Abstract
The pharmacological effects of Bothrops neuwiedi pauloensis venom on mouse phrenic nerve-diaphragm (PND) preparations were studied. Venom (20 mug/ml) irreversibly inhibited indirectly evoked twitches in PND preparations (60 ± 10% inhibition, mean ± SEM; p<0.05; n=6). At 50 mug/ml, the venom blocked indirectly and directly (curarized preparations) evoked twitches in mouse hemidiaphragms. In the absence of Ca2+, venom (50 mug/ml), produced partial blockade only after an 80 min incubation, which reached 40.3 ± 7.8% (p<0.05; n=3) after 120 min. Venom (20 mug/ml) increased (25 ± 2%, p< 0.05) the frequency of giant miniature end-plate potentials in 9 of 10 end-plates after 30 min and the number of miniature end-plate potentials which was maximum (562 ± 3%, p<0.05) after 120 min. During the same period, the resting membrane potential decreased from - 81 ± 1.4 mV to - 41.3 ± 3.6 mV 24 fibers; p<0.01; n=4) in the end-plate region and from - 77.4 ± 1.4 to -44.6 ± 3.9 mV (24 fibers; p<0.01; n=4) in regions distant from the end-plate. These results indicate that B. n. pauloensis venom acts primarily at presynaptic sites. They also suggest that enzymatic activity may be involved in this pharmacological action.
giant MEPPs; myotoxicity; neurotoxicity; presynaptic action
ORIGINAL PAPERS
Neuromuscular activity of Bothrops neuwiedi pauloensis snake venom in mouse nerve-muscle preparations
Durigon A. M.I; Borja-Oliveira C. R.I; Dal Belo C. A.I; Oshima-Franco Y.I; Cogo J. C.II; Lapa A. J.III; Souccar C.III; Rodrigues-Simioni L.I
IDepartamento de Farmacologia, Faculdade de Ciências Médicas, Universidade Estadual de Campinas (UNICAMP), Campinas, SP, Brasil
IIInstituto de Pesquisa e Desenvolvimento (IP&D), Universidade do Vale do Paraíba (UNIVAP), São José dos Campos, SP, Brasil
IIIDepartamento de Farmacologia, Escola Paulista de Medicina, Universidade Federal de São Paulo (UNIFESP), São Paulo, SP, Brasil
Correspondence Correspondence to L. Rodrigues Simioni Departamento de Farmacologia, Faculdade de Ciências Médicas, Universidade Estadual de Campinas (UNICAMP) Cidade Universitária Zeferino Vaz Caixa Postal 6111, 13083-970 Campinas, SP, Brasil Phone: 55-19-3788-9534; Fax: 55-19-3289-2968 Email: simioni@obelix.unicamp.br
ABSTRACT
The pharmacological effects of Bothrops neuwiedi pauloensis venom on mouse phrenic nerve-diaphragm (PND) preparations were studied. Venom (20 mg/ml) irreversibly inhibited indirectly evoked twitches in PND preparations (60 ± 10% inhibition, mean ± SEM; p<0.05; n=6). At 50 mg/ml, the venom blocked indirectly and directly (curarized preparations) evoked twitches in mouse hemidiaphragms. In the absence of Ca2+, venom (50 mg/ml), produced partial blockade only after an 80 min incubation, which reached 40.3 ± 7.8% (p<0.05; n=3) after 120 min. Venom (20 mg/ml) increased (25 ± 2%, p< 0.05) the frequency of giant miniature end-plate potentials in 9 of 10 end-plates after 30 min and the number of miniature end-plate potentials which was maximum (562 ± 3%, p<0.05) after 120 min. During the same period, the resting membrane potential decreased from 81 ± 1.4 mV to 41.3 ± 3.6 mV 24 fibers; p<0.01; n=4) in the end-plate region and from 77.4 ± 1.4 to 44.6 ± 3.9 mV (24 fibers; p<0.01; n=4) in regions distant from the end-plate. These results indicate that B. n. pauloensis venom acts primarily at presynaptic sites. They also suggest that enzymatic activity may be involved in this pharmacological action.
Key words: giant MEPPs, myotoxicity, neurotoxicity, presynaptic action.
INTRODUCTION
According to the Brazilian Ministry of Health, from 1990 to 1993, bites by Bothrops spp. accounted for 90.5% of the accidents involving snakes in which the genus was known or suspected and for about 70% venomous snakebites in Brazil (14). The main complications in lethal cases are acute renal failure, shock, acute respiratory failure, and sepsis (2, 5, 6, 15). In some cases, respiratory failure has been associated with pulmonary edema (15). Although Bothrops venoms do not produce signs of neurotoxicity after snakebite, in recent years venoms of several Bothrops species, including B. jararacussu (17), B. insularis (8, 9), B. neuwiedi (3, 4, 21, 22), and B. pirajai (10) have been found to affect neuromuscular transmission in avian and mammalian isolated nerve-muscle preparations and in chick in vivo.
B. neuwiedi venom caused head-drop, loss of balance and respiratory failure in chicks in vivo, and inhibited twitch-tension response in biventer cervicis preparations, without decreasing responses to acetylcholine or KCl, all of which suggested a presynaptic action (21). More recently, Borja-Oliveira et al. (3) investigated the neurotoxicity of several lots of B. neuwiedi venoms in biventer cervicis and observed that not all lots exhibited neurotoxicity in vitro. B. n. pauloensis venom neurotoxicity in these preparations is temperature-dependent (4). In this work, the initial observations on neuromuscular action of B. n. pauloensis venom in chick preparations were extended using myographic and electrophysiological techniques in mouse nerve-muscle preparations.
MATERIALS AND METHODS
Venom and reagents
B. n. pauloensis venom, collected from snakes caught in São Paulo State, was provided by the Instituto Butantan (São Paulo, SP, Brazil). D-tubocurarine chloride was from Abbot Laboratórios do Brasil Ltda and neostigmine methylsulfate from Roche (Basel, Switzerland).
Mouse phrenic nerve-diaphragm preparation
Adult male Swiss white mice (28-35 g) were supplied by the University Animal House. Phrenic nerve and diaphragm were obtained from mice anesthetized with chloral hydrate (300 mg/kg, IP) and sacrificed by exsanguination. Nerve-muscle preparation was mounted as described by Bülbring (7). Hemidiaphragms and phrenic nerves were mounted in 5 ml tissue baths containing Tyrode solution (pH 7.4, 37° C) of the following composition (mM): NaCl 137, KCl 2.7, CaCl2 1.8, MgCl2 0.49, NaH2PO4 0.42, NaHCO3 11.9 and glucose 11.1. The preparations were mounted on an electrode that enabled indirect (supramaximal voltage, 0.1 Hz, 0.2 ms) and direct (50 V, 0.1 Hz, 2 ms) muscle stimulation with impulses delivered from a Grass S4 stimulator. The resulting muscle tension was recorded using a force displacement transducer (BG 25 GM Kulite) coupled to a Gould RS 3400 recorder. The preparation was allowed to stabilize for at least 15 min before venom addition (10, 20, 50, or 100 mg/ml). In some cases, neostigmine (7.4 mM) was used to reverse venom effects. Before direct stimulation, the preparations were curarized (d-tubocurarine, 7.3 mM). In some experiments, the Ca2+ (1.8 mM) in the Tyrode solution was replaced with Sr2+ (4 mM).
Electrophysiology
Membrane resting potentials and miniature end-plate potentials (MEPPs) were recorded in mouse hemidiaphragm muscle using conventional microelectrode techniques. The dissected muscle was mounted in a lucite chamber containing aerated (95% O2, 5% CO2) Tyrode solution (pH 7.4) at 30° C. The resting potential and MEPPs were recorded using glass microelectrodes filled with 3 M KCl (resistance 10-20 MW ) inserted into the muscle fiber and the recordings were displayed on a Tektronix oscilloscope. Measurements were obtained 15, 30, 60, 90, and 120 min after the addition of venom (20 mg/ml) and were recorded on a Gould RS 3400. The resting potential was measured at or distant from the end-plate regions.
Statistical analysis
Each experiment was repeated at least three times. The results were expressed as mean ± S.E.M. Student's t-test was used for statistical analysis of data. Values of p < 0.05 were considered significant.
RESULTS
Venom effect on indirectly and directly stimulated mouse diaphragm
B. n. pauloensis venom (20-100 mg/ml) produced concentration-dependent neuromuscular blockade in indirectly stimulated mouse nerve-muscle preparation (Table 1 and Figure 1). At 20 mg/ml, the venom produced 60 ± 10% blockade (p<0.05; n = 6) after 120 min (Figure 1A, B); a concentration of 50 mg/ml or 100 mg/ml caused complete twitch-tension blockade (Figure 1A, C, D). In the absence of Ca2+, venom (50 mg/ml), produced partial blockade only after an 80 min incubation, which reached 40.3 ± 7.8% (p<0.05; n = 3) after 120 min. In Ca2+-free solution, the results differed significantly (p<0.05) from that obtained in experiments with Ca2+ after 35 min incubation with venom (Figure 2). There were no venom-induced contractures. The twitch inhibition was not reverted by neostigmine (not shown) or by several washes with Tyrode solution (Figure 1B-D). No significant neuromuscular blockade was observed at concentrations < 20 mg/ml (not shown).
Venom concentration of 50 mg/ml inhibited contractions induced by direct muscle stimulation in curarized preparations (80 ± 8% inhibition after 120 min; p<0.05; n = 6) (Figure 3B). No significant neuromuscular blockade was observed at concentrations < 50 mg/ml (Figure 3A).

Venom effect (20 mg/ml) on the resting membrane potential and MEPPs
Venom caused a decrease in the resting membrane potential from 81 ± 1.4 mV to 41.3 ± 3.6 mV (24 fibers; p<0.01; n = 4) in the end-plate region and from 77.4 ± 1.4 to 44.6 ± 3.9 mV (24 fibers; p<0.01; n = 4) in regions distant from the end-plate (Figure 4).
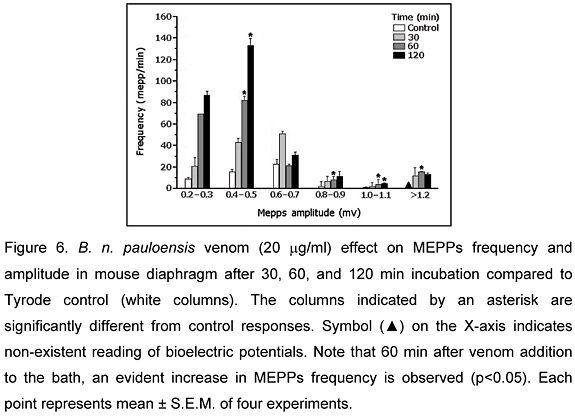
Changes in MEPPs amplitude and frequency were observed in preparations incubated with B. n. pauloensis venom when compared to preparations incubated with Tyrode solution (control). There was also an increase (25 ± 2%, p< 0.05) in the number of giant miniature end-plate potentials observed in 9 of 10 end-plates after a 30 min incubation (Figure 5). The intracellular recordings showed an increase in MEPPs frequency, which was maximum (562 ± 3%, p<0.05) after 120 min incubation with venom (20 mg/ml). There was also a visible increase in the frequency of 0.6-0.7 mV potentials after 30 min and 0.4-0.5 mV after 120 min, and the generation of MEPPs with large amplitude (>1.2 mV) during 120 min incubation, which was not observed in control experiments (Figure 6).
DISCUSSION
Although Bothrops venoms produce no clinical signs of neurotoxicity after snakebite, the venoms of several species can cause neuromuscular blockade in vitro and produce signs of peripheral muscular weakness in chicks. B. n. pauloensis venom produced neuromuscular blockade which was not reversed by neostigmine or washing the preparations. The MEPPs frequency of mouse isolated hemidiaphragm preparations was increased by venom and giant MEPPs were observed. Effects on muscle membrane were also observed, including the inhibition of muscle contractions in response to direct stimulation (seen at high venom concentrations - 50 mg/ml) and extrajunctional depolarizations in mouse preparations. These results suggest that B. n. pauloensis venom neuromuscular blocking action was caused mainly by neurotoxic presynaptic and myotoxic components.
B. neuwiedi venom neuromuscular activity has been the subject of previous reports. Soares et al. (21) described the effect of the venom and an isolated component, a Lys49 myotoxic phospholipase A2 homolog, on chick biventer cervicis preparations, in which inhibition of twitch-tension and KCl-induced contractures was observed. High concentrations of venom and of its myotoxic component were necessary to cause only partial neuromuscular blockade. Indeed, as reported by Harvey et al. (11), low venom concentrations frequently reveal the presence of neurotoxins, while high concentrations are required to demonstrate the presence of myotoxic components. Borja-Oliveira et al. (3) reported intraspecific variation in B. neuwiedi venom neuromuscular activity in chick biventer cervicis preparations based on 17 venom lots from various regions of southeastern Brazil. The samples which had the highest neuromuscular potency also had an additional electrophoretic band in relation to the other venoms and, at low concentrations, most of the venoms reduced the twitch-tension without abolishing the contracture to exogenous acetylcholine, suggesting a presynaptic action. B. n. pauloensis venom shows individual variation in its composition and there may be occasional complete lack of some toxins (16,21). Several myotoxic variants have been identified in B. neuwiedi venoms from different geographic regions (16).
B. neuwiedi venom is not the first of the Bothrops genus to show presynaptic activity. B. insularis venom also has a presynaptic action (8) which probably involves the Ca2+- and temperature-dependent presynaptic phospholipase A2 present in this venom (9). The observation that B. n. pauloensis venom neuromuscular effect was Ca2+-dependent indicated that enzymatic activity was necessary for neuromuscular action. Several divalent ions, including Sr2+, can bind to the same site, allowing neuromuscular transmission, but cannot substitute for Ca2+ in catalysis. For this reason, they are useful phospholipase A2 activity inhibitors (20).
Similar to B. insularis venom (8), B. n. pauloensis venom caused the early appearance of giant MEPPs that may be considered as the initial manifestation of a presynaptic action. This early agrees with the action of this venom on membrane resting potential seen at the same time (30 min). Crotoxin, the major neurotoxin from Crotalus durrissus terrificus venom, also causes an initial fall in MEPPs frequency in frog neuromuscular junction, followed by a secondary rise which was characterized by the appearance of large spontaneous potentials, i.e., giant MEPPs (12, 18). At frog neuromuscular junction, giant MEPPs are described as spontaneous potentials with amplitude of more than twice the modal MEPPs average and a slower rising phase (1). A recent comparative study about the pharmacological activities of B. insularis, B. neuwiedi, and C. d. terrificus in chick neuromuscular preparations (19) indicated that bothropic venoms may contain components which act presynaptically at the skeletal neuromuscular junction in a manner similar to C. d. terrificus venom and its main toxin, crotoxin.
The neuromuscular blocking action of some bothropic venoms reported in recent years appears to involve Ca2+-independent (B. jararacussu) and Ca2+-dependent (B. insularis and B. n. pauloensis) pathways. The first pathway is intriguing because high Ca2+ inhibits the neuromuscular blockade seen with bothropstoxin-I from B. jararacussu venom (13), the second pathway, which requires Ca2+ for its toxic action, may involve PLA2 activity of some as yet unidentified venom constituents. Thus, the classic concept of neurotoxicity involving mainly elapid and some crotalid venoms may need reexamination in view of the presynaptic actions of some bothropic venoms.
ACKNOWLEDGEMENTS
The authors thank Gildo Bernardo Leite for technical assistance and Instituto Butantan for providing the venom sample. This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), Conselho Nacional de Desenvolvimento Científico e Tecnlógico (CNPq), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and Fundação de Apoio ao Ensino e à Pesquisa (FAEP-UNICAMP).
Received: May 28, 2003
Accepted: August 11, 2003
Published online: February 11, 2005.
- 1 ALKADHI KA. Emetine increases giant miniature endplate potential population at the frog neuromuscular junction. Brain Res., 1988, 447, 293-8.
- 2 AMARAL CFS., DA SILVA OA., GODOY P., MIRANDA D. Renal cortical necrosis following Bothrops jararaca and B. jararacussu snake bite. Toxicon, 1985, 23, 877-85.
- 3 BORJA-OLIVEIRA CR., SOARES AM., ZAMUNER SR., HYSLOP S., GIGLIO JR., PRADO-FRANCESCHI J., RODRIGUES-SIMIONI L. Intraspecific variation in the neurotoxic and myotoxic activities of Bothrops neuwiedi snake venoms. J. Venom. Anim. Toxins, 2002, 8, 88-101.
- 4 BORJA-OLIVEIRA CR., DURIGON AM., VALLIN ACC., TOYAMA MH., SOUCCAR C., MARANGONI S., RODRIGUES-SIMIONI L. The pharmacological effects of Bothrops neuwiedi pauloensis (jararaca-pintada) snake venom on avian neuromuscular transmission. Braz. J. Med. Biol. Res, 2003, 36, 617-24.
- 5 BRASIL. Ministério da Saúde. Manual de diagnóstico e tratamento de acidentes por animais peçonhentos Brasília: Fundação Nacional de Saúde, 1998. 131p.
- 6 BUCARETCHI F., HERRERA SR., HYSLOP S., BARACAT EC., VIEIRA RJ. Snakebites by Crotalus durissus ssp in children in Campinas, São Paulo, Brazil. Rev. Inst. Med. Trop. São Paulo, 2002, 44, 133-8.
- 7 BÜLBRING E. Observations on the isolated phrenic nerve diaphragm preparation of the rat. Br. J. Pharmacol, 1946, 1, 38-61.
- 8 COGO JC., PRADO-FRANCESCHI J., CRUZ-HÖFLING MA., CORRADO AP., RODRIGUES-SIMIONI L. Effect of Bothrops insularis venom on the mouse and chick nerve-muscle preparation. Toxicon, 1993, 31, 1237-47.
- 9 COGO JC., PRADO-FRANCESCHI J., GIGLIO JR., CORRADO AP., CRUZ-HÖFLING MA., DONATO JL., LEITE GB., RODRIGUES-SIMIONI L. An unusual presynaptic action of Bothrops insularis snake venom mediated by phospholipase A2 fraction. Toxicon, 1998, 36, 1323-32.
- 10 COSTA PD., TOYAMA MH., MARANGONI S., RODRIGUES-SIMIONI L., CRUZ-HÖFLING MA. Effects of Bothrops pirajai venom on the mouse extensor digitorum longus (EDL) muscle preparation. Toxicon, 1999, 37, 1143-53.
- 11 HARVEY AL., BARFARAZ A., THOMPSON E., FAIZ A., PRESTON S. Screening of snake venoms for neurotoxic and myotoxic effects using simple in vitro preparations from rodents and chicks. Toxicon, 1994, 32, 257-65.
- 12 HAWGOOD BJ., SMITH ICH., STRONG PN. Early induction by crotoxin of biphasic frequency changes and giant miniature endplate potentials in frog muscle. Br. J. Pharmacol, 1988, 94, 765-72.
- 13 HELUANY NF., HOMSI-BRANDEBURGO MI., GIGLIO JR., PRADO-FRANCESCHI J., RODRIGUES-SIMIONI L. Effects induced by bothropstoxin, a component from Bothrops jararacussu snake venom, on mouse and chick muscle preparations. Toxicon, 1992, 30, 1203-10.
- 14 HOMSI-BRANDEBURGO MI., QUEIROZ LS., SANTO-NETO H., RODRIGUES-SIMIONI L., GIGLIO JR. Fractionation of Bothrops jararacussu snake venom: partial chemical characterization and biological activity of bothropstoxin. Toxicon, 1988, 26, 615-27.
- 15 RIBEIRO LA., ALBUQUERQUE MJ., DE CAMPOS VA., KATZ G., TAKAOKA NY., LEBRAO ML., JORGE MT. Óbitos por serpentes peçonhentas no Estado de São Paulo: avaliação de 43 casos, 1988/93. Rev. Assoc. Med. Bras, 1998, 44, 312-8.
- 16 RODRIGUES VM., SOARES AM., MANCIN AC., FONTES MRM., HOMSI-BRANDEBURGO MI., GIGLIO JR. Geopraphic variations in the composition of myotoxins from Bothrops neuwiedi snake venoms: biochemical characterization and biological activity. Comp. Biochem. Physiol. A, 1998, 121, 215-22.
- 17 RODRIGUES-SIMIONI L., BORGES EN., CECCARELLI B. The effects of Bothrops jararacussu venom and its components on frog nerve-muscle preparation. Neuroscience, 1983, 10, 475-89.
- 18 RODRIGUES-SIMIONI L., HAWGOOD BJ., SMITH ICH. Properties of the early phases of crotoxin poisoning at frog neuromuscular junctions. Toxicon, 1990, 28, 1479-89.
- 19 RODRIGUES-SIMIONI L., ZAMUNER S., COGO JC., BORJA-OLIVEIRA CR., PRADO-FRANCESCHI J., CRUZ-HÖFLING MA., CORRADO AP. Pharmacological evidence for a crotoxin-like action of venoms from Bothrops insularis (jararaca-ilhôa) and Bothrops neuwiedi (jararaca-pintada). Toxicon (in press).
- 20 SCHIAVO G., MATTEOLI M., MONTECUCCO C. Neurotoxins affecting neuroexocytosis. Physiol. Rev, 2000, 80, 717-66.
- 21 SOARES AM., GUERRA-SÁ R., BORJA-OLIVEIRA CR., RODRIGUES VM., RODRIGUES-SIMIONI L., RODRIGUES V., FONTES MRM., LOMONTE B., GUTIÉRREZ JM., GIGLIO JR. Structural and functional characterization of BnSP-7, a lys49 myotoxic phospholipase A2 homologue from Bothrops neuwiedi venom. Arch. Biochem. Biophys, 2000, 378, 201-9.
- 22 ZAMUNER SR., PRADO-FRANCESCHI J., RODRIGUES-SIMIONI L. The screening of Bothrops venoms for neurotoxic activity using the chick biventer cervicis preparation. Toxicon, 1996, 34, 314-5.
Publication Dates
-
Publication in this collection
03 May 2005 -
Date of issue
Mar 2005
History
-
Accepted
11 Feb 2005 -
Received
28 May 2003 -
Reviewed
11 Aug 2003