Resumos
Utilizando-se fragrâncias artificiais como iscas, foram estudadas a riqueza, composição, abundância e sazonalidade de espécies de Euglossina em duas áreas (mata e duna) na Área de Proteção Ambiental da Barra do Rio Mamanguape, PB, entre agosto de 2002 e julho de 2004. As abelhas foram atraídas com chumaços de papel absorvente contendo cada uma das fragrâncias: acetato de benzila, beta ionona, escatol, eucaliptol, eugenol e vanilina, e capturadas com rede entomológica. Foram coletados 3.132 machos de nove espécies de Euglossina. Euglossa cordata (L.) e Eulaema nigrita Lepeletier ocorreram durante todo o período de estudo e foram as espécies mais abundantes na mata e a primeira foi a espécie mais abundante na duna. Quanto à composição, as áreas de Mata Atlântica do estado da Paraíba foram mais similares entre si, o mesmo ocorrendo para as áreas de dunas da Paraíba e Bahia. Na área de mata, as espécies apresentaram maior sazonalidade, sendo mais abundantes no período mais seco, destacando-se E. cordata. Na duna, as espécies distribuíram-se homogeneamente nos períodos seco e chuvoso.
Abelha das orquídeas; Euglossa; Eulaema; Exaerete
By using artificial fragrances as baits, we studied richness, composition, abundance and seasonality of Euglossina species in two areas (forest and dune) in the Environmental Protection Area of the Mamanguape River Bar, State of Paraiba, Brazil, between August 2002 and July 2004. Bees were attracted with wads of absorbent paper containing each of the fragrances: benzyl acetate, ionone beta, skatole, eucalyptol, eugenol and vanillin, and captured with insect net. We collected a total of 3,132 males of nine species of Euglossina. On both areas, Euglossa cordata (L.) and Eulaema nigrita Lepeletier were present throughout the year and were the most abundant species in the forest and the first one was the most abundant specie at the dune. Concerning to the composition, the Atlantic Rainforest areas in Paraíba State were more similar among themselves, the same occurring to the dune areas in Paraíba and Bahia States. In the forest, Euglossina species showed higher seasonality, being more abundant during the drier period, specially E. cordata. At the dune, species were homogeneously distributed in the dry and rainy periods.
Orchid bee; Euglossa; Eulaema; Exaerete
ECOLOGY, BEHAVIOR AND BIONOMICS
Composição e sazonalidade de espécies de Euglossina (Hymenoptera: Apidae) em mata e duna na Área de Proteção Ambiental da Barra do Rio Mamanguape, Rio Tinto, PB
Composition and seasonality of Euglossina Species (Hymenoptera: Apidae) in forest and dune in the Environmental Protection Area of the Mamanguape River Bar, PB
Rodrigo C.A.P. FariasI; Maria C. Madeira-da-SilvaII; Maria H. Pereira-PeixotoI; Celso F. MartinsI
ILab. Entomologia, Departamento de Sistemática e Ecologia, Centro de Ciências Exatas e da Natureza Univ. Federal da Paraíba (UFPB), Cidade Universitária, 58051-900, João Pessoa, PB rodrigoento@gmail.com; helenametal@yahoo.com.br; cmartins@dse.ufpb.br
IICentro Federal de Educação Tecnológica da Paraíba CEFET-PB. Av. 1o de Maio, 720 58015-430, João Pessoa, PB; madeiradasilva@yahoo.com.br
RESUMO
Utilizando-se fragrâncias artificiais como iscas, foram estudadas a riqueza, composição, abundância e sazonalidade de espécies de Euglossina em duas áreas (mata e duna) na Área de Proteção Ambiental da Barra do Rio Mamanguape, PB, entre agosto de 2002 e julho de 2004. As abelhas foram atraídas com chumaços de papel absorvente contendo cada uma das fragrâncias: acetato de benzila, beta ionona, escatol, eucaliptol, eugenol e vanilina, e capturadas com rede entomológica. Foram coletados 3.132 machos de nove espécies de Euglossina. Euglossa cordata (L.) e Eulaema nigrita Lepeletier ocorreram durante todo o período de estudo e foram as espécies mais abundantes na mata e a primeira foi a espécie mais abundante na duna. Quanto à composição, as áreas de Mata Atlântica do estado da Paraíba foram mais similares entre si, o mesmo ocorrendo para as áreas de dunas da Paraíba e Bahia. Na área de mata, as espécies apresentaram maior sazonalidade, sendo mais abundantes no período mais seco, destacando-se E. cordata. Na duna, as espécies distribuíram-se homogeneamente nos períodos seco e chuvoso.
Palavras-Chave: Abelha das orquídeas, Euglossa, Eulaema, Exaerete
ABSTRACT
By using artificial fragrances as baits, we studied richness, composition, abundance and seasonality of Euglossina species in two areas (forest and dune) in the Environmental Protection Area of the Mamanguape River Bar, State of Paraiba, Brazil, between August 2002 and July 2004. Bees were attracted with wads of absorbent paper containing each of the fragrances: benzyl acetate, ionone beta, skatole, eucalyptol, eugenol and vanillin, and captured with insect net. We collected a total of 3,132 males of nine species of Euglossina. On both areas, Euglossa cordata (L.) and Eulaema nigrita Lepeletier were present throughout the year and were the most abundant species in the forest and the first one was the most abundant specie at the dune. Concerning to the composition, the Atlantic Rainforest areas in Paraíba State were more similar among themselves, the same occurring to the dune areas in Paraíba and Bahia States. In the forest, Euglossina species showed higher seasonality, being more abundant during the drier period, specially E. cordata. At the dune, species were homogeneously distributed in the dry and rainy periods.
Key Words: Orchid bee, Euglossa, Eulaema, Exaerete
As abelhas Euglossina estão distribuídas exclusivamente na Região Neotropical, sendo encontradas na faixa compreendida entre 30o de latitude norte e 32o de latitude sul, com algumas espécies passando além dos trópicos (Rebêlo 2001). A diversidade é maior em florestas úmidas, com poucas espécies ocorrendo em savanas e florestas de galeria (Dressler 1982). Ramírez et al. (2002) listam o total de 184 espécies mas de acordo com Roubik & Hanson (2004) esse número pode ultrapassar 200. Estimativas indicam que as euglossinas compreendem cerca de 25% de todas as espécies de abelhas que residem em qualquer floresta de baixa a média elevação nos úmidos trópicos americanos (Roubik & Hanson 2004), e o Brasil é o segundo país com maior diversidade de Euglossina, sendo a Colômbia o primeiro (Ramírez et al. 2002).
Os machos de Euglossina coletam substâncias aromáticas nas plantas, dentre as quais as orquídeas se destacam com centenas de espécies visitadas. Um grande passo para o melhor conhecimento das Euglossina foi dado quando as fragrâncias passaram a ser sintetizadas em laboratório. Dodson et al. (1969) descobriram como atrair um grande número de machos a iscas contendo compostos aromáticos. Williams & Whitten (1983) apresentaram uma lista de fragrâncias que atraem machos de Euglossina, classificadas de acordo com a sua atratividade e as espécies de orquídeas que as produzem. A utilização dessas substâncias como iscas permitiu a realização de levantamentos faunísticos, abrindo caminho para os mais diversos estudos em toda a Região Neotropical (Rebêlo 2001). No Brasil, já foram realizados estudos sobre a diversidade e ecologia de Euglossina em vários ambientes. No Nordeste, podemos citar os trabalhos de Bezerra & Martins (2001) em remanescentes de Mata Atlântica em João Pessoa, PB, Brito & Rêgo (2001) numa floresta secundária no Maranhão, Neves & Viana (1999) em matas de Caatinga na Bahia, Rebêlo & Cabral (1997) numa área de Cerrado na Baixada Litorânea do Maranhão, Souza et al. (2005) em remanescente de Mata Atlântica em Mamanguape, PB, e Viana et al. (2002) em dunas litorâneas na Bahia.
A coleta de fragrâncias pelos machos tem atraído o interesse em trabalhos de campo a fim de se monitorarem a abundância, a dinâmica e outros aspectos das populações de Euglossina, tanto local como regionalmente (Roubik 2004). O padrão diário e sazonal de visitação às iscas pode ser semelhante àquele observado na planta visitada. Assim, pode-se obter informações preciosas sobre os padrões de diversidade, densidade populacional, variação sazonal e geográfica, longevidade, estruturas de idade, além de sua especificidade pelos compostos químicos.
A subtribo Euglossina é um dos grupos de abelhas neotropicais mais estudados, utilizado como modelo biológico em trabalhos relacionados à fisiologia, coevolução planta-polinizador, dinâmica de populações, mimetismo além de aspectos ecoetológicos (Roubik 1989). Isto, associado a sua grande diversificação ecológica e taxonômica, abundância e importância em alguns ecossistemas, faz desses insetos candidatos a indicadores das qualidades ambientais de áreas naturais ou de conservação (Brown 1991).
Este trabalho teve como objetivos estudar a composição, riqueza, abundância e a sazonalidade de espécies de Euglossina em mata e duna na Área de Proteção Ambiental da Barra do Rio Mamanguape e realizar comparações com estudos prévios regionais visando verificar a relação entre a diversidade local e a similaridade entre tipos principais de hábitats.
Material e Métodos
Área de estudo. A Área de Proteção Ambiental da Barra do Rio Mamanguape (APA), com 14.640 ha, foi criada pelo Decreto Federal no. 924 de 10 de setembro de 1993. Localiza-se no litoral norte da Paraíba, a cerca de 80 km de João Pessoa, entre as coordenadas geográficas de 6o45 e 6o50" S e 34o56" e 35o05" W, na mesorregião da Zona da Mata (Brasil 1993). O clima da região é tropical e chuvoso (Am, na classificação de Köeppen) e a temperatura média anual varia entre 24oC e 27oC (Governo do Estado da Paraíba 1985).
A Barra do Rio Mamanguape é uma das áreas prioritárias para conservação (Ministério do Meio Ambiente 2002). A baixada litorânea, onde se situa a APA, é formada por terrenos sedimentares que podem atingir até 10 m e estão representados por praias, dunas, restingas, manguezais e formações recifais (Carvalho 1982). Os fragmentos de mata presentes na APA encontram-se em estado secundário de crescimento, cuja vegetação é de porte médio (árvores de até 10 m de altura) e estão espalhados desde áreas litorâneas até locais circundados por canaviais. Aqueles fragmentos que não estão sob influência das atividades canavieiras estão adjacentes a áreas habitadas por seres humanos e, por isso, sofrem alterações com a abertura de clareiras para o plantio de espécies utilizadas na alimentação bovina ou aquelas de importância comercial.
Amostragem. Os pontos de coleta consistiram de uma duna na formação vegetal aberta de restinga localizada às margens do estuário do Rio Mamanguape (6o 46 21" S e 34o 55 36" W) e uma clareira artificial em um fragmento de mata localizado a cerca de 200 m da praia do Oiteiro (6o 51 36" S e 34o5508" W).
As coletas foram realizadas no período de agosto de 2002 a julho de 2004, mensalmente, das 8:00h às 14:00h, e em dois dias consecutivos, sendo um dia para cada área. Utilizou-se o método descrito por Rebêlo & Garófalo (1991), que consiste no uso de chumaço de papel absorvente amarrado com barbante e pendurado nas ramagens das árvores, e cada chumaço sendo embebido de uma fragrância. Foram utilizadas seis fragrâncias artificiais para atrair os machos: acetato de benzila, beta-ionona, escatol, eucaliptol, eugenol e vanilina. As iscas foram instaladas a cerca de 1,5 m do solo, ao abrigo da insolação direta e distando pelo menos 1,5 m uma da outra. As iscas mais visitadas eram reabastecidas de fragrâncias a cada 2h. Ao pousarem na isca as abelhas eram capturadas com rede entomológica e agrupadas por horário de coleta e iscas visitadas. A partir de janeiro de 2004 as abelhas não foram mais sacrificadas. Após serem capturadas com a rede, eram anestesiadas por resfriamento, marcadas com canetas Edding 791 e liberadas. Todavia, indivíduos de espécies consideradas raras para as áreas de estudo foram coletados.
A cada hora era a feita a leitura da temperatura. Os dados da precipitação mensal foram fornecidos pelo Laboratório de Meteorologia, Recursos Hídricos e Sensoriamento Remoto da Paraíba (LMRS-PB) e os períodos de chuva e estiagem foram estabelecidos de acordo com os valores mensais de precipitação nos meses em que foram feitos este estudo.
Em laboratório, as abelhas foram montadas, etiquetadas e colocadas em estufa por 48h, conforme método padrão de coleções entomológicas. Os exemplares encontram-se depositados na Coleção Entomológica do Departamento de Sistemática e Ecologia (DSEC) da Universidade Federal da Paraíba.
Análise dos dados. Foram feitas análises faunísticas considerando-se a abundância relativa, riqueza de espécies, sazonalidade e relações com fatores abióticos. Os resultados obtidos foram comparados com os de outros levantamentos faunísticos de Euglossina realizados em áreas de Mata Atlântica, Dunas Litorâneas, Matas Ciliares sob a influência de Caatinga e Cerrado da região Nordeste do Brasil. Para a análise dos dados foi calculado o índice de similaridade de Sorenson, utilizando o programa Bio-Dap. A fim de comparar a similaridade entre hábitats, a matriz de similaridade foi transformada em uma matriz de distâncias, subtraindo-se 1 de cada índice, e elaborado um dendrograma de distâncias através do método UGMA (método não ponderado de agrupamento aos pares por médias aritméticas) utilizando o programa Statistica 6.0. O índice de Sorenson foi utilizado porque considera para a ocorrência de espécies apenas as concordâncias positivas e as discordâncias, ignorando as concordâncias negativas (duplos zeros), e também porque não leva em conta a abundância, minimizando diferenças de esforço amostral nos diferentes estudos realizados por autores diversos. Também foram calculados índices de correlação de Spearman (rs) com fatores abióticos (pluviosidade e temperatura), através do programa BioEstat (versão 2.0). A variação na abundância de machos de Euglossina ao longo do ano nas duas áreas foi analisada levando-se em conta o número total de indivíduos, os valores mensais de precipitação e as médias de temperatura dos horários de coleta. Porém, tendo em vista que, em ambas as áreas, as duas espécies mais abundantes totalizaram mais de 70% do total da amostra, a análise da sazonalidade também foi feita individualmente para cada uma das espécies mais abundantes nas duas áreas.
Resultados e Discussão
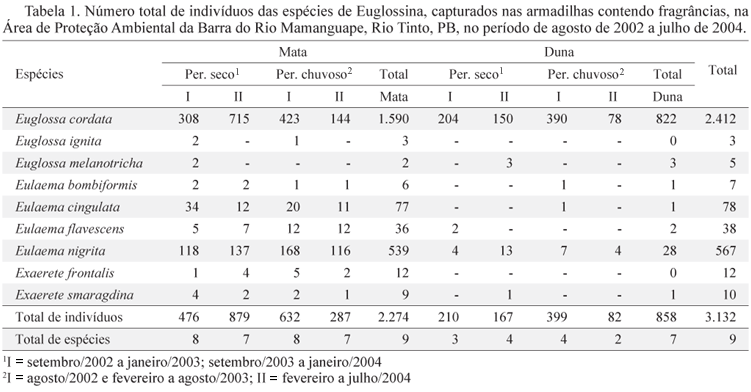
Composição, riqueza e abundância de Euglossina. Nas duas áreas de estudo foram amostrados 3.132 machos de Euglossina, sendo 2.274 na mata, pertencentes a três gêneros e nove espécies, e 858 na restinga, pertencentes a três gêneros e sete espécies. As duas áreas tiveram em comum sete espécies. Foram exclusivas da mata as espécies Euglossa ignita Smith e Exaerete frontalis (Guérin). E. cordata foi a mais abundante tanto na mata quanto na duna. E. nigrita e E. cingulata foram abundantes apenas na mata (Tabela 1).
Foi observada maior similaridade na composição de espécies entre as áreas de mata da APA e a mata de João Pessoa, formando a seguir um grupo com a mata da Reserva Biológica Guaribas, todas localizadas no estado da Paraíba (Fig. 1). Do mesmo modo, as duas áreas de dunas, da Paraíba e da Bahia, também apresentaram maior similaridade em sua composição, além de riqueza menor. Esses dois grupos formaram um grupo maior com cerca de 80% de similaridade, ao qual se ligam, sucessivamente, com menor semelhança, as áreas de cerrado e mata do estado do Maranhão e da caatinga baiana.
Como esperado, as áreas de Mata Atlântica foram mais similares entre si, o mesmo ocorrendo para as áreas de dunas. A área de mata ciliar localizada na caatinga baiana apresenta menor riqueza e similaridade, provavelmente devido às diferenças climáticas e composição florística desse ambiente quando comparada com as áreas de mata do litoral. Por outro lado, as áreas maranhenses sofrem grande influência da fauna amazônica, apresentando maior riqueza e composição distinta das outras áreas nordestinas (Rebêlo & Cabral 1997, Rebêlo 2001, Silva e Rebêlo 2002).
Nos levantamentos realizados nos estados da Paraíba e Bahia utilizando-se fragrâncias têm-se amostrado entre sete e 13 espécies, em ambientes de duna, restinga, mangue e floresta atlântica (e.g. Neves & Viana 1997, Bezerra & Martins 2001, Souza et al. 2005), enquanto em Minas Gerais e no Rio de Janeiro, em áreas de Mata Atlântica, ocorrem, respectivamente, ca. 20 e 21 espécies (Nemésio & Silveira 2006, Tonhasca Jr. et al. 2002). Todavia, estudos na Amazônia e América Central revelam comunidades mais ricas com ca. 50 espécies (Roubik 2004). Portanto, o presente estudo mostra uma riqueza e composição de espécies semelhantes às de outras localidades do Nordeste (com exceção daquelas com influência amazônica, como o Maranhão), inclusive com a predominância de espécies comuns e de ampla distribuição. Nesse sentido, nota-se a grande abundância de E. cordata e E. nigrita na mata e da primeira em duna. A grande abundância de E. nigrita, espécie considerada de áreas abertas ou degradadas (Peruquetti et al. 1999), pode estar relacionada com a "qualidade ambiental", considerando o tamanho de fragmentos e efeito de borda (Nemésio & Silveira 2007). Entretanto, como sua abundância é elevada mesmo em áreas relativamente conservadas (Bezerra & Martins 2001), para testar essa hipótese são necessários estudos comparativos com áreas em excelente estado de conservação e história de baixa influência antrópica. Desse modo, a hipótese alternativa de que a dominância de E. nigrita reflete a estrutura populacional original dessas áreas não pode ser descartada. Por outro lado, é interessante notar que na área de duna a abundância de E. nigrita foi baixa e a espécie dominante foi E. cordata, fato também observado nas dunas de Abaeté, Bahia, por Viana et al. (2002). Talvez, nas altas temperaturas desse ambiente, E. nigrita tenha maior dificuldade de termoregulação do que as abelhas menores de E. cordata.
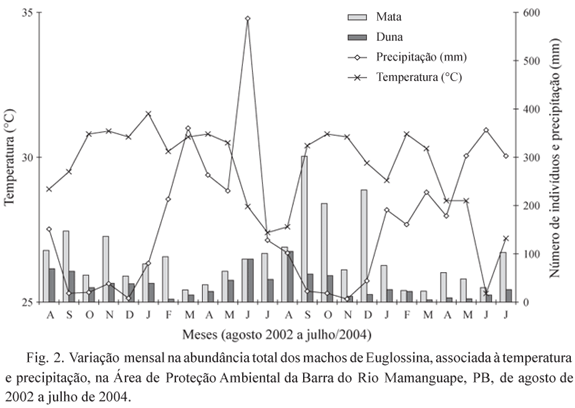
Sazonalidade. Na área de mata houve maior abundância de indivíduos entre os meses de agosto/2002 e fevereiro/2003 e entre julho/2003 e dezembro/2003, ambos os períodos correspondendo ao fim da estação chuvosa, estação seca e início da estação chuvosa seguinte (Tabela 1 e Fig. 2). E. cordata esteve presente em todos os meses, porém foi mais abundante no período seco. E. nigrita também esteve presente em todos os meses e foi mais abundante nos meses de novembro/2002, fevereiro/2003, janeiro/2004 e julho/2004, não mostrando relação clara da abundância com as estações. E. cingulata apresentou maior abundância nos meses de novembro/2002, janeiro/2003 e fevereiro/2003 (meados da estação seca e início da estação chuvosa). As demais espécies foram coletadas esparsamente, não demonstrando nenhuma relação aparente com as estações chuvosa e seca.
Na duna, E. cordata (que corresponde a 95,9% dos indivíduos coletados) foi mais abundante entre agosto/2002 e janeiro/2003 e entre maio/2003 e outubro/2003, homogeneamente distribuída nas duas estações (Tabela 1 e Fig. 2).
Em síntese, apenas na mata observou-se correlação negativa significativa entre a abundância de E. cordata e a precipitação (rs = -0,488; P = 0,015). Não foram observadas correlações significativas com os dados de temperatura.
As análises de sazonalidade têm se mostrado bastante variáveis nos trabalhos realizados no Nordeste. Rebêlo & Cabral (1997), Brito & Rêgo (2001), Silva & Rebêlo (2002) e Viana et al. (2002) observaram maior atividade dos machos na estação chuvosa; Neves & Viana (1999) nos meses mais secos. Entretanto, Neves & Viana (1997) e Bezerra & Martins (2001) observaram correlação apenas com a variação térmica, apesar da ocorrência de grande atividade nos períodos mais secos.
É de se esperar que haja variação sazonal na abundância de Euglossina (Roubik 2001). Até mesmo nos trópicos, as comunidades de Euglossina modificam-se ao longo do ano (Roubik & Hanson 2004). Mesmo a maioria das espécies estando presentes durante todo o ano, elas são dinâmicas e têm picos distintos e períodos onde há menor número de indivíduos. Ackerman (1983) explica que flutuações sazonais na abundância decorrem provavelmente dos padrões locais de nidificação e emergência das abelhas e que as variações na disponibilidade de recursos podem ser responsáveis pelas mudanças sazonais na comunidade de Euglossina. Embora alguns indivíduos realmente se desloquem por longas distâncias, isso não é responsável pela grande maioria do fenômeno sazonal observado nas comunidades de Euglossina (Ackerman 1983). Para Rebêlo & Garófalo (1991) a ocorrência de algumas espécies com pequeno número de machos amostrados num determinado período de tempo pode se dever à fraca associação entre elas e as fragrâncias ou simplesmente reflete a própria estrutura das populações locais.
Apesar da grande variabilidade na sazonalidade das espécies no Nordeste, os dados deste estudo, assim como dados de dissertações não publicadas obtidos em ambientes no estado da Paraíba, mostram que, de modo geral, as espécies apresentam maior abundância nos períodos mais secos em áreas de mata e são homogeneamente distribuídas nos períodos seco e chuvoso em áreas de dunas.
Agradecimentos
Agradecemos aos moradores da APA da Barra do Rio Mamanguape pelo acolhimento e apoio durante os trabalhos de campo, aos administradores da APA pelo apoio logístico, aos amigos do curso de graduação em Ciências Biológicas da UFPB pela ajuda nas coletas e ao CNPq pelas bolsas concedidas.
Received 20/XII/06. Accepted 09/X/07.
- Ackerman, J.D. 1983. Diversity and seasonality of male euglossine bees (Hymenoptera: Apidae) in central Panama. Ecology 64: 274-283.
- Bezerra, C.P. & C.F. Martins. 2001. Diversidade de Euglossinae (Hymenoptera, Apidae) em dois fragmentos de Mata Atlântica localizados na região urbana de João Pessoa, Paraíba, Brasil. Rev. Bras. Zool. 18: 823-835.
- Brasil. 1993. Decreto no 924, de 10 de setembro de 1993. Cria a Área de Proteção Ambiental da Barra do Rio Mamanguape, no Estado da Paraíba e dá outras providências. Diário Oficial [da] República Federativa do Brasil, Poder Executivo, Brasília, DF.
- Brito, C.M.S. de & M.M.C. Rêgo. 2001. Community of male Euglossini bees (Hymenoptera: Apidae) in a secondary Forest, Alcântara, MA, Brazil. Braz. J. Biol. 61: 631-638.
- Brown Jr., K.S. 1991. Conservation of neotropical environments: Insects as indicators, p.349-404. In N.M. Collins & J.A. Thomas (eds.), The conservation of insects and their habitats. London, Academic Press, 430p.
- Carvalho, M.G.R.F. de. 1982. Estado da Paraíba Classificação geomorfológica. João Pessoa, Editora Universitária/Universidade Federal da Paraíba, 72p.
- Dodson, C.H., R.L. Dressler, H.G. Hills, R.M. Adams & N.H. Williams. 1969. Biologically active compounds in orchid fragrances. Science 164: 1243-1249.
- Dressler, R.L. 1982. Biology of the orchid bees (Euglossinae). Annu. Rev. Ecol. Syst. 13: 373-394.
- Governo do Estado da Paraíba. 1985. Atlas geográfico do Estado da Paraíba. Secretaria de Educação. João Pessoa, Universidade Federal da Paraíba, Grafes.
- Janzen, D.H., P.J.Devries, M.L. Higgins & L.S.Kimsey. 1982. Seasonal and site variation in Costa Rica euglossine bees at chemical baits in lowland deciduous and evergreen forests. Ecology 63: 66-74.
- Michener, C.D. 2000. The bees of the world. Baltimore, Johns Hopkins University Press, xiv + 913p.
- Ministério do Meio Ambiente. 2002. Avaliação e identificação de áreas e ações prioritárias para conservação, utilização sustentável e repartição de benefícios da biodiversidade brasileira. Brasília, MMA/SBF, 404p.
- Nemésio, A. & F.A. Silveira. 2006. Edge effects on the orchid bee fauna (Hymenoptera: Apidade: Apini: Euglossina) at a large remnant of Atlantic Rain Forest in southeastern Brazil. Neotrop. Entomol. 35: 313-323.
- Nemésio, A. & F.A. Silveira. 2007. Orchid bee fauna (Hymenoptera: Apidade: Euglossina) of Atlantic Forest fragments inside an urban area in southeastern Brazil. Neotrop. Entomol. 36: 186-191.
- Neves, E.L. das & B.F. Viana. 1997. Inventário da fauna de Euglossinae (Hymenoptera, Apidae) do baixo sul da Bahia, Brasil. Rev. Bras. Zool. 14:831-837.
- Neves, E.L. das & B.F. Viana. 1999. Comunidade de machos de Euglossinae (Hymenoptera, Apidae) das Matas Ciliares da margem esquerda do Médio Rio São Francisco, Bahia. An. Soc. Entomol. Bras. 28: 201-210.
- Peruquetti, R.C., L.A.O. Campos, C.D.P. Coelho, C.V.M. Abrantes & L.C.V. Lisboa. 1999. Abelhas Euglossini (Apidae) de áreas de Mata Atlântica: Abundância, riqueza e aspectos biológicos. Revta. Bras. Zool. 16 (Supl. 2): 101-118.
- Ramírez, S., R.L. Dressler & M. Ospina. 2002. Abejas euglossinas (Hymenoptera: Apidae) de la Región Neotropical: Listado de especies com notas sobre su biología. Biota Colombiana 3: 7-118.
- Rebêlo, J.M.M. 2001. História natural das Euglossíneas As abelhas das orquídeas. São Luís, Maranhão, Lithograf Editora, 152p.
- Rebêlo, J.M.M. & A.J.M. Cabral. 1997. Abelhas Euglossinae de Barreirinhas, zona do litoral da Baixada Oriental Maranhense. Acta Amazônica 27: 145-152.
- Rebêlo, J.M.M. & C.A. Garófalo. 1991. Diversidade e sazonalidade de machos de Euglossini (Hymenoptera, Apidae) e preferências por iscas-odores em um fragmento de floresta no Sudeste do Brasil. Rev. Bras. Biol. 51:787-799.
- Roubik, D.W. 1989. Ecology and natural history of tropical bees. Cambridge, University Press, 514p.
- Roubik, D.W. 2001. Ups and downs in pollinator populations: When is there a decline? Conserv. Ecol. 5 [online] URL: http://www.consecol.org/vol5/iss1/art2/ Acesso em 19/10/2006.
- Roubik, D.W. 2004. Long-term studies of solitary bees: What the orchid bees are telling us, p.97-103. In B.M. Freitas & J.O.P. Pereira (eds.), Solitary bees: Conservation, rearing and management for pollination. Fortaleza, CE, Imprensa Universitária, 285p.
- Roubik, D.W. & P.E. Hanson. 2004. Abejas de orquídeas de la America tropical - Biología y guía de campo. Costa Rica, Editorial INBio, 370p.
- Silva, F.S. & J.M.M. Rebêlo. 1999. Euglossine bees (Hymenoptera: Apidae) of Buriticupu, Amazonia of Maranhão, Brazil. Acta Amazonica 29: 587-599.
- Silva, F.S. & J.M.M. Rebêlo. 2002. Population dynamics of Euglossinae bees (Hymenoptera, Apidae) in an early second-growth forest of Cajual Island, in the state of Maranhão, Brazil. Braz. J. Biol. 62: 15-23.
- Souza, A.K.P., M.I.M. Hernández & C.F. Martins. 2005. Riqueza, abundância e diversidade de Euglossina (Hymenoptera, Apidae) em três áreas da Reserva Biológica Guaribas, Paraíba, Brasil. Rev. Bras. Zool. 22: 320-325.
- Tonhasca Jr., A., J.L. Blackmer & G.S. Albuquerque. 2002. Abundance and diversity of euglossine bees in the fragmented landscape of the Brazilian Atlantic Forest. Biotropica 34: 416-422.
- Viana, B.F., A.M.P. Kleinert & E.L. das Neves. 2002. Comunidade de Euglossini (Hymenoptera, Apidae) das dunas litorâneas do Abaeté, Salvador, Bahia, Brasil. Rev. Bras. Entomol. 46: 539-545.
- Williams, N.H. & W.M. Whitten. 1983. Orchid floral fragrances and male euglossine bees: Methods and advances in last sesquidecade. Biol. Bull. 164: 355-395.
Datas de Publicação
-
Publicação nesta coleção
14 Jul 2008 -
Data do Fascículo
Jun 2008
Histórico
-
Aceito
09 Out 2007 -
Recebido
20 Dez 2006