Resumos
Foram analisados 95 ninhos de Centris (Heterocentris) terminata Smith, fundados em ninhos-armadilha, entre novembro de 2001 e janeiro de 2003, em dois fragmentos urbanos (CFO-UFBA e PZGV) de Mata Atlântica secundária, em Salvador, BA (13º01’ W e 38º30’ S). O maior número de nidificações ocorreu de dezembro a fevereiro (verão), não havendo ninhos fundados nos meses de agosto, setembro e outubro (inverno-primavera). Emergiram 280 adultos de C. terminata (n = 347 células), sendo 164 machos e 116 fêmeas, resultando em razão sexual de 1: 0,42. Durante esse período, a emergência de machos foi significativamente superior a de fêmeas (c2 = 9,342; gl =10; P < 0,05). Os ninhos foram fundados em ninhos-armadilha de madeira com 6, 8, 10 mm de diâmetro, sendo que 84,2% deles foram construídos com 8 e 10 mm. Cada ninhos possui de uma a sete células, dispostas em série em série linear e partições construídas com uma mistura de areia e resina ou óleo. O macho é significativamente menor que a fêmea, sendo que esta última emerge a partir das primeiras células construídas. A mortalidade de imaturos foi baixa, ocorrendo em 14,1% das células (n= 49), sendo 13,0% por falhas no desenvolvimento e 1,2% por parasitas Coelioxys sp. (Hymenoptera: Megachilidae) e Tetraonyx sp. (Coleoptera: Meloidae). Na área estudada, fatores meteorológicos, especialmente a pluviosidade, mostraram-se mais determinantes da abundância temporal do que inimigos naturais. O longo período de atividade anual, abundância local e adequação aos ninhos-armadilha sugerem bom potencial de C. terminata para manejo visando a polinização de plantas nativas e cultivadas.
Abelha solitária; ninho-armadilha; biologia de nidificação; arquitetura de ninhos
Ninety-five nests of Centris (Heterocentris) terminata Smith were collected in trap-nests, during November/2001 and January/2003, at two fragments (PZGV e CFO-UFBA) of secondary Atlantic Forest, in Salvador, Bahia State (13º01’ W e 38º30’ S). The highest nest frequencies occurred from December to February (summer), with no nests foundations from August to October (winter - early spring). Two-hundred eight adults emerged from 347 brood cells, being 164 males and 116 females (1: 0.42). During the study period sex ratio was male biased (c2 = 9.342; gl = 10; P < 0.05). C. terminata nested in holes with diameters 6, 8, 10 mm, but 84,2% were constructed in 8 and 10 mm. nests had one to seven cells arranged in a linear series with the cell’s partitions built with a mixture of sand and resin or oil. Male is significantly smaller than female, which emerges from the first cells constructed. Immature mortality occurred in 14.1% of brood cells (n = 49), of which 13.0% were due fail in development and 1.2% due to parasitism of Coelioxys sp. (Hymenoptera: Megachilidae) e Tetraonyx sp. (Coleoptera: Meloidae). In the study site, weather, mainly pluviosity, rather than natural enemies influenced seasonal population abundance. The long period of nesting activity, local abundance and usage of trap nests, suggest the potential of C. terminata for management aiming at pollination of native and cultivated plants.
Solitary bee; nesting biology; nest architecture
ECOLOGY, BEHAVIOR AND BIONOMICS
Ninhos de Centris (Heterocentris) terminata Smith (Hymenoptera: Apidae, Centridini) em fragmentos de Mata Atlântica secundária, Salvador, BA
Trap-nests used by Centris (Heterocentris) terminata Smith (Hymenoptera: Apidae, Centridini) at secondary Atlantic Forest fragments, in Salvador, Bahia State
Patrícia DrummontI; Fabiana O. da SilvaII, III; Blandina F. VianaI
ILab. Biologia e Ecologia de Abelhas (Labea), Depto. Zoologia, Instituto de Biologia, Univ. Federal da Bahia Rua Barão de Geremoabo s/n, Campus Universitário de Ondina, 40170-110, Salvador, BA
IIFaculdade Tecnologia e Ciências (FTC). Av. Luís Viana Filho, 8812, Paralela, 41820-785 Salvador, BA
IIIFaculdades Jorge Amado. Av. Luís Viana Filho, 6775, Paralela, 41745-130, Salvador, BA
RESUMO
Foram analisados 95 ninhos de Centris (Heterocentris) terminata Smith, fundados em ninhos-armadilha, entre novembro de 2001 e janeiro de 2003, em dois fragmentos urbanos (CFO-UFBA e PZGV) de Mata Atlântica secundária, em Salvador, BA (13o01 W e 38o30 S). O maior número de nidificações ocorreu de dezembro a fevereiro (verão), não havendo ninhos fundados nos meses de agosto, setembro e outubro (inverno-primavera). Emergiram 280 adultos de C. terminata (n = 347 células), sendo 164 machos e 116 fêmeas, resultando em razão sexual de 1: 0,42. Durante esse período, a emergência de machos foi significativamente superior a de fêmeas (c2 = 9,342; gl =10; P < 0,05). Os ninhos foram fundados em ninhos-armadilha de madeira com 6, 8, 10 mm de diâmetro, sendo que 84,2% deles foram construídos com 8 e 10 mm. Cada ninhos possui de uma a sete células, dispostas em série em série linear e partições construídas com uma mistura de areia e resina ou óleo. O macho é significativamente menor que a fêmea, sendo que esta última emerge a partir das primeiras células construídas. A mortalidade de imaturos foi baixa, ocorrendo em 14,1% das células (n= 49), sendo 13,0% por falhas no desenvolvimento e 1,2% por parasitas Coelioxys sp. (Hymenoptera: Megachilidae) e Tetraonyx sp. (Coleoptera: Meloidae). Na área estudada, fatores meteorológicos, especialmente a pluviosidade, mostraram-se mais determinantes da abundância temporal do que inimigos naturais. O longo período de atividade anual, abundância local e adequação aos ninhos-armadilha sugerem bom potencial de C. terminata para manejo visando a polinização de plantas nativas e cultivadas.
Palavras-Chave: Abelha solitária, ninho-armadilha, biologia de nidificação, arquitetura de ninhos
ABSTRACT
Ninety-five nests of Centris (Heterocentris) terminata Smith were collected in trap-nests, during November/2001 and January/2003, at two fragments (PZGV e CFO-UFBA) of secondary Atlantic Forest, in Salvador, Bahia State (13o01 W e 38o30 S). The highest nest frequencies occurred from December to February (summer), with no nests foundations from August to October (winter - early spring). Two-hundred eight adults emerged from 347 brood cells, being 164 males and 116 females (1: 0.42). During the study period sex ratio was male biased (c2 = 9.342; gl = 10; P < 0.05). C. terminata nested in holes with diameters 6, 8, 10 mm, but 84,2% were constructed in 8 and 10 mm. nests had one to seven cells arranged in a linear series with the cells partitions built with a mixture of sand and resin or oil. Male is significantly smaller than female, which emerges from the first cells constructed. Immature mortality occurred in 14.1% of brood cells (n = 49), of which 13.0% were due fail in development and 1.2% due to parasitism of Coelioxys sp. (Hymenoptera: Megachilidae) e Tetraonyx sp. (Coleoptera: Meloidae). In the study site, weather, mainly pluviosity, rather than natural enemies influenced seasonal population abundance. The long period of nesting activity, local abundance and usage of trap nests, suggest the potential of C. terminata for management aiming at pollination of native and cultivated plants.
Key Words: Solitary bee, nesting biology, nest architecture
O gênero Centris é tipicamente tropical com espécies divididas em 12 subgêneros (Snelling 1984, Ayala 1998). As informações acumuladas sobre esse gênero indicam grande diversidade de hábitos de nidificação, com as maiores diferenças aparecendo em nível subgenérico (Coville et al. 1983). Algumas espécies de diferentes subgêneros escavam seus ninhos no solo enquanto outras nidificam em madeiras que podem ser troncos ou galhos, enquanto poucas espécies constroem ninhos expostos (Roubik 1989). Há, ainda, aquelas espécies que nidificam em cavidades naturais preexistentes (orifícios existentes em árvores, galerias feitas em madeira por besouros ou outros insetos, gomos de bambu, ou até mesmo ninhos abandonados por outras espécies), neste último grupo encontram-se os subgêneros Hemisiella, Heterocentris e Xanthemisia (Coville et al. 1983).
Essa característica comportamental exibida por esses três subgêneros tem facilitado o estudo das espécies solitárias, pois através da disponibilização de cavidades artificiais previamente preparadas (os chamados ninhos-armadilha) é possível obter ninhos de fêmeas que eventualmente usam essas cavidades quando dispostas no campo (Krombein 1967, Camillo et al. 1993). Uma vez recolhidos os ninhos podem ser estudados quanto às características estruturais e comportamentais.
No Brasil, observa-se a predominância de espécies do gênero Centris em número de ninhos fundados em ninhos-armadilha em diferentes habitats (Morato & Campos 2000, Viana et al. 2001, Aguiar et al. 2005). Embora, alguns estudos (p.ex., Vinson & Frankie 1988) revelem uma tendência geral das espécies de Centris em nidificar em habitats florestados, ninhos de algumas das espécies desse gênero (p. ex., Centris tarsata Smith) são comumente encontrados em habitats quentes e iluminados (Pérez-Maluf 1993, Silva et al. 2001, Viana et al. 2001).
Apesar da ampla distribuição e predominância, aspectos da biologia de nidificação de algumas espécies de Centris são ainda pouco conhecidos (Morato et al. 1999, Jesus & Garófalo 2000, Silva et al. 2001). Dada a grande variabilidade nos hábitos de nidificação das espécies de Centris (Vinson et al. 1987, 1991; Frankie et al. 1993) e considerando que as informações sobre a arquitetura dos ninhos das espécies do gênero são ainda escassas (Morato et al. 1999), fazem-se necessários estudos abordando tais aspectos, em um número maior de espécies e em ambientes diferentes.
Um estudo realizado por Morato et al. (1999) na Amazônia Central registrou a ocorrência Centris terminata Smith, em locais abertos e iluminados. A espécie foi predominante em estudos com ninho-armadilha realizado em fragmentos urbanos de Mata Atlântica secundária, em Salvador, indicando tolerância à perda de hábitat de floresta (Viana et al. 2006).
A despeito da abundância local de C. terminata, informações sobre a biologia de nidificação dessa espécie são mencionadas apenas por Morato et al. (1999). Desse modo, este trabalho visa contribuir para o conhecimento da biologia de C. terminata, descrevendo a estrutura do ninho e os inimigos naturais associados. O número de nidificações foi relacionado com o período do ano e a razão sexual com o diâmetro e o período de nidificações nos ninhos-armadilha coletados.
Material e Métodos
Os ninhos de C. terminata analisados foram coletados em fragmentos urbanos de Mata Atlântica secundária no Parque Zoobotânico Getúlio Vargas e Campus Federação/Ondina da Universidade Federal da Bahia, localizados no bairro de Ondina (13o 01 S e 38o 31 W) em Salvador, BA. O Parque Zoobotânico Getúlio Vargas e o Campus Federação/Ondina da UFBA ocupam áreas de 302.175,9 m2 e 240.000 m2, respectivamente, e estão circundadas por uma zona densamente urbanizada, desprovida de coberturas vegetais de tamanho significativo.
A fisionomia dominante no Parque Zoobotânico Getúlio Vargas é a de Mata Atlântica secundária enquanto a área do Campus Federação/Ondina da UFBA é um mosaico formado por vegetação herbácea heterogênea, mata secundária arbustiva arbórea, bosque de mangueiras (Mangifera indica L.), bambuzal (Bambusa vulgaris Schrad. ex J.C. Wendl.). Em ambas ocorrem jardins com plantas ornamentais e campo de gramíneas, podendo haver árvores espaças, prédios e estradas. O clima local é tropical quente e úmido (Af na tipologia climática de Köppen) com pluviometria anual média em torno de 1800 mm e temperatura média anual de 25 oC, com poucas variações (Conder 1994).
Cada ninho-armadilha constituiu-se de duas peças de madeira (15 x 30 x 150 mm) unidas com fita adesiva e com perfuração longitudinal de 100 mm. Para a observação ao longo de toda a cavidade, foi feito um corte da fita adesiva em uma das laterais da peça. Os ninhos-armadilha foram agrupados em blocos contendo 20 unidades unidas por uma borracha, sendo quatro ninhos-armadilha de cada diâmetro de cavidade (6, 8, 10, 15 e 20 mm).
Em cada área foram selecionadas sete árvores aleatoriamente, colocando-se em cada árvore dois blocos com ninhos-armadilha suspensos, em posição horizontal, com cordão de náilon preso ao galho a 1,5 m e 8 m de altura, cada. Nos blocos suspensos a 8 m de altura o cordão de náilon passou por uma roldana de metal para facilitar a inspeção periódica dos ninhos. No total foram disponibilizados 560 ninhos-armadilha dispostos em 26 blocos, nas duas áreas.
Quinzenalmente, os ninhos-armadilha foram inspecionados com auxílio de otoscópio, entre novembro de 2001 e janeiro de 2003. Os ninhos-armadilha ocupados foram removidos e substituídos por outros vazios. No laboratório, os ninhos-ocupados foram mantidos em caixas de vidro (15 x 15 x 12 cm) cobertas com tela, até a emergência do adulto. Os espécimes estão depositados na coleção de referência do Laboratório de Biologia e Ecologia de Abelhas (LABEA), do IB-UFBA.
Os ninhos foram abertos, descritos e fotografados e as principais características estruturais foram medidas. O volume, largura e comprimento das células, espessura das divisórias e o espaço existente entre a última célula construída e a entrada do ninho, foram medidos em 80 ninhos. O volume das células foi calculado considerando-se o volume do cilindro. Em todos os ninhos coletados foram registradas a razão sexual, o número de células, presença ou não de células vestibulares, características das partições, tipo de material utilizado para construção e aprovisionamento do ninho, e mortalidade. O tamanho dos adultos foi avaliado pela largura máxima da cabeça (LC), com o auxílio de um paquímetro digital.
Os parâmetros medidos nos ninho fundados foram submetidos às seguintes análises: (1) comparou-se o comprimento, largura e volume das células onde emergiram machos e fêmeas (teste "t" student); (2) o tamanho relativo dos machos e das fêmeas (teste "t" de Student e o teste de Mann-Whitney); (3) o comprimento, largura e volume das células com emergência de machos e fêmeas, nos diferentes diâmetros (5) a proporção de machos e fêmeas nas diferentes classes de diâmetro (ANOVA seguida do teste de comparações múltiplas (paramétrico) de Tukey-Kramer); (5) A razão sexual foi determinada como a proporção de fêmeas adultas, calculada pela divisão do número de fêmeas pelo total de adultos obtidos durante o período de amostragem; (6) a influência do diâmetro do orifício sobre a razão sexual e (7) se a distribuição das células que originaram machos e fêmeas foi ao acaso (análise de contingência - Tabela de Contingência).
Resultados
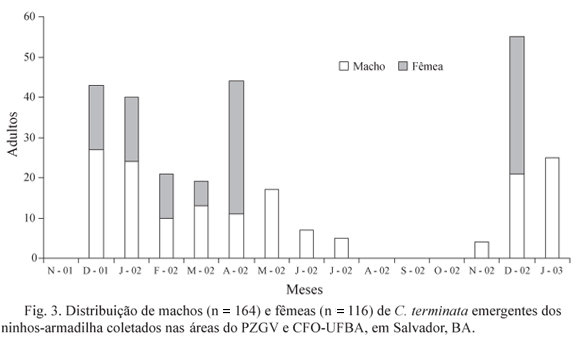
Distribuição temporal dos ninhos. Durante o período de amostragem foram fundados 95 ninhos de C. (Heterocentris) terminata, sendo 77 (81,1%) no Parque Zoobotânico Getúlio Vargas e 18 (19,0%) no Campus Federação/Ondina-UFBA. O total de 87,4% das nidificações (n = 83 ninhos) ocorreu em ninhos-armadilha dispostos a 1,5 m, enquanto 12,6% (n = 12 ninhos) ocorreram a 8 m de altura, tanto no Parque Zoobotânico Getúlio Vargas quanto no Campus Federação/Ondina-UFBA. O maior número de nidificações ocorreu nos meses de dezembro, janeiro e fevereiro, não havendo ninhos fundados nos meses de agosto, setembro e outubro (Fig. 1).
Ninhos-armadilha utilizados e estrutura do ninho. C. terminata construiu ninhos nos diâmetros de 6, 8 e 10 mm, sendo 16 (16,8%), 49 (51,6%) e 30 (31,6%) ninhos, respectivamente. Não houve nidificações nas classes de diâmetro de 15 mm e 20 mm. A número total de células construídas nas cavidades com diâmetro de 6, 8 e 10mm foram, respectivamente, 43, 181 e 123.
As células possuíam a parte posterior côncava e apresentaram-se sempre dispostas em série linear (Fig. 2). A presença de células vestibulares foi registrada em 8,4% (n = 8) dos ninhos. O material utilizado para a construção das células constitui-se de uma mistura de areia e uma substância aglutinante oleosa, cuja composição não foi determinada. As divisórias das células tinham espessura entre 1,20 mm e 3,60 mm (0,80 ± 0,20, n = 80 ninhos). Nos ninhos analisados, o comprimento das células variou entre 9,50 mm e 32,65 mm (15,13 ± 3,03); e o volume variou entre 0,25 mm3 e 1,86 mm3 (0,74 ± 0,27). O espaço existente entre a última célula construída e a entrada do ninho variou entre 0,0 mm e 3,75 mm (1,50 ± 1,10) de comprimento.
O volume das células nas três classes de diâmetro foi significativamente diferente (kw = 40,426; P < 0,0001). No entanto, a diferença ocorreu entre os diâmetros de 6 e 10 mm e entre 8 e 10 mm, não sendo significativa a diferença entre os diâmetros 6 e 8 mm. A largura também foi significativa entre as três classes de diâmetro (kw = 74,176; P < 0,0001) e todas diferiram entre si. O comprimento das células nas três classes de diâmetro foi o único parâmetro, entre as principais medidas das células, que não mostrou diferenças significativas entre os diâmetros de 6, 8 e 10 mm (Tabela 1).
Emergência dos adultos. Foram construídas nos ninhos analisados 347 células de cria (n = 95 ninhos), sendo que a partir de 18 delas os adultos já haviam emergido na ocasião das inspeções. Durante o monitoramento em laboratório emergiram 280 adultos de C. (Heterocentris) terminata, dos quais 164 eram machos (58,6%) e 116 eram fêmeas (41,4%), resultando na razão sexual de 1: 0,42. Durante os meses amostrados, a emergência de machos foi significativamente superior à emergência de fêmeas (c2 = 9,342; gl = 10; P < 0,05), sendo que em maio, junho, julho e novembro/02 e janeiro/03 houve emergência exclusiva de machos. Apenas em abril e dezembro/02 a emergência de fêmeas foi superior à de machos (Fig. 3).
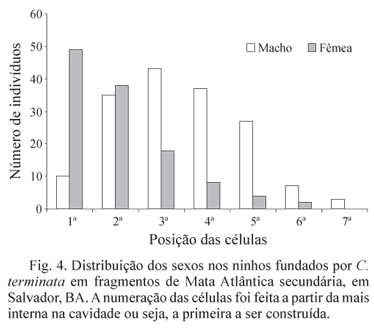
O período transcorrido entre a coleta dos ninhos e emergência dos adultos foi, em média, de 32,3 ± 10,38 dias. A partir da 1ª célula emergiram predominantemente fêmeas, enquanto que a partir da 2ª célula construída emergiram fêmeas e machos, em proporções semelhantes. Nas demais células a emergência de machos predominou em relação à emergência de fêmeas (Fig. 4). A diferença na proporção de machos e fêmeas entre as classes de diâmetro ocupadas foi considerada significativa apenas entre 6 e 8 mm (q = 7.424; P < 0,05), não sendo observada diferença entre 6 e 10 mm (q = 3.558; P > 0,05) e 8 e 10 mm (q = 3,865; P > 0,05) (Tabela 2).
A largura da cabeça dos machos variou de 3,08 a 4,71 mm, enquanto que a largura da cabeça das fêmeas variou de 3,37 a 5,12 mm sendo, portanto, menores que as fêmeas (Fig. 5). O tamanho dos machos e das fêmeas diferiu significativamente nos três diâmetros de ninhos analisados: 6 mm (t = 3,043; gl = 32; P = 0,0047), 8 mm (U = 4684.0; P < 0,0001) e 10 mm (t = 4,859; gl = 67; P < 0,0001).
Mortalidade de imaturos e inimigos naturais associados. Em seis ninhos (6,1%) não houve emergência de adultos de C. terminata, e em quatro ninhos, ocorreu falha parcial de desenvolvimento. A mortalidade total observada nos ninhos de C. terminata representou 14,1% (n = 49 células). Em 13,0% a mortalidade foi atribuída a falha de desenvolvimento de origem endógena e em 1,4% (n = 4 células) foi causada por parasitismo. A presença de cleptoparasita do gênero Coelioxys sp. (Hymenoptera: Megachilidae) causou mortalidade em quatro células e, em uma célula a mortalidade foi causada por Tetraonyx sp. (Coleoptera: Meloidae).
Discussão
A ocorrência de variações anuais na freqüência de ninhos produzidos é comum entre espécies solitárias (Pereira et al. 1999, Viana et al. 2001, Aguiar et al. 2005). O ciclo reprodutivo de C. terminata foi longo e, considerando o período transcorrido entre a coleta dos ninhos e a emergência do adulto, devem ocorrer de 4 a 5 gerações por ano, não tendo sido evidenciada a ocorrência de diapausa.
Durante o período de estudo, a atividade de nidificação em C. terminata foi sazonal, com maior freqüência de nidificações durante o verão e início do outono, sugerindo que mudanças climáticas influenciam a atividade de nidificação de C. terminata, na área de estudo. Durante o inverno, a alta pluviosidade não favoreceu a ocupação das armadilhas, visto que os mesmos não estavam protegidos da chuva e a madeira retém umidade; além disso, a pluviosidade influencia a baixa produção de flores, reduzindo a oferta potencial de recursos para essa espécie.
Estudos utilizando a técnica de ninhos-armadilha demonstram a predominância de Centris em número de ninhos coletados, em diferentes regiões do Brasil (Camillo et al. 1995, Morato & Campos 2000), sendo também constatado para a região nordeste (Viana et al. 2001, Aguiar & Martins 2002, Aguiar et al. 2005). O número de ninhos de C. terminata obtidos neste estudo assemelha-se ao encontrado em levantamentos de espécies desse gênero em ambientes florestados (Morato et al. 1999, Jesus & Garófalo 2000). Os dados revelam que fragmentos urbanos, embora pequenos, são capazes de abrigar populações de abelhas e contribuir para a manutenção de espécies encontradas em área de floresta nativa.
A despeito do predomínio de arbóreas no Parque Zoobotânico Getúlio Vargas, o maior número de ninhos fundados a 1,5 m de altura corrobora a tendência encontrada para abelhas e vespas solitárias utilizando ninhos-armadilha (Krombein 1967, Peres-Maluf 1993), já que havia igual quantidade de cavidades disponíveis a 1,5 m e 8 m de altura. A maior freqüência de nidificações registrada para a área do Parque Zoobotânico Getúlio Vargas, onde as condições do sub-bosque são mais estáveis (Viana et al. 2006), evidenciam que C. terminata é sensível a modificações na estrutura do ambiente, e que as condições microclimáticas do sub-bosque são importantes para a seleção dos locais de nidificação.
Os diâmetros preferencialmente ocupados por C. terminata no presente estudo estão incluídos na variação de diâmetro (4,8; 9,5 e 12,7 mm) dos ninhos-armadilha utilizados por Morato et al. (1999). O predomínio de ninhos em cavidades de 8 mm e 10 mm de diâmetro é uma tendência comum em Centris, tendo sido registrada para espécies de Hemisiella (Silva et al. 2001, Aguiar & Garófalo 2004, Aguiar et al. 2005) e Heterocentris (Frankie et al. 1993, Jesus & Garófalo 2000) que utilizam cavidades pré-existentes.
Vários aspectos podem influenciar a variação no diâmetro da cavidade nidificada como: tamanho das fêmeas, disponibilidade de cavidades adequadas (Krombein 1967), gasto energético para a coleta do material de construção do ninho (Jesus & Garófalo 2000, Aguiar & Garófalo 2005) e a abundância local de recursos tróficos para provisionamento das células (Aguiar & Garófalo 2004). A continuidade dos estudos sobre a biologia de nidificação de Centris poderá elucidar se existe um fator preponderante para cada espécie ou se predomina uma interação entre os mesmos.
A arquitetura dos ninhos de C. terminata assemelha-se ao padrão descrito para C. terminata por Morato et al. (1999) em fragmentos e em mata contínua, na Amazônia Central, bem como outras espécies de Centris (Coville et al. 1983, Pereira et al. 1999, Jesus & Garófalo 2000, Silva et al. 2001). Dentre as semelhanças destacam-se as características das células (disposição linear, formato e revestimento) e o material usado para a construção das partições em ninhos de C. dichrootricha Moure (Morato et al. 1999), C. analis (Jesus & Garófalo 2000) e C. tarsata (Silva et al. 2001, Aguiar & Garófalo 2004, Aguiar et al. 2005) e C. trigonoides (Aguiar et al. 2006).
Ao contrário de outras espécies (Jesus & Garófalo 2000, Aguiar et al. 2005), a variação observada em número, largura e comprimento de células construídas nos ninhos de C. terminata, não é influenciada pelo aumento do diâmetro da cavidade do ninho-armadilha. Em nenhum dos ninhos fundados a cavidade foi completamente utilizada indicando que o comprimento das cavidades disponíveis também não foi o principal determinante do número ou dimensões das células construídas por C. terminata. Morato et al. (1999) também não observaram diferença significativa na largura e no comprimento das células.
A forte correlação entre o sexo do emergente e a posição da célula no ninho de C. terminata corrobora resultados obtidos diversos autores com abelhas solitárias (Krombein 1967, Jesus & Garófalo 2000, Silva et al. 2001). A menor duração do período de desenvolvimento dos machos permite a emergência das fêmeas a partir das células mais internas (Jesus & Garófalo 2000).
A razão sexual encontrada foi semelhante à encontrada por Morato et al. (1999) para essa espécie. Ao longo do período de estudo, a forte tendência à produção de machos foi compensada pela maior produção de fêmeas em gerações seguintes, neste estudo evidenciada nos meses de abril e dezembro/02, confirmando a hipótese de Torchio & Terpedino (1980).
Outro fator também relacionado à razão sexual é o tamanho do diâmetro das cavidades (Pérez-Maluf 1993), onde o esperado é que nos diâmetros menores sejam produzidos preferencialmente machos, talvez, devido ao menor tamanho corporal dos mesmos, e um aumento na produção de fêmeas com o aumento do diâmetro do orifício. Essa tendência não foi confirmada neste estudo, como também não foi confirmada por Silva et al. (2001) em ninhos de C. tarsata.
Nos ninhos de C. terminata a mortalidade de imaturos resultantes de falhas no desenvolvimento foi mais relevante do que o ataque de parasitas. Contudo, a mortalidade registrada neste estudo foi bem menor em relação a outros estudos com abelhas e vespas solitárias empregando ninhos-armadilha (Krombein 1967).
Neste estudo, o número reduzido de células vestibulares (8,4%) e parasitas encontrados nos ninhos não permitem relacionar a presença de células vestibulares à redução na taxa de parasitismo nos ninhos, como sugerido por Pérez-Maluf (1993). Os parasitas encontrados nos ninhos de C. terminata, no presente estudo, Coelioxys Latreille e Tetraonyx sp., também foram observados em ninhos da mesma espécie na Amazônia Central (Morato et al.1999). Coelioxys também foi registrado em ninhos de C. tarsata (Aguiar & Garófalo 2004), sendo responsável por 11,3% da mortalidade das células e C. analis (Jesus & Garófalo 2000), causando mortalidade em 30% das células.
A ocorrência dessas populações em fragmentos urbanos revela sua resistência a perturbações ambientais e capacidade de uso de diversos materiais de nidificação. Na área estudada, fatores meteorológicos, especialmente pluviosidade, mostraram-se mais determinantes da abundância temporal do que inimigos naturais. O longo período de atividade anual, abundância local e adequação aos ninhos-armadilha sugerem bom potencial de C. terminata para manejo visando a polinização de plantas nativas e cultivadas. Embora muitas características biológicas revelem-se semelhantes em nível genérico, estudos mais detalhados sobre a biologia de nidificação de C. terminata em áreas naturais e urbanas, são necessários para melhor entendimento desses aspectos.
Agradecimentos
Ao PROCES/CAPES (Programa de Capacitação para o ensino superior) e ao CNPq pelas bolsas concedidas a primeira e terceira autora, respectivamente. Ao professor Pe. Jesus Santiago Moure pela identificação da espécie estudada. À equipe do Laboratório de Biologia e Ecologia de Abelhas (LABEA) pelo auxílio nos trabalhos de campo. Aos revisores anônimos que contribuíram com criticas e sugestões para a melhoria da versão final do manuscrito.
Received 21/III/06. Accepted 09/X/07.
- Aguiar, C.M.L. & C.A. Garófalo. 2004. Nesting biology of Centris (Hemisiella) tarsata Smith (Hymenoptera, Apidae, Centridini). Rev. Bras. Zool. 21: 477-486.
- Aguiar, C.M.L., C.A. Garófalo & G.F. Almeida. 2005. Abelhas (Hymenoptera, Apoidea) que nidificam em ninhos-armadilha em áreas de floresta semi-decídua e caatinga, Bahia, Brasil. Rev. Bras. Zool. 22: 1030-1038.
- Aguiar, C.M.L., C.A. Garófalo & G.F. Almeida. 2006. Nesting biology of Centris (Hemisiella) trigonoides Lepeletier (Hymenoptera, Apidae, Centridini). Rev. Bras. Zool. 23: 323-330.
- Aguiar, C.M.L & C.F. Martins. 2002. Abelhas e vespas solitárias em ninhos-armadilha na Reserva Biológica Guaribas (Mamanguape, Paraíba, Brasil). Rev. Bras. Zool. 19: 101-116.
- Ayala, R.B. 1998. Sistematica de los taxas supraespecificos de lãs abejas de la tribu Centridini (Hymenoptera: Anthophoridae). Tesis, Doctorado. Universidad Nacional Autônoma de México, México, 280p.
- Camillo, E., C.A. Garófalo & J.C. Serrano. 1993. Hábitos de nidificação de Melitoma segmentaria, Centris collaris, Centris fuscata e Paratetrapedia gigantea (Hymenoptera, Anthophoridae). Rev. Bras. Entomol. 37: 145-156.
- Camillo, E., C.A. Garófalo, J.C. Serrano & G. Muccillo. 1995. Diversidade e abundância sazonal de abelhas e vespas solitárias em ninhos armadilhas (Hymenoptera, Apocrita, Aculeata). Rev. Bras. Entomol. 39: 459-470.
- Conder. 1994. Informações básicas dos municípios baianos: Região Metropolitana de Salvador. 1 v. il., tab., gráf. 267p.
- Coville, R.E., G.W. Frankie & S.B. Vinson. 1983. Nests of Centris segregata (Hymenoptera: Anthophoridae) with a review of nesting habitats of the genus. J. Kans. Entomol. Soc. 56: 109-122.
- Frankie, G.W., L.E. Newstrom, S.B. Wilson, S.B. Vinson & J.F. Barthell. 1993. Nesting habitat preferences of selected Centris bees species in Costa Rican dry forest. Biotropica 25: 322-33.
- Jesus, B.M.V. & C.A. Garófalo. 2000. Nesting behaviour of Centris (Heterocentris) analis (Fabricius) in southeastern Brazil (Hymenoptera, Apidae, Centridini). Apidologie 31: 503-515.
- Krombein, K.V. 1967. Trap-nesting wasps and bees: Life, histories, nests and associates. Washington, Smithsonian Press, 570p.
- Michener, C.D. 2000. The bees of the world. Baltimore, John Hopkins University Press, 913p.
- Morato, E.F. & L.A.O. Campos. 2000. Efeitos da fragmentação florestal sobre vespas e abelhas solitárias em uma área da Amazônia Central. Rev. Bras. Zool. 17: 429-444.
- Morato, E.F., M.V.B. Garcia & L.A.O. Campos. 1999. Biologia de Centris Fabricius (Hymenoptera, Anthophoridae, Centridini) em matas contínuas e fragmentadas na Amazônia Central. Rev. Bras. Zool. 16: 1213-1222.
- Pereira, M., C.A. Garófalo, E. Camillo & J.C. Serrano. 1999. Nesting biology of Centris (Hemisiella) vittata Lepeletier in southeastern Brazil (Hymenoptera, Apidae, Centridini). Apidologie 30: 327-338.
- Pérez-Maluf, R. 1993. Biologia de vespas e abelhas solitárias, em ninhos-armadilhas, em Viçosa - MG. Dissertação de mestrado, Universidade Federal de Viçosa, Viçosa, 87p.
- Roubik, D.W. 1989. Ecology and natural history of tropical bees. New York, Cambridge University Press, 514p.
- Siegel, S. & N.J. Castellan Jr. 1988. Nonparametric statistics for the behavior sciences. New York, McGraw-Hill, 350p.
- Silva, F.O., B.F. Viana & E.L. Neves. 2001. Biologia e arquitetura de ninhos de Centris (Hemisiella) tarsata Smith (Hymenoptera: Apidae: Centridini). Neotrop. Entomol. 30: 541-545.
- Snelling, R.R. 1984. Studies on the taxonomy and distribuition American Centridine bees (Hymenoptera: Anthophoridae). Contrib. Sci. (Los Angel.) 347: 1-69.
- Torchio, P.F. & V.J. Terpedino. 1980. Sex ratio, body size and seasonality in solitary bees, Osmia lignaria propinqua Cresson (Hymenoptera: Megachilidae). Evolution 34: 993-1003.
- Viana, B.F., A.M.C. Melo & P.D. Drumond. 2006. Variação na estrutura do habitat afetando a composição de abelhas e vespas solitárias em remanescentes florestais urbanos de Mata Atlântica no Nordeste do Brasil. Sitientibus ser. Ciênc. Biol. 6: 282-295.
- Viana, B.F., F.O. Silva & A.M.P. Kleinert. 2001. Diversidade e sazonalidade de abelhas solitárias (Hymenoptera: Apoidea) em dunas litorâneas no Nordeste do Brasil. Neotrop. Entomol. 30: 245-251.
- Vinson, S.B. & G.W. Frankie. 1988. A comparative study of the ground nests of Centris flavifrons and Centris aethiocesta (Hymenoptera: Anthophoridae). Entomol. Exp. Appl. 49: 181-187.
- Vinson, S.B. & G.W. Frankie. 1991. Nest variability in Centris aethytera (Hymenoptera: Anthophoridae) in response to nesting site conditions. J. Kans. Entomol. Soc. 64: 156-162.
- Vinson, S.B., G.W. Frankie & R.E. Coville. 1987. Nesting habitats of Centris flavofasciata Friese (Hymenoptera: Apoidea: Anthophoridae) in Costa Rica. J. Kans. Entomol. Soc. 64: 249-263.
Datas de Publicação
-
Publicação nesta coleção
14 Jul 2008 -
Data do Fascículo
Jun 2008
Histórico
-
Aceito
09 Out 2007 -
Recebido
21 Mar 2006