, eds.) Blackwell Science, Oxford, p.361-383.
- GOTTSBERGER, G. & MORAWETZ, W. 1986. Floristic and phytogeographic analysis of the savannas of Humaitá (Amazonas). Flora 178:41-71.
- HARIDASAN, M. 1992. Observations on soils, foliar nutrient concentrations and floristic composition of cerrado sensu stricto and cerradão communities in central Brazil. In Nature and dynamics of forest-savanna boundaries (P.A. Furley, J. Proctor & J.A. Ratter, eds.) Chapman & Hall, London, p.171-184.
- HOFFMANN, W.A. & MOREIRA, A.G. 2002. The role of fire in population dynamics of woody plants. In The Cerrado of Brazil (P.S. Oliveira & R.J. Marquis, eds.) Columbia University Press, New York, p.159-177.
- HUECK, K. 1972. As florestas da América do Sul. Editora Universidade de Brasília, Brasília, Polígono, São Paulo.
- JELTSCH, F., MILTON, S.J., DEAN, W.J., ROOYEN, N.V. & MOLONEY, K.A. 1998. Modelling the impact of small-scale heterogeneities on tree-grass coexistence in semi-arid savannas. Journal of Ecology 86:780-793.
- KUBITSKY, K. 1979. Ocorrência de Kielmeyera nos campos de 'Humaitá' e a natureza dos 'campos' Flora da Amazônia. Acta Amazonica 9:401-404.
- KUBITSKY, K. 1983. Dissemination biology in the savanna vegetation of Amazonia. Sonderbände des naturwissenshaftlichen Vereins in Hamburg 7:353-357.
- LEDOUX, P. 1968. Estudos sobre Hancornia speciosa Gom. (mangabeira: Apocynaceae) na região equatorial amazônica. Ciência e Cultura 20:504-505.
- LEDOUX, P. 1969. Fitotopos, nas savanas equatoriais do Amapá, com desenvolvimento arbóreo ao máximo do potencial específico. Ciência e Cultura 21:433-444.
- LONGMAN, K.A. & JENIK, J. 1992. Forest-savanna boundaries: general considerations. In Nature and dynamics of forest-savanna boundaries (P.A. Furley, J. Proctor & J.A. Ratter, eds) Chapman & Hall, London, p.3-21.
- MAGNANI, A. 1952. As regiões naturais do Amapá. Revista Brasileira de Geografia 14:243-301.
- MAGNUSSON, W.E., SANAIOTTI, T.M., LIMA, A.P., MARTINELLI, L.A., VICTORIA, R.L., ARAÚJO, M.C. & ALBERNAZ, A.L. 2002. A comparison of d13C ratios of surface soils in savannas and forests in Amazonia. Journal of Biogeography 29:857-863.
- MANDELBROT, B.B. 1967. How long is the coast of Britain? Statistical self similarity and fractional dimension. Science 156:636-638.
- MANTOVANI, W. & MARTINS, F.R. 1990. O método de pontos. Acta Botânica Brasilica 4:95-122.
- McCUNE, B. & GRACE, J. B. 2002. Analysis of ecological communities. MjM Software Design, Oregon.
- MILLIKEN, W. & RATTER, J.A. 1989. The vegetation of the Ilha de Maracá. Royal Botanic Garden, Edinburgh.
- MIRANDA, I.S. 1993. Estrutura do estrato arbóreo do cerrado amazônico em Alter do Chão, Pará, Brasil. Revista Brasileira de Botânica 16:143-150.
- MIRANDA, I.S. 1995. Fenologia do estrato arbóreo de uma comunidade de cerrado em Alter-do-Chão, PA. Revista Brasileira de Botânica 18:235-240.
- MIRANDA, I.S. & CARNEIRO FILHO, A. 1994. Similaridade florística de algumas savanas amazônicas. Boletim Museu Paraense Emílio Goeldi, série Botânica 10:249-267.
- MIRANDA, I.S., ABSY, M.L. & REBÊLO, H. 2002. Community structure of woody plants of Roraima savannas, Brazil. Plant Ecology 164:109-123.
- MORI, S.A., SILVA, L.A.M., LISBOA, G. & CORADIN, L. 1989. Manual de manejo do herbário fanerogâmico. CEPLAC, Ilhéus.
- RADAMBRASIL. 1974. Projeto Radam. Levantamento de recursos naturais. Ministério de Minas e Energia-DNPN, Rio de Janeiro.
- RATTER, J.A. & DARGIE, T.C.D. 1992. An analysis of the floristic composition of 26 cerrado areas in Brazil. Edinburgh Journal of Botany 49:235-250.
- RIBEIRO, J.F., SANO, S.M., MACEDO, J. & DA SILVA, J.A. 1983. Os principais tipos fitofisionômicos da região dos cerrados. EMBRAPA/CPAC (Empresa Brasileira de Pesquisa Agropecuária Centro de Pesquisa Agropecuária do Cerrado) Boletim de Pesquisa 21: 1-28.
- RUOKOLAINEN, K., LINNA, A. & TUOMITSO, H. 1997. Use of Melastomataceae and pteridophytes for revealing phytogeographical patterns in Amazonian rain forests. Journal of Tropical Ecology 13:243-256.
- SANAIOTTI, T.M. & MAGNUSSON, W.E. 1995. Effects of annual fires on the production of fleshy fruits eaten by birds in a Brazilian Amazonian savanna. Journal of Tropical Ecology 11:53-65.
- SARMIENTO, G. & MONASTERIO, M. 1985. A critical consideration of the environmental conditions associated with the occurrence of savanna ecosystems in tropical America. In Tropical ecological systems (F.B. Golley & E. Medina, eds.) Springer-Verlag, Berlin, p.223-250.
- SCHOLES, R.J. & ARCHER, S.R. 1997. Tree-grass interactions in savannas. Annual review of Ecology and Systematics 28:517-544.
- SILLBAUER-GOTTESBERGER, I. & GOTTESBERGER, G. 1984. Cerrado-cerradão: a comparison with respect to number of species and growth forms. Phytocoenologia 12:293-303.
- TAKEUCHI, M. 1960. A estrutura da vegetação na Amazônia II. As savanas do norte da Amazônia. Boletim do Museu Paraense Emílio Goeldi nova série Botânica 7:1-14.
- VELOSO, H.P., RANGEL FILHO, A.L. & LIMA, J.C.A. 1991. Classificação da vegetação brasileira adaptada a um sistema universal. IBGE, Rio de Janeiro.
Datas de Publicação
-
Publicação nesta coleção
19 Jun 2008 -
Data do Fascículo
Mar 2008
Histórico
-
Aceito
03 Mar 2008 -
Recebido
22 Dez 2005