ABSTRACT
This study aimed to investigate the impact of nonionic surfactants on the efficacy of fluorine degradation by Polyporus sp. S133 in a liquid culture. Fluorene was observed to be degraded in its entirety by Polyporus sp. S133 subsequent to a 23-day incubation period. The fastest cell growth rate was observed in the initial 7 days in the culture that was supplemented with Tween 80. The degradation process was primarily modulated by the activity of two ligninolytic enzymes, laccase and MnP. The highest laccase activity was stimulated by the addition of Tween 80 (2443 U/L) followed by mixed surfactant (1766 U/L) and Brij 35 (1655 U/L). UV-vis spectroscopy, TLC analysis and mass spectrum analysis of samples subsequent to the degradation process in the culture medium confirmed the biotransformation of fluorene. Two metabolites, 9-fluorenol (λmax 270, tR 8.0 min and m/z 254) and protocatechuic acid (λmax 260, tR 11.3 min and m/z 370), were identified in the treated medium.
Keywords:
Metabolites; Ligninolytic enzymes; Polyporus sp. S133; Non-ionic surfactants; Solubilization
Introduction
Fungal biotransformation is considered as one of the most important methods for removing pollutant from the ecosystem. In case of organic pollutants, the primary issue that impedes their clearance is their characteristically low solubility in water. The inherent hydrophobic nature of these organic pollutants results in their selective partitioning in the soil matrix such that their bioavailability is seriously compromised. This in turn jeopardizes the success of the bioremediation treatment. There is an urgent need for the development of alternative treatment modalities which will help remove these pollutants from the environment by making them more available for the degradation by microorganisms. Surfactant-mediated biodegradation is one such promising process. This method has the potential to increase the solubility of hydrophobic compounds, such as PAHs, thereby increasing their bioavailability and expediting their removal from the environment.11 Kristanti RA, Hadibarata T. Biodegradation and identification of transformation products of fluorene by ascomycete fungus. Water Air Soil Pollut. 2015;226(12):1-6.
2 Khudhair AB, Hadibarata T, Yusoff ARM, The ZC, Adnan LA, Kamyab H. Pyrene metabolism by new species isolated from soil Rhizocotonia zeae SOL3. Water Air Soil Pollut. 2015;226(186):1-8.-33 Li JL, Chen BH. Surfactant-mediated biodegradation of polycyclic aromatic hydrocarbons. Materials. 2009;2:76-94. Previous studies have shown that depending on the surface of the substance and the type of the surfactant used, the process of surfactant sorption to microorganisms can result in either an increase or a decrease in adhesion.44 Fuchedzhieva N, Karakashev D, Angelidaki I. Anaerobic biodegradation of fluoranthene under methanogenic conditions in presence of surface-active compounds. J Hazard Mater. 2008;153:123-127.
Micelles are the solution aggregates formed by surfactant molecules in water when their concentration is above the critical micelle concentration or CMC. Micelles have dual nature: their hydrophilic outer surface is exposed to the solvent whereas the core is composed of hydrophobic moieties. Several researchers are actively conducting studies in an attempt to investigate the effect of surfactants on microbial transformation of pollutants; all these studies are based on surfactant utilization at concentrations above CMC. Hydrocarbons are known to be partitioned into the hydrophobic core of the micelles.55 Kim IS, Park JS, Kim KW. Enhanced biodegradation of polycyclic aromatic hydrocarbons using nonionic surfactants in soil slurry. Appl Geochem. 2001;16:1419-1429.,66 Zhao BW, Zhu LZ, Li W, et al. Solubilization and biodegradation of phenanthrene in mixed anionic-nonionic surfactant solutions. Chemosphere. 2005;58:33-40. But, it is well established that extent of solubilization is influenced by a multitude of factors such as type and concentration of the surfactant, the interaction between the surfactant and soil, the contact time between the contaminant and soil as well as the hydrophobicity of the compound in question.77 Mulligan CN, Yong RN, Gibbs BF. Surfactant-enhanced remediation of contaminated soil: a review. Eng Geol. 2001;60:371-380.,88 Zhou M, Rhue RD. Screening commercial surfactants suitable for remediating DNAPL source zones by solubilization. Environ Sci Technol. 2000;34:1985-1990.
Fluorene is a three-ring PAH found ubiquitously in the environment.99 Seo JS, Keum YS, Hu Y, et al. Phenanthrene degradation in Arthrobacter sp. P1-1: initial 1,2-, 3,4- and 9,10-dioxygenation, and meta- and ortho-cleavages of naphthalene-1, 2-diol after its formation from naphthalene-1, 2-dicarboxylic acid and hydroxyl naphthoic acids. Chemosphere. 2006;65:2388-2394. It is produced as a result of an incomplete combustion of certain products. Fluorene leaches into the groundwater either as a result of direct contamination from polluted surface waters or indirectly through the soil. Fluorene is not considered as a genotoxic agent but the carcinogenic aspect of its molecular structure has been popularly used as an indicator for the evaluation of PAH-containing pollutants.1010 Kweon O, Kim SJ, Jones RC, et al. A polyomic approach to elucidate the fluoranthene degradative pathway in Mycobacterium vanbaalenii PYR-1. J Bacteriol. 2007;189:4635-4647. White-rot fungi have been frequently employed for the degradation of various organic environmental pollutants. Their exceptional capacity to degrade and biotransform organic contaminants can be attributed to the production of ligninolytic enzymes such as MnP, LiP, laccase, and versatile peroxidase.1111 Hadibarata T, Adnan LA, Yusoff ARM, et al. Microbial decolorization of an azo dye reactive black 5 using white-rot fungus Pleurotus eryngii F032. Water Air Soil Pollut. 2013;224:1595-1604.
12 Kristanti RA, Hadibarata T, Toyama T, et al. Bioremediation of crude oil by white rot fungi Polyporus sp. S133. J Microbiol Biotechnol. 2011;21:995-1000.-1313 Riva S. Laccases: blue enzymes for green chemistry. Trends Biotechnol. 2006;24:219-226. The ligninolytic enzymes are responsible for the depolymerization of PAHs, which results in the production of various types of phenol. They also oxidize substrates in the presence of a suitable co-substrate.1414 Novotny C, Svobodova K, Erbanova P, et al. Ligninolytic fungi in bioremediation: extracellular enzyme production and degradation rate. Soil Biol Biochem. 2004;36:1545-1551. Extensive studies have been undertaken in order to investigate the potential of white-rot fungus to degrade PAHs.1515 Teh ZC, Hadibarata T. Enhanced degradation of pyrene and metabolites identification by Pleurotus eryngii F032. Water Air Soil Pollut. 2014;225(1909):1-8.,1616 Kristanti RA, Toyama T, Hadibarata T, et al. Bioaugmentation involving a bacterial consortium isolated from the rhizosphere of Spirodela polyrhiza for treating water contaminated with a mixture of four nitrophenol isomers. RSC Adv. 2014;4:1616-1621. However, the impact of solubilization of fluorene with single and mixed nonionic surfactants on the degradation rate remains unexplored.
This study attempts to fill the above mentioned lacuna by evaluating the biodegradation of fluorene by the Polyporus species in liquid cultures in the presence of two synthetic surfactants. Our study also quantifies the resultant biomass produced, the extracellular enzymes and the glucose consumption involved in the process. Furthermore, in an attempt to comprehensively study the degradation process, metabolites formed during fluorene degradation have been identified and characterized using a variety of chromatographic techniques. Two nonionic surfactants, Tween 80 and Brij 35, were chosen to study the influence of the surfactant structure on the biodegradation and biotransformation of fluorene by the fungal biomass. Studies have established that nonionic surfactants are more effective on account of their higher adsorption capacity in clay fractions as compared to anionic or cationic surfactants.66 Zhao BW, Zhu LZ, Li W, et al. Solubilization and biodegradation of phenanthrene in mixed anionic-nonionic surfactant solutions. Chemosphere. 2005;58:33-40. The relationship between enzyme activity, glucose consumption, fluorene utilization and biomass production in the presence of Tween 80 and Brij 35 was also established. The data generated by this study will contribute toward defining a new approach for biotransformation of PAHs by improving our understanding about the influence of surfactants in the process. It is hoped that the results of our study will be a significant contribution to the field of biodegradation especially surfactant enhanced biodegradation (Table 1).
Materials and methods
Chemicals and microorganisms
Fluorene and an internal standard, 4-chlorobiphenyl, were obtained from Sigma-Aldrich (Milwaukee, WI). Both were supplied at >97% purity. Nonionic surfactants such as Brij 35 and Tween 80, and extraction solvents such as ethyl acetate and dichloromethane were obtained from Acros Chemicals. Malt extract as well as other chemicals required for liquid culture were procured from Difco (Detroit, USA). All the solutions obtained or prepared utilized analytical grade and high-purity reagents. The fungus used in the study was isolated from Matsuyama, Japan. The Polyporus sp. S133 was cultivated in a 100-mL Erlenmeyer flask containing mineral salt medium (MSM). The composition of the growth medium MSM is given in Table 2. The sterility of MSM was ensured by autoclaving the growth medium prior to use. Fluorene (10 mg/L), Tween 80 and Brij 35 solutions were sterilized by vacuum filtration in order to prevent any conversion of the compounds.1515 Teh ZC, Hadibarata T. Enhanced degradation of pyrene and metabolites identification by Pleurotus eryngii F032. Water Air Soil Pollut. 2014;225(1909):1-8.,1717 Hadibarata T, Kristanti RA. Effect of surfactant and identification of metabolites on the biodegradation of fluoranthene by basidiomycetes fungal isolate Armillaria sp. F022. Bioproc Biosys Eng. 2014;28:920-926. For estimation of enzyme activity, the culture was supplemented with the desired amount of fluorene and incubated for specific time intervals. All experiments were conducted in triplicate.
Enzyme assays and biomass determination
The enzyme analysis of the fungus was conducted as per the protocol described earlier.1818 Hadibarata T, Teh ZC. Optimization of pyrene degradation by white-rot fungus Pleurotus pulmonarius F043 and characterization of its metabolites. Bioproc Biosys Eng. 2014;37:1679-1684. Laccase activity was determined by ABTS oxidation and assayed spectrophotometrically at 420 nm. MnP activity was determined by the oxidation of malonate and dimethoxyphenol in an MnSO4 solution and assayed by monitoring changes in absorbance at 270 nm. The enzyme production was expressed in U L-1. Biomass production was estimated by centrifuging the culture followed by filtration through a Whatman No. 1 filter paper. The biomass of the fungi was expressed in g L-1.
Solubilization test
The solubilization test was performed as described previously.1919 Chan LC, Lee JJ, Park JW. Solubilization of PAH mixtures by three different anionic surfactants. Environ Pollut. 2002;118:307-313.,2020 Zhu L, Chiou CT. Water solubility enhancements of pyrene by single and mixed surfactant solutions. J Environ Sci. 2002;13:491-496. Single nonionic surfactants were used in the following concentrations: 0.1, 0.5, 1, 2, 4 mM. Mixed surfactant solutions (Tween 80 and Brij 35) were prepared at same concentration in a 1:1 ratio.
Instrumental analysis
Agilent 5975E FID GC-MS (DB-1 capillary column; 0.25 µm ID; 0.25 mm diameter; 30 m length) was used to identify and characterize the metabolites present in the incubated culture. As described in our previous study, trimethylchlorosilane was used in the silylation procedure employed for detecting samples containing hydroxyl and carboxyl groups.1818 Hadibarata T, Teh ZC. Optimization of pyrene degradation by white-rot fungus Pleurotus pulmonarius F043 and characterization of its metabolites. Bioproc Biosys Eng. 2014;37:1679-1684. The GC temperature was maintained as follows: initial 70 °C (1 min), increased at the rate of 18 °C min-1 till 150 °C, increased at the rate of 28 °C min-1 till 330 °C, held at 330 °C for 10 min. The injector and interface temperatures were set at 260 °C with a splitless time of 2 min. The injection concentration was 1 µL in the splitless condition and flow rate of the helium was adjusted to 1 mL/min. The EI ionization mode was used with electron energy of 1.3 eV and a mass range of 50-500 amu. The mass spectra of the samples were compared with authentic standard compounds as well as the database library (Wiley 275L).2121 Hadibarata T, Zubir MMFA, Rubiyatno Chuang TZ. Microbial transformation and sorption of anthracene in liquid culture. Bioproc Biosys Eng. 2013;36:1229-1233.
Results and discussion
Solubilization of fluorene
Fig. 1 shows the change in the solubility of fluorene as affected by the addition of single and mixed nonionic surfactants. It was observed that the solubility of fluorene was significantly enhanced by addition of surfactants and that the solubility of the pollutant was directly proportional to the surfactant concentrations. The highest solubility noted in this study was achieved by the addition of Tween 80 (39.4 mg L-1); the addition of mixed solutions resulted in a moderate rise in the solubility, whereas the lowest solubility values were observed upon addition of Brij 35 (20.4 g L-1). It is hypothesized that the addition of fluorene in combination with the nonionic surfactant solution facilitates the formation of mixed micelles which in turn improves the contact between microorganisms and surfactant.
Solubilization of fluorene by single and mixed surfactant: Tween 80; Brij 35; Tween 80-Brij 35.
Biodegradation of fluorene by Polyporus sp. S133
It is evident from the data available in the literature that fluorene is likely to be absorbed onto the fungal biomass surface prior to being accumulated, degraded, and metabolized by the fungus. In order to identify the impact of single and mixed surfactants on fluorene removal, the growth medium of Polyporus sp. was supplemented with the addition of Tween 80, Brij 35, and mixtures of Tween 80 and Brij 35. S133 culture was undertaken in order to estimate both biomass production as well as fluorene degradation. Results indicated that both the cell growth as well as the degradation of fluorene was influenced by the presence of surfactants (Fig. 2). As is shown in Fig. 2, the presence of Tween 80 significantly enhanced the removal of fluorene as well as the biomass production. Fluorene was found to be degraded in entirety by Polyporus sp. S133 after an incubation period of 23 d. The fastest growth rate was experienced in the initial 7 days (0.6 g L-1 per day) in the culture that was supplemented with Tween 80. Tween 80 increased the sorption extent and no distinction was found between surfactant and mycelia. Fig. 2 suggests that the addition of Tween 80 and mixed surfactants enhanced the fluorene degradation and biomass production by Polyporus sp. S133. It was observed after the 23-day incubation period that the cellular production and generation slowly decreased largely because most of the fluorene had already been degraded. The results show that the maximum fluorene degradation was observed for the Tween 80 culture (0.4 mg per day), which 0.2 and 0.27 mg/d compared to was mixed surfactant and Brij 35.
Utilization of fluorene and biomass production in single and mixed surfactant: Tween 80 (A); Brij 35 (B); Tween 80-Brij 35 (C).
Dioxygenase, peroxidase and laccase enzymes play a critical role in the biotransformation of fluorene by Polyporus sp. S133. Enzymes such as 1,2-dioxygenase, 2,3-dioxygenase, MnP, LiP, and laccase were studied with the aim of gaining additional insight into the mechanism of decolorization (Table 3). It was noted that the degradation process was primarily determined by the activity of two ligninolytic enzymes: laccase and MnP. In order to assay the impact of nonionic detergents on the enzyme activity as well as on the fluorene degradation process, activity of the enzyme was plotted against glucose consumption for different concentrations of the surfactants (Fig. 3).
Effect of non-ionic surfactant on enzyme activities and glucose consumption. Tween 80 (A); Brij 35 (B); Tween 80-Brij 35 (C).
It was observed that for all surfactants, the maximum laccase production was seen on day 30 of the incubation; it was also seen that the activity declined significantly at day 15 for Brij 35 and at day 23 for mixed surfactants. Growth medium supplemented with Tween 80 exhibited the highest activity (2443 U L-1) for laccase followed by the mixed surfactant (1766 U L-1) and lastly by Brij 35 (1655 U L-1). A similar analysis of the MnP activity with varying combinations of the surfactant proved that MnP activity decreased after 7-15 days of incubation and the downward trend continued till the end of the experimental period. The glucose consumption was significantly high in the first 7 days of incubation but was observed to decrease slowly after that till the end of the experimental period. The highest consumption of glucose was seen in the growth medium supplemented by Brij 35 (3.8 g L-1) after 30 days of incubation. From this result it can be concluded that when the growth medium is supplemented by Brij 35, glucose is the preferred source of carbon for fungal growth. The function of glucose in the culture was to induce the enzymatic system of the fungus. From the results obtained above, we theorize that the production of ligninolytic enzymes such as laccase and peroxidase was suppressed by addition of fluorene and surfactant. Our results are in accord with a previous study that analyzed PAH degradation by various white-rot fungi. The authors of the study reported that the degradation rates were found to be associated both with the activity of ligninolytic enzymes as well as with the type of surfactant used. The authors also proposed that the enzymes that play an important role in PAHs degradation, namely laccase and MnP, are also implicated in the metabolism of some PAHs. Laccase is also considered to be an important enzyme for oxidizing PAHs in liquid culture.1515 Teh ZC, Hadibarata T. Enhanced degradation of pyrene and metabolites identification by Pleurotus eryngii F032. Water Air Soil Pollut. 2014;225(1909):1-8.,1616 Kristanti RA, Toyama T, Hadibarata T, et al. Bioaugmentation involving a bacterial consortium isolated from the rhizosphere of Spirodela polyrhiza for treating water contaminated with a mixture of four nitrophenol isomers. RSC Adv. 2014;4:1616-1621. The function of MnP in the fluorene removal process is similar to that of laccase in the early weeks of incubation and it sustains till the end of the incubation period.
It was observed that the microorganisms grew at a faster rate when the liquid culture was supplemented with a carbon source such as glucose. Glucose was supplemented into the fungal culture to fulfill the function of a co-metabolic substrate in the degradation process. Previous studies have established that glucose plays an important role in the depolymerization of long chain fatty acids in the biodegradation process. Also, several reports in literature claim that many organic pollutants were more quickly transformed when glucose was supplemented into the culture as a co-substrate for growth.2222 Adosinda M, Martins M, Ferreira IC, et al. Biodegradation of bioaccessible textile azo dyes by Phanerochaete chrysosporium. J Biotechnol. 2001;89:91-98.,2323 Li JL, Chen BH. Solubilization of model polycyclic aromatic hydrocarbons by nonionic surfactants. Chem Eng Sci. 2002;57:2825-2835. Other researchers have proved that for most white-rot fungi, simple carbon sources, such as glucose, provide nutrients that are crucial for the production of extracellular enzymes and biomass production. Although ligninolytic enzymes are produced during the secondary metabolism and are strongly influenced by nutrient limitation, the actual pollutant degradation process occurs after the production of enzymes.2424 Wesenberg D, Kyriakides I, Agathos SN. White-rot fungi and their enzymes for the treatment of industrial dye effluents. Biotechnol Adv. 2003;22:161-187.
Fluorene metabolites
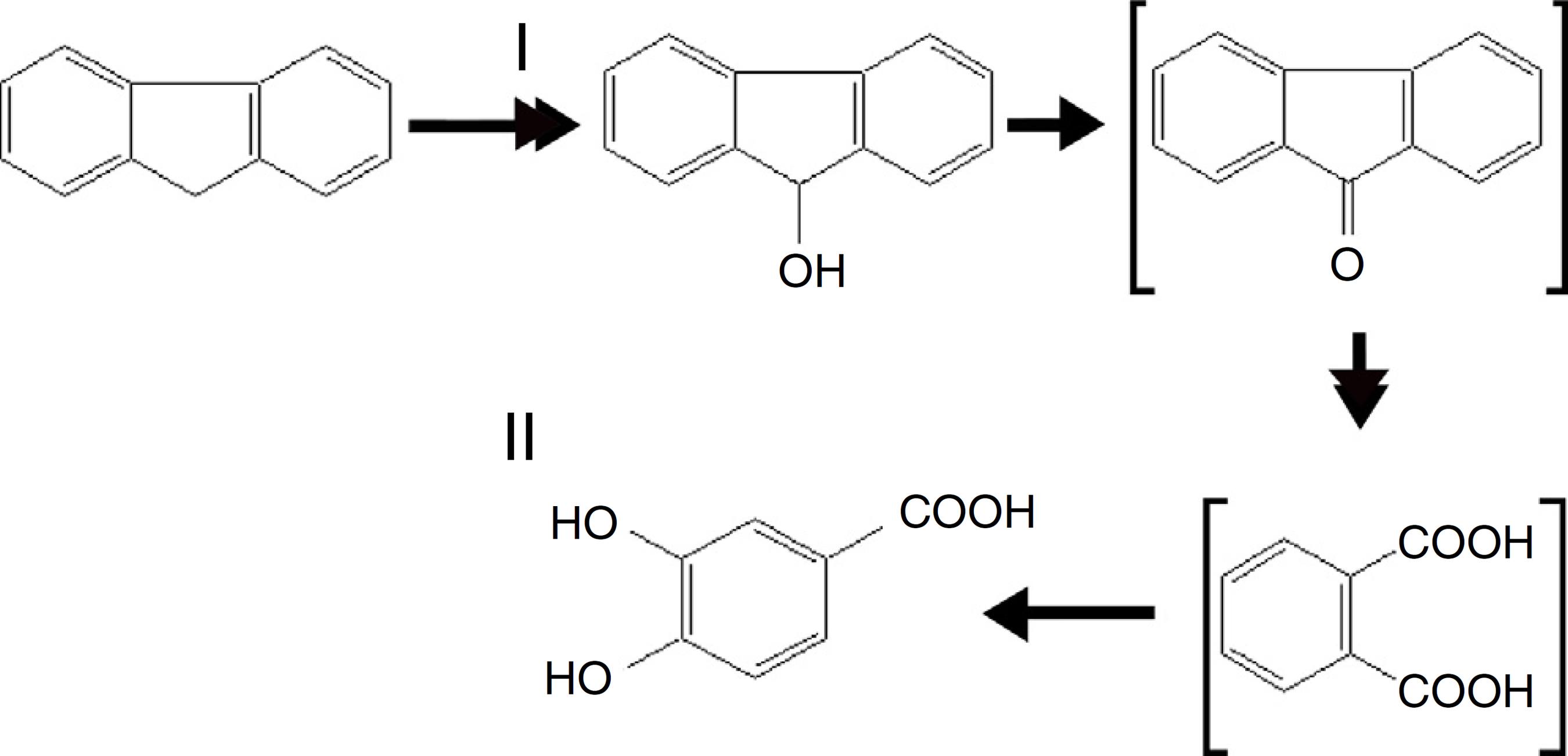
The culture was acidified with HCl in order to stop the fungal growth following which the sample was extracted with ethyl acetate. TLC and UV absorbance of the sample extracts revealed the presence of three fluorine metabolites (Table 4). GC-MS analyses of the ethyl acetate-extracted samples were found to have two major chromatographic peaks (Table 4). Metabolite I (Rf 0.35) exhibited λmax at 270 nm which corresponds to the 9-fluorenol standard. The spectrum of Compound I (tR of 8.0 min) show maxima m/z 254, M+, a fragment ion at m/z 239 (M+-15) representing loss of methyl, as well as the expected fragment ion at m/z 165 (M+-89) which could be accounted for by the sequential loss of OSi(CH3)3, and at 73, 178, and 254. Further metabolite characterization was done by growing Polyporus sp. S133 in a 9-flurenol acid culture. In the culture, we identified metabolite II (tR of 11.3 min), whose spectra was found to have a main ion at m/z 370 (M+), a fragment ion at m/z 355 (M+-15) representing loss of methyl along with expected fragment ions at m/z 181 [M+-89; -OSi(CH3)3], 73 [(CH3)3Si], 165 and 193. Based on the mass spectral data obtained above, we identified compound II as protocatechuic acid.
UV absorbance and mass spectral analysis of the metabolic product produced from fluorene by Polyporus sp. S133.
Polyporus sp. S133 is a white-rot fungus that has the capacity to eliminate and transform many types of PAHs. In this study, we established that the fungus succeeded in producing the ligninolytic enzyme important for the degradation of fluorene. Two major enzymes, laccase and MnP, were presumed to be vital for the transformation of the fluorene. During the laccase-mediated oxidation of PAHs, quinone is formed during the initial step of degradation while 9-fluorenol-1-carboxylic acid is formed at a later stage. In the next step the carboxylic acid compound undergoes a ring cleavage to form 9-fluorenol and protocatechuic acid which is finally converted to tricarboxylic acid. Unfortunately, fluorene quinone and 9-fluorenol-1-carboxylic acid was not identified in any of the extracts obtained in this study (Fig. 4). It is known that certain metabolites produced as a by-product of PAHs metabolism are hazardous in nature. For this reason fragmentary metabolism of fluorene has the potential to pose a grave threat to the ecosystem because the characteristic of intermediate metabolites may actually be more harmful than that of the parent compound.2525 Kim SJ, Kweon O, Jones RC, et al. Complete and integrated pyrene degradation pathway in Mycobacterium vanbaalenii PYR-1 based on systems biology. J Bacteriol. 2007;189:464-472.
26 Lopez Z, Vila J, Minguillon C, et al. Metabolism of fluoranthene by Mycobacterium sp. strain AP1. Appl Microbiol Biotechnol. 2006;70:747-756.
27 Story SP, Parker SH, Hayasaka SS, et al. Convergent and divergent points in catabolic pathways involved in utilization of fluoranthene, naphthalene, anthracene, and phenanthrene by Sphingomonas paucimobilis var. EPA505. J Ind Microbiol Biotechnol. 2001;26:369-382.-2828 Zhong Y, Luan T, Lin L, et al. Production of metabolites in the biodegradation of phenanthrene, fluoranthene and pyrene by the mixed culture of Mycobacterium sp. and Sphingomonas sp. Bioresour Technol. 2011;102:2965-2972.
Proposed pathway of fluorene metabolism by Polyporus sp. S133. Bracket-compounds were unidentified in our culture extract.
Conclusions
From the results presented above it can be confidently concluded that fluorene, a three-ring PAH, was degraded in its entirety by a fungus Polyporus sp. S133 following a 23-day incubation in the liquid culture format. The positive relationship between biomass, enzyme and degradation established that the fluorene bioavailability was significantly increased by surfactant mediated bioaccumulation enhancement especially that by Tween 80. Nonionic surfactants were observed to increase the sorption rate of fluorene on the biomass and thus improve the removal process. Two important ligninolytic enzymes, laccase and MnP, were found to play a significant role in the complex process of fluorene transformation by Polyporus sp. S133. To conclude, our study makes a powerful argument in support of the utility and importance of Tween 80 in the PAHs-polluted soil bioremediation process.
-
Associate Editor: Cynthia Canêdo da Silva
Acknowledgements
A part of this research was financially supported by a Fundamental Research Grant Scheme (FRGS) of Ministry of High Education Malaysia (R.J130000.7809.4F465) and Research University Grant of Universiti Teknologi Malaysia (Q.J130000.2522.10H17).
REFERENCES
-
1Kristanti RA, Hadibarata T. Biodegradation and identification of transformation products of fluorene by ascomycete fungus. Water Air Soil Pollut 2015;226(12):1-6.
-
2Khudhair AB, Hadibarata T, Yusoff ARM, The ZC, Adnan LA, Kamyab H. Pyrene metabolism by new species isolated from soil Rhizocotonia zeae SOL3. Water Air Soil Pollut 2015;226(186):1-8.
-
3Li JL, Chen BH. Surfactant-mediated biodegradation of polycyclic aromatic hydrocarbons. Materials 2009;2:76-94.
-
4Fuchedzhieva N, Karakashev D, Angelidaki I. Anaerobic biodegradation of fluoranthene under methanogenic conditions in presence of surface-active compounds. J Hazard Mater 2008;153:123-127.
-
5Kim IS, Park JS, Kim KW. Enhanced biodegradation of polycyclic aromatic hydrocarbons using nonionic surfactants in soil slurry. Appl Geochem 2001;16:1419-1429.
-
6Zhao BW, Zhu LZ, Li W, et al. Solubilization and biodegradation of phenanthrene in mixed anionic-nonionic surfactant solutions. Chemosphere 2005;58:33-40.
-
7Mulligan CN, Yong RN, Gibbs BF. Surfactant-enhanced remediation of contaminated soil: a review. Eng Geol 2001;60:371-380.
-
8Zhou M, Rhue RD. Screening commercial surfactants suitable for remediating DNAPL source zones by solubilization. Environ Sci Technol 2000;34:1985-1990.
-
9Seo JS, Keum YS, Hu Y, et al. Phenanthrene degradation in Arthrobacter sp. P1-1: initial 1,2-, 3,4- and 9,10-dioxygenation, and meta- and ortho-cleavages of naphthalene-1, 2-diol after its formation from naphthalene-1, 2-dicarboxylic acid and hydroxyl naphthoic acids. Chemosphere 2006;65:2388-2394.
-
10Kweon O, Kim SJ, Jones RC, et al. A polyomic approach to elucidate the fluoranthene degradative pathway in Mycobacterium vanbaalenii PYR-1. J Bacteriol 2007;189:4635-4647.
-
11Hadibarata T, Adnan LA, Yusoff ARM, et al. Microbial decolorization of an azo dye reactive black 5 using white-rot fungus Pleurotus eryngii F032. Water Air Soil Pollut 2013;224:1595-1604.
-
12Kristanti RA, Hadibarata T, Toyama T, et al. Bioremediation of crude oil by white rot fungi Polyporus sp. S133. J Microbiol Biotechnol 2011;21:995-1000.
-
13Riva S. Laccases: blue enzymes for green chemistry. Trends Biotechnol 2006;24:219-226.
-
14Novotny C, Svobodova K, Erbanova P, et al. Ligninolytic fungi in bioremediation: extracellular enzyme production and degradation rate. Soil Biol Biochem 2004;36:1545-1551.
-
15Teh ZC, Hadibarata T. Enhanced degradation of pyrene and metabolites identification by Pleurotus eryngii F032. Water Air Soil Pollut 2014;225(1909):1-8.
-
16Kristanti RA, Toyama T, Hadibarata T, et al. Bioaugmentation involving a bacterial consortium isolated from the rhizosphere of Spirodela polyrhiza for treating water contaminated with a mixture of four nitrophenol isomers. RSC Adv 2014;4:1616-1621.
-
17Hadibarata T, Kristanti RA. Effect of surfactant and identification of metabolites on the biodegradation of fluoranthene by basidiomycetes fungal isolate Armillaria sp. F022. Bioproc Biosys Eng 2014;28:920-926.
-
18Hadibarata T, Teh ZC. Optimization of pyrene degradation by white-rot fungus Pleurotus pulmonarius F043 and characterization of its metabolites. Bioproc Biosys Eng 2014;37:1679-1684.
-
19Chan LC, Lee JJ, Park JW. Solubilization of PAH mixtures by three different anionic surfactants. Environ Pollut 2002;118:307-313.
-
20Zhu L, Chiou CT. Water solubility enhancements of pyrene by single and mixed surfactant solutions. J Environ Sci 2002;13:491-496.
-
21Hadibarata T, Zubir MMFA, Rubiyatno Chuang TZ. Microbial transformation and sorption of anthracene in liquid culture. Bioproc Biosys Eng 2013;36:1229-1233.
-
22Adosinda M, Martins M, Ferreira IC, et al. Biodegradation of bioaccessible textile azo dyes by Phanerochaete chrysosporium J Biotechnol 2001;89:91-98.
-
23Li JL, Chen BH. Solubilization of model polycyclic aromatic hydrocarbons by nonionic surfactants. Chem Eng Sci 2002;57:2825-2835.
-
24Wesenberg D, Kyriakides I, Agathos SN. White-rot fungi and their enzymes for the treatment of industrial dye effluents. Biotechnol Adv 2003;22:161-187.
-
25Kim SJ, Kweon O, Jones RC, et al. Complete and integrated pyrene degradation pathway in Mycobacterium vanbaalenii PYR-1 based on systems biology. J Bacteriol 2007;189:464-472.
-
26Lopez Z, Vila J, Minguillon C, et al. Metabolism of fluoranthene by Mycobacterium sp. strain AP1. Appl Microbiol Biotechnol. 2006;70:747-756.
-
27Story SP, Parker SH, Hayasaka SS, et al. Convergent and divergent points in catabolic pathways involved in utilization of fluoranthene, naphthalene, anthracene, and phenanthrene by Sphingomonas paucimobilis var. EPA505. J Ind Microbiol Biotechnol 2001;26:369-382.
-
28Zhong Y, Luan T, Lin L, et al. Production of metabolites in the biodegradation of phenanthrene, fluoranthene and pyrene by the mixed culture of Mycobacterium sp. and Sphingomonas sp. Bioresour Technol 2011;102:2965-2972.
Publication Dates
-
Publication in this collection
Jul-Sep 2016
History
-
Received
11 July 2015 -
Accepted
22 Dec 2015