Abstracts
In this review we re-visit and discuss the current knowledge on ecophysiology of citrus trees, addressing the influence of environmental conditions on citrus photosynthesis. Knowledge of physiological responses of citrus trees to their surrounding environment is essential in order to improve crop production and plant development, both being consequences of appropriate horticultural management in citrus orchards. In this context, citrus photosynthesis is addressed as the primary source of carbon and energy for plant growth and development. The photosynthetic activity on both a daily and a seasonal scale is reviewed, taking into consideration the physiological aspects related to seasonal variation of photochemical and biochemical activities, stomatal conductance and leaf water potential. These aspects are treated for citrus plants growing in subtropical climates with varying environmental conditions, such as moderate to severe drought during the winter season. In addition, the possible inhibitory/stimulatory effects of carbohydrate metabolism on citrus photosynthesis are discussed with regard to the source-sink relationship. Field experimentation that enhances knowledge concerning citrus ecophysiology in subtropical climates is highlighted. Among interesting subjects to be unraveled by future research, we may point out the effects of low temperatures on citrus photosynthesis and water relations, the nature of the relationship between leaf carbohydrate content and photosynthesis, and the significance of photosynthesis in different canopy layers and positions in relation to the total carbon gain in mature citrus trees.
carbohydrate; Citrus; chlorophyll fluorescence; gas exchange; seasonality
Objetivou-se re-visitar e discutir o conhecimento atual sobre a ecofisiologia dos citros, tratando da influência das condições ambientes na fotossíntese. O conhecimento das respostas fisiológicas dos citros ao ambiente que os circundam é essencial para aumentar a produção de frutos e o desenvolvimento das plantas, conseqüências do manejo agrícola apropriado em pomares de citros. Nesse contexto, a fotossíntese dos citros é tratada como a fonte primária de carbono e energia para o crescimento e desenvolvimento vegetal. A atividade fotossintética, tanto numa escala diária como sazonal, é revisada em árvores de citros, considerando-se aspectos fisiológicos relacionados à variação sazonal das atividades fotoquímica e bioquímica, condutância estomática e potencial da água na folha. Esses aspectos são abordados em plantas cultivadas em climas subtropicais com condições ambientais diversas, tal como seca durante o inverno, variando de moderada a severa. Ainda, possíveis efeitos inibitórios/estimulantes do metabolismo de carboidratos na fotossíntese dos citros são discutidos com ênfase na relação fonte-dreno. Experimentação de campo para aumentar o conhecimento sobre a ecofisiologia dos citros é ressaltada. Dentre assuntos interessantes para serem elucidados por pesquisas futuras, são indicados os efeitos das baixas temperaturas na fotossíntese e relações hídricas dos citros, a natureza da relação entre o conteúdo foliar de carboidratos e a fotossíntese, e a significância da fotossíntese nas diferentes camadas e posições do dossel em relação ao ganho total de carbono em plantas adultas.
carboidrato; Citrus; fluorescência da clorofila; sazonalidade; trocas gasosas
REVIEW
Some aspects of citrus ecophysiology in subtropical climates: re-visiting photosynthesis under natural conditions
Alguns aspectos da ecofisiologia dos citros em climas subtropicais: revisitando a fotossíntese sob condições naturais
Rafael V. Ribeiro* * Corresponding author: rafael@iac.sp.gov.br ; Eduardo C. Machado
Setor de Fisiologia Vegetal, Centro de Pesquisa e Desenvolvimento de Ecofisiologia e Biofísica, Instituto Agronômico (IAC), Caixa Postal 28, 13012-970 Campinas, SP, Brasil
ABSTRACT
In this review we re-visit and discuss the current knowledge on ecophysiology of citrus trees, addressing the influence of environmental conditions on citrus photosynthesis. Knowledge of physiological responses of citrus trees to their surrounding environment is essential in order to improve crop production and plant development, both being consequences of appropriate horticultural management in citrus orchards. In this context, citrus photosynthesis is addressed as the primary source of carbon and energy for plant growth and development. The photosynthetic activity on both a daily and a seasonal scale is reviewed, taking into consideration the physiological aspects related to seasonal variation of photochemical and biochemical activities, stomatal conductance and leaf water potential. These aspects are treated for citrus plants growing in subtropical climates with varying environmental conditions, such as moderate to severe drought during the winter season. In addition, the possible inhibitory/stimulatory effects of carbohydrate metabolism on citrus photosynthesis are discussed with regard to the source-sink relationship. Field experimentation that enhances knowledge concerning citrus ecophysiology in subtropical climates is highlighted. Among interesting subjects to be unraveled by future research, we may point out the effects of low temperatures on citrus photosynthesis and water relations, the nature of the relationship between leaf carbohydrate content and photosynthesis, and the significance of photosynthesis in different canopy layers and positions in relation to the total carbon gain in mature citrus trees.
Ke ywords: carbohydrate, Citrus, chlorophyll fluorescence, gas exchange, seasonality
RESUMO
Objetivou-se re-visitar e discutir o conhecimento atual sobre a ecofisiologia dos citros, tratando da influência das condições ambientes na fotossíntese. O conhecimento das respostas fisiológicas dos citros ao ambiente que os circundam é essencial para aumentar a produção de frutos e o desenvolvimento das plantas, conseqüências do manejo agrícola apropriado em pomares de citros. Nesse contexto, a fotossíntese dos citros é tratada como a fonte primária de carbono e energia para o crescimento e desenvolvimento vegetal. A atividade fotossintética, tanto numa escala diária como sazonal, é revisada em árvores de citros, considerando-se aspectos fisiológicos relacionados à variação sazonal das atividades fotoquímica e bioquímica, condutância estomática e potencial da água na folha. Esses aspectos são abordados em plantas cultivadas em climas subtropicais com condições ambientais diversas, tal como seca durante o inverno, variando de moderada a severa. Ainda, possíveis efeitos inibitórios/estimulantes do metabolismo de carboidratos na fotossíntese dos citros são discutidos com ênfase na relação fonte-dreno. Experimentação de campo para aumentar o conhecimento sobre a ecofisiologia dos citros é ressaltada. Dentre assuntos interessantes para serem elucidados por pesquisas futuras, são indicados os efeitos das baixas temperaturas na fotossíntese e relações hídricas dos citros, a natureza da relação entre o conteúdo foliar de carboidratos e a fotossíntese, e a significância da fotossíntese nas diferentes camadas e posições do dossel em relação ao ganho total de carbono em plantas adultas.
Palavras-chave: carboidrato, Citrus, fluorescência da clorofila, sazonalidade, trocas gasosas
INTRODUCTION
As a perennial and evergreen crop, citrus trees are subjected to large seasonal variation of environmental conditions throughout the annual cycle. Considering the environmental changes that naturally occur in the Brazilian subtropical climates, we may emphasize two periods: (a) the autumn-winter season with low soil water availability and low temperatures; and (b) the spring-summer season with an abundant rainfall, high temperature and high radiation loading (Ribeiro et al., 2005a; 2006a; Machado et al., 2007). Depending on the growing areas, these environmental conditions could be more or less limiting for citrus growth and development.
Although the physiological response of citrus trees to natural environmental changes is a very interesting subject with regard to improved orchard management (Silva et al., 2005; Ribeiro, 2006) and essential for the advance of our understanding of the developmental patterns of the species, little is known about the ecophysiology of citrus trees grown in the Brazilian subtropical climates. The lack of information on the physiological aspects of field-grown plants is probably due to the high complexity of the interaction of citrus trees with their environment, ranging from soil temperature and root metabolism to the variation in light exposure of leaves. A significant number of citrus-related papers are available; however, most of the results and conclusions concern the physiological responses of plants to a specific environmental factor obtained under controlled or semi-controlled conditions, such as drought, temperature and evaporative demand (Ahrens and Ingram, 1988; Vu and Yelenosky, 1989; Habermann et al., 2003a,b; Machado et al., 2005; Pimentel et al., 2004; Ribeiro et al., 2004; Souza et al., 2004; Guo et al., 2006).
Significant efforts have been made to clarify the physiological responses of citrus plants to water deficit, high temperature and high irradiance conditions (Vu and Yelenosky, 1988a,b; Brakke and Allen Jr., 1995; Savé et al., 1995; Machado et al., 1999; Medina et al., 2002; Jifon and Syvertsen, 2003; Raveh et al., 2003; Ribeiro et al., 2004; Guo et al., 2006). On the other hand, the influence of low temperature either in the soil or air has been largely placed aside, and only a few reports are available concerning the influence of this important environmental constraint on citrus physiology (Syvertsen et al., 1983; Vu and Yelenosky, 1987; Ribeiro, 2006). Systematic experimentation with field-grown citrus plants is needed to uncover ecophysiological patterns under natural conditions; however, studies of this kind are not widespread in Brazil (Ribeiro, 2006; Machado et al., 2007).
Regarding plant metabolism, photosynthesis is an important process that may be addressed from an ecophysiological point of view, since the early photochemical events are driven by light availability while the final product (carbohydrate) is a key element in the source-sink relationship. In citrus trees growing in a subtropical climate, both seasonal drought and low temperatures affect photosynthetic activity during the winter season, whereas high temperatures and high evaporative demands are the most limiting factors during the summer season (Ribeiro et al., 2005a; 2006a; Ribeiro, 2006). In such conditions, high light availability may also be an overabundant resource that may affect the photochemical reactions, since the original habitat of citrus species is the shaded environment of the forest understorey (Syvertsen and Lloyd, 1994; Davies and Albrigo, 1994; Spiegel-Roy and Goldschmidt, 1996). With regard to the interaction between photosynthesis and carbohydrate content, an inhibitory effect of high levels of leaf carbohydrate on the photosynthetic rates of citrus leaves has been suggested (Iglesias et al., 2002), revealing the source-sink relationship in citrus trees as an important ecophysiological theme.
In this review we shall re-visit and discuss the current knowledge concerning the ecophysiology of citrus trees under the Brazilian subtropical conditions, focusing on the interaction between environment and photosynthesis, the primary source of energy and carbon for plant growth and development.
PHOTOCHEMISTRY OF FIELD-GROWN PLANTS
Despite the limited data concerning chlorophyll fluorescence of citrus leaves under field conditions, some evidence has suggested that citrus plants are well acclimated or have a good potential for acclimation to the radiation regimes of subtropical climates (Syvertsen, 1984; Syvertsen and Smith, 1984). Firstly, a lack of chronic photoinhibition has been reported in sweet orange plants in areas with distinct environmental conditions (Machado et al., 2006; Ribeiro, 2006). Secondly, citrus leaves may potentially dissipate excessive light energy by non-photochemical processes (Blanke, 2000; Ribeiro, 2006).
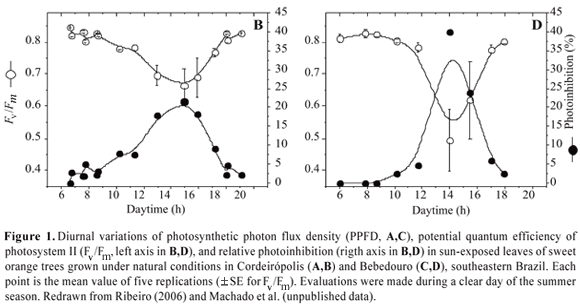
Citrus plants have large or small seasonal variation of the potential quantum efficiency of PSII (as assessed by the variable-to-maximum chlorophyll fluorescence ratio, Fv/Fm), depending on the environmental conditions of the growing regions (Figure 1). In general, sweet orange plants exhibit the lowest Fv/Fm values during the summer season, found normally at midday and early afternoon (Figure 1B,D). Reduction of Fv/Fm to values below 0.725 suggests the occurrence of photoinhibition (Critchley, 1998), as found in citrus plants (Veste et al., 2000; Jifon and Syvertsen, 2003; Machado et al., 2006; Ribeiro, 2006). In fact, photoinhibition observed in citrus is a protective mechanism, reducing energetic pressure at PSII level and avoiding photodamage and consequent photooxidation. This photoprotective nature of photoinhibition, referred to as dynamic photoinhibition by Osmond (1994), is indicated by the rapid recovery of Fv/Fm when the photosynthetic photon flux density (PPFD) decreases, a physiological feature that is observed under both field and greenhouse conditions (Blanke, 2000; Medina et al., 2002; Jifon and Syvertsen, 2003; Machado et al., 2006; Ribeiro, 2006). Higher dynamic photoinhibition can be observed in warmer regions during the summer season (Figure 1) when high radiation load and high air temperature occur, particularly at midday.

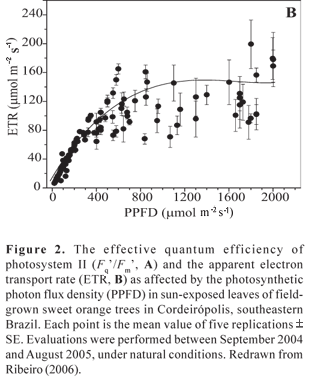
Photoinhibition of citrus is also a consequence of the low-light saturation of photosynthesis (Vu et al., 1986; Habermann et al., 2003a; Ribeiro et al., 2003a; Machado et al., 2005; Ribeiro et al., 2006b). Most light energy intercepted by leaves but not used in photochemistry (generation of ATP and NADPH) is dissipated in non-photochemical reactions, such as heat generation at the PSII level (Demmig-Adams and Adams, 1996; Horton et al., 1996; Ort, 2001). Excessive light energy at PSII is commonly attained under subtropical conditions, where daily-integrated global solar radiation may reach values around 35 MJ m-2 d-1 during the summer season (Ribeiro et al., 2005a). As observed in other species, citrus shows a significant and progressive reduction in the effective quantum efficiency of PSII (Fq'/Fm') as PPFD increases up to ca. 750 µmol m-2 s-1 (Figure 2A) and Medina et al., 2002. This response reflects the closing of PSII reaction centers and is directly related to electron flow between PSII and PSI (Maxwell and Johnson, 2000), that is, apparent electron transport rate, ETR (Figure 2B). At PPFD above 1000 µmol m-2 s-1, Fq'/Fm' does not show significant alteration (Figure 2B) and the relative light energy excess reaches the maximum values (Ribeiro, 2006), as given by the relation between Fv/Fm and Fq'/Fm' (see Bilger et al., 1995). The rapid response of Fq'/Fm' to PPFD changes can be considered as an adaptive mechanism that maintains reasonable light energy pressure at the PSII level either under high (low Fq'/Fm') or low (high Fq'/Fm') light conditions.

The non-photochemical quenching of chlorophyll fluorescence (NPQ) is an essential mechanism for plant protection against excessive light energy (Horton et al., 1996; Ort, 2001; Ribeiro et al., 2003b). The NPQ is closely related to the DpH formation across the thylakoid membrane and to the xanthophyll cycle activity, having an important role in photoprotection during environmental stresses (Horton et al., 1996) such as high temperature in citrus leaves (Guo et al., 2006). Citrus plants exhibit a linear increase of NPQ with increasing relative light energy excess at the PSII level, regardless of additional abiotic or biotic stresses (Ribeiro et al., 2003b). Alternative electron sinks are also an important mechanism to deal with excess light energy in citrus (Ribeiro et al., 2003a; 2004; Guo et al., 2006). Photorespiration, water-water cycle, nitrogen and sulphur metabolism are physiological processes that use photochemical products and then indirectly protect leaf tissues from photodamage (Osmond and Grace, 1995; Guo et al., 2006; Baker et al., 2007). The photochemical performance of citrus under field conditions is poorly understood, especially if we consider the seasonal variation of related physiological processes. However, we assume that photochemical activity does not limit CO2 assimilation in plants grown under subtropical conditions, since there is sufficient ETR to support the photosynthetic rates during the winter season and leaves do not exhibit chronic photoinhibition during the annual cycle (Ribeiro, 2006).
Light exposure of citrus leaves is another important aspect regarding energy availability and photochemistry. Most studies under field or controlled conditions address sun-exposed leaves of the citrus canopy, which represent around 20% of the total leaf area in a cross-section of an orange tree canopy (Cohen and Fuchs, 1987). The internal canopy layer does not receive more than 10% of the total PPFD available at the external canopy layer (Greene and Gerber, 1967; Davies and Albrigo, 1994). In this context, there is a large leaf area that is not subjected to excessive light energy and may be a source of carbon and energy for plant growth and development. While excess light energy is present at the external canopy layer, the PPFD at deep layers is probably higher than the light compensation point and promotes low CO2 assimilation (Davies and Albrigo, 1994). Shaded citrus leaves have potential for acclimation to low PPFD, showing increased chlorophyll content and consequently increased light-absorption efficiency (Syvertsen and Smith, 1984).
Environmental stresses that cause dysfunction in CO2 assimilation and hence decreased ATP and NADPH consumption may potentially affect plant photo-chemistry (Schreiber and Bilger, 1987; Baker and Rosenqvist, 2004). Even low PPFD can represent excessive light energy at the PSII level under stressful conditions (Demmig-Adams and Adams, 1996; Horton et al., 1996; Ort, 2001). However, photochemical events are relatively tolerant to water deficit and high temperature, being commonly down-regulated under such conditions by reduced CO2 assimilation (Schreiber and Bilger, 1987). Although high temperatures are responsible for significant inhibition of photochemical activity (Schreiber and Bilger, 1987; Georgieva, 1999; Guo et al., 2006), this environmental factor is probably not limiting for citrus leaves under subtropical conditions. In fact, tropical species have a critical temperature for photochemical damage that is close to 46ºC (Smillie and Nott, 1979) whereas citrus leaves reach maximum temperatures of around 42ºC during the summer season (Machado et al., unpublished data; Ribeiro et al., 2005a). In addition, citrus plants may potentially acclimate to varying growth temperature, showing small decreases in photochemical performance at high temperature (up to 40ºC) under warmer conditions (Ribeiro et al., 2004; 2006b). Therefore, the negative effects of high temperature on citrus photosynthesis are probably due to diffusive and/or to biochemical limitations.
SEASONAL CHANGES IN SHOOT WATER POTENTIAL AND STOMATAL CONDUCTANCE
Plant physiological variables such as leaf or stem water potential, stomatal conductance or resistance and leaf or canopy transpiration have been often studied in citrus plants under natural conditions (Cohen et al., 1997; Machado et al., 2002; 2007; Raveh et al., 2003; Angelocci et al., 2004; Rana et al., 2005; Silva et al., 2005; Ribeiro, 2006; Hu et al., 2007; Oguntunde et al., 2007). According to Syvertsen and Lloyd (1994), variations in plant water relations are the principal plant responses to their surrounding and changing environment. Since shoot water status can regulate stomatal aperture, leaf water relations are deeply related to gas exchanges since the former affects the influx of CO2 to the mesophyll as well as leaf transpiration, a well-known cooling processes (Nobel, 1999).
Under subtropical conditions, temperature and water availability are the most important environmental elements affecting citrus water relations. Light energy availability is not directly considered as an environmental constraint since stomatal aperture is maximal at relatively low PPFD (Cohen and Cohen, 1983; Vu et al., 1986; Machado et al., 2005). However, stomatal conductance significantly fluctuates over the seasons for a same PPFD, even in well-hydrated plants (Figure 3). This finding suggests that other environmental factors are limiting stomatal aperture during the winter season, such as low temperature (Elfving et al., 1972; Moreshet and Green, 1984). The influence of light availability on stomatal behavior is also indirect via changes in thermal regimes at the leaf level, determining the leaf-to-air vapor pressure difference (VPDL).
In subtropical climates, citrus plants exhibit higher stem water potential during the summer rainy season; in contrast, significant reduction in stem water potential is found in the winter dry season (Figure 4). In addition, leaf or stem water potential has a significant daily variation, reaching the lowest values during the afternoon (Figure 4C; Elfving et al., 1972; Syvertsen and Albrigo, 1980a,b; Syvertsen et al., 1981; Moreshet and Green, 1984; González-Altozano and Castel, 1999). As transpiration is higher than water uptake, an imbalance is generated in the water continuum through the plant body and then the shoot water potential is reduced in the afternoon, even in well-irrigated plants. During the wet period, the higher the diurnal transpiration rates the lower will be the minimum (measured at 1400 h) stem water potential, as found in 'Valencia' orange trees (Figure 5A). This pattern suggests that water availability allows transpiration of citrus plants and then stem water potential is decreased as a consequence. On the other hand, transpiration correlates better with pre-dawn stem water potential during the dry season (autumn-winter), indicating that citrus plants achieve higher diurnal transpiration as they are less affected by water deficit (Figure 5B). In 'Tahiti' lime trees subjected to water deficit, midday and predawn leaf water potential differed by 10% and 40%, respectively, when comparing irrigated and non-irrigated plants (Silva et al., 2005). Therefore, pre-dawn leaf or stem water potential is more appropriated to describe the watering treatment effects on citrus plants under natural conditions.


Medina et al. (2005) have suggested that citrus plants follow an anysohydric pattern, which is characterized by (i) a significant decrease in leaf water potential over the course of the day due to the evaporative demand and by (ii) lower leaf water potential in drought-stressed plants when compared with well-watered individuals (Tardieu and Simonneau, 1998). Accordingly, field-grown citrus trees showed significant diurnal reduction in stem water potential during the winter season, when non-irrigated plants exhibited lower stem water potential than irrigated plants regardless of daytime (Ribeiro, 2006).
Diurnal reductions in leaf or stem water potential between -1.2 and -1.8 MPa in citrus plants during the winter have been registered, depending on the growing region (Habermann, 2004; Ribeiro, 2006; Machado et al., 2007). Over the course of the year, stem water potential varied from -0.20 (at pre-dawn) to -1.75 MPa (at 1400 h) in irrigated plants, reaching -2.30 MPa in non-irrigated plants (Ribeiro, 2006; Machado et al., 2007). In regions with high evaporative demand and thermal regimes, leaf abscission in citrus plants was noticed when predawn leaf water potentials were around -2.75 ± 0.07 MPa (Pires et al., unpublished data). Predawn stem water potentials lower than -0.7 MPa have been found in field-grown citrus plants during the winter season (Ribeiro, 2006), a sufficient value to impair leaf gas exchange of 'Tahiti' lime trees under field conditions (Silva et al., 2005).
There is appreciable stomatal closure even in irrigated plants with high stem water potential during the winter (Ribeiro, 2006), which reveals that the general assumption that seasonal and diurnal patterns of stomatal aperture are coupled to the variations of the leaf/stem water potential (leaf turgor) may be an oversimplification. In fact, stomata can be considered as a multi-sensorial organ, sensing changes in both environmental and physiological factors (Zeiger et al., 1987). High stomatal conductance is commonly reported in leaves with high water potential; as the latter decreases during the diurnal cycle the stomatal aperture also reduces. As a result, the highest stomatal conductances are found around mid-morning (between 0900 and 1030 h) under subtropical conditions, regardless of the season (Machado et al., 2002, 2006, 2007; Ribeiro, 2006). During a clear day, increases in transpiration in response to the air evaporative demand causes reduction in leaf/stem water potential, which in turn decreases stomatal conductance, a feedback stomatal response (Cohen and Cohen, 1983; Syvertsen and Lloyd, 1994; Nobel, 1999). The feedforward stomatal response in citrus is not commonly noticed under controlled conditions; however, this response was observed under high evaporative demand (VPDL ~ 4.5 kPa) in field-grown plants during the summer (Ribeiro, 2006), when a halving in stomatal conductance caused a decrease in leaf transpiration (~ 43%) in plants with stem water potential relatively high (around -1.25 MPa at 1400 h). Recently, Hu et al. (2007) also reported a feedforward response in citrus plants under field conditions.
At early morning and late evening, light availability seems to be the principal regulatory element affecting stomatal aperture of citrus plants. When PPFD is non-limiting, VPDL seems to be the chief environmental factor regulating stomatal aperture under natural conditions (Ribeiro et al., unpublished data). Even in well-irrigated plants, stomata tend to close at VPDL above 1.5 kPa (Figure 6; see also Habermann et al., 2003b; Jifon and Syvertsen, 2003; Machado et al., 2005). For temperature, there seems to be an optimum temperature range rather than a single optimum point for maximum stomatal aperture in citrus leaves (Syvertsen and Lloyd, 1994; Machado et al., 2005). In any case, stomata sensitivity to temperature is low between 25 and 40ºC (Machado et al., 2005) but outside this range [it should be noted that fluctuations in leaf temperatures between 15 (winter) and 42ºC (summer) in field-grown orange trees have been found under the subtropical Brazilian conditions (Ribeiro et al., 2005a; Machado et al., unpublished data)], stomata aperture is probably affected, as occurs during the winter season. In fact, there is sufficient evidence that low night temperature and low soil temperature play an important role in the stomatal mechanism in citrus plants under natural conditions (Elfving et al., 1972; Moreshet and Green, 1984; Lloyd and Howie, 1989; Angelocci et al., 2004; Ribeiro, 2006). Although diurnal environmental conditions were not limiting for maximum stomatal aperture during the winter season (Ribeiro, 2006), maximum stomatal conductance in irrigated 'Valencia' citrus trees was about 65% lower in winter than in spring, i.e. ~ 0.08 against ~ 0.22 mol m-2 s-1 (Table 1).
Low stomatal conductance during winter season may be induced by hydraulic and/or chemical signals. Regarding hydraulic signals as those referred to changes in shoot water potential, it is known that low soil temperature causes reduction in both water uptake by citrus roots due to decreases in root permeability (Elfving and Kaufmann, 1972) and root and tree hydraulic conductivity (Syvertsen et al., 1983; Moreshet and Green, 1984). Changes in pH and ions of the xylem sap (Wan et al., 2004), reductions in leaf concentration of cytokinins (Veselova et al., 2005) and increases in ABA content (Davies and Zhang, 1991; Tardieu and Simonneau, 1998; Gomes et al., 2003) have been invoked as potential chemical signals affecting stomatal aperture. However, this topic should be further studied in citrus plants under field conditions to clarify how low temperature and water deficit change the citrus water relations and stomatal conductance during the winter season. In general, low stomatal conductance affects photosynthetic rates by decreasing CO2 availability at the mesophyll and carboxylation sites. Anyway, as stomatal conductance is low even under high stem water potential (> -1.5 MPa at 1400 h) during the winter season (Ribeiro, 2006), we believe that chemical signals play a key role in regulating stomatal aperture in subtropical climates with moderate seasonal water deficits.
An important aspect to be considered in seasonal changes of plant water relations is the influence of rootstocks, which have different physiological characteristics and cause significant differences in shoot hydration (Eissenstat, 1991; Syvertsen and Lloyd, 1994; Medina et al., 1998; 2005; Barry et al., 2004; Cerqueira et al., 2004). Therefore, rootstocks may differentially affect citrus sensitivity to varying environmental conditions. For example, efficient scion/rootstock combinations may improve the water-use efficiency of citrus plants (relationship between photosynthesis and transpiration) (Khairi and Hall, 1976; Brakke and Allen Jr., 1995; Medina et al., 1999; Machado et al., 2002; 2005) and thus their adaptive responses to drought stress, which is a current matter for the rational use of water in agriculture. In general, plants should show increases in water-use efficiency at early drought stages since a reduction in stomatal conductance affects transpiration to a greater extent than photosynthesis (Nobel, 1999). However, since significant reductions in water-use efficiency have been noticed in field-grown citrus plants during the winter season, even in irrigated plants (Ribeiro, 2006), it may be suggested that (i) photosynthesis may be more affected than transpiration and (ii) environmental factors other than water availability may be more directly involved in the regulation of leaf gas exchange in citrus. This response pattern of water-use efficiency often differs in warmer regions, where severe seasonal drought and relatively high temperatures are common during the winter (Machado et al., 2007).
SEASONAL VARIATION IN LEAF CO2 ASSIMILATION
As an evergreen species with C3 photosynthetic metabolism, citrus trees show low photosynthetic rates under natural conditions, with maximum values around 13 µmol m-2 s-1 being found during the spring season (Figure 7B). The low CO2 assimilation in citrus leaves is probably a consequence of low CO2 partial pressure at the carboxylation sites, this being about 8 Pa lower than the intercellular CO2 concentration (Lloyd et al., 1992). Syvertsen and Lloyd (1994) and Spiegel-Roy and Goldschmidt (1996) have reported maximum photosynthetic rates ranging from 8 to 12 µmol m-2 s-1. Such differences are probably related to the environmental conditions of growing regions, mainly temperature affecting biochemistry and evaporative demand, which in turn affects stomatal conductance. Larger rates of photosynthesis may be expected in subtropical climates with adequate water availability (to support transpiration) and with a reasonable temperature range to sustain photosynthesis during most of the year. The diurnal-integrated leaf CO2 assimilation is a physiological variable that permits more adequate evaluation of how environment affects citrus trees. This assumption is based on the fact that the total CO2 gain throughout the diurnal period can integrate the influence of any environmental constraint occurring during the daytime on leaf photosynthesis. In fact, Machado et al. (2007) verified significant differences between CO2 assimilation of healthy and diseased citrus trees under field conditions when using this approach. In addition, the seasonal variation of citrus photosynthesis is more accentuated and evident when evaluating the diurnal-integrated values (Figure 7). As observed with the maximum photosynthetic rates, maximum diurnal-integrated values were also found during the spring season, reaching around 320 mmol CO2 m-2 d-1 in sun-exposed leaves (Figure 7B). During the annual cycle, higher photosynthesis is observed during the spring season followed by the summer and then by the winter season (Figure 7).
An important point is that these maximum photosynthetic rates are commonly measured in sun-exposed leaves and thus they may not be directly related to plant performance since most portions of canopy leaves are not sun-exposed, as discussed previously. Plant canopy position is also relevant in relation to photosynthetic rates since canopy layers may be subjected to different environmental conditions (Angelocci et al., 2004; Ribeiro et al., 2005a). For example, in a north-south-oriented citrus orchard, the east canopy position received direct solar radiation during the morning when VPDL and leaf temperature were low; in contrast, the west position received direct solar radiation during the afternoon when VPDL and leaf temperature were high (Ribeiro et al., 2005a). Therefore, as expected, the east position exhibits higher photosynthesis than the west position, regardless of water availability and seasons (Table 2). In addition, irrigation was more effective in increasing photosynthesis in leaves of the west canopy position and in decreasing the differences between canopy faces (Table 2). Ecophysiological instrumentation to measure entire citrus canopy gas exchange is desired and necessary to reveal the whole canopy photosynthetic patterns related to crop production.
After discussing some aspects related to photochemical and diffusive regulation of photosynthesis, we will focus on the other environmental factors that constrain the biochemical activity of citrus trees under natural conditions. Under field conditions, temperature frequently imposes limitation to CO2 fixation to a greater degree than PPFD availability. In fact, the photosynthetic enzyme ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) is saturated and shows around 90% of its activation under a PPFD close to 1000 µmol m-2 s-1 (Vu, 1999). In warmer regions where leaf temperature reaches values above 35ºC, the carboxylase activity of the Rubisco is probably reduced by increases in the photorespiration, i.e. oxygenase activity (Berry and Björkman, 1980; Hällgren et al., 1991). Under high temperatures, the relative solubility between CO2 and O2 is altered due to a higher decrease in CO2 solubility, which reduces the CO2 partial pressure at the Rubisco sites and thereby favors the oxygenase activity (Berry and Björkman, 1980; Jordan and Ogren, 1984). Non-stomatal limitation plays an important role in reducing citrus photosynthesis at leaf temperatures above 40ºC, with a relative importance overcoming the stomatal limitation (Jifon and Syvertsen, 2003; Syvertsen et al., 2003; Hu et al., 2007). On the other hand, low night temperatures may also affect the photosynthetic activity through modifications in photochemical activity, inhibition of CO2 carboxylation, changes in stomatal control of leaf gas exchange and impairment of carbohydrate metabolism (Allen et al., 2000; Allen and Ort, 2001). Therefore, the negative effects of high temperature are expected during the summer season, whereas the low temperature influence is likely during the winter season.
Vu (1999) reported that low temperatures caused biochemical impairment due to decreases in the carboxylation activity of Rubisco or to changes in the expression of genes related to the carbon metabolism in citrus plants. In fact, the maximum rate of Rubisco carboxylation (Vc,max) was significantly reduced during the winter when compared to the summer season, even in well-hydrated citrus plants (Table 3). The maximum rate of electron transport driving RuBP regeneration (Jmax) was also reduced during the winter (Table 3). There are a few reports concerning these biochemical variables in citrus species (Vu and Yelenosky, 1988a,b; Lloyd et al., 1992; Syvertsen and Lloyd, 1994), mainly under natural conditions. In these reports, Vc,max varied from 50 to 120 µmol m-2 s-1 and Jmax reached 140 µmol m-2 s-1. During the winter season, the decrease in Vc,max was higher than in Jmax in citrus leaves under natural conditions, which causes an increase in the Jmax:Vc,max ratio (around 3.0) and represents an imbalance between both biochemical processes (Table 3). On the other hand, Ribeiro et al. (2005a) and Ribeiro (unpublished data) were unable to detect biochemical impairment of photosynthesis during the summer season in citrus plants; they showed that reduction of CO2 assimilation was primarily caused by stomatal effects in a subtropical area with maximum midday temperatures between 30 and 35ºC. In warmer regions, however, higher temperature is likely to directly impair CO2 assimilation in exposed citrus leaves.
It is noteworthy that evergreen species like citrus may potentially acclimate their photosynthetic apparatus to increasing growth temperature. Some evidence for this was reported by Ribeiro et al. (2004), who showed significant changes in leaf gas exchange and photochemical performance when changing growth temperature from 25/20ºC to 35/20ºC (day/night). Such photosynthetic acclimation was also suggested by Syvertsen and Lloyd (1994) and Ribeiro et al. (2006b). In addition, Veste et al. (2000) reported photochemical adaptation in citrus plants subjected to extreme high temperatures under field conditions. These adaptive responses may include changes in chloroplast membrane properties leading to thermal stability as well as increases in stability of photosynthetic enzymes under high temperature (Berry and Björkman, 1980). Severe heat injury in citrus leaves is unlikely to occur under natural conditions in subtropical climates since the lethal temperatures are around 55ºC (Ahrens and Ingram, 1988). Maximum leaf temperatures noticed in field-grown citrus trees were around 36ºC in Florida State, USA (Ahrens and Ingram, 1988) and 42ºC in São Paulo State, Brazil (Ribeiro et al., 2005a; Machado et al., unpublished data). Although these temperatures were not lethal for citrus leaves, significant negative effects are expected in leaf CO2 gain at temperatures higher than 30ºC (Khairi and Hall, 1976; Ribeiro et al., 2004; Machado et al., 2005; Guo et al., 2006), mainly due to increases in photorespiration. During midday, the difference between leaf and air temperature may reach values greater than 5ºC during the summer season (Syvertsen and Albrigo, 1980b; Ribeiro et al., 2005a), suggesting that the cooling capacity of transpiration was insufficient or was reduced by stomatal closure.
During the winter season, drought may impair photosynthesis of citrus trees due to the partial inactivation of Rubisco as well as reduction in Rubisco amount (Vu and Yelenosky, 1988a,b). The biochemical limitation of photosynthesis during stressful conditions such as water deficit is suggested to cause reduction in leaf carbohydrate (starch and sucrose) contents (Vu and Yelenosky, 1989), which can potentially decrease plant growth and crop production. Even in a growing area with moderate seasonal drought, significant reduction in leaf carbohydrate (starch and reducing sugars) content was observed in field-grown citrus trees during the winter season (Ribeiro, 2006). Such drought influence is probably more severe in warmer regions, where the accumulated water deficiency during the winter season is commonly above 130 mm (Ciiagro, 2007). The diurnal-integrated CO2 assimilation of citrus trees grown in areas with contrasting environmental characteristics can reveal significant differences in seasonal susceptibility of photosynthetic activity to a specific environmental constraint (Table 4). It is important to mention that such responses represent concomitant stomatal and non-stomatal limitations to photosynthesis, as discussed previously.
INTERACTION BETWEEN CARBOHYDRATE CONCENTRATION AND PHOTOSYNTHESIS
Citrus growth and the presence of generative structures (buds, flowers and/or fruits) represent significant sinks of leaf reserves throughout the annual cycle (Bevington and Castle, 1985; García-Luis et al., 1988; 1995; Bustan and Goldschmidt, 1998). Such reserves are essential for crop production in citrus orchards when photoassimilate synthesis is unable to supply the energetic and carbon demand during key developmental stages such as flowering and fruit set (Syvertsen and Lloyd, 1994; Goldschmidt and Koch, 1996; Bustan and Goldschmidt, 1998; Goldschmidt, 1999; Ruiz et al., 2001; Iglesias et al., 2003; Prado et al., 2007). As a survival strategy, citrus trees have to accumulate reserve compounds such as carbohydrates to decrease the influence of the seasonal variation of photoassimilate supply and to maintain reproductive and vegetative development. As suggested by Syvertsen and Lloyd (1994), the year-to-year fluctuation in carbohydrate availability may be related to the alternate bearing in some citrus varieties. Although the root has an essential role as a reservoir of carbohydrates (Goldschmidt and Golomb, 1982; Goldschmidt and Koch, 1996; Li et al., 2003), leaves also have significant carbohydrate contents (García-Luis et al., 1995; Ruiz et al., 2001; Iglesias et al., 2003; Syvertsen et al., 2003). Considering the non-structural carbohydrates (soluble sugars and starch), citrus leaves reached a total sugar content of around 275 mg g-1 just before the reproductive flush of the spring season under subtropical conditions. In this situation, starch was the main carbohydrate in citrus leaves, reaching almost 200 mg g-1 (Ribeiro, 2006), representing the principal storage carbohydrate in citrus tree organs (Goldschmidt and Golomb, 1982; Goldschmidt and Koch, 1996).
Significant differences were noticed when comparing the seasonal dynamics of leaf carbohydrate contents in 'Valencia' orange trees grown under distinct environmental conditions (Jones and Steinacker, 1951; Ribeiro, 2006). Whereas in California/USA (the Northern hemisphere) both starch and soluble sugar contents were minimal during the summer season (Jones and Steinacker, 1951), under the Brazilian subtropical conditions citrus trees showed similar contents of total non-structural carbohydrates in the winter and summer seasons (Ribeiro, 2006). However, the highest leaf carbohydrate contents in both growth conditions were found in early spring, just before the flowering/vegetative flush (Jones and Steinacker, 1951; Ribeiro, 2006).
The lowest photosynthetic rates and, as a consequence, the lowest photoassimilate supply occur during the winter (Figure 7; Machado et al., 2002; Ribeiro, 2006), when citrus plants have reduced metabolic activity with reduced carbohydrate consumption (Davenport, 1980; Syvertsen and Lloyd, 1994; Goldschmidt and Koch, 1996). Under such conditions an increase in leaf carbohydrate content can be expected, this being a result of low sink demand rather than greater photoassimilate supply (Goldschmidt and Koch, 1996). However, this assumption was not found to be true for both young and mature 'Valencia' orange trees under natural conditions, since their leaf reserves during the winter were actually lower than or similar to those for the summer (Ribeiro et al., 2005b; Ribeiro, 2006). A large decrease in leaf reserves is observed during the flowering/fruiting period, i.e. spring season, which is related to the high sink strength of developing flowers (Jones and Steinacker, 1951; García-Luis et al., 1988; Ruiz et al., 2001; Ribeiro, 2006; Prado et al., 2007).
The continuous photosynthetic activity of citrus trees (Figure 7) enables the plants to accumulate leaf reserves during periods of low carbon and/or energetic demand (Goldschmidt and Koch, 1996). However, an important aspect is that the carbon sink strength varies according to the fruit developmental stage (Huang et al., 1992) and citrus variety, with late ones (e.g. 'Valencia') showing stronger sinks during the winter as a consequence of the longer presence of fruit (Goldschmidt and Koch, 1996). The vegetative growth also varies during the annual cycle, with appreciable alternation between shoot and root growth (Bevington and Castle, 1985). As shoot growth during the winter season is negligible (Ribeiro, 2006), we may hypothesize the occurrence of root growth under non-limiting conditions of soil temperature and water availability. Therefore, the sink strength produced by the presence of fruit and by root growth is probably related to the non-accumulation of leaf reserves during the winter season. It seems consensual that both vegetative and reproductive development drives the seasonality of sink strength in citrus trees under natural conditions.
Accumulation of leaf reserves during periods of reduced sink demand can decrease photosynthetic activity (Nafziger and Koller, 1976; Azcón-Bieto, 1983; Foyer, 1988; Goldschmidt and Huber, 1992; Nakano et al., 2000; Paul and Pellny, 2003). Some evidence has indirectly indicated that this inhibitory mechanism occurs in citrus leaves, as observed using girdled branches (Goldschmidt and Huber, 1992; Iglesias et al., 2002), de-fruited trees (Iglesias et al., 2002; Syvertsen et al., 2003), shoot removal (Syvertsen, 1994; Iglesias et al., 2002) and from sugar-feeding studies (Iglesias et al., 2002). In field-grown citrus plants, Iglesias et al. (2002) proposed that inhibition of photosynthesis was caused by increases in leaf carbohydrate content per se. Among the possible mechanisms of photosynthesis inhibition by carbohydrate content, we may speculate that this may involve a decrease in re-cycling of Pi to the chloroplast (Foyer, 1988), decreases in ATP and NADPH consumption/production and reduced regeneration of RuBP (Azcón-Bieto, 1983), impairment in expression of photosynthesis-related genes (Paul and Pellny, 2003), changes in cycling of sucrose and hexose sugars (Goldschmidt and Huber, 1992; Nakano et al., 2000), increased mesophyll resistance to CO2 diffusion (Nafziger and Koller, 1976; Nakano et al., 2000) and damage to thylakoid structure and chlorophyll degradation under extreme carbohydrate accumulation (Schaffer et al., 1986). However, Goldschmidt and Koch (1996) have pointed out that the control of citrus photosynthesis by sink demand is not clear under natural conditions. In addition, Syvertsen and Lloyd (1994) reported that it is not known if starch accumulates, both on a seasonal and daily basis, to a sufficient extent in citrus leaves to inhibit photosynthesis.
High photosynthesis in citrus leaves has been found to occur simultaneously with high leaf carbohydrate contents in plants under natural conditions (Figure 8), suggesting that the inhibitory effect of carbohydrate concentration on photosynthesis is not directly related to the amount of leaf carbohydrate per se. Probably, the photosynthesis of citrus plants is regulated by dynamic aspects of leaf carbohydrate rather than by the absolute carbohydrate content. The higher photosynthesis of citrus plants during the summer season was related to intense starch metabolism during the nocturnal period due to a great export of photoassimilate from the leaves (Table 5). In the summer season, total leaf area and photoassimilate export were five times higher than in the winter (Table 5). Recently, Li et al. (2003) reported that the inhibition of the expression of genes related to carbohydrate metabolism was apparently related to the decreasing trend in sugar concentration rather than absolute sugar levels. Accordingly, Goldschmidt and Huber (1992) have suggested that the inhibition of photosynthesis by end-product accumulation is related to changes in soluble sugar metabolism. Citrus plants probably respond to changes in carbon availability through the 'acclimatory' responses that balance the carbon supply and demand to optimize the capacity for sustained sink demand as a conservative response (Smith and Stitt, 2007). This is accomplished by regulating the partitioning of photoassimilates, starch degradation at night and plant growth in a changing environment, as occurs during the seasonal variation of in light and water availability, day length and temperature (Smith and Stitt, 2007). Changes in sugar contents (sucrose and hexoses) affect the expression of several genes encoding enzymes of photosynthesis and carbohydrate metabolism (Smith et al., 2004).
Based on the above discussion, we may suggest that a large decrease of leaf carbohydrate content can stimulate the photosynthetic rate (source) during the onset of citrus flowering (sink), as found in both mature and young 'Valencia' orange trees under natural conditions (Ribeiro et al., 2005b; Ribeiro, 2006). Therefore, the influence of daily changes in leaf carbohydrate content is likely to be more important to the source-sink relationship than the leaf carbohydrate concentration per se in citrus trees under natural conditions, i.e. without artificial modification of canopy and plant metabolism.
CURRENT AND FUTURE RESEARCH
The seasonal variation of citrus photosynthesis is evident and strongly regulated by the annual dynamics of environmental factors under subtropical conditions. With basic information about the response of citrus trees to their surrounding environment becoming available, the modeling of citrus photosynthesis can be improved for application in specific growing areas (Syvertsen and Lloyd, 1994), where drought/low temperature and high temperature/high VPDL are the strongest environmental constraints during the winter and summer season, respectively. Currently, the limited comprehension of physiological mechanisms triggered by environmental constraints in field-grown citrus trees delays and even impedes the advance of models for predicting crop production and techniques for improving productivity of citrus orchards under subtropical conditions. As a stimulating question, we may ask what is the relative importance of low temperatures (in both soil and air) in relation to seasonal drought in the regulation of citrus photosynthesis under subtropical conditions? An additional question: how important is the understanding of water relations in field-grown citrus plants? The knowledge of how plants respond to the seasonal variation of water availability is essential in order to improve and generate effective strategies for maintaining reproductive/vegetative development and/or reducing negative impacts of seasonal drought on citrus transpiration and photosynthesis. As an example, the irrigation management for increasing citrus flowering and thus fruit production based on shoot water potential has been studied under field conditions (Pires et al., unpublished data). A general model for water management in citrus orchards is not a simple matter since (i) there are various citrus scion and rootstock combinations (Medina et al., 1998, 2005; Machado et al., 2002; Barry et al., 2004; Cerqueira et al., 2004) and (ii) the environmental conditions differ significantly among growing areas and show considerable year-to-year variation even in a same region (Ribeiro et al., 2006a).
Another relevant point is that citrus leaves have a distinct form of exposure to sunlight, which suggests sun-exposed leaves are likely to be more constrained by high VPDL and high temperature than leaves in deep layers of the plant canopy. In addition, a large number of leaves are not exposed to sunlight and probably have different photosynthetic characteristics when compared to exposed leaves. Therefore, one should consider the non-exposed leaves to evaluate citrus canopy photosynthesis and its relation to citrus production, which will probably improve photosynthesis models for field-grown citrus. In relation to the 'hidden' citrus canopy, the photosynthetic traits such as leaf gas exchange, photochemical reactions and carbohydrate dynamics are poorly understood in mature plants under subtropical conditions. Considering the leaf reserves, we believe that this topic should be addressed from a systemic point of view, with attention to the seasonal patterns of carbohydrate production/consumption and their influence on citrus photosynthesis under natural environmental conditions. Such studies should also consider the crop load and plant growth in 'on' and 'off' years, when significant changes can be expected in the annual carbohydrate dynamics in citrus organs (Goldschmidt and Golomb, 1982).
In conclusion, the ecophysiology of citrus trees is a multi disciplinary task, in which only a cooperative effort among specialists of various areas, such as crop science and physiology, soil and irrigation sciences, agricultural meteorology and molecular biology, will improve our knowledge about the interaction of citrus plants with their surrounding environment under subtropical conditions.
Acknowledgments: The authors gratefully acknowledge the Fundação de Amparo à Pesquisa do Estado de São Paulo (Fapesp, Brazil) for the continuous financial support (since 1995 grants nº 95/8956-6, 98/16259-1, 00/02325-4, 02/08480-7, 03/02281-5, 04/06203-1 and 05/57862-8). E.C. Machado is also grateful to the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, Brazil) for the fellowship granted. Both authors are indebted to the Estação Experimental de Citricultura de Bebedouro (Brazil) and the Centro Apta Citros "Sylvio Moreira"/IAC (Brazil) for the use of experimental fields and facilities.
Received: 14 November 2007; Accepted: 12 December 2007
- Ahrens MJ, Ingram DL (1988) Heat tolerance of citrus leaves. Hort Science 23:747-748.
- Allen DJ, Ort DR (2001) Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends Plant Sci. 6:36-42.
- Allen DJ, Ratner K, Giller YE, Gussakovsky EE, Shahak Y, Ort DR (2000) An overnight chill induces a delayed inhibition of photosynthesis at midday in mango (Mangifera indica L.). J. Exp. Bot. 51:1893-1902.
- Angelocci LR, Marin FR, Oliveira RF, Righi EZ (2004) Transpiration, leaf diffusive conductance, and atmospheric water demand relationship in an irrigated acid lime orchard. Braz. J. Plant Physiol. 16:53-64.
- Azcón-Bieto J (1983) Inhibition of photosynthesis by carbohydrates in wheat leaves. Plant Physiol. 73:681-686.
- Baker NR, Rosenqvist E (2004) Application of chlorophyll fluorescence can improve crop production strategies: an examination of future possibilities. J. Exp. Bot. 55:1607-1621.
- Baker NR, Harbinson J, Kramer DM (2007) Determining the limitations and regulation of photosynthetic energy transduction in leaves. Plant Cell Environ. 30:1107-1125.
- Barry GH, Castle WS, Davies FS (2004) Rootstocks and plant water relations affect sugar accumulation of citrus fruit via osmotic adjustment. J. Am. Soc. Hort. Sci. 129:881-889.
- Berry J, Björkman O (1980) Photosynthetic response and adaptation to temperature in higher plants. Annu. Rev. Plant Physiol. 31:491-543.
- Bevington KB, Castle WS (1985) Annual root growth pattern of young citrus trees in relation to shoot growth, soil temperature and soil water content. J. Am. Soc. Hort. Sci. 110:840-845.
- Bilger W, Schreiber U, Bock M (1995) Determination of the quantum efficiency of photosystem II and of non-photochemical quenching of chlorophyll fluorescence in the field. Oecologia 102:425-432.
- Blanke MM (2000) Repair mechanisms following photoinhibition in Citrus Acta Hort. 531:151-153.
- Brakke M, Allen Jr LH (1995) Gas exchange of Citrus seedlings at different temperatures, vapor-pressure deficits, and soil water contents. J. Am. Soc. Hort. Sci. 120:497-504.
- Bustan A, Goldschmidt EE (1998) Estimating the cost of flowering in a grapefruit tree. Plant Cell Environ. 21:217-224.
- Ciiagro - Centro Integrado de Informações Agrometeorológicas. Boletim Agrometeorológico SP - Available in: <http://www.ciiagro.sp.gov.br/>. Accessed in: 10 Feb. 2007.
- Cerqueira EC, Castro Neto T, Peixoto CP, Soares Filho WS, Ledo CAS, Oliveira JG (2004) Resposta de porta-enxertos de citros ao déficit hídrico. Rev. Bras. Frutic. 26:515-519.
- Cohen S, Cohen Y (1983) Field studies of leaf conductance response to environmental variables in citrus. J. Appl. Ecol. 20:561-570.
- Cohen S, Fuchs M (1987) The distribution of leaf area, radiation, photosynthesis and transpiration in a Shamouti orange hedgerow orchard. Part I. Leaf area and radiation. Agric. For. Meteorol. 40:123-144.
- Cohen S, Moreshet S, Le Guilou L, Simon J, Cohen M (1997) Response of citrus trees to modified radiation regime in semi-arid conditions. J. Exp. Bot. 48:35-44.
- Critchley C (1998) Photoinhibition. In: Raghavendra AS (ed), Photosynthesis: a Comprehensive Treatise, pp.264-272. Cambridge University Press, Cambridge.
- Davenport TL (1990) Citrus flowering. Hort. Rev. 12:349-408.
- Davies FS, Albrigo LG (1994) Citrus. CAB International, Wallingford.
- Davies WJ, Zhang J (1991) Root signals and the regulation of growth and development of plants in drying soils. Annu. Rev. Plant Physiol. Plant Mol. Biol. 42:55-76.
- Demmig-Adams B, Adams WW (1996) The role of xanthophyll cycle carotenoids in the protection of photosynthesis. Trends Plant Sci. 1:21-26.
- Eissenstat DM (1991) On the relationship between specific root length and the rate of root proliferation: a field study using citrus rootstocks. New Phytol. 118:63-68.
- Elfving DC, Kaufmann MR (1972) Diurnal and seasonal effects of environment on plant water relations and fruit diameter of citrus. J. Am. Soc. Hort. Sci. 97:566-570.
- Elfving DC, Kaufmann MR, Hall AE (1972) Interpreting leaf water potential measurements with a model of the soil-plant-atmosphere continuum. Physiol. Plant. 27:161-168.
- Foyer CH (1988) Feedback inhibition of photosynthesis through source-sink regulation in leaves. Plant Physiol. Biochem. 26:483-492.
- García-Luis A, Fornes F, Sanz A, Guardiola JL (1988) The regulation of flowering and fruit set in citrus: relationship with carbohydrate levels. Isr. J. Bot. 37:189-201.
- García-Luis A, Fornes F, Guardiola JL (1995) Leaf carbohydrates and flower formation in Citrus J. Am. Soc. Hort. Sci. 120:222-227.
- Georgieva K (1999) Some mechanisms of damage and acclimation of the photosynthetic apparatus due to high temperature. Bulg. J. Plant Physiol. 25:89-99.
- Goldschmidt EE (1999) Carbohydrate supply as a critical factor for citrus fruit development and productivity. HortScience 34:1020-1024.
- Goldschmidt EE, Golomb A (1982) The carbohydrate balance of alternate-bearing citrus trees and the significance of reserves for flowering and fruiting. J. Am. Soc. Hort. Sci. 107:206-208.
- Goldschmidt EE, Huber SC (1992) Regulation of photosynthesis by end-product accumulation in leaves of plants storing starch, sucrose, and hexose sugars. Plant Physiol. 99:1443-1448.
- Goldschmidt EE, Koch KE (1996) Citrus. In: Zamski E, Schaffer AA (eds), Photoassimilate Distribution in Plants and Crops: Source-Sink Relationships, pp.797-823. Marcel Dekker, New York.
- Gomes MMA, Lagôa AMMA, Machado EC, Medina CL (2003) Abscisic acid and indole-3-acetic acid contents in orange trees infected by Xylella fastidiosa and submitted to cycles of water stress. Plant Growth Regul. 39:263-270.
- González-Altozano P, Castel JR (1999) Regulated deficit irrigation in 'Clementina de Nules' citrus trees. I. Yield and fruit quality effects. J. Hort. Sci. Biotechnol. 74:706-713.
- Greene BA, Gerber JF (1967) Radiation energy distribution in citrus trees. Proc. Am. Soc. Hort. Sci. 90:77-85.
- Guo YP, Zhou HF, Zhang LC (2006) Photosynthetic characteristics and protective mechanisms against photooxidation during high temperature stress in two citrus species. Sci. Hort. 108:260-267.
- Habermann G (2004) Trocas gasosas, relações hídricas e hormônios vegetais em laranjeira-doce infectada com Xylella fastidiosa em três regiões do Estado de São Paulo. Botucatu, Universidade Estadual Paulista. PhD thesis.
- Habermann G, Machado EC, Rodrigues JD, Medina CL (2003a) CO2 assimilation, photosynthetic light response curves, and water relations of 'Pera' sweet orange plants infected with Xylella fastidiosa Braz. J. Plant Physiol. 15:79-87.
- Habermann G, Machado EC, Rodrigues JD, Medina CL (2003b) Gas exchange rates at different vapor pressure deficits and water relations of 'Pera' sweet orange plants with citrus variegated chlorosis (CVC). Sci. Hort. 98:233-245.
- Hällgren J-E, Strand M, Lundmark T (1991) Temperature stress. In: Raghavendra AS (ed), Physiology of Trees, pp.301-335. John Wiley, New York.
- Horton P, Ruban AV, Walters RG (1996) Regulation of light harvesting in green plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 47:655-684.
- Hu LM, Xia RX, Xiao ZY, Huang RH, Tan ML, Wang MY, Wu QS (2007) Reduced leaf photosynthesis at midday in citrus leaves growing under field or screenhouse conditions. J. Hort. Sci. Biotechnol. 82:387-392.
- Huang T-B, Darnell RL, Koch KE (1992) Water and carbon budgets of developing citrus-fruit. J. Am. Soc. Hort. Sci. 117:287-293.
- Iglesias DJ, Lliso I, Tadeo FR, Talon M (2002) Regulation of photosynthesis through source-sink imbalance in citrus is mediated by carbohydrate content in leaves. Physiol. Plant. 116:563-572.
- Iglesias DJ, Tadeo FR, Primo-Millo E, Talon M (2003) Fruit dependence on carbohydrate availability in citrus trees. Tree Physiol. 23:199-204.
- Jifon JL, Syvertsen JP (2003) Moderate shade can increase net gas exchange and reduce photoinhibition in citrus leaves. Tree Physiol. 23:119-127.
- Jones WW, Steinacker ML (1951) Seasonal changes in concentration of sugars and starch in leaves and twigs of citrus trees. Proc. Am. Soc. Hort. Sci. 58:1-4.
- Jordan DB, Ogren WL (1984) The CO2/O2 specificity of ribulose-1,5-bisphosphate carboxylase/oxygenase: dependence on ribulose-bisphosphate concentration, pH and temperature. Planta 161:308-313.
- Khairi MM, Hall AE (1976) Comparative studies of net photosynthesis and transpiration of some citrus species and relatives. Physiol. Plant. 36:35-39.
- Li CY, Weiss D, Goldschmidt EE (2003) Effects of carbohydrate starvation on gene expression in citrus root. Planta 217:11-20.
- Lloyd J, Howie H (1989) Salinity, stomatal responses and whole tree conductivity of orchard Washington Navel orange. Aust. J. Plant Physiol. 16:169-179.
- Lloyd J, Syvertsen JP, Kriedemann PE, Farquhar GD (1992) Low conductances for CO2 diffusion from stomata to the sites of carboxylation in leaves of woody species. Plant Cell Environ. 15:873-899.
- Machado EC, Medina CL, Gomes MMA (1999) Teor de água no substrato de crescimento e fotossíntese em laranjeira 'Valência'. Bragantia 58:217-226.
- Machado EC, Medina CL, Gomes MMA, Habermann G (2002) Variação sazonal da fotossíntese, condutância estomática e potencial da água na folha de laranjeira 'valência'. Sci. Agric. 59:53-58.
- Machado EC, Schmidt PT, Medina CL, Ribeiro RV (2005) Respostas da fotossíntese de três espécies de citros a fatores ambientais. Pesq. Agropec. Bras. 40:1161-1170.
- Machado EC, Oliveira RF, Ribeiro RV, Medina CL, Stuchi ES, Pavani LC (2007) Deficiência hídrica agrava os sintomas fisiológicos da clorose variegada dos citros em laranjeira 'Natal'. Bragantia 66:373-379.
- Machado EC, Oliveira RF, Ribeiro RV, Medina CL, Stuchi ES, Marin FR, Silva JAB, Silva SR (2006) Fluxo de seiva e fotossíntese em laranjeira 'Natal' com clorose variegada dos citros. Pesq. Agropec. Bras. 41:911-918.
- Maxwell K, Johnson GN (2000) Chlorophyll fluorescence Exp. Bot. 54:539-547.
- Pimentel C, Ribeiro RV, Santos MG, Oliveira RF, Machado EC (2004) Effects of changes in the photosynthetic photon flux density on net gas exchange of Citrus sinensis and Nicotiana tabacum Braz. J. Plant Physiol. 16:77-82.
- Prado AKS, Machado EC, Medina CL, Machado DFSP, Mazzafera P (2007) Florescimento e frutificação em laranjeiras 'Valência' com diferentes cargas de frutos e submetidas ou não à irrigação. Bragantia 66:173-182.
- Rana G, Katerji N, De Lorenzi F (2005) Measurement and modelling of evapotranspiration of irrigated citrus orchard under Mediterranean conditions. Agric. For. Meteorol. 128:199-209.
- Raveh E, Cohen S, Raz T, Yakir D, Grava A, Goldschmidt EE (2003) Increase growth of young citrus trees under reduced radiation load in a semi-arid climate. J. Exp. Bot. 54:365-373.
- Ribeiro RV (2006) Variação sazonal da fotossíntese e relações hídricas de laranjeira 'Valência'. Piracicaba, Universidade de São Paulo. PhD thesis.
- Ribeiro RV, Machado EC, Oliveira RF (2003a) Early photosynthetic responses of sweet orange plants infected with Xylella fastidiosa Physiol. Mol. Plant Pathol. 62:167-173.
- Ribeiro RV, Machado EC, Oliveira RF (2004) Growth- and leaf-temperature effects on photosynthesis of sweet orange seedlings infected with Xylella fastidiosa Plant Pathol. 53:334-340.
- Ribeiro RV, Machado EC, Santos MG (2005a) Leaf temperature in sweet orange plants under field conditions: influence of meteorological elements. Rev. Bras. Agrometeorol. 13:353-368.
- Ribeiro RV, Machado EC, Brunini O (2006a) Ocorrência de condições ambientais para a indução do florescimento de laranjeiras no Estado de São Paulo. Rev. Bras. Frutic. 28:247-253.
- Ribeiro RV, Machado EC, Oliveira RF (2006b) Temperature response of photosynthesis and its interaction with light intensity in sweet orange leaf discs under non-photorespiratory condition. Ciênc. Agrotecnol. 30:670-678.
- Ribeiro RV, Machado EC, Oliveira RF, Pimentel C (2003b) High temperature effects on the response of photosynthesis to light in sweet orange plants infected with Xylella fastidiosa Braz. J. Plant Physiol. 15:89-97.
- Ribeiro RV, Machado EC, Santos MG, Oliveira RF (2005b) Variação sazonal e diária do conteúdo de carboidratos em folhas de laranjeira 'Valência'. In: Anais of the X Congresso Brasileiro de Fisiologia Vegetal and the XII Congresso Latino Americano de Fisiologia Vegetal, Recife, Brazil (Extended abstract on CD-ROM).
- Ruiz R, García-Luis A, Monerri C, Guardiola JL (2001) Carbohydrate availability in relation to fruitlet abscission in Citrus Ann. Bot. 87:805-812.
- Savé R, Biel C, Domingo R, Ruiz-Sánchez MC, Torrecillas A (1995) Some physiological and morphological characteristics of citrus plants for drought tolerance. Plant Sci. 110:167-172.
- Schaffer AA, Lin KC, Goldschmidt EE, Boyer CD, Goren R (1986) Citrus leaf chlorosis induced by sink removal: starch, nitrogen and chloroplast ultrastructure. J. Plant Physiol. 124:111-121.
- Schreiber U, Bilger W (1987) Rapid assessment of stress effects on plant leaves by chlorophyll fluorescence measurements. In: Tenhunen JD, Catarino FM, Lange OL, Oechel WC (eds), Plant Response to Stress, pp.27-53. Springer-Verlag, Berlin.
- Silva CR, Folegatti MV, Silva TJA, Alves Jr. J, Souza CF, Ribeiro RV (2005) Water relations and photosynthesis as criteria for adequate irrigation management in 'Tahiti' lime trees. Sci. Agric. 62:415-422.
- Smillie RM, Nott R (1979) Heat injury in leaves of alpine temperature and tropical plants. Aust. J. Plant Physiol. 6:135-141.
- Smith AM, Stitt M (2007) Coordination of carbon supply and plant growth. Plant Cell Environ. 30:1126-1149.
- Smith SM, Fulton DC, Chia T, Thorneycroft D, Chapple A, Dunstan H, Hylton C, Zeeman SC, Smith AM (2004) Diurnal changes in the transcriptome encoding enzymes of starch metabolism provide evidence for both transcriptional and post-transcriptional regulation of starch metabolism in Arabidopsis leaves. Plant Physiol. 136:2687-2699.
- Souza GM, Ribeiro RV, Pincus SM (2004) Changes in network connectance and temporal dynamics of gas exchange in Citrus sinensis under different evaporative demands. Braz. J. Plant Physiol. 16:119-130.
- Spiegel-Roy P, Goldschmidt EE (1996) Biology of Citrus. Cambridge University Press, Cambridge.
- Syvertsen JP (1984) Light acclimation in citrus leaves: II. CO2 assimilation and light, water, and nitrogen use efficiency. J. Am. Soc. Hort. Sci. 109:812-817.
- Syvertsen JP (1994) Partial shoot removal increases net CO2 assimilation and alters water relations of Citrus seedlings. Tree Physiol. 14:497-508.
- Syvertsen JP, Albrigo LG (1980a) Seasonal and diurnal citrus leaf and fruit water relations. Bot. Gaz. 141:440-446.
- Syvertsen JP, Albrigo LG (1980b) Some effects of grapefruit tree canopy position on microclimate, water relations, fruit yield, and juice quality. J. Am. Soc. Hort. Sci. 105:454-459.
- Syvertsen JP, Lloyd J (1994) Citrus. In: Schaffer B, Andersen PC (eds), Handbook of Environmental Physiology of Fruit Crops: Sub-Tropical and Tropical Crops, pp.65-99. CRC Press, Boca Raton.
- Syvertsen JP, Smith Jr ML (1984) Light acclimation in citrus leaves. I. Changes in physical characteristics, chlorophyll, and nitrogen content. J. Am. Soc. Hort. Sci. 109:807-812.
- Syvertsen JP, Smith Jr. ML, Allen C (1981) Growth rate and water relations of citrus leaf flushes. Ann. Bot. 47:97-105.
- Syvertsen JP, Goñi C, Otero A (2003) Fruit load and canopy shading affect leaf characteristics and net gas exchange of 'Spring' navel orange trees. Tree Physiol. 23:899-906.
- Syvertsen JP, Zablotowicz RM, Smith Jr. ML (1983) Soil temperature and flooding effects on two species of citrus. 1. Plant growth and hydraulic conductivity. Plant Soil 72:3-12.
- Tardieu F, Simonneau T (1998) Variability among species of stomatal control under fluctuating soil water status and evaporative demand: modelling isohydric and anisohydric behaviours. J. Exp. Bot. 49:419-432.
- Veselova SV, Farhutdinov RG, Veselov SY, Kudoyarova GR, Veselov DS, Hartung W (2005) The effect of root cooling on hormone content, leaf conductance and root hydraulic conductivity of durum wheat seedlings (Triticum durum L.). J. Plant Physiol. 162:21-26.
- Veste M, Ben-Gal A, Shani U (2000) Impact of thermal stress and high VPD on gas exchange and chlorophyll fluorescence of Citrus grandis under desert conditions. Acta Hort. 531:143-149.
- Vu JCV (1999) Photosynthetic responses of citrus to environmental changes. In: Pessarakli M (ed), Handbook of Plant and Crop Stress, pp.947-961. Marcel Dekker, New York.
- Vu JCV, Yelenosky G (1987) Photosynthetic characteristics in leaves of 'Valencia' orange (Citrus sinensis (L.) Osbeck) grown under high and low temperature regimes. Environ. Exp. Bot. 27:279-287.
- Vu JCV, Yelenosky G (1988a) Water deficit and associated changes in some photosynthetic parameters in leaves of 'Valencia' orange (Citrus sinensis osbeck). Plant Physiol. 88:375-378.
- Vu JCV, Yelenosky G (1988b) Solar irradiance and drought stress effects on the activity and concentration of ribulose bisphosphate carboxylase in 'Valencia' orange leaves. Isr. J. Bot. 37:245-256.
- Vu JCV, Yelenosky G (1989) Non-structural carbohydrate concentrations in leaves of 'Valencia' orange subjected to water deficits. Environ. Exp. Bot. 29:149-154.
- Vu JCV, Yelenosky G, Bausher MG (1986) CO2 exchange rate, stomatal conductance and transpiration in attached leaves of Valencia orange. HortScience 21:143-144.
- Wan X, Landhäusser SM, Zwiazek JJ, Lieffers VJ (2004) Stomatal conductance and xylem sap properties of aspen (Populus tremuloides) in response to low soil temperature. Physiol. Plant. 122:79-85.
- Zeiger E, Farquhar GD, Cowan IR (1987) Stomatal Function. Stanford University Press, Stanford.
Publication Dates
-
Publication in this collection
05 May 2008 -
Date of issue
Dec 2007
History
-
Accepted
12 Dec 2007 -
Received
14 Nov 2007