Abstract
'Bordô' grapevines (Vitis labrusca) have great relevance to viticulture due to the quality they can impart to wines and juices. However, this cultivar has high variation in yield, ranging from 6 to 11 t ha-1. The use of clones with superior genetic potential related to scions currently marketed may increase crop profitability and revitalize its cultivation. The aim of this study was to evaluate the agronomical responses of twelve clones of the Bordô cultivar selected over a period of 15 years according to yield and quality. The vineyard was planted in 2008. Grape plants were grafted onto '1103 Paulsen' rootstock and trained on vertical shoot positioning. The agronomical evaluations, performed in the 2011, 2012 and 2013 seasons, covered the duration of their phenological cycles, shoot growth, yield per plant, estimated total yield and physicochemical characteristics. Differences were found between clones in terms of phenology, yield components, and berry composition. Clone 6 had the lowest yield, averaging 5.0 t ha-1 whereas clone 13 was the most productive with 14.9 t ha-1. Based on the most productive vineyards in the region (10.8 t ha-1), the adoption of more productive clones can generate an increase in yield of around 38 %.
Vitis labrusca; productivity; quality; composition; anthocyanins
CROP SCIENCE
Agronomic evaluation of 'Bordô' grapevine (Ives) clones
Lidiane Carla Vilanova MiottoI; Renata Vieira da MotaII, * * Corresponding author < rvmota@epamig.br> ; Claudia Rita de SouzaII; Danilo Vieira Cardozo FrançaII; Frederico Alcântara Novelli DiasI; Rodrigo Meirelles de Azevedo PimentelII; Marite Carlin Dal'OstoII; Murillo de Albuquerque ReginaII
IFederal University of Lavras - Dept. of Agriculture, C.P. 3037 - 37200-000 - Lavras, MG - Brazil
IIInstitute of Agronomical Research of Minas Gerais - Tecnological Center of Grape and Wine Research, C.P. 33 - 37780-000 - Caldas, MG - Brazil
ABSTRACT
'Bordô' grapevines (Vitis labrusca) have great relevance to viticulture due to the quality they can impart to wines and juices. However, this cultivar has high variation in yield, ranging from 6 to 11 t ha-1. The use of clones with superior genetic potential related to scions currently marketed may increase crop profitability and revitalize its cultivation. The aim of this study was to evaluate the agronomical responses of twelve clones of the Bordô cultivar selected over a period of 15 years according to yield and quality. The vineyard was planted in 2008. Grape plants were grafted onto '1103 Paulsen' rootstock and trained on vertical shoot positioning. The agronomical evaluations, performed in the 2011, 2012 and 2013 seasons, covered the duration of their phenological cycles, shoot growth, yield per plant, estimated total yield and physicochemical characteristics. Differences were found between clones in terms of phenology, yield components, and berry composition. Clone 6 had the lowest yield, averaging 5.0 t ha-1 whereas clone 13 was the most productive with 14.9 t ha-1. Based on the most productive vineyards in the region (10.8 t ha-1), the adoption of more productive clones can generate an increase in yield of around 38 %.
Keywords: Vitis labrusca, productivity, quality, composition, anthocyanins
Introduction
Amongst the American grapevines cultivated in Brazil, the Bordô cultivar (Vitis labrusca L, also known as Ives) distinguished by rusticity and intense coloration of the berries, can present constant yield loss and annual yield fluctuations due to the decrease in the fruit-set. Orlando et al. (2003) and Norberto et al. (2008) evaluated management techniques that increase vineyard yields. These authors observed that grafting was responsible for a marked increase in yield but the use of different rootstocks or canopy management had little effect on increasing yield, mainly due to poor fruit-set. In an organic production trial under plastic cover in a subtropical highland climate in the south of Brazil, Botelho et al. (2011) observed yields of 1.4 and 2.6 t ha-1 in Bordô vines grafted onto '1103 Paulsen' rootstock in the 2007/2008 and 2008/2009 seasons, respectively, with no difference in yield regardless of the rootstook.
Somatic variation in grapevines plays a major role as a source of phenotypic variation. This variation results from genetic alterations, epigenetic differences or changes in pathogen content that can take place in a single cell, being spread to one apical meristem of a shoot and then to the whole plant by vegetative propagation. Somatic variants, usually observed in cultivars with a long history of cultivation or a large extension of growth, potentially represent a commercial interest if they improve an important agronomical trait or allow for product innovation (Torregrosa et al., 2011).
The phenotypic heterogeneity of the vineyards becomes a new source of variation for genetic improvement of grape cultivars through clonal selection. Such selection may improve productivity as observed by Anderson et al. (2008) with 'Pinot noir' vines in California.
Due to the national importance of common grapes and especially the 'Bordô' grapevine, in 1994 the Institute of Agronomical Research of Minas Gerais, Brazil, (EPAMIG) initiated a program of clonal selection of 'Bordô', in order to identify healthy genetic material with high and constant productivity (Villa et al., 2010). The selection of clones that are more adapted and productive can result in an increase in profitability for the grapevine grower and dissemination of the cultivar. The objective of this study was to evaluate the yield and berry quality of clones of the 'Bordô' grapevine selected for cultivation in subtropical viticultural zones.
Materials and Methods
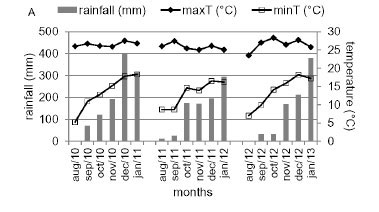
The experiment was carried out during the 2011, 2012 and 2013 seasons in a 158 m2 plot set up in 2008 in Caldas, MG (22°55' S, 46°23' W and altitude of 1,150 m). The climate is of the Cwb type, warm, temperate and rainy, also known as subtropical climate of altitude with mean minimum temperatures of 12 oC and maximum of 25 °C, and 1,500 mm of mean annual rainfall. Viticultural climatic indices for this region in January are as follows: Heliothermal index (HI) 2300; Cold night index (CI) 17 and Dryness index (DI) 170 (Tonietto et al., 2006). These indices classify the region at the ripening stage as a humid, warm with warm nights viticultural climate (Embrapa Uva e Vinho, 2013). Figure 1 presents the climate data during the experimental period.
The vegetative clonal material of the Bordô cultivar grapevine was selected from ungrafted vineyards over 20 years of age. At the first stage, 19 vegetative clonal materials were selected based on the performance of the whole vine and a higher yield. These clones were subjected to sanitary evaluation by the ELISA test to detect the presence of six viruses (Grapevine virus A - GVA; Grapevine virus B - GVB; Grapevine leafroll virus - GFRV, Grapevine fanleaf virus - GFLV and Grapevine leafroll associated virus - GLRaV-1 and -3 (Villa et al., 2010). At this second stage, 12 superior and healthy clones were selected for agronomical evaluation in commercial management conditions adopted by viticulturists in this region. The stocks were produced by the table grafting technique using the '1103 Paulsen' (Vitis berlandieri × V. rupestris) as rootstock. The grapevines were trained in vertical shoot positions on a bilateral Royat Cordon, 1.0 m above ground with shoots positioned upwards by three foliage wires.
Each experimental area consisted of six rows with 40 plants each, spaced at 2.5 m between rows and 1.0 m within rows. The design was completely randomized, consisting of twelve treatments with four replications of five plants for a total of 48 plots. Shoot growth length was obtained by weekly measurements of four shoots previously selected and identified per plot. The evaluations were made with a metric tape, from the beginning of shoot development until summer trimming.
Phenology was evaluated weekly and expressed in days after pruning (DAP) using the following grades: beginning of budburst, when at least two grapevine buds are in the second stage of initial development, which means the moment of scale rupture and appearance of a wooly bud; end of budburst, when 50 % of the gems reach the second stage; flowering start, when anthesis has occurred in at least two grapevine flower buds from different inflorescences; flowering end, when anthesis has occurred in all flower buds, but two from different inflorescences; beginning of veraison, when at least two berries from different clusters have changed color, which means turning green to red; end of veraison; when at least two clusters had berries that were still green.
Mean cluster mass was determined by the production and number of clusters from the three central plants of each treatment replication. To evaluate the number of berries per cluster, berries were counted in 25 clusters for each of the four replications. Estimated yield of each treatment was determined by using mean plant production data multiplied by 4,000, which refers to the adopted plant density. The Ravaz index was calculated by dividing total yield per vine by the pruning weight recorded during the winter following the 2012 season. At harvest composite samples of 100 berries were collected from clusters in its upper, middle lower regions, external and internal side in each replication for the evaluation of must composition (pH, soluble solids content, titratable acidity, and skin phenolic compounds content).
Total Soluble Solids were determined by a hand held digital refractometer and the results were expressed in °Brix. Titratable acidity was determined by titration with 0.1M NaOH, with 1 % phenolphthalein and the results were expressed in g L-1 tartaric acid. Skins were separated manually, dried, weighed and ground in liquid nitrogen. Total monomeric anthocyanins were determined by the pH-differential method (Giusti and Wrolstad, 2000) and total phenolics by the Folin-Ciocalteau method, using gallic acid for the calibration curve (Falcão et al., 2008). Data were submitted to ANOVA and means compared by the Tukey test (p < 0.05).
Results and Discussion
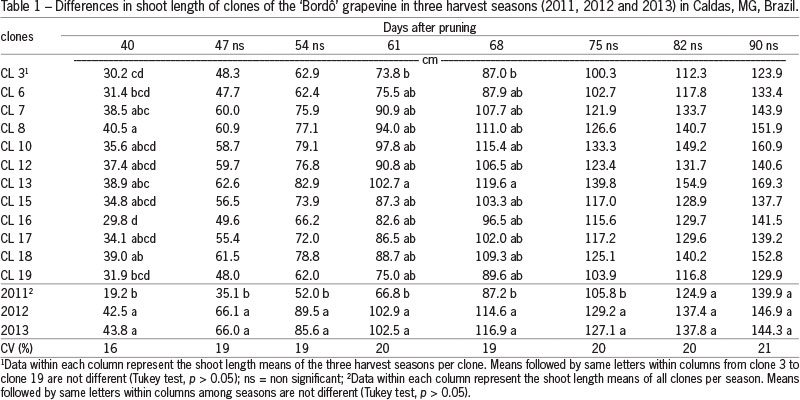
Clone growth vigor was estimated by evaluation of the shoot growth (Table 1). Data from the 2011 season had the slowest shoot growth from 40 to 75 DAP probably because of the more juvenile stage of the vines (Bates et al., 2002). Clone 8 showed the highest shoot growth at 40 DAP which was overtaken by clone 13 in subsequent evaluations. After 75 DAP all clones had similar vigor. The mean values of shoot size from clones observed in the three harvest seasons at 75 (120.7 cm), 82 (133.4 cm) and 90 (143.7 cm) DAP were superior to the mean value of 114.5 cm at 97 DAP in 4 and 5 years old plants from the Bordô cultivar under different training systems (Orlando et al., 2003) or of 120 cm at 84 DAP in vertical shoot position of the Bordô cultivar (Norberto et al., 2008).
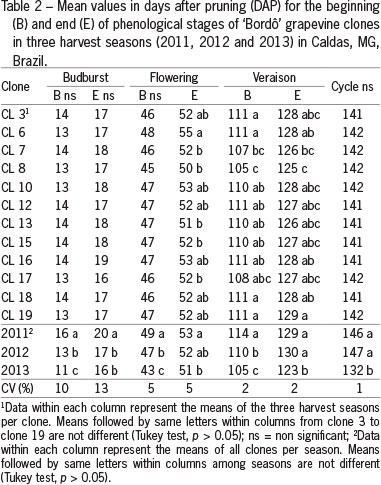
The mean production cycle (from start of budburst to harvest) in the three harvest seasons was 142 days (Table 2), similar to the 149 days observed by Orlando et al. (2003) and superior to the 128 days observed by Botelho et al. (2011). The highest temperatures and lowest rainfall observed in the 2013 growing season may have contributed to the reduction of the production cycle of the clones during this season (Smart and Robinson, 1991).
The phenological stages differed only at the end of flowering and veraison, when clone 8 displayed the shortest growth period and ripened, on average, 5 days earlier in comparison with later clones. Little difference in the duration of phenological phases was also observed for clones 169 and 685 of Cabernet Sauvignon grapes by Burin et al. (2011).
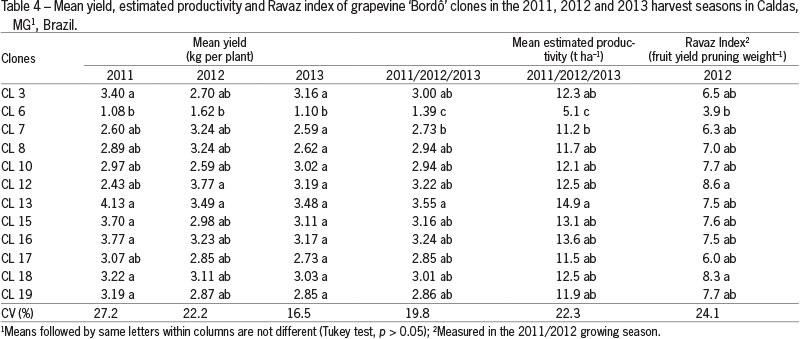
With the exception of clone 6, all clones had mean estimated productivity over 11 t ha-1 and a Ravaz Index over 6, which corresponds to balanced vines (Smart and Robinson, 1991). Clone 13 presented the highest mean yield among treatments, with cluster mass and number of berries per cluster of 112 % and 98 %, respectively, higher than clone 6 and 10 % and 11 %, respectively, higher than the mean of the other clones (Tables 3 and 4). Mean yield for clone 13 was 14.9 t ha-1 which was 28 % superior to the mean of 11.6 t ha-1 of the other clones and 49 % superior to the mean yield (9 to 10 t ha-1) observed by other authors (Norberto et al., 2008;Maia et al., 2013).
Production per plant and estimated production was lower for clone 6 mainly because of the differences observed in cluster mass and number of berries per cluster, since there were no differences in number of clusters, berry size or mass, which points to a poor fruit-set for this clone (Table 3).
Considering the same management for all clones based on the techniques currently adopted by viticulturists, the differences in yield are mainly due to an increase of fruit-set observed in some clones and consequently their higher cluster mass. Santos et al. (2011) also observed an increase in yield with the use of clones selected in the vineyard region of the state of São Paulo. The 'Barberinha' clone resulted in a 68 % yield increase compared to the mean yield of 'Bordô' grapevines. Mean yield of 3.3 kg per plant of clone 'Barberinha' was similar to the mean values of the most productive clones selected in this study (Table 4), which corroborates the idea of increasing yield of 'Bordô' vineyards with clonal selection programs.
Yields of 7.0 and 5.1 t ha-1 of 'Bordô' grapes under the "pergola system" in the harvest seasons of 2002 and 2003, respectively, were reported by Orlando et al. (2003), which were close to the yield of 5.1 t ha-1 obtained by the least productive clone evaluated in this study. According to Orlando et al. (2003), the low productivities are related to the poor fruit-set observed in plants trained in the "pergola system", which can promote more vigor in the plants when compared to the vertical shoot position system.
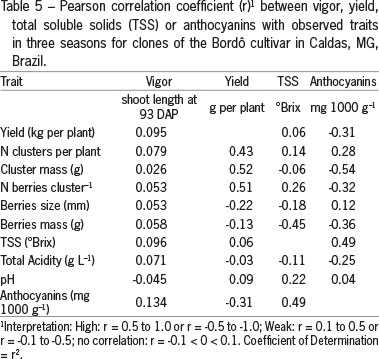
One of the causes of poor fruit-set in grapevines is the excess of vigor (Dry et al., 2010). Under the conditions of this study, however, there was no direct relationship between vigor and fruit set (Table 5). Clones did not differ in shoot length in the three harvest seasons (Table 1), and the Ravaz Index showed that vines were balanced with the exception of clone 6 due to its having the lowest yield (Table 4).
Another factor that causes poor fruit-set is adverse weather conditions with low temperatures and high rainfall during flowering (Jackson, 1991). Data on grape production of Ebadi et al. (1996) showed that temperature effects are related to genotype. A temperature decline from 25°/20 °C (day/night) to 12°/9 °C (day/night) decreased fruit-set between 10 % and 6 % in Chardonnay and between 14 % and 12 % in Syrah grapes. Low temperatures between 12 °C and 9 °C two days before flowering that persist for one week, also affect flower ovule development and give rise to abnormalities (Ebadi et al., 1995).
Rainfall may also lead to poor fruit-set by restraining stamen liberation when the soaked cap is kept attached to the flower. These flowers are called encapsulated, since the stamens cannot disperse the pollen and pollination does not occur (Lebon et al., 2008). Climactic data observed during the period of this study (Figure 1) suggest that there may have been no influence from rainfall on the fruit-set. The 2011 harvest season presented higher incidence of rainfall during flowering with no loss in the number of berries per cluster. However, temperature seems to be related to poor fruit-set. In the 2012 and 2013 harvest seasons, the temperature at the beginning of flowering was below 12 °C, which resulted in a decrease in the number of berries per cluster (Figure 1B, Table 3). Despite the weather conditions, clone 13 showed superior results in the three seasons evaluated.
Total soluble solids content ranged from 15.6 to 17.0 °Brix considering all the harvest seasons (Table 6). These values are in accordance with the 15.3 °Brix found by Orlando et al. (2003), higher than the 13.8 and 14.4°Brix observed by Botelho et al. (2011) but lower than the 19.0° Brix observed at Bswh climate regions by Leão et al. (2010). Clone 8 distinguished itself by presenting the highest values for total soluble solids. The longer period at the ripening stage (in average five days longer than other clones) may explain its higher TSS content at harvest. Burin et al. (2011) also observed slight increases in °Brix values of clone 169 of Cabernet Sauvignon grapes which showed higher veraison in the harvest cycle. There was no correlation between yield and TSS (Table 5). The mean value of must acidity on average, 5.16 g L-1 tartaric acid for all clones and pH 3.3 differed from that observed by Orlando et al (2003) who found must acidity of 6.18 g L-1 and pH 3.15.
With regard to the anthocyanins, values obtained in this study from the 'Bordô' clones varied between 113 and 166 mg 100 g-1 and were similar to the findings of Yang et al. (2009) for the 'Concord' grapes, 80.6 mg cyanidin 3-glucoside 100 g-1. The lower number of berries observed in the clusters of clone 6 could be associated with the elevation of total anthocyanin content of 1,657 mg 1,000 g-1 of berries and total phenolic compounds of 2,181 mg 1,000 g-1 of berries found in the grape skin (Table 7) although correlation data between yield and anthocyanin content showed a weak negative correlation of 10 % (Table 5).
Grapes from 'Bordô' clones possess typical cultivar coloration. The higher yield did not affect negatively the must composition or the anthocyanin contents of the skin, thus clonal selection can be an effective technique for obtaining grapevines with higher yield and berry quality or other interesting agronomic characteristics.
Conclusions
Genetic variability played a major role in productivity. Yield increases resulted in no impairment of berry quality. The use of clone 13, selected in this trial, may contribute to increasing the productivity of 'Bordô' vineyards, with good berry quality and similar production costs of vineyards implanted with non-selected scions.
Acknowledgements
The authors are grateful to the Minas Gerais State Foundation for Research Support (FAPEMIG) and to the Brazilian National Council for Scientific and Technological Development (CNPq) for financial support.
Received November 21, 2013
Accepted April 23, 2014
- Anderson, M.M.; Smith, R.J.; Williams, M.A.; Wolpert, J.A. 2008. Viticultural evaluation of French and Californian Pinot Noir clones grown for production of sparkling wine. American Journal of Enology and Viticulture 59: 181-193.
- Bates, T.R.; Dunst, R.M.; Joy, P. 2002. Seasonal dry matter, starch, and nutrient distribution in 'Concord' grapevine roots. HortScience 37: 313-316.
- Botelho, R.V.; Pavanello, A.P.; Pires, E.J.P.; Terra, M.M.; Müller, M.M.L. 2011. Organic production of red wine grapes under plastic cover in subtropical region of Brazil. Ciência e Agrotecnologia 35: 1186-1195.
- Burin, V.M.; Silva, A.L.; Malinovski, L.I.; Rosier, J.P.; Falcão, L.D.; Bordignon-Luiz, M.T. 2011. Characterization and multivariate classification of grapes and wines of two Cabernet Sauvignon clones. Pesquisa Agropecuária Brasileira 46: 474-481.
- Dry, P.R.; Longbottom, M.L.; McLoughlin, S.; Johnson, T.E.; Collins, C. 2010. Classification of reproductive performance of ten winegrape varieties. Australian Journal of Grape and Wine Research 16: 47-55.
- Ebadi, A.; Coombe, B.G.; May, P. 1995. Fruit-set on small Chardonnay and Shiraz vines grown under varying temperature regimes between budburst and flowering. Australian Journal of Grape and Wine Research 1: 3-10.
- Ebadi, A.; Coombe, B.G.; May, P. 1996. Effect of short-term temperature and shading on fruit-set, seed and berry development in model vines of Vitis vinifera, cvs Chardonnay and Shiraz. Australian Journal of Grape and Wine Research 2: 2-9.
- Embrapa Uva E Vinho. 2013. Methodology of the geoviticulture MCC system. Available at: http://www.cnpuv.embrapa.br/tecnologias/ccm/met.en.html [Accessed Feb 21, 2014]
- Falcão, L.D.; Chaves, E.S.; Burin, V.M.; Falcão, A.P.; Gris, E.F.; Bonin, V.; Bordignon-Luiz, M.T. 2008. Maturity of Cabernet Sauvignon berries from grapevines grown with two different training systems in a new growing region in Brazil. Ciencia e Investigación Agraria 35: 271-282.
- Giusti, M.M.; Wrosltad, R.E. 2000. Characterization and Measurement of Anthocyanins by uv-Visible Spectroscopy. John Willey, New York, NY, USA. (Current Protocols in Food Analytical Chemistry).
- Jackson, D.I. 1991. Environmental and hormonal effects on development of early bunch stem necrosis. American Journal of Enology and Viticulture 42: 290-294.
- Leão, P.C.S.; Cruz, C.D.; Motoike, S.Y. 2010. Genetic diversity of a Brazilian wine grape germplasm collection based on morphoagronomic traits. Revista Brasileira de Fruticultura 32: 1164-1172.
- Lebon, G.; Wojnarowiez, G.; Holzapfel, B.; Fontaine, F.; Vaillant-Gaveau, N.; Clément, C. 2008. Sugars and flowering in the grapevine (Vitis vinifera L.). Journal of Experimental Botany 59: 2565-2578.
- Maia, A.J.; Schwan-Estrada, K.R.F.; Botelho, R.V.; Jardinetti, V.A.; Faria, C.M.D.R.; Batista, A.F.; Costa, W.F. 2013. Bud break and enzymatic activity in buds of grapevine cv. Ives treated with Gallesia integrifolia hydrolate. Acta Physiologiae Plantarum 35: 2727-2735.
- Norberto, P.M.; Regina, M.A.; Chalfun, N.N.J.; Soares, A.M.; Fernandes, V.B. 2008. Influence of the training system in the yield and in the quality of vine fruits 'Folha de Figo' and 'Niágara Rosada' in Caldas, MG. Ciência e Agrotecnologia 32: 450-455 (in Portuguese, with abstract in English).
- Orlando, T.G.S.; Regina, M.A.; Soares, A.M.; Chalfun, N.N.J.; Souza, C.M.; Freitas, G.F.; Toyota, M. 2003. Agronomic characterization of grapevines cultivars (Vitis labrusca L.) in different training systems. Ciência e Agrotecnologia 27: 1460-1469 (in Portuguese, with abstract in English).
- Santos, A.O.; Hernandes, J.L.; Pedro Junior, M.J.; Rolim, G.S. 2011. Plant parameters and microclimatic conditions for wine grapes cultivated under sequential double pruning. Revista Brasileira de Engenharia Agrícola e Ambiental 15: 1251-1256 (in Portuguese, with abstract in English).
- Smart, R.; Robinson, M. 1991. Sun Light into Wine: A Handbook for Winegrape Canopy Management. Winetitles, Adelaide, Australia.
- Tonietto, J.; Vianello, R.L.; Regina, M.A. 2006. Macroclimatic characterization and enological potential of different regions with viticultural inclination in Minas Gerais = Caracterização macroclimática e potencial enológico de diferentes regiões com vocação vitícola em Minas Gerais. Informe Agropecuário 27: 32-55 (in Portuguese).
- Torregrosa, L.; Fernandez, L.; Bouquet, A.; Boursiquot, J-M; Pelsy, F.; Martinez-Zapater, J-M. 2011. Origins and consequences of somatic variation in grapevine. p. 68-92. In: Kole, C., ed. Genomics of fruits and vegetables crops. Science Publishers, Enfield, NH, USA.
- Villa, F.; Regina, M.A.; Alvarenga, A.A.; Pasqual, M.; Stopa, R.A. 2010. Clonal prospection and viruses occurrence in the grapevine cv. 'Folha de Figo' at Caldas, MG, Brazil. Scientia Agraria 11: 155-161 (in Portuguese, with abstract in English).
- Yang, J.; Martinson, T.E.; Liu, R.H. 2009. Phytochemical profiles and antioxidant activities of wine grapes. Food Chemistry 116: 332-339.
Publication Dates
-
Publication in this collection
15 Dec 2014 -
Date of issue
Dec 2014
History
-
Accepted
23 Apr 2014 -
Received
21 Nov 2013