Abstract
Envenomations caused by different species of Bothrops snakes result in severe local tissue damage, hemorrhage, pain, myonecrosis, and inflammation with a significant leukocyte accumulation at the bite site. However, the activation state of leukocytes is still unclear. According to clinical cases and experimental work, the local effects observed in envenenomation by Bothrops alternatus are mainly the appearance of edema, hemorrhage, and necrosis. In this study we investigated the ability of Bothrops alternatus crude venom to induce macrophage activation. At 6 to 100 ¼g/mL, BaV is not toxic to thioglycollate-elicited macrophages; at 3 and 6 ¼g/mL, it did not interfere in macrophage adhesion or detachment. Moreover, at concentrations of 1.5, 3, and 6 ¼g/mL the venom induced an increase in phagocytosis via complement receptor one hour after incubation. Pharmacological treatment of thioglycollate-elicited macrophages with staurosporine, a protein kinase (PKC) inhibitor, abolished phagocytosis, suggesting that PKC may be involved in the increase of serum-opsonized zymosan phagocytosis induced by BaV. Moreover, BaV also induced the production of anion superoxide (O2_) by thioglycollate-elicited macrophages. This BaV stimulated superoxide production was abolished after treating the cells with staurosporine, indicating that PKC is an important signaling pathway for the production of this radical. Based on these results, we suggest that phagocytosis and reactive oxygen species are involved in the pathogenesis of local tissue damage characteristic of Bothrops spp. envenomations.
snake venom; Bothrops; macrophages; phagocytosis; superoxide; protein kinase C; inflammation
ORIGINAL PAPERS
Effect of Bothrops alternatus snake venom on macrophage phagocytosis and superoxide production: participation of protein kinase C
Setubal SSI; Pontes ASI; Furtado JLI; Kayano AMI; Stábeli RGI,II; Zuliani JPI,II
ILaboratory of Biochemistry and Biotechnology and Laboratory of Cell Culture and Monoclonal Antibodies, Tropical Pathology Research Institute (Ipepatro), Oswaldo Cruz Foundation (Fiocruz), Porto Velho, Rondônia State, Brazil
IICenter of Biomolecules Applied to Medicine, Department of Medicine, Federal University of Rondônia (UNIR), Porto Velho, Rondônia State, Brazil.
Correspondence to Correspondence to: Juliana Pavan Zuliani Instituto de Pesquisas em Patologias Tropicais Laboratório de Bioquímica e Biotecnologia Laboratório de Cultivo Celular e Anticorpos Monoclonais Rua da Beira, 7671, BR 364, km 3,5 Porto Velho, RO, 78912-000, Brasil Phone: +55 69 3219 6010 Fax: +55 69 3219 6000 Email: jzuliani@pq.cnpq.br or jzuliani@ipepatro.org.br.
ABSTRACT
Envenomations caused by different species of Bothrops snakes result in severe local tissue damage, hemorrhage, pain, myonecrosis, and inflammation with a significant leukocyte accumulation at the bite site. However, the activation state of leukocytes is still unclear. According to clinical cases and experimental work, the local effects observed in envenenomation by Bothrops alternatus are mainly the appearance of edema, hemorrhage, and necrosis. In this study we investigated the ability of Bothrops alternatus crude venom to induce macrophage activation. At 6 to 100 ¼g/mL, BaV is not toxic to thioglycollate-elicited macrophages; at 3 and 6 ¼g/mL, it did not interfere in macrophage adhesion or detachment. Moreover, at concentrations of 1.5, 3, and 6 ¼g/mL the venom induced an increase in phagocytosis via complement receptor one hour after incubation. Pharmacological treatment of thioglycollate-elicited macrophages with staurosporine, a protein kinase (PKC) inhibitor, abolished phagocytosis, suggesting that PKC may be involved in the increase of serum-opsonized zymosan phagocytosis induced by BaV. Moreover, BaV also induced the production of anion superoxide (O2_) by thioglycollate-elicited macrophages. This BaV stimulated superoxide production was abolished after treating the cells with staurosporine, indicating that PKC is an important signaling pathway for the production of this radical. Based on these results, we suggest that phagocytosis and reactive oxygen species are involved in the pathogenesis of local tissue damage characteristic of Bothrops spp. envenomations.
Key words: snake venom, Bothrops, macrophages, phagocytosis, superoxide, protein kinase C, inflammation.
INTRODUCTION
The majority of envenomations in Brazil are caused by species of the genus Bothrops from the Viperidae family (1). Bothrops snakes have received a large amount of attention due to the high incidence of accidents, with over 90% of officially reported cases (2). The incidence of snakebites reported between 2000 and 2007, was 1920, of which 27% (703 cases) were recorded in the northern region, 28% in the southeast, 24% in the northeast and respectively 11 and 10% in the south and central-west regions (1). These accidents are frequently related to climatic factors and human activity in rural areas where their concentration is higher (3-5).
Bothrops alternatus snake (now in the genus Rhinocerophis) is found in northern and northeastern Argentina, southern Paraguay and the southern, southeastern and western part of central Brazil (6, 7). Its preferred habitat is humid tropical, subtropical, and temperate forests. Its average size is about one meter long, but it can reach two meters.
The severity of poisoning caused by this specie can be partly explained by the large quantity of venom it produces. The maximum venom yield per specimen is 380 mg (dry weight) if compared with an average of 200 mg (dry weight) of Bothrops atrox, Bothrops insularis, Bothrops moojeni, and Bothrops jararacussu (8-10). In common with other Bothrops species, Bothrops alternatus venom contains a variety of proteins and enzymes including L-amino acid oxidase (LAAO), phospholipase A2 (PLA2), thrombin-like enzyme (such as balterobin), thrombin inhibitor (bothroalternin), metalloproteinases, disintegrins, and phosphodiesterases (11-23).
Furthermore, although Bothrops alternatus is aggressive and dangerous, its venom is not as well-known as other Bothrops venoms, especially Bothrops jararaca, which has been widely studied for its clinical importance in terms of snakebite incidence.
In fact, the local effects observed in B. alternatus envenomation, according to clinical cases and experimental studies, mainly refer to the appearance of edema, hemorrhage, and necrosis (24-27). The literature also shows that B. alternatus venom causes systemic disturbances in blood coagulation cascade in humans and other animals such as dogs, and 97% of accidents have alterations in coagulation times (24). Its toxicity is similar to other Bothrops species (9, 28).
Envenomation caused by different Bothrops species result in severe local tissue damage with hemorrhage, pain, myonecrosis, and inflammation (8, 29-33). In addition, Bothrops venoms induce a significant leukocyte accumulation at the bite site (32-35). However, the activation state of these cells is still unclear.
Macrophages are important cells of the innate immune system. They play a central role in a wide variety of processes associated with tissue maintenance, antigen presentation, inflammation, and tissue repair (36). Resident macrophages are one of the first lines of host defense and are found in many tissues. Upon stimulation, these quiescent cells become active and display diverse cellular functions such as the production of numerous active substances. As macrophage activation can play beneficial and deleterious roles in tissues, these phenomena may have an impact on the local pathological alterations induced by snake venoms.
One of the most important macrophage responses against infections is phagocytosis. This event consists of the uptake and destruction of invading microorganisms which is initiated by the engagement of receptors on the surface of phagocytes, macrophages, and neutrophils, which express a broad spectrum of receptors that participate in target cell recognition and internalization of ligands (37). Complement receptor type 3 (CR3, also called CD11b/CD18, Mac-1, and integrin αMβ2) recognizes complement-coated particles (37-41).
CR3 requires an additional activation signal for efficient particle binding and phagocytosis. These activation signals can be provided by a number of stimuli such as phorbol esters, lipopolysaccharides (LPS), cytokines, growth factors, and chemokines, which activate protein kinase C (PKC) and induce "inside-out" signaling to integrin receptors (42-46). The inside-out signaling relies on the small GTP-binding protein Rap1 and the actin-binding protein talin (47, 48). This signal is translated to changes in integrin tertiary structure and clustering, which enhances receptor affinity and valency (49, 50).
Subsequent to phagocytosis, one of the most immediate macrophage responses is abrupt production of the potent oxygen free radical, superoxide anion, referred to as oxidative burst. The enzyme complex primarily responsible for producing this highly reactive oxygen species is NADPH oxidase complex. In resting cells, the oxidase is dormant and its components exist separately in the membrane and cytosol. When macrophages are stimulated, these components associate on the plasma or phagosomal membranes to form the active oxidase complex (51, 52, 53). The resulting reactive oxygen species (ROS), such as superoxide anion and hydrogen peroxide, are used by macrophages to kill ingested microorganisms, and are thus an essential component of the innate immune system (51-53). This reaction parallels the release of a variety of inflammatory mediators that play crucial roles in host defense by microbial killing, but may also cause injury to surrounding tissues (36, 51, 52, 54).
Despite the well-documented importance of macrophages in tissue maintenance, antigen presentation, inflammation, and tissue repair, the activation state of these cells is still unclear in snake envenomation. This study was therefore designed to evaluate the effects of crude Bothrops alternatus venom (BaV) on isolated thioglycollate macrophages, particularly phagocytosis and superoxide anion production.
MATERIALS AND METHODS
Chemicals and Reagents
Trypan blue, RPMI-1640, L-glutamine, gentamicin, halothane, zymosan, staurosporine (Sts), phorbol myristate acetate (PMA), DMSO, Giemsa and nitroblue tetrazolium (NBT) were purchased from Sigma (USA). Fetal bovine serum (FBS) was obtained from Cultilab (Brazil). Low endotoxin or endotoxin-free grades of salts and reagents were obtained from Merck (Germany).
Animals and Venom
Venom from Bothrops alternatus was acquired from the Bioactive Protein Serpentarium, Batatais, São Paulo state, Brazil. Male Swiss mice (18-20 g) were housed in temperature-controlled rooms and received water and food ad libitum until experiments were carried out. Animal care was in accordance with the guidelines of the Brazilian College for Animal Experimentation (COBEA) and the present study was approved by the Tropical Pathology Research Institute Ethics Committee on Animal Use.
Harvesting of Macrophages
Thioglycollate-elicited macrophages were harvested four days after intraperitoneal (i.p.) injection of 1mL 3% thioglycollate. Animals were euthanized under halothane and exsanguined. Then, peritoneal lavage was performed, after a gentle massage of the abdominal wall, using 3mL cold phosphate-buffered saline (PBS, 14 mM NaCl, 2 mM NaH2PO4H2O, 7 mM Na2HPO412H2O, pH 7.2). Peritoneal fluid, containing thioglycollate-elicited macrophages, was collected. Total peritoneal cell counts were determined in a Neubauer's chamber. The cell population consisted of more than 95% macrophages, as determined by morphological criteria.
Cytotoxic Assay
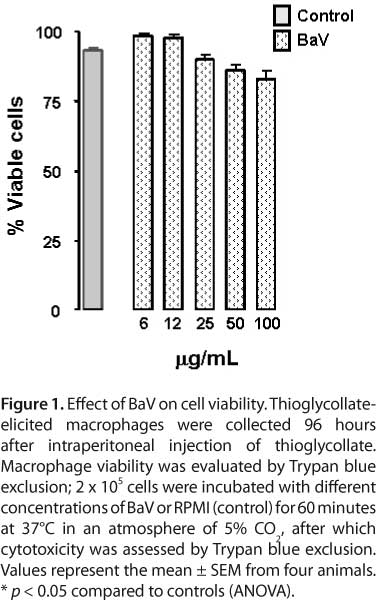
Cell viability was measured by Trypan blue exclusion according to Zuliani et al. (55). In brief, thioglycollate-elicited macrophages were centrifuged at 200 g for five minutes at 4°C and the cell pellets were resuspended in 1 mL of RPMI-1640 medium supplemented with 100 μg/mL of gentamicin, 2 mM L-glutamine and 10% fetal bovine serum (FBS). After counting, 2 x 105 macrophages/80 μL were added to plastic vials and incubated with 20 μL at different concentrations of venom from B. alternates (6, 12, 25, 50, and 100 μg/mL) diluted in assay medium or RPMI (control), for one hour at 37°C in a humidified atmosphere (5% CO2). Then, 20 μL of 0.1% Trypan blue was added to 100 μL of thioglycollate-elicited macrophage suspension. Viable cell index was determined in a Neubauer's chamber by counting a total of 100 cells. Results were expressed as percentage of viable cells.
Adhesion Assay
Macrophage adhesion was assayed according to the procedure described by Rosen and Gordon (56). Thioglycollate-elicited macrophages (2 x 105 cells/well) were cultured for 60 minutes with BaV (6 and 12 μg/mL), diluted in assay medium or RPMI (control) at 37°C in a humidified atmosphere (5% CO2). After incubation, the plates were washed three times with PBS and adherent cells fixed with methanol. After staining with 0.1% Giemsa solution for 40 minutes, the plates were washed with water, and the remaining dye was solubilized with methanol. Absorbance was determined spectrophotometrically at 550 nm. The control group was considered 100% adhered.
Detachment Assay
Thioglycollate-elicited macrophages were placed on 96-well plates at 2 x 105 cells/well to attach for 24 hours at 37°C in a humidified atmosphere (5% CO2). After adhesion, the adhered macrophages were incubated with BaV (6 and 12 μg/mL), diluted in assay medium or RPMI (control) for 60 minutes at 37°C in a humidified atmosphere (5% CO2). The end of the process was performed as previously described for the adhesion assay.
Phagocytic Activity of Elicited Peritoneal Macrophages
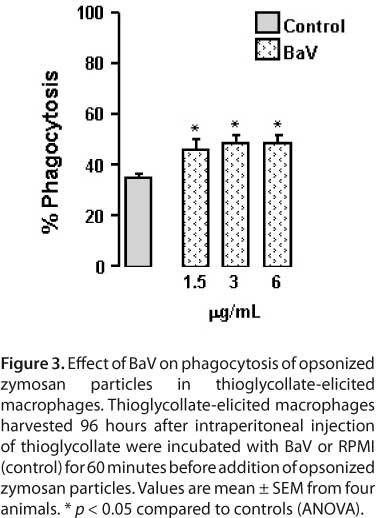
Phagocytic activity was determined according to Zuliani et al. (55). In brief, thioglycollate-elicited macrophages were placed on 13-mm diameter glass coverslips (Glass Tecnica, Brazil) in 24-well plates at 2 x 105 cells per coverslip and allowed to attach for 40 minutes at 37°C under a 5% CO2 atmosphere. Non-adherent cells were removed by washing with PBS. Cell monolayers were cultured for one hour with RPMI-1640 supplemented with 100 μg/mL of gentamicin, 2 mM L-glutamine and 10% FBS at 37°C and 5% CO2, and then challenged with RPMI (control) or different concentrations of BaV (1.5, 3, and 6 mg/mL) diluted in RPMI.
After washing in cold PBS, the monolayers were incubated for 40 minutes at 37°C and 5% CO2 with serum-opsonized zymosan, prepared as described below, and unbound particles were removed by washing with PBS. Cells were fixed with 2.5% glutaraldehyde for 15 minutes at room temperature and the coverslips were mounted on microscope slides. The extent of phagocytosis was quantified by contrast phase microscopic observation. At least 200 macrophages were counted in each determination and those containing three or more internalized particles were considered positive for phagocytosis. Results were presented as the percentage of phagocytosis positive cells.
To prepare serum-opsonized zymosan particles of zymosan obtained from yeast cell walls were suspended in PBS at 5.7 mg/mL. For opsonization, 2 mL of zymosan particles were mixed with 2 mL of normal mouse serum and incubated for 30 minutes at 37°C (57). The serum-opsonized zymosan particles were then centrifuged at 200 g for ten minutes and suspended in PBS for the phagocytosis assay.
Pharmacological Modulation of Phagocytosis
To assess the different signal transduction pathways involved in phagocytosis of serum-opsonized zymosan particles, thioglycollate-elicited peritoneal macrophages were treated with 14 nM/mL of staurosporine, a PKC inhibitor, 15 minutes before adding the venom or RPMI alone with the vehicle used for the inhibitor. Inhibitor concentration did not have an adverse effect on cell viability during the course of the assays, and was based on concentrations used elsewhere in the literature. Control cells were treated with the same concentration of the carrier in which the inhibitor had been dissolved.
Superoxide Anion Production Assay
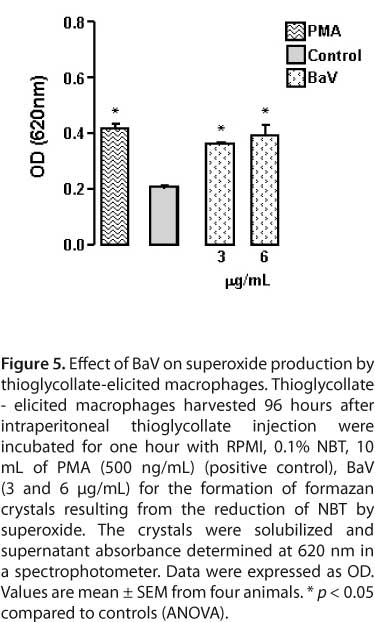
In this assay, superoxide generation was estimated by reducing nitroblue tetrazolium (NBT), a yellow liposoluble compound that becomes insoluble and blue in its reduced form (58). In brief, thioglycollate-elicited macrophages concentrations were adjusted to 2 x 105/100 ΜL and were incubated with 100 μL RPMI (control) or phorbol myristate acetate (PMA-positive control) containing NBT 0.1% or 100 μL of different BaV concentrations (3 and 6 μg/mL), diluted in RPMI containing NBT 0.1%, and incubated for 1h at 37°C in a humidified atmosphere (5% CO2). At the end of the incubation period, the vials were centrifuged for 30 seconds at 800 g and cells washed twice with PBS. The reduced NBT deposited inside the cells was then dissolved, first by adding 90 μL 2 M KOH to solubilize cell membranes, and then by adding 110 μL of DMSO to dissolve blue formazan with gentle shaking for ten minutes at room temperature. The dissolved NBT solution was then transferred to a 96-well plate and absorbance was read on a microplate reader at 620 nm. Data were expressed as absorbance.
Pharmacological Modulation of Superoxide Production
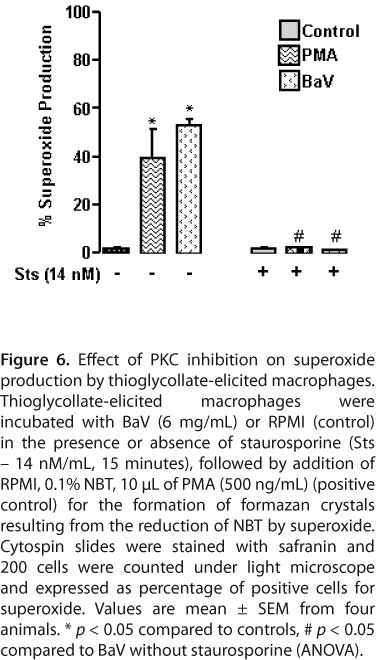
To assess the different signal transduction pathways involved in superoxide production, thioglycollate-elicited peritoneal macrophages were treated with 14 nM/mL of staurosporine 15 minutes before adding the venom or RPMI alone with the vehicle used for the inhibitor.
Statistical Analysis
Mean and standard error of the mean (SEM) for all data were obtained and compared by two-way ANOVA, followed by the Tukey's test with significance levels set at p < 0.05.
RESULTS
Effect of BaV on Macrophage Viability
BaV toxicity on isolated thioglycollate-elicited macrophages was investigated by one-hour incubation of several venom concentrations. Figure 1 shows that incubation of BaV at concentrations of 6 to 100 μg/mL did not affect thioglycollate-elicited peritoneal macrophages.
Effect of BaV on Macrophage Adhesion and Detachment
To examine whether BaV would interfere in adhesion and detachment of thioglycollate-elicited macrophages from support, adhesion and detachment capacity was tested on thioglycollate-elicited macrophages treated with non-cytotoxic concentrations of BaV or RPMI (control). Figure 2 shows that incubation of macrophages with BaV (3 and 6 μg/mL) did not interfere in thioglycollate-elicited macrophage adhesion or detachment.
Effect of BaV on Phagocytosis by Macrophages
To assess the ability of BaV to stimulate complement receptor-mediated phagocytosis, serum-opsonized zymosan particle uptake was determined in adherent thioglycollate-elicited macrophages treated with non-cytotoxic concentrations of BaV or RPMI (control). Figure 3 shows that average phagocytosis of serum-opsonized zymosan particles in thioglycollate-elicited macrophages incubated with RPMI was 35%. Macrophage incubation with BaV resulted in 48.58 ± 2.81%, 48.5 ± 3.2%, and 46 ± 4.02% of phagocytosis of serum-opsonized zymosan at concentrations of 1.5, 3 and 6 μg/mL, respectively. These values were significantly higher than controls.
Effect of PKC Inhibition on BaV-Induced Macrophage Phagocytosis of Serum-Opsonized Zymosan
Staurosporine was used to examine the role of PKC on signaling pathways of BaV-stimulated macrophage phagocytosis. The toxicity of staurosporine on isolated thioglycollate-elicited macrophages was tested by one-hour incubation. Results indicated that staurosporine at 14 nM/mL was not toxic to macrophages (data not shown). Figure 4 shows that thioglycollate-elicited macrophages incubated with RPMI had an average phagocytosis for serum-opsonized zymosan particles of 35%; this was significantly higher after stimulation with BaV at 6 μg/mL. Moreover, preincubation of macrophages with staurosporine significantly reduced the BaV-induced phagocytic index.
Effect of BaV on Superoxide (O2-) Production by Macrophages
To investigate the ability of BaV to induce the production of superoxide by thioglycollate-elicited macrophages, cells were incubated with non-cytotoxic concentrations of BaV or RPMI (control) in the presence of NBT. Figure 5 shows that thioglycollate-elicited macrophages incubated with RPMI produced basal superoxide. Incubation of macrophages with BaV at non-cytotoxic concentrations induced a significant production of O2- in thioglycollate-elicited macrophages.
Effect of PKC Inhibition on Macrophage Superoxide Production Induced by BaV
Staurosporine was used to examine the role of PKC on signaling pathways in BaV-stimulated macrophage superoxide production. Figure 6 shows thioglycollate-elicited macrophages incubated with RPMI produced basal superoxide, which was significantly higher after stimulation with BaV at 3 and 6 μg/mL. Preincubation of macrophages with staurosporine (14 nM/mL) significantly reduced BaV-induced superoxide production.
DISCUSSION
After a snake bite, venom enters tissues and affects resident cells in various ways. Firstly, direct cytotoxicity leading to necrosis is one output of envenomation. However, non-cytotoxic concentrations of snake venom may reach other cells; in these cases, other cellular responses may occur which are distinct from cell death, and may contribute to the observed tissue alterations.
One of the cells that may be reached by non-cytotoxic concentrations of snake venom, and may contribute to tissue homeostasis, is the macrophage. This cell is multifunctional and plays an important role in immune responses exerting their host defense function through various activities, which include phagocytosis, production of microbicidal agents, secretion of cytokines and other inflammatory mediators, and antigen processing and presentation.
Our initial results demonstrated that different concentrations of Bothrops alternatus venom were not toxic to thioglycollate-elicited macrophages. Cellular toxicity screening assays have shown that Bothrops neuwiedii mattogrossensis and Bothrops leucurus venoms presented higher toxicity than Bothrops atrox and Bothrops alternatus venoms for Vero cells (59). Moreover, Moreira et al. (60) verified that Bothrops asper venom only affects the viability of both macrophages and neutrophils at concentrations up to 8 μg/mL.
The macrophage is a multifunctional cell which plays an important role in inflammation by migrating from blood vessels and phagocytosing foreign agents. For effective occurrence of phagocytosis, macrophages must adhere to a substrate before modifying their cell morphology from a round to a flat or spread shape (61). We therefore investigated the ability of BaV to interfere in the adhesion and detachment of thioglycollate-elicited macrophages. Our results showed that BaV did not affect either the adhesion or detachment of these cells from their support. Based on these data we conducted assays to verify whether BaV affects adhered thioglycollate-elicited macrophage phagocytosis via complement receptor using opsonized zymosan particles. Our results showed that BaV stimulated macrophage phagocytosis. In line with our results, Zamuner et al. (33) observed increased phagocytosis of opsonized zymosan particles by peritoneal leukocytes induced by Bothrops asper and Bothrops jararaca venom 12 and 48 hours after intraperitoneal injection in mice.
The fact that snake venom activates the phagocytosis process suggests that leukocyte function is an important event for the elimination of venom in envenomed individuals. Thus, the literature shows that both resident macrophages and macrophages recruited to the injured muscle, perform phagocytosis of cellular debris and facilitate the regeneration of damaged myofibrils. It is believed that these cells, through the production of several cytokines and growth factors derived from platelets, induce chemotaxis of muscle cell and other leukocyte precursors by stimulating endothelial and muscle cell proliferation and promoting muscle regeneration (62).
The phagocytic process triggered by complement receptor 3 (CR3) requires an additional activation signal for efficient particle binding and subsequent internalization. These stimuli activate PKC and induce signaling from the inside out culminating in the internalization of the particle by the mechanism of invagination (37, 42-46).
Accordingly, we designed experiments to analyze the participation of PKC in the BaV stimulated process of opsonized zymosan particle phagocytosis. Our results showed that PKC is involved in BaV-induced phagocytosis as the use of staurosporine, a PKC inhibitor, abolished thioglycollate-elicited macrophage phagocytosis via complement receptor.
PKC is a large family of protein kinases with serine/threonine domains consisting of ten protein isotypes encoded by nine genes (63). The PKC isotypes exhibit distinct patterns of tissue expression and intracellular location. However, the current study did not determine which isoforms are involved in BaV induced phagocytosis. The literature shows that different PKC isoforms are translocated to membranes during phagocytosis in response to different stimuli and during cell surface receptor stimulation, each PKC isotype is translocated or targeted to particular intracellular compartments, such as plasma membrane, Golgi complex, mitochondria, nucleus, and cytoskeleton (64-66). Still, the destiny of the enzyme can be targeted by lipid mediators and perhaps by phosphorylation.
Recent studies have demonstrated that lipids produced in the membrane after receptor stimulation can participate in PKC activation. These lipids include e free fatty acids, especially arachidonic acid, phosphatidic acid, lysophospholipids, phosphatidylinositol, and diacylglycerol (DAG) (67). Translocation and subsequent binding to the membrane is due to the presence of specific lipid binding regions, including DAG, arachidonic acid, and ceramide (68). In quiescent conditions, PKC is cytoplasmic and inactive. When stimulated, it is translocated to the membrane, where it phosphorylates its substrate in selected residues of serine or threonine (69, 70).
Concomitantly with phagocytosis, there is an increase in oxidative leukocyte metabolism. The high oxygen consumption associated with rapid activation of NADPH oxidase triggers a respiratory burst, with the subsequent production of superoxide ions and hydrogen peroxide (71). The enzyme responsible for superoxide production is a multicomponent NADPH oxidase or "burst" respiratory oxidase (51, 72). In phagocytes, NADPH and the production of reactive oxygen species (ROS) are developing a key role in host defense against microbial pathogens, as illustrated by a human genetic disorder known as chronic granulomatous disease (CGD), characterized by the absence of ROS production due a deficiency in one component of NADPH oxidase (73,74).
The NADPH enzyme complex consists of several separate proteins between the membrane and the cytosol (72). Cytosolic proteins p47phox, p67phox, and p40phox interact with each other to form a complex, and small G proteins Rac 1 (in monocytes) or Rac 2 (in neutrophil) and the membrane components are associated with the glycosylated 91 kDa protein (gp91phox or NOX 2) and subunit (p22phox), which together form flavocytocrome b558 (45, 75). The spatial separation of NADPH oxidase components ensures that the enzyme is dormant in quiescent cells. However in response to stimuli, the cytosolic components migrate almost instantly to the membrane where they meet flavocytocrome b558 to form the active enzyme, a process highly regulated by protein-protein interactions and p47phox phosphorylation (76-79). Thus macrophage NADPH oxidase plays an important role in host defense against microbial pathogens by generating the superoxide anion and other ROS molecules.
The excessive release of superoxide may damage tissue involved in inflammation. We therefore evaluated the effect of BaV on the release of superoxide anions by thioglycollate-elicited macrophages. In this work, our findings showed that BaV induced a significant release of superoxide anion indicating that BaV is able to stimulate macrophages to activate respiratory burst. Moreover, the literature shows that the injection of Bothrops asper and Bothrops jararaca venom into the peritoneal cavity of mice induce hydrogen peroxide production by peritoneal leukocytes, thus indicating that they are capable of priming leukocytes for respiratory burst (33).
The mechanism by which BaV stimulates superoxide anion generation has not been fully clarified. It is therefore possible that the venom activates NADPH oxidase by a direct mechanism or indirectly through the activation of signaling pathways such as PKC, which culminate in NADPH oxidase activation. The present results clearly showed that stimulatory action by the venom was abolished after treating cells with staurosporine, indicating that PKC is an important signaling pathway for superoxide anion production in thioglycollate-elicited macrophages. Reactive oxygen species can also be generated by mechanisms independent of NADPH oxidase, via mitochondria. The production of superoxide and hydrogen peroxide in mitochondria can be stimulated by activation of small conductance calcium-activated potassium channels (80). It is noteworthy that BaV may act through different mechanisms.
Taken together, our data support the conclusion that BaV does not interfere with the adhesion or detachment of adherent macrophages from the support. The venom stimulates phagocytosis of opsonized zymosis particles by activating CR3 through molecular signaling mechanisms that involve PKC. However, the mechanism by which BaV interacts with macrophages to trigger this signaling pathway has not been established. Even so, venom did induce superoxide production by a PKC-dependent pathway. Although results from this study showed that the venom action mechanism on phagocytosis and superoxide production by macrophages occurs by a PKC-dependent signaling pathway, additional studies are required to determine which isoforms are involved in these processes and how activation of this kinase occurs. Finally, such studies have added knowledge to the action mechanisms of Bothrops alternatus venom, contributing to better characterization of macrophage functionality.
ACKNOWLEDGMENTS
We are extremely grateful to The National Council for Scientific and Technological Development (CNPq), the State of Rondônia Planning Secretariat (SEPLAN-RO) and the Coordination for the Improvement of Higher Education Personnel (CAPES) for their financial support; and to Caroline Vargas Xavier and Fabianne Lacouth da Silva for technical assistance and critical reading of the manuscript.
Submission status
Received: March 31, 2011.
Accepted: June 1, 2011.
Abstract: published online: June 2, 2011.
Full paper published online: November 30, 2011.
Conflicts of interest
There are no conflicts of interest.
Financial source
The National Council for Scientific and Technological Development (CNPq) and the State of Rondônia Planning Secretariat (SEPLAN-RO) provided financial support. Moreover, Sulamita da Silva Setúbal was the beneficiary of a CNPq fellowship.
Ethics committee approval
This study was approved by the Tropical Pathology Research Institute Ethics Committee on Animal Use under the protocol number 2008/8. Furthermore, animal tests were in accordance with the guidelines of the Brazilian College for Animal Experimentation (COBEA).
- 1. Oliveira RC, Wen FH, Sifuentes DN. Epidemiologia dos acidentes por animais peçonhentos. In: Cardoso JL, Haddad-Jr V, França FOS, Wen FH, Malaque CMS, editors. Animais peçonhentos do Brasil: biologia, clínica e terapêutica. 2nd ed. São Paulo: Sarvier; 2009. p. 6-21.
-
2Brasil. Ministério da Saúde. Manual de diagnóstico e tratamento de acidentes por animais peçonhentos. 2nd ed. Brasília: Fundação Nacional de Saúde; 2008.
- 3. Amorim MF, Franco de Mello, R, Saliba F. Envenenamento botrópico e crotálico. Mem Inst Butantan. 1951;23:108.
- 4. Barraviera B. Acidentes por serpentes do gênero Crotalus Arq Bras Med. 1990;64:14-20.
- 5. Barraviera B. Estudo clínico dos acidentes ofídicos. J Bras Med. 1993;65(4):209-50.
- 6. Campbell JA, Lamar WW. The venomous reptiles of Latin America. Cornell Univ. Press: Ithaca; 1989. p. 425.
- 7. Fenwick AM, Gutberlet RL Jr, Evans JA, Christopher LP. Morphological and molecular evidence for phylogeny and classiûcation of South American pitvipers, genera Bothrops, Bothriopsis, and Bothrocophias (Serpentes: Viperidae). Zool J Linn Soc. 2009;156(3):617-40.
- 8. Rosenfeld G. Symptomatology, pathology and treatment of snakes bites in South America. In: Burchel, W, Buckley, EE, editors. Venomous animals and their venoms. New York: Academic Press; 1971. p. 345-403.
- 9. Furtado MF, Maruyama M, Kamiguti AS, Antonio LC. Comparative study of nine Bothrops snake venoms from adult female snakes and their offspring. Toxicon. 1991;29(2):219-26.
- 10. de Roodt AR, Dolab JA, Galarce PP, Gould E, Litwin S, Dokmetjian JC, et al. A study on the venom yield of venomous snake species from Argentina. Toxicon. 1998;36(12):1949-57
- 11. Mebs D. A comparative study of enzyme activities in snake venoms. Int J Biochem. 1970;1(3):335-42.
- 12. Tan NH, Ponnudurai G. A comparative study of the biological properties of some venoms of snacks of the genus Bothrops (American lance-headed viper). Comp Biochem Physiol. 1991;100(2):361-5.
- 13. Stábeli RG, Marcussi S, Carlos GB, Pietro RC, Selistre-de-Araújo HS, Giglio JR, et al. Platelet aggregation and antibacterial effects of an L-amino acid oxidase purified from Brothops alternatus snake venom. Bioorg Med Chem. 2004;12(11):2881-6.
- 14. Nisenbom HE, Seki C, Vidal JC. Phospholipase A2 from Bothrops alternatus (víbora de la cruz) venom. Purification and some characteristic properties. Toxicon. 1986;24(3):259-72.
- 15. Smolka MB, Marangoni S, Oliveira B, Novello JC. Purification and partial characterization of a thrombin-like enzyme, balterobin, from the venom of Bothrops alternatus Toxicon. 1998;36(7):1059-63.
- 16. Castro HC, Dutra DL, Oliveira-Carvalho AL, Zingali RB. Bothroalternin, a thrombin inhibitor from the venom of Bothrops alternatus Toxicon. 1998;36(12):1903-12.
- 17. Souza DH, Iemma MR, Ferreira LL, Faria JP, Oliva ML, Zingali RB, et AL. The disintegrin-like domain of the snake venom metalloprotease alternagin inhibits alpha2beta1 integrin-mediated cell adhesion. Arch Biochem Biophys. 2000;384(2):341-50.
- 18. Mariano-Oliveira A, Coelho AL, Terruggi CH, Selistre-de-Araújo HS, Barja-Fidalgo C, De Freitas MS. Alternagin-C, a nonRGD-disintegrin, induces neutrophil migration via integrin signaling. Eur J Biochem. 2003;270(24):4799-808.
- 19. Cominetti MR, Ribeiro JU, Fox JW, Selistre-de-Araujo, HS. BaG, a new dimeric metalloproteinase/disintegrin from the Bothrops alternatus snake venom, that interacts with alpha5beta1 integrin. Arch Biochem Biophys. 2003;416(2):171-9.
- 20. Cominetti MR, Terrugi CHB, Ramos OHP, Fox JW, Figueiredo CC, Mariano-Oliveira A, et al. Alternagin-C, a disintegrin like protein, induces vascular endothelial cell growth factor (VEGF) expression and endothelial cell proliferation in vitro J Biol Chem. 2004;279(18):18247-55.
- 21. Gay CC, Leiva IC, Maruñak S, Teibler P, Acosta de Pérez O. Proteolytic, edematogenic and myotoxic activities of a hemorrhagic metalloproteinase isolated from Bothrops alternatus venom. Toxicon. 2005;46(5):546-54.
- 22. Selistre-de-Araújo HS, Cominetti MR, Terruggi CH, Mariano-Oliveira A, De Freitas MS, Crepin M, et al. Alternagin-C, a disintegrin-like protein from the venom of Bothrops alternatus, modulates alpha2beta1 integrin-mediated cell adhesion, migration and proliferation. Braz J Med Biol Res. 2005;38(10):1505-11.
- 23. Valério AA, Corradini AC, Panunto PC, Mello SM, Hyslop S. Purification and characterization of a phosphodiesterase from Bothrops alternatus snake venom. J Protein Chem. 2002;21(8):495-503.
- 24. Bauab FA, Junqueira GR, Corradini MC, Silveira PV, Nishioka S de A. Clinical and epidemiological aspects of the 'urutu' lance-headed viper (Bothrops alternatus) bite in a Brazilian hospital. Trop Med Parasitol. 1994;45(3):243-5.
- 25. Maruñak SL, Acosta De Pérez O, Ruíz De Torrent RM, Teibler GP. Hemorrhagic, edema-forming, proteolytic and myonecrotic activities of viper venoms of Bothrops alternatus (Vibora de la Cruz). Acta Physiol Pharmacol Ther Latinoam. 1999;49(3):149-54.
- 26. de Roodt AR, Dolab JA, Dokmetjian JC, Litwin S, Segre L, Vidal JCA. A comparison of different methods to assess the hemorrhagic activity of Bothrops venoms. Toxicon. 2000;38(6):865-73.
- 27. Queiroz LS, Santo-Neto H, Rodrigues-Simioni L, Prado-Franceschi J. Muscle necrosis regeneration after envenomation by Bothrops jararacussu snake venom. Toxicon. 1984;22(3):339-46.
- 28. Sanchez EF, Freitas TV, Ferreira-Alves DL, Velarde DT, Diniz MR, Cordeiro MN. Biological activities of venoms from South American snakes. Toxicon. 1992;30(1):95-103.
- 29. Gutiérrez JM, Ownby CL, Odell GV. Skeletal muscle regeneration after myonecrosis induced by crude venom and a myotoxin from the snake Bothrops asper (Fer-de-Lance). Toxicon. 1984;22(5):719-31
- 30. Gutiérrez JM, Chaves F, Cerdas L. Inflammatory infiltrate in skeletal muscle injected with Bothrops asper venom. Rev Biol Trop. 1986;34(2):209-19.
- 31. Trebien HA, Calixto JB. Pharmacological evaluation of rat paw oedema induced by Bothrops jararaca venom. Agents Actions.1989;26(3-4):292-300.
- 32. Flores CA, Zappellini A, Prado-Franceschi J. Lypoxygenase-derived mediators may be involved in in vivo neutrophil migration induced by Bothrops erythromelas and Bothrops alternatus venom. Toxicon. 1993;31(12):1551-9.
- 33. Zamuner SR, Gutiérrez JM, Muscará MN, Teixeira SA, Teixeira CF. Bothrops asper and Bothrops jararaca snake venoms trigger microbicidal functions of peritoneal leukocytes in vivo Toxicon.2001;39(10):1505-13.
- 34. Acosta de Pérez O, Teibler P, Koscinczuk P, Sánchez Negrette M, Trulls H, Maruñak S. Edema and myonecrosis induced by Bothrops jararaca venom of Argentina in mice. Acta Physiol Pharmacol Ther Latinoam. 1996;46(4):233-8.
- 35. Farsky SH,Walber J, Costa-Cruz JW, Cury Y, Teixeira CF. Leukocyte response induced by Bothrops jararaca crude venom. In vivo and in vitro studies. Toxicon. 1997;35(2):185-93.
- 36. Naito M. Macrophage heterogeneity in development and differentiation. Arch Histol Cytol. 1993;56(4):331-51.
- 37. Aderem A, Underhill DM. Mechanisms of phagocytosis in macrophages. Annu Rev Immunol. 1999;17:593-23.
- 38. Brown GD, Gordon S. Immune recognition. A new receptor for beta-glucans. Nature. 2001; 413: 36-7.
- 39. Brown WJ, Chambers K, Doody A. Phospholipase A2 (PLA2) enzymes in membrane trafficking: mediators of membrane shape and function. Traffic. 2003;4(4):214-21.
- 40. Herre J, Gordon S, Brown GD. Dectin-1 and its role in the recognition of beta-glucans by macrophages. Mol Immunol. 2004;40(12):869-76.
- 41. Herre J, Marshall AS, Caron E, Edwards AD, Williams DL, Schweighoffer E, et al. Dectin-1 uses novel mechanisms for yeast phagocytosis in macrophages. Blood. 2004;104(13):4038-45.
- 42. Wright SD, Silverstein SC. Tumor-promoting phorbol esters stimulate C3b and C3b' receptor-mediated phagocytosis in cultured human monocytes. J Exp Med. 1982;156(4):1149-64.
- 43. Griffin FM Jr, Mullinax PJ. Effects of differentiation in vivo and of lymphokine treatment in vitro on the mobility of C3 receptors of human and mouse mononuclear phagocytes. J Immunol. 1985;135(5):3394-7.
- 44. Ross GD, VĔtvicka V. CR3 (CD11b, CD18): a phagocyte and NK cell membrane receptor with multiple ligand specificities and functions. Clin Exp Immunol. 1993;92(2):181-4.
- 45. Williams MA, Solomkin JS. Integrin-mediated signaling in human neutrophil functioning. J Leukoc Biol. 1999;65(6):725-36.
- 46. Schmidt A, Caron E, Hall A. Lipopolysaccharide-induced activation of beta2-integrin function in macrophages requires Irak kinase activity, p38 mitogen-activated protein kinase, and the Rap1 GTPase. Mol Cell Biol. 2001;21(2):438-48.
- 47. Caron E. Cellular functions of the Rap 1 GTP-binding protein: a pattern emerges. J Cell Sci. 2003;116(Pt 3)435-40.
- 48. Lim J, Wiedemann A, Tzircotis G, Monkley SJ, Critchley DR, Caron E. An essential role for talin during alpha(M)beta(2)-mediated phagocytosis. Mol Biol Cell. 2007;18(3):976-85.
- 49. Wright SD, Griffin FM Jr. Activation of phagocytic cells' C3 receptors for phagocytosis. J Leukoc Biol. 1985;38(2):327-39.
- 50. Van Kooyk Y, Figdor CG. Avidity regulation of integrins: the driving force in leukocyte adhesion. Curr Opin Cell Biol. 2000;12(5):542-7.
- 51. Babior BM. NADPH oxidase: an update. Blood. 1999;93(4):1464-76.
- 52. Babior BM. NADPH oxidase. Curr Opin Immunol. 2004;16(1):42-7.
- 53. Babior BM, Lambeth, JD, Nauseef W. The neutrophil NADPH oxidase. Arch Biochem Biophys. 2002;397(2):342-4.
- 54. Forman HJ, Torres M. Reactive oxygen species and cell signaling: Respiratory burst in macrophage signaling. Am J Respir Crit Care Med. 2002;166(12 Pt 2):S4-8.
- 55. Zuliani JP, Gutiérrez JM, Casais e Silva LL, Sampaio SC, Teixeira CFP. Activation of cellular functions in macrophages by venom secretory Asp-49 and Lys-49 Phospholipases A2. Toxicon. 2005;46(3):523-32.
- 56. Rosen H, Gordon S. Monoclonal antibody to the murine type 3 complement receptor inhibits adhesion of myelomonocytic cells in vitro and anflamatory cell recruitment in vivo J Exp Med. 1987;166:1685-701.
- 57. Costa Rosa LF, Saû DA, Curi R. Effect of thioglycollate and BCG stimuli on glucose and glutamine metabolism in rat macrophages. J Leukoc Biol. 1994; 56(1):10-4.
- 58. Madhavi N, Das VN. Effects os n-6 and n-3 fatty acids on the survival of vincristine sensitive and resistant human cervical carcinoma cells, in vitro Cancer Lett. 1994;84(1):31-41.
- 59. Oliveira JC, de Oca HM, Duarte MM, Diniz CR, Fortes-Dias CL. Toxicity of South American snake venoms measured by an in vitro cell culture assay. Toxicon. 2002. 40(3):321-5.
- 60. Moreira V, Gutiérrez JM, Amaral RB, Zamunér SR, Teixeira C de F. Effects of Bothrops asper snake venom on the expression of cyclooxygenases and production of prostaglandins by peritoneal leukocytes in vivo, and by isolated neutrophils and macrophages in vitro Prostaglandins Leukot Essent Fatty Acids. 2009. 80(2-3):107-14.
- 61. Taylor AC. Attachment and spreading of cells in culture. Exp Cell Res. 1961; 8:154-73.
- 62. Robertson TA, Maley MA, Grounds MD, Papadimitriou JM. The role of macrophages in skeletal muscle regeneration with particular reference to chemotaxis. Exp Cell Res. 1993;207(2):321-31.
- 63. Ohno S, Nishizuka Y. Protein kinase C isotypes and their specific functions: prologue. J Biochem. 2002;132(4):509-11.
- 64. Allen LA, Aderem A. Molecular definition of distinct cytoesqueletal structures involved in complement-and Fc receptor-mediated phagocytosis in macrophages. J Exp Med. 1996;184(2):627-37.
- 65. Larsen EC, Di Gennaro JA, Saito N, Metha S, Loegering, DJ, Mazurkiewicz JM, et al. Differential requirement for classic and novel PKC isoforms in respiratory burst and phagocytosis in RAW 264.7 cells. J Immunol. 2000;165(5):2809-17.
- 66. Yedovitzky M, Mochly-Rosen D, Johnson JA, Gray MO, Ron D, Abramovitchi E, et al. Translocation inhibitors define specificity of PKC isoenzymes in pancreatic β cells. J Biol Chem. 1997;272(3):1417-20.
- 67. Nishizuka Y. Protein kinase C and lipid signaling for sustained cellular responses. FASEB J. 1995;9(7):484-96.
- 68. Kashiwagi K, Shirai Y, Kuriyama M, Sakai N, Saito N. Importance of C1B domain for lipid messenger-induced targeting of protein kinase C. J Biol Chem. 2002;277(20):18037-45.
- 69. Newton AC. Protein Kinase C: structural and spatial regulation by phosphorilatyon, cofactors, and macromolecular interactions. Chem Rev. 2001;101(8):2353-64.
- 70. Newton AC. Regulation of the ABC kinases by phosphorylation: protein kinase C as a paradigm. Biochem J. 2003;370(Pt 2):361-71.
- 71. Babior BM, Kipnes RS, Curcnutte JT. Biological defense mechanism. The production by leukocytes of superoxide, a potential bactericidal agent. J Clin Invest.1973;52(3):741-4.
- 72. El-Benna J, Dang PM, Gougerot-Pocidalo MA, Elbim C. Phagocyte NADPH oxidase: a multicomponent enzyme essential for host defenses. Arch Immunol Ther Exp (Warsz). 2005;53(3):199-206.
- 73. Meischl C, Roos D. The molecular basis of chronic granulomatous disease. Springer Semin Immunopathol. 1998;19(4):417-34.
- 74. Kannengiesser C, Gérard B, El Benna J, Henri D, Kroviarski Y, Chollet-Martin S, et al. Molecular epidemiology of chronic granulomatous disease in a series of 80 kindreds: identification of 31 novel mutations. Hum Mutat. 2008;29(9):132-49.
- 75. Vignais PV. The superoxide-generating NADPH oxidase: structural aspects and activation mechanism. Cell Mol Life Sci. 2002;59(9):1428-59.
- 76. Groemping Y, Rittinger K. Activation and assembly of the NADPH-oxidase: a structural perspective. Biochem J. 2005;386(Pt 3):401-16.
- 77. el Benna J, Faust LP, Babior BM. The phosphorylation of the respiratory burst oxidase component p47phox during neutrophil activation. Phosphorylation of sites recognized by protein kinase C and by proline directed kinases. J Biol Chem.1994;269(38):23431-6.
- 78. Faust LR, el Benna J, Babior BM., Chanock SJ. The phosphorylation targets of p47phox, a subunit of the respiratory burst oxidase. Functions of the individual target serines as evaluated by site-directed mutagenesis. J Clin Invest. 1995;96(3):1499-505.
- 79. Quinn MT, Gauss KA. Structure and regulation of the neutrophil respiratory burst oxidase: comparison with nonphagocyte oxidases. J Leukoc Biol. 2004;76(4):760-81.
- 80. Fay AJ, Qian X, Jan YN, Jan LY. SK channels mediate NADPH oxidase-independent reactive oxygen species production and apoptosis in granulocytes. Proc Natl Acad Sci USA. 2006; 103(46):17548-53.
Publication Dates
-
Publication in this collection
05 Dec 2011 -
Date of issue
2011
History
-
Received
31 Mar 2011 -
Accepted
01 June 2011